
The Role of Dysregulated miRNAs in the Pathogenesis, Diagnosis and Treatment of Age-Related Macular Degeneration

 , , , ,
, , , ,  ,
,  and
and
Abstract
:
1. Introduction
2. Retinal, Circulating and Vitreous Body miRNAs Found in Human Studies (2016–2021)
2.1. Retinal miRNA Expression
2.2. Vitreous Body miRNA Expression
2.3. Circulating miRNA Expression
2.4. Circulating miRNA Expression in Wet and Dry AMD
2.5. Relationship between Expression of miRNAs and Polymorphisms in Genes Associated with AMD
2.6. Interplay between miRNAs and Physical Examination
2.7. Summary and Limitations of the Studies
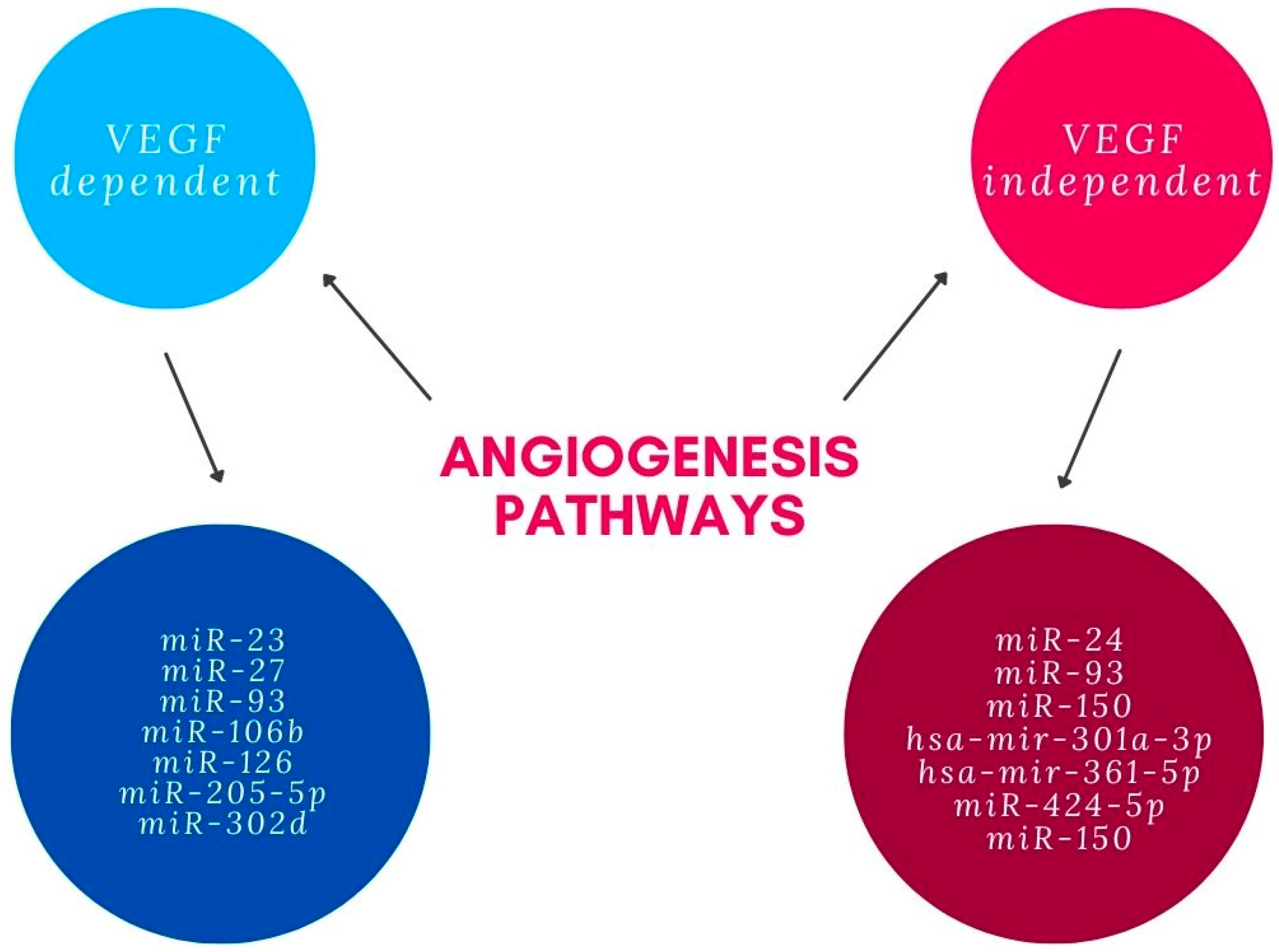
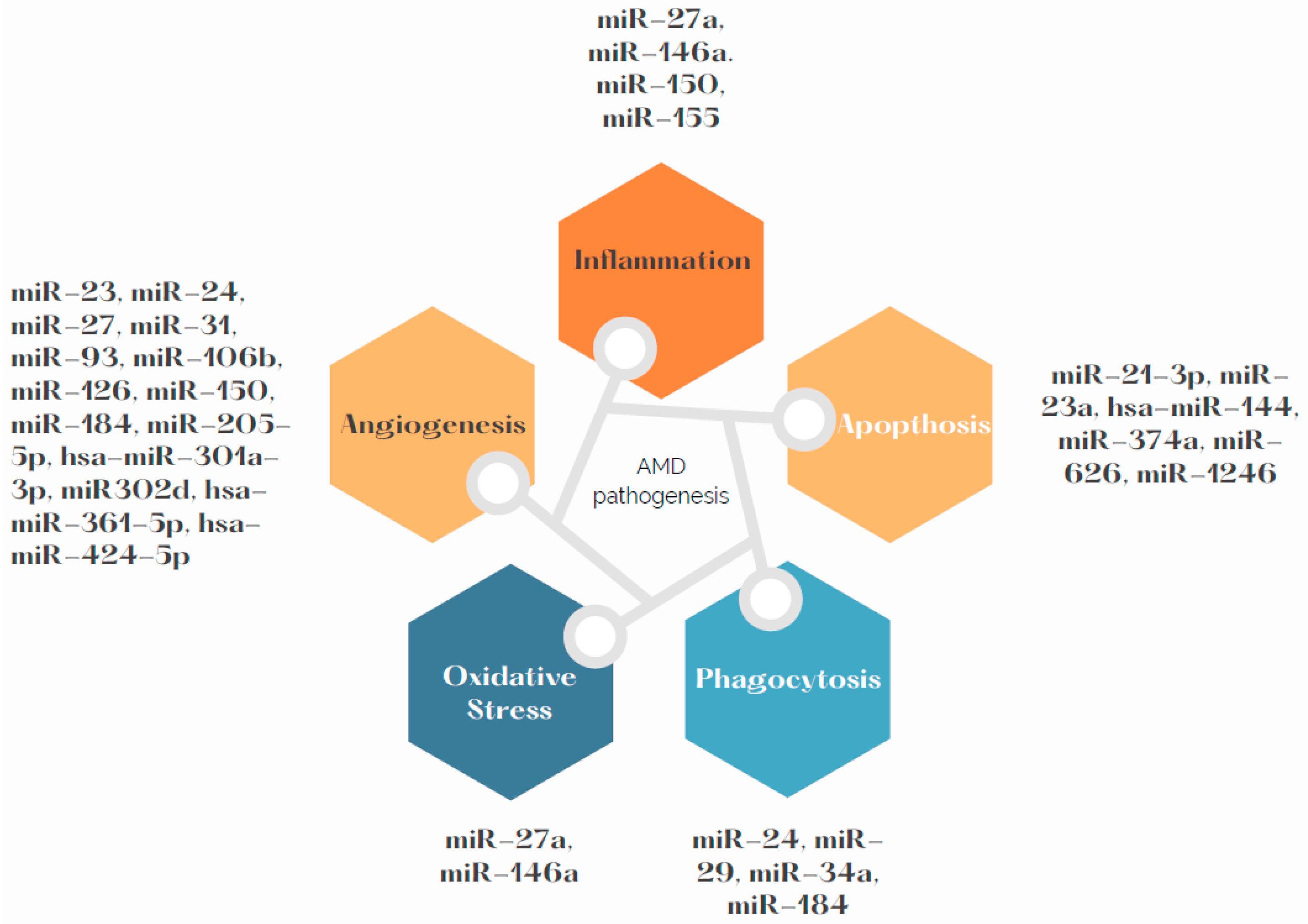
3. Influence of miRNAs on Angiogenesis in AMD Pathogenesis
4. Influence of miRNAs on Phagocytosis in AMD Pathogenesis
5. Influence of miRNAs on Apoptosis in AMD Pathogenesis
6. Influence of miRNAs on Inflammation and Oxidative Stress in AMD Pathogenesis
7. miRNAs as Potential Therapeutic Targets
8. Potential Role of miRNAs in Clinical Practice
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Ethical conduct of research
Data Availability Statement
Conflicts of Interest
Abbreviations
| AMD | Age-related macular degeneration |
| CNV | choroidal neovascularization |
| miRNA | microRNA |
| NV AMD | neovascular age-related macular degeneration |
| VEGFA | Vascular endothelial factor A |
| CFH | Complement Factor H |
| ARMS | age-related maculopathy susceptibility |
| VEGF | vascular endothelial growth factor |
| MAPK | mitogen-activated protein kinase |
| VEGFR2 | vascular endothelial growth factor receptor 2 |
| PI3K | phosphatidylinositol-3-kinase |
| PKB | Protein kinase B |
| PERK | protein kinase RNA-like endoplasmic reticulum kinase |
| HIF1A | hypoxia-inducible factor-α |
| TGFβ | Transforming Growth Factor β |
| ARPE-19 | Adult Retinal Pigment Epithelial cell line-19 |
| TGFβR2 | Transforming Growth Factor β Receptor 2 |
| KDR | kinase insert domain receptor |
| VEGFR-2 | vascular endothelial growth factor receptor-2 |
| SPRED-1 | Sprouty-related EVH1 domain-containing protein 1 |
| RPE | Retinal pigment epithelium |
| HUVECS | Human Umbilical Vein Endothelial Cells |
| Limk2 | LIM domain kinase 2 |
| Pak4 | p21-Activated kinase 4 |
| Diaph1 | homologs of Drosophila diaphanous |
| mTOR | mammalian target of rapamycin |
| Scd2 | stearoyl-CoA desaturase-2 |
| TREM2 | Triggering receptor expressed in myeloid/microglial cells-2 |
| CNS | Central nervous system |
| NF-κB | nuclear factor kappa-light-chain enhancer of activated B cells |
| CHI3L1 | Chitinase-3-like protein 1 |
| ERK | extracellular signal-regulated kinase |
| mTORC1 | mammalian target of rapamycin complex 1 |
| EZR | ezrin |
| LAMP-1 | lysosomal-associated membrane protein 1 |
| NaIO3 | Sodium iodate |
| Keap1 | Kelch-like ECH-associated protein 1 |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| FasL | Fas Ligand |
| LPS | lipopolysaccharide |
| RGS4 | regulator of G protein signaling 4 |
| IL-6 | interleukin 6 |
| MCP-1 | monocyte chemoattractant protein-1 |
| UVB | Ultraviolet B |
| iASPP | inhibitor of apoptosis stimulating protein of p53 |
| apoB | apolipoprotein B |
| FOXO1 | forkhead box protein O1 |
| ASO | antisense oligonucleotides |
| TNFα | Tumor necrosis factor α |
| HIF | hypoxia-inducible factor |
| OCT | Optical Coherence Tomography |
References
- Arruabarrena, C.; Toro, M.D.; Onen, M.; Malyugin, B.E.; Rejdak, R.; Tognetto, D.; Zweifel, S.; Giglio, R.; Teus, M.A. Impact on Visual Acuity in Neovascular Age Related Macular Degeneration (NAMD) in Europe Due to COVID-19 Pandemic Lockdown. J. Clin. Med. 2021, 10, 3281. [Google Scholar] [CrossRef] [PubMed]
- Fleckenstein, M.; Keenan, T.D.L.; Guymer, R.H.; Chakravarthy, U.; Schmitz-Valckenberg, S.; Klaver, C.C.; Wong, W.T.; Chew, E.Y. Age-Related Macular Degeneration. Nat. Rev. Dis. Prim. 2021, 7, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.; Liew, G.; Gopinath, B.; Wong, T.Y. Age-Related Macular Degeneration. Lancet 2018, 392, 1147–1159. [Google Scholar] [CrossRef]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.G.; Klein, R.; Cheng, C.Y.; Wong, T.Y. Global Prevalence of Age-Related Macular Degeneration and Disease Burden Projection for 2020 and 2040: A Systematic Review and Meta-Analysis. Lancet. Glob. Health 2014, 2, E106–E116. [Google Scholar] [CrossRef] [Green Version]
- Bourne, R.R.A.; Steinmetz, J.D.; Saylan, M.; Mersha, A.M.; Weldemariam, A.H.; Wondmeneh, T.G.; Sreeramareddy, C.T.; Pinheiro, M.; Yaseri, M.; Yu, C.; et al. Causes of Blindness and Vision Impairment in 2020 and Trends over 30 Years, and Prevalence of Avoidable Blindness in Relation to VISION 2020: The Right to Sight: An Analysis for the Global Burden of Disease Study. Lancet. Glob. Health 2021, 9, e144–e160. [Google Scholar] [CrossRef]
- Russo, A.; Ragusa, M.; Barbagallo, C.; Longo, A.; Avitabile, T.; Uva, M.G.; Bonfiglio, V.; Toro, M.D.; Caltabiano, R.; Mariotti, C.; et al. MiRNAs in the Vitreous Humor of Patients Affected by Idiopathic Epiretinal Membrane and Macular Hole. PLoS ONE 2017, 12, e0174297. [Google Scholar] [CrossRef]
- Toro, M.D.; Reibaldi, M.; Avitabile, T.; Bucolo, C.; Salomone, S.; Rejdak, R.; Nowomiejska, K.; Tripodi, S.; Posarelli, C.; Ragusa, M.; et al. MicroRNAs in the Vitreous Humor of Patients with Retinal Detachment and a Different Grading of Proliferative Vitreoretinopathy: A Pilot Study. Transl. Vis. Sci. Technol. 2020, 9, 1–13. [Google Scholar] [CrossRef]
- Natoli, R.; Fernando, N. MicroRNA as Therapeutics for Age-Related Macular Degeneration. Adv. Exp. Med. Biol. 2018, 1074, 37–43. [Google Scholar] [CrossRef]
- Wang, K.; Long, B.; Liu, F.; Wang, J.X.; Liu, C.Y.; Zhao, B.; Zhou, L.Y.; Sun, T.; Wang, M.; Yu, T.; et al. A Circular RNA Protects the Heart from Pathological Hypertrophy and Heart Failure by Targeting MiR-223. Eur. Heart J. 2016, 37, 2602a–2611a. [Google Scholar] [CrossRef]
- Chen, S.; Li, T.; Zhao, Q.; Xiao, B.; Guo, J. Using Circular RNA Hsa_circ_0000190 as a New Biomarker in the Diagnosis of Gastric Cancer. Clin. Chim. Acta 2017, 466, 167–171. [Google Scholar] [CrossRef]
- Cao, D.D.; Li, L.; Chan, W.Y. MicroRNAs: Key Regulators in the Central Nervous System and Their Implication in Neurological Diseases. Int. J. Mol. Sci. 2016, 17, 842. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Liu, X.F.; Pant, O.P.; Zhou, D.D.; Hao, J.L.; Lu, C.W. Circular RNAs: Novel Promising Biomarkers in Ocular Diseases. Int. J. Med. Sci. 2019, 16, 513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.J.; Chen, X.; Li, C.P.; Li, X.M.; Liu, C.; Liu, B.H.; Shan, K.; Jiang, Q.; Zhao, C.; Yan, B. Identification and Characterization of Circular RNAs as a New Class of Putative Biomarkers in Diabetes Retinopathy. Investig. Ophthalmol. Vis. Sci. 2017, 58, 6500–6509. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, H.; Terasaki, H. Biological Involvement of MicroRNAs in Proliferative Vitreoretinopathy. Transl. Vis. Sci. Technol. 2017, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Ragusa, M.; Caltabiano, R.; Russo, A.; Puzzo, L.; Avitabile, T.; Longo, A.; Toro, M.D.; Di Pietro, C.; Purrello, M.; Reibaldi, M. MicroRNAs in Vitreus Humor from Patients with Ocular Diseases. Mol. Vis. 2013, 19, 430–440. [Google Scholar] [PubMed]
- Ragusa, M.; Barbagallo, C.; Statello, L.; Caltabiano, R.; Russo, A.; Puzzo, L.; Avitabile, T.; Longo, A.; Toro, M.D.; Barbagallo, D.; et al. MiRNA Profiling in Vitreous Humor, Vitreal Exosomes and Serum from Uveal Melanoma Patients: Pathological and Diagnostic Implications. Cancer Biol. Ther. 2015, 16, 1387–1396. [Google Scholar] [CrossRef]
- Romano, G.L.; Platania, C.B.M.; Drago, F.; Salomone, S.; Ragusa, M.; Barbagallo, C.; Di Pietro, C.; Purrello, M.; Reibaldi, M.; Avitabile, T.; et al. Retinal and Circulating MiRNAs in Age-Related Macular Degeneration: An In Vivo Animal and Human Study. Front. Pharmacol. 2017, 8, 168. [Google Scholar] [CrossRef] [Green Version]
- Shewmaker, F.; McGlinchey, R.P.; Wickner, R.B. Structural Insights into Functional and Pathological Amyloid. J. Biol. Chem. 2011, 286, 16533. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, S.; Zhao, Y.; Dua, P.; Rogaev, E.I.; Lukiw, W.J. MicroRNA-34a-Mediated Down-Regulation of the Microglial-Enriched Triggering Receptor and Phagocytosis-Sensor TREM2 in Age-Related Macular Degeneration. PLoS ONE 2016, 11, e0150211. [Google Scholar] [CrossRef]
- Pogue, A.I.; Lukiw, W.J. Up-Regulated Pro-Inflammatory MicroRNAs (MiRNAs) in Alzheimer’s Disease (AD) and Age-Related Macular Degeneration (AMD). Cell. Mol. Neurobiol. 2018, 38, 1021–1031. [Google Scholar] [CrossRef]
- Ménard, C.; Rezende, F.A.; Miloudi, K.; Wilson, A.; Tétreault, N.; Hardy, P.; SanGiovanni, J.P.; De Guire, V.; Sapieha, P.; Ménard, C.; et al. MicroRNA Signatures in Vitreous Humour and Plasma of Patients with Exudative AMD. Oncotarget 2016, 7, 19171–19184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blasiak, J.; Watala, C.; Tuuminen, R.; Kivinen, N.; Koskela, A.; Uusitalo-Järvinen, H.; Tuulonen, A.; Winiarczyk, M.; Mackiewicz, J.; Zmorzyński, S.; et al. Expression of VEGFA-Regulating MiRNAs and Mortality in Wet AMD. J. Cell. Mol. Med. 2019, 23, 8464–8471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbay, A.; Ercan, Ç.; Akbaş, F.; Bulut, H.; Ozdemir, H. Three New Circulating MicroRNAs May Be Associated with Wet Age-Related Macular Degeneration. Scand. J. Clin. Lab. Investig. 2019, 79, 388–394. [Google Scholar] [CrossRef]
- Ren, C.; Liu, Q.; Wei, Q.; Cai, W.; He, M.; Du, Y.; Xu, D.; Wu, Y.; Yu, J. Circulating MiRNAs as Potential Biomarkers of Age-Related Macular Degeneration. Cell. Physiol. Biochem. 2017, 41, 1413–1423. [Google Scholar] [CrossRef] [PubMed]
- Litwińska, Z.; Sobuś, A.; Łuczkowska, K.; Grabowicz, A.; Mozolewska-Piotrowska, K.; Safranow, K.; Kawa, M.P.; Machaliński, B.; Machalińska, A. The Interplay Between Systemic Inflammatory Factors and MicroRNAs in Age-Related Macular Degeneration. Front. Aging Neurosci. 2019, 11, 286. [Google Scholar] [CrossRef]
- Ulańczyk, Z.; Sobuś, A.; Łuczkowska, K.; Grabowicz, A.; Mozolewska-Piotrowska, K.; Safranow, K.; Kawa, M.P.; Pałucha, A.; Krawczyk, M.; Sikora, P.; et al. Associations of MicroRNAs, Angiogenesis-Regulating Factors and CFH Y402H Polymorphism—An Attempt to Search for Systemic Biomarkers in Age-Related Macular Degeneration. Int. J. Mol. Sci. 2019, 20, 5750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshelmani, H.; Wride, M.A.; Saad, T.; Rani, S.; Kelly, D.J.; Keegan, D. Identification of Novel Serum MicroRNAs in Age-Related Macular Degeneration. Transl. Vis. Sci. Technol. 2020, 9, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshelmani, H.; Brennan, I.; Kelly, D.J.; Keegan, D. Differential Circulating MicroRNA Expression in Age-Related Macular Degeneration. Int. J. Mol. Sci. 2021, 22, 2321. [Google Scholar] [CrossRef]
- Elshelmani, H.; Wride, M.A.; Saad, T.; Rani, S.; Kelly, D.J.; Keegan, D. The Role of Deregulated MicroRNAs in Age-Related Macular Degeneration Pathology. Transl. Vis. Sci. Technol. 2021, 10, 12. [Google Scholar] [CrossRef]
- Strafella, C.; Errichiello, V.; Caputo, V.; Aloe, G.; Ricci, F.; Cusumano, A.; Novelli, G.; Giardina, E.; Cascella, R. The Interplay between MiRNA-Related Variants and Age-Related Macular Degeneration: Evidence of Association of MIR146A and MIR27A. Int. J. Mol. Sci. 2019, 20, 1578. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Gallagher, R.; Ufret-Vincenty, R.; Li, X.; Olson, E.N.; Wang, S. Regulation of Angiogenesis and Choroidal Neovascularization by Members of MicroRNA-23~27~24 Clusters. Proc. Natl. Acad. Sci. USA 2011, 108, 8287–8292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oltra, M.; Vidal-Gil, L.; Maisto, R.; Sancho-Pelluz, J.; Barcia, J.M. Oxidative Stress-induced Angiogenesis Is Mediated by MiR-205-5p. J. Cell. Mol. Med. 2020, 24, 1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ménard, C.; Wilson, A.M.; Dejda, A.; Miloudi, K.; Binet, F.; Crespo-Garcia, S.; Parinot, C.; Pilon, F.; Juneau, R.; Andriessen, E.M.; et al. MiR-106b Suppresses Pathological Retinal Angiogenesis. Aging 2020, 12, 24836. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lee, A.Y.W.; Wigg, J.P.; Peshavariya, H.; Liu, P.; Zhang, H. MiRNA Involvement in Angiogenesis in Age-Related Macular Degeneration. J. Physiol. Biochem. 2016, 72, 583–592. [Google Scholar] [CrossRef]
- Fuchs, H.R.; Meister, R.; Lotke, R.; Framme, C. The MicroRNAs MiR-302d and MiR-93 Inhibit TGFB-Mediated EMT and VEGFA Secretion from ARPE-19 Cells. Exp. Eye Res. 2020, 201, 108258. [Google Scholar] [CrossRef]
- Wang, L.; Lee, A.Y.W.; Wigg, J.P.; Peshavariya, H.; Liu, P.; Zhang, H. MiR-126 Regulation of Angiogenesis in Age-Related Macular Degeneration in CNV Mouse Model. Int. J. Mol. Sci. 2016, 17, 895. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Xie, P.; Sun, R.; Sun, X.; Liu, G.; Ding, S.; Zhu, M.; Yan, B.; Liu, Q.; Chen, X.; et al. C-Jun-Mediated MicroRNA-302d-3p Induces RPE Dedifferentiation by Targeting P21Waf1/Cip1. Cell Death Dis. 2018, 9, 451. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Anderson, C.; Zhang, H.; Li, X.; Inglis, F.; Jayagopal, A.; Wang, S. Repression of Choroidal Neovascularization through Actin Cytoskeleton Pathways by MicroRNA-24. Mol. Ther. 2014, 22, 378–389. [Google Scholar] [CrossRef] [Green Version]
- Grassmann, F.; Schoenberger, P.G.A.; Brandl, C.; Schick, T.; Hasler, D.; Meister, G.; Fleckenstein, M.; Lindner, M.; Helbig, H.; Fauser, S.; et al. A Circulating Microrna Profile Is Associated with Late-Stage Neovascular Age-Related Macular Degeneration. PLoS ONE 2014, 9, e107461. [Google Scholar] [CrossRef]
- Lin, J.B.; Moolani, H.V.; Sene, A.; Sidhu, R.; Kell, P.; Lin, J.B.; Dong, Z.; Ban, N.; Ory, D.S.; Apte, R.S. Macrophage MicroRNA-150 Promotes Pathological Angiogenesis as Seen in Age-Related Macular Degeneration. JCI Insight 2018, 3, e120157. [Google Scholar] [CrossRef]
- Lian, C.; Lou, H.; Zhang, J.; Tian, H.; Ou, Q.; Xu, J.Y.; Jin, C.; Gao, F.; Zhang, J.; Wang, J.; et al. MicroRNA-24 Protects Retina from Degeneration in Rats by down-Regulating Chitinase-3-like Protein 1. Exp. Eye Res. 2019, 188, 107791. [Google Scholar] [CrossRef]
- Cai, J.; Zhang, H.; Zhang, Y.F.; Zhou, Z.; Wu, S. MicroRNA-29 Enhances Autophagy and Cleanses Exogenous Mutant AB-Crystallin in Retinal Pigment Epithelial Cells. Exp. Cell Res. 2019, 374, 231–248. [Google Scholar] [CrossRef] [PubMed]
- Murad, N.; Kokkinaki, M.; Gunawardena, N.; Gunawan, M.S.; Hathout, Y.; Janczura, K.J.; Theos, A.C.; Golestaneh, N. MiR-184 Regulates Ezrin, LAMP-1 Expression, Affects Phagocytosis in Human Retinal Pigment Epithelium and Is Downregulated in Age-Related Macular Degeneration. FEBS J. 2014, 281, 5251–5264. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Lin, H.; Deng, X.; Li, S.; Zhang, J. MiR-1246 Promotes Anti-Apoptotic Effect of Mini-AA in Oxidative Stress-Induced Apoptosis in Retinal Pigment Epithelial Cells. Clin. Exp. Ophthalmol. 2020, 48, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Z.; Tang, Y.; Cheng, L.B.; Yao, J.; Jiang, Q.; Li, K.R.; Zhen, Y. fang Targeting Keap1 by MiR-626 Protects Retinal Pigment Epithelium Cells from Oxidative Injury by Activating Nrf2 Signaling. Free Radic. Biol. Med. 2019, 143, 387–396. [Google Scholar] [CrossRef]
- Garcia, T.Y.; Gutierrez, M.; Reynolds, J.; Lamba, D.A. Modeling the Dynamic AMD-Associated Chronic Oxidative Stress Changes in Human ESC and IPSC-Derived RPE Cells. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7480–7488. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Qian, J.; Castillo, A.C.; Long, B.; Keyes, K.T.; Chen, G.; Ye, Y. Effect of MiR-23 on Oxidant-Induced Injury in Human Retinal Pigment Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6308–6314. [Google Scholar] [CrossRef]
- Tasharrofi, N.; Kouhkan, F.; Soleimani, M.; Soheili, Z.S.; Kabiri, M.; Mahmoudi Saber, M.; Dorkoosh, F.A. Survival Improvement in Human Retinal Pigment Epithelial Cells via Fas Receptor Targeting by MiR-374a. J. Cell. Biochem. 2017, 118, 4854–4861. [Google Scholar] [CrossRef]
- Liu, J.; Ma, Z.; Ran, Z. MiR-21–3p Modulates Lipopolysaccharide-Induced Inflammation and Apoptosis via Targeting TGS4 in Retinal Pigment Epithelial Cells. Clin. Exp. Pharmacol. Physiol. 2019, 46, 883–889. [Google Scholar] [CrossRef]
- Yan, J.; Qin, Y.; Yu, J.; Peng, Q.; Chen, X. MiR-340/IASPP Axis Affects UVB-Mediated Retinal Pigment Epithelium (RPE) Cell Damage. J. Photochem. Photobiol. B Biol. 2018, 186, 9–16. [Google Scholar] [CrossRef]
- Berber, P.; Grassmann, F.; Kiel, C.; Weber, B.H.F. An Eye on Age-Related Macular Degeneration: The Role of MicroRNAs in Disease Pathology. Mol. Diagn. Ther. 2017, 21, 31. [Google Scholar] [CrossRef] [Green Version]
- Alexandrov, P.N.; Dua, P.; Lukiw, W.J. Up-Regulation of MiRNA-146a in Progressive, Age-Related Inflammatory Neurodegenerative Disorders of the Human CNS. Front. Neurol. 2014, 5, 181. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Irani, S.; Sirwi, A.; Hussain, M.M. MicroRNAs Regulating Apolipoprotein B-Containing Lipoprotein Production. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2016, 1861, 2062–2068. [Google Scholar] [CrossRef]
- Jun, S.; Datta, S.; Wang, L.; Pegany, R.; Cano, M.; Handa, J.T. The Impact of Lipids, Lipid Oxidation, and Inflammation on AMD, and the Potential Role of MiRNAs on Lipid Metabolism in the RPE. Exp. Eye Res. 2019, 181, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Toomey, C.B.; Kelly, U.; Saban, D.R.; Rickman, C.B. Regulation of Age-Related Macular Degeneration-like Pathology by Complement Factor H. Proc. Natl. Acad. Sci. USA 2015, 112, E3040–E3049. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.; Hu, W.; Wei, Q.; Cai, W.; Jin, H.; Yu, D.; Liu, C.; Shen, T.; Zhu, M.; Liang, X.; et al. MicroRNA-27a Promotes Oxidative-Induced RPE Cell Death through Targeting FOXO1. BioMed Res. Int. 2021, 2021, 6666506. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Koster, K.M.; He, Y.; Zhou, Q. MiRNAs as Potential Therapeutic Targets for Age-Related Macular Degeneration. Future Med. Chem. 2012, 4, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyttinen, J.M.T.; Blasiak, J.; Felszeghy, S.; Kaarniranta, K. MicroRNAs in the Regulation of Autophagy and Their Possible Use in Age-Related Macular Degeneration Therapy. Ageing Res. Rev. 2021, 67, 101260. [Google Scholar] [CrossRef]
- Gnanaguru, G.; Wagschal, A.; Oh, J.; Saez-Torres, K.L.; Li, T.; Temel, R.E.; Kleinman, M.E.; Näär, A.M.; D’Amore, P.A. Targeting of MiR-33 Ameliorates Phenotypes Linked to Age-Related Macular Degeneration. Mol. Ther. 2021, 29, 2281–2293. [Google Scholar] [CrossRef]
- Roblain, Q.; Louis, T.; Yip, C.; Baudin, L.; Struman, I.; Caolo, V.; Lambert, V.; Lecomte, J.; Noël, A.; Heymans, S. Intravitreal Injection of Anti-MiRs against MiR-142–3p Reduces Angiogenesis and Microglia Activation in a Mouse Model of Laser-Induced Choroidal Neovascularization. Aging 2021, 13, 12359. [Google Scholar] [CrossRef]
- Bajan, S.; Hutvagner, G. RNA-Based Therapeutics: From Antisense Oligonucleotides to MiRNAs. Cells 2020, 9, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Authors | Samples | AMD Group | Control Group | miRNA Expression |
|---|---|---|---|---|
| Ménard et al. (2016) [21] | Vitreous body | 13 patients with wet AMD | 13 patients | Increased miR-548a, miR146a-5p Decreased miR-106b, miR-152, miR-205 |
| Blood plasma | Increased miR-146a Decreased miR-106b, miR-152 | |||
| Bhattacharjee et al. (2016) [19] | Retina | 12 AMD retinas | 9 control | Increased miRNA-34a |
| Ren et al. (2017) [24] | Blood | 126 patients with AMD | 140 patients | Increased miR-27a-3p, miR-29b-3p and miR-195-5p |
| Romano et al. (2017) [17] | Blood serum | 11 patients with AMD | 11 patients | Increased miR-9, miR-23a, miR-27a, miR-34a, miR-126, and miR-146a Decreased miR-155 |
| Pogue et al. (2018) [20] | Retina | 17 AMD retinas | 10 control | Increased miR-7 and the Let-7 cluster, miR-23a and the miR-27a cluster, miR-9, miR-34a, miR-125b, miR-155, miR-146a |
| Blasiak et al. (2019) [22] | Blood | 76 patients with wet AMD | 70 patients | Decreased miR-34a-5p, miR-126-3p, miR-145-5p and miR-205-5p |
| Litwińska et al. (2019) [25] | Blood plasma | 354 patients with AMD (179 with wet AMD, 175 with dry AMD) | 121 patients | Wet AMD: increased miR-23a-3p, miR-30b, mir-191-5p, decreased miR-16-5p, miR-17-3p, miR-150-5p and miR-155-5p. Dry AMD: increased miR-23a-3p, miR-126-3p, miR-126-5p, miR-146a, miR-191-5p, decreased miR-16-5p, miR-17-3p and miR-17-5p |
| Strafella et al. (2019) [30] | Blood | 976 patients with wet AMD | 1000 patients | Polymorphisms in DNA—MIR146A and MIR27A are significantly associated with AMD |
| Ulańczyk et al. (2019) [26] | Blood plasma | 354 patients (175 patients with dry AMD, 179 patients with wet AMD) | 121 patients | Increased miR-23a-3p, miR-126-5p, miR-16-5p, miR-17-3p, miR-17-5p, miR-223-3p and miR-93 Decreased: miR-21-3p. miR-155-5p |
| Elbay et al. (2019) [23] | Blood—serum | 70 patients with wet AMD | 50 patients | Increased: miR-486-5p and miR-626 Decreased: miR-885-5p |
| ElShelmani et al. (2020) [27] | Blood | 60 patients (30 patients with dry AMD, 30 patients with wet AMD) | 30 patients | 46 miRNAs increased in wet AMD group |
| 4 miRNAs increased in dry AMD | ||||
| 7 miRNA increased in both groups | ||||
| Potential role of miR-126, miR-410, and miR-19a as biomarkers | ||||
| ElShelmani et al. (2021) [29] | Blood serum | 80 patients (40 with wet AMD, 40 with dry AMD | 40 patients | Overexpression of miR-126, miR-19a and miR-410 |
| ElShelmani at al. (2021) [28] | Blood serum | 26 patients (12 patients with dry AMD, 14 with wet AMD) | 10 patients | Increased in dry AMD: hsa-let-7a-5p, hsa-let-7d-5p, hsa-miR-23a-3p, hsa-miR-301a-3p, hsa-miR-361-5p, hsa-miR-27b-3p, hsa-miR-874-3p, hsa-miR-19b-1-5p |
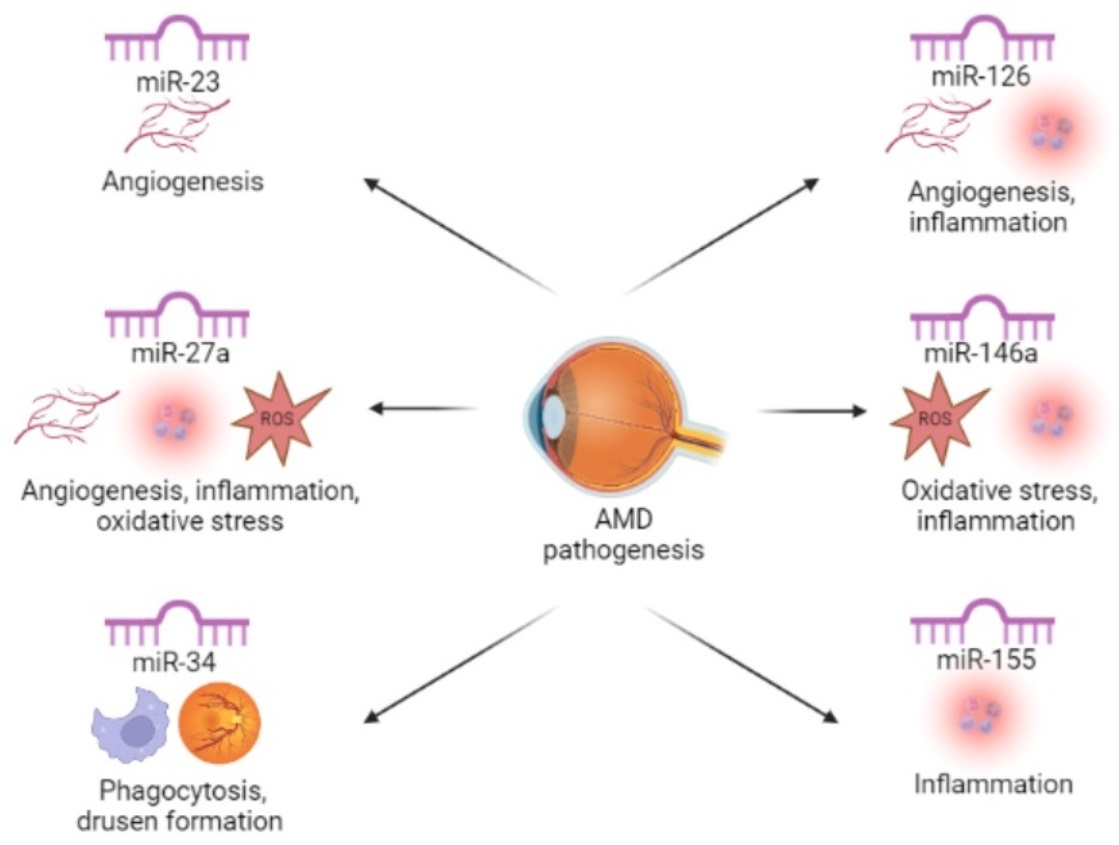
| The Most Frequently Dysregulated miRNAs | The Role in AMD Pathogenesis |
|---|---|
| miR-23 | angiogenesis |
| miR-27a | angiogenesis, inflammation, oxidative stress |
| miR-34 | phagocytosis, drusen formation |
| miR-126 | angiogenesis, inflammation |
| miR-146a | inflammation, oxidative stress |
| miR-155 | inflammation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbańska, K.; Stępień, P.W.; Nowakowska, K.N.; Stefaniak, M.; Osial, N.; Chorągiewicz, T.; Toro, M.D.; Nowomiejska, K.; Rejdak, R. The Role of Dysregulated miRNAs in the Pathogenesis, Diagnosis and Treatment of Age-Related Macular Degeneration. Int. J. Mol. Sci. 2022, 23, 7761. https://doi.org/10.3390/ijms23147761
Urbańska K, Stępień PW, Nowakowska KN, Stefaniak M, Osial N, Chorągiewicz T, Toro MD, Nowomiejska K, Rejdak R. The Role of Dysregulated miRNAs in the Pathogenesis, Diagnosis and Treatment of Age-Related Macular Degeneration. International Journal of Molecular Sciences. 2022; 23(14):7761. https://doi.org/10.3390/ijms23147761
Chicago/Turabian StyleUrbańska, Karolina, Piotr Witold Stępień, Katarzyna Natalia Nowakowska, Martyna Stefaniak, Natalia Osial, Tomasz Chorągiewicz, Mario Damiano Toro, Katarzyna Nowomiejska, and Robert Rejdak. 2022. "The Role of Dysregulated miRNAs in the Pathogenesis, Diagnosis and Treatment of Age-Related Macular Degeneration" International Journal of Molecular Sciences 23, no. 14: 7761. https://doi.org/10.3390/ijms23147761
APA StyleUrbańska, K., Stępień, P. W., Nowakowska, K. N., Stefaniak, M., Osial, N., Chorągiewicz, T., Toro, M. D., Nowomiejska, K., & Rejdak, R. (2022). The Role of Dysregulated miRNAs in the Pathogenesis, Diagnosis and Treatment of Age-Related Macular Degeneration. International Journal of Molecular Sciences, 23(14), 7761. https://doi.org/10.3390/ijms23147761