
Role of Group I Metabotropic Glutamate Receptors in Spike Timing-Dependent Plasticity




{kind=link}
Abstract
:1. Introduction
2. The STDP Phenomena and mGluRs Involvement
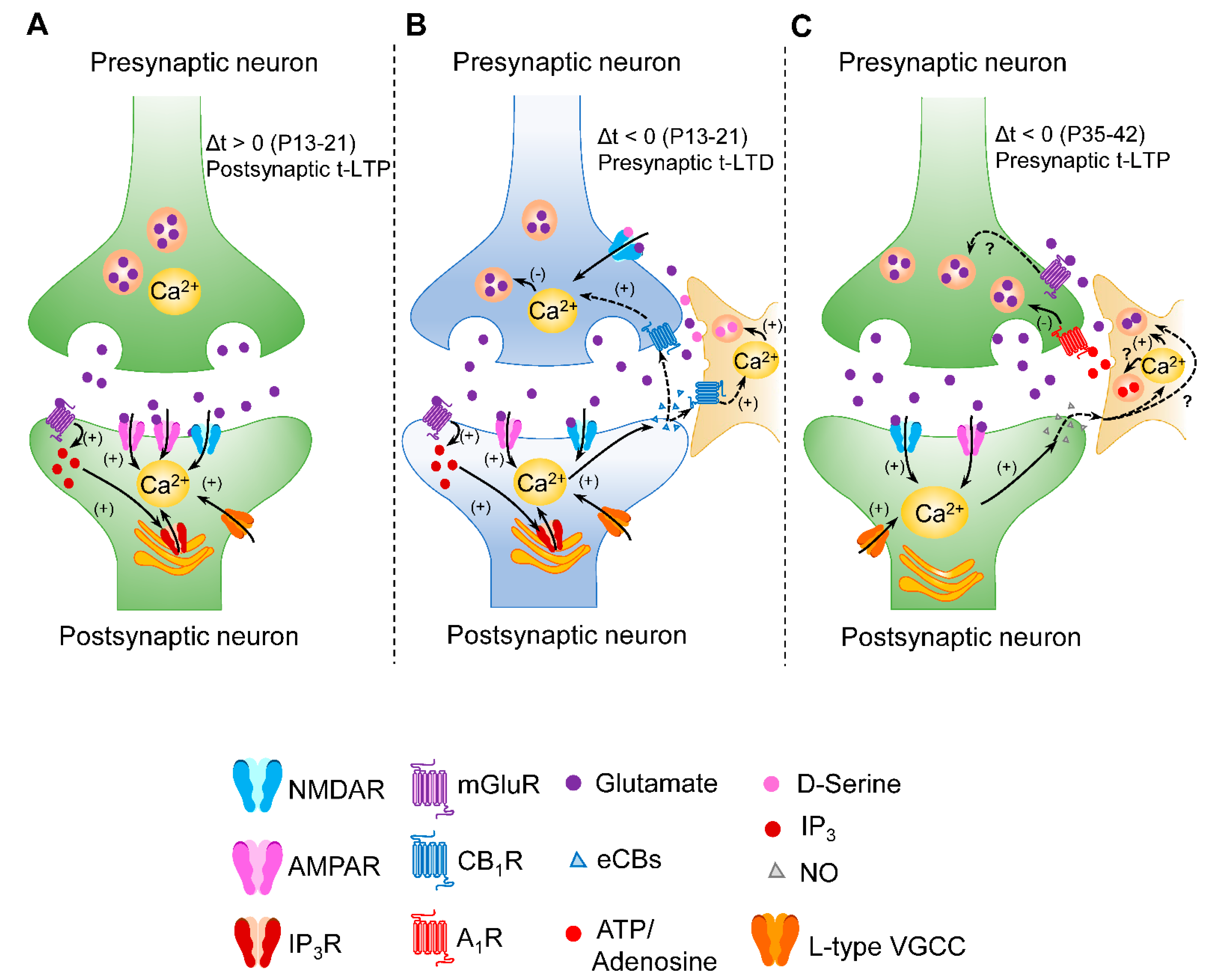
3. Group I mGluR Involvement in the Coincidence Detection for STDP
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bodzęta, A.; Scheefhals, N.; MacGillavry, H.D. Membrane trafficking and positioning of mGluRs at presynaptic and postsynaptic sites of excitatory synapses. Neuropharmacology 2021, 200, 108799. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, S. Molecular Diversity of Glutamate Receptors and Implications for Brain Function. Science 1992, 258, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Kroon, T.; Dawitz, J.; Kramvis, I.; Anink, J.; Obermayer, J.; Verhoog, M.B.; Wilbers, R.; Goriounova, N.A.; Idema, S.; Baayen, J.C.; et al. Group I mGluR-mediated activation of martinotti cells inhibits local cortical circuitry in human cortex. Front. Cell. Neurosci. 2019, 13, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niswender, C.M.; Conn, P.J. Metabotropic glutamate receptors: physiology, pharmacology, and disease. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 295–322. [Google Scholar] [CrossRef] [Green Version]
- Collingridge, G.L.; Abraham, W.C. Glutamate receptors and synaptic plasticity: The impact of Evans and Watkins. Neuropharmacology 2021, 206, 108922. [Google Scholar] [CrossRef]
- Reiner, A.; Levitz, J. Glutamatergic Signaling in the Central Nervous System: Ionotropic and Metabotropic Receptors in Concert. Neuron 2018, 98, 1080–1098. [Google Scholar] [CrossRef] [Green Version]
- Mateos-Aparicio, P.; Rodríguez-Moreno, A. The Impact of Studying Brain Plasticity. Front. Cell. Neurosci. 2019, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Dolphin, A.C.; Errington, M.L.; Bliss, T. V Long-term potentiation of the perforant path in vivo is associated with increased glutamate release. Nature 1982, 297, 496–498. [Google Scholar] [CrossRef]
- Bliss, T.V.P.; Collingridge, G.L. A synaptic model of memory: long-term potentiation in the hippocampus. Nature 1993, 361, 31–39. [Google Scholar] [CrossRef]
- Bi, G.; Poo, M. Synaptic Modifications in Cultured Hippocampal Neurons: Dependence on Spike Timing, Synaptic Strength, and Postsynaptic Cell Type. J. Neurosci. 1998, 18, 10464–10472. [Google Scholar] [CrossRef]
- Malenka, R.C.; Bear, M.F. LTP and LTD: an embarrassment of riches. Neuron 2004, 44, 5–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neves, G.; Cooke, S.F.; Bliss, T.V.P. Synaptic plasticity, memory and the hippocampus: a neural network approach to causality. Nat. Rev. Neurosci. 2008, 9, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Talavera, Y.; Rodríguez-Moreno, A. Synaptic Plasticity and Oscillations in Alzheimer’s Disease: A Complex Picture of a Multifaceted Disease. Front. Mol. Neurosci. 2021, 14, 696476. [Google Scholar] [CrossRef] [PubMed]
- Park, P.; Kang, H.; Sanderson, T.M.; Bortolotto, Z.A.; Georgiou, J.; Zhuo, M.; Kaang, B.K.; Collingridge, G.L. The Role of Calcium-Permeable AMPARs in Long-Term Potentiation at Principal Neurons in the Rodent Hippocampus. Front. Synaptic Neurosci. 2018, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bashir, Z.I.; Bortolotto, Z.A.; Davies, C.H.; Berretta, N.; Irving, A.J.; Seal, A.J.; Henley, J.M.; Jane, D.E.; Watkins, J.C.; Collingridge, G.L. Induction of LTP in the hippocampus needs synaptic activation of glutamate metabotropic receptors. Nature 1993, 363, 347–350. [Google Scholar] [CrossRef]
- Bortolotto, Z.A.; Bashir, Z.I.; Davies, C.H.; Collingridge, G.L. A molecular switch activated by metabotropic glutamate receptors regulates induction of long-term potentiation. Nature 1994, 368, 740–743. [Google Scholar] [CrossRef]
- Tigaret, C.M.; Chamberlain, S.E.L.; Sadowski, J.H.L.P.; Hall, J.; Ashby, M.C.; Mellor, J.R. Convergent Metabotropic Signaling Pathways Inhibit SK Channels to Promote Synaptic Plasticity in the Hippocampus. J. Neurosci. 2018, 38, 9252–9262. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, K.; Manabe, T.; Takahashi, T. Presynaptic Long-Term Depression at the Hippocampal Mossy Fiber—CA3 Synapse. Science 1996, 273, 648–650. [Google Scholar] [CrossRef]
- Negrete-Díaz, J.V.; Sihra, T.S.; Delgado-García, J.M.; Rodríguez-Moreno, A. Kainate receptor-mediated presynaptic inhibition converges with presynaptic inhibition mediated by Group II mGluRs and long-term depression at the hippocampal mossy fiber-CA3 synapse. J. Neural Transm. 2007, 114, 1425–1431. [Google Scholar] [CrossRef]
- Lyon, L.; Borel, M.; Carrión, M.; Kew, J.N.C.; Corti, C.; Harrison, P.J.; Burnet, P.W.J.; Paulsen, O.; Rodríguez-Moreno, A. Hippocampal mossy fiber long-term depression in Grm2/3 double knockout mice. Synapse 2011, 65, 945–954. [Google Scholar] [CrossRef]
- Mercier, M.S.; Lodge, D. Group III metabotropic glutamate receptors: pharmacology, physiology and therapeutic potential. Neurochem. Res. 2014, 39, 1876–1894. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A.; Lim, Y.J.; Kumar, K.; Baby, N.; Pang, K.L.K.; Benoy, A.; Behnisch, T.; Sajikumar, S. Group III metabotropic glutamate receptors gate long-term potentiation and synaptic tagging/capture in rat hippocampal area CA2. Elife 2020, 9, e55344. [Google Scholar] [CrossRef]
- Lüscher, C.; Huber, K.M. Group 1 mGluR-Dependent Synaptic Long-Term Depression: Mechanisms and Implications for Circuitry and Disease. Neuron 2010, 65, 445–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, S.M. The function of metabotropic glutamate receptors in thalamus and cortex. Neuroscientist 2014, 20, 136–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.F.; Daw, N.W. Metabotropic glutamate receptors potentiate responses to NMDA and AMPA from layer V cells in rat visual cortex. J. Neurophysiol. 1996, 76, 808–815. [Google Scholar] [CrossRef]
- Mannaioni, G.; Marino, M.J.; Valenti, O.; Traynelis, S.F.; Conn, P.J. Metabotropic glutamate receptors 1 and 5 differentially regulate CA1 pyramidal cell function. J. Neurosci. 2001, 21, 5925–5934. [Google Scholar] [CrossRef] [Green Version]
- Guérineau, N.C.; Bossu, J.L.; Gähwiler, B.H.; Gerber, U. Activation of a nonselective cationic conductance by metabotropic glutamatergic and muscarinic agonists in CA3 pyramidal neurons of the rat hippocampus. J. Neurosci. 1995, 15, 4395–4407. [Google Scholar] [CrossRef]
- Guérineau, N.C.; Gähwiler, B.H.; Gerber, U. Reduction of resting K+ current by metabotropic glutamate and muscarinic receptors in rat CA3 cells: mediation by G-proteins. J. Physiol. 1994, 474, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Jones, O.D. Do group I metabotropic glutamate receptors mediate LTD? Neurobiol. Learn. Mem. 2017, 138, 85–97. [Google Scholar] [CrossRef]
- Feldman, D.E. The Spike-Timing Dependence of Plasticity. Neuron 2012, 75, 556–571. [Google Scholar] [CrossRef] [Green Version]
- Corlew, R.; Brasier, D.J.; Feldman, D.E.; Philpot, B.D. Presynaptic NMDA receptors: newly appreciated roles in cortical synaptic function and plasticity. Neuroscientist 2008, 14, 609–625. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Moreno, A.; Banerjee, A.; Paulsen, O. Presynaptic NMDA Receptors and Spike Timing-Dependent Depression at Cortical Synapses. Front. Synaptic Neurosci. 2010, 2, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markram, H.; Gerstner, W.; Sjöström, P.J. Spike-Timing-Dependent Plasticity: A Comprehensive Overview. Front. Synaptic Neurosci. 2012, 4, 2010–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade-Talavera, Y.; Duque-Feria, P.; Paulsen, O.; Rodríguez-Moreno, A. Presynaptic Spike Timing-Dependent Long-Term Depression in the Mouse Hippocampus. Cereb. Cortex 2016, 26, 3637–3654. [Google Scholar] [CrossRef] [Green Version]
- Min, R.; Nevian, T. Astrocyte signaling controls spike timing-dependent depression at neocortical synapses. Nat. Neurosci. 2012, 15, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; González-Rueda, A.; Sampaio-Baptista, C.; Paulsen, O.; Rodríguez-Moreno, A. Distinct mechanisms of spike timing-dependent LTD at vertical and horizontal inputs onto L2/3 pyramidal neurons in mouse barrel cortex. Physiol. Rep. 2014, 2, e00271. [Google Scholar] [CrossRef]
- Fino, E.; Paille, V.; Cui, Y.; Morera-Herreras, T.; Deniau, J.-M.; Venance, L. Distinct coincidence detectors govern the corticostriatal spike timing-dependent plasticity. J. Physiol. 2010, 588, 3045–3062. [Google Scholar] [CrossRef] [PubMed]
- Sgritta, M.; Locatelli, F.; Soda, T.; Prestori, F.; D’Angelo, E.U. Hebbian Spike-Timing Dependent Plasticity at the Cerebellar Input Stage. J. Neurosci. 2017, 37, 2809–2823. [Google Scholar] [CrossRef]
- Banerjee, A.; Meredith, R.M.; Rodríguez-Moreno, A.; Mierau, S.B.; Auberson, Y.P.; Paulsen, O. Double dissociation of spike timing-dependent potentiation and depression by subunit-preferring NMDA receptor antagonists in mouse barrel cortex. Cereb. Cortex 2009, 19, 2959–2969. [Google Scholar] [CrossRef]
- Duguid, I.; Sjöström, P.J. Novel presynaptic mechanisms for coincidence detection in synaptic plasticity. Curr. Opin. Neurobiol. 2006, 16, 312–322. [Google Scholar] [CrossRef]
- Kwag, J.; Paulsen, O. The timing of external input controls the sign of plasticity at local synapses. Nat. Neurosci. 2009, 12, 1219–1221. [Google Scholar] [CrossRef] [PubMed]
- Brzosko, Z.; Schultz, W.; Paulsen, O. Retroactive modulation of spike timing-dependent plasticity by dopamine. Elife 2015, 4, e09685. [Google Scholar] [CrossRef]
- Brzosko, Z.; Mierau, S.B.; Paulsen, O. Neuromodulation of Spike-Timing-Dependent Plasticity: Past, Present, and Future. Neuron 2019, 103, 563–581. [Google Scholar] [CrossRef] [PubMed]
- Falcón-Moya, R.; Pérez-Rodríguez, M.; Prius-Mengual, J.; Andrade-Talavera, Y.; Arroyo-García, L.E.; Pérez-Artés, R.; Mateos-Aparicio, P.; Guerra-Gomes, S.; Oliveira, J.F.; Flores, G.; et al. Astrocyte-mediated switch in spike timing-dependent plasticity during hippocampal development. Nat. Commun. 2020, 11, 4388. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Gallego, I.; Pérez-Rodríguez, M.; Coatl-Cuaya, H.; Flores, G.; Rodríguez-Moreno, A. Adenosine and astrocytes determine the developmental dynamics of spike timing-dependent plasticity in the somatosensory cortex. J. Neurosci. 2022. ahead of print. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, M.; Arroyo-García, L.E.; Prius-Mengual, J.; Andrade-Talavera, Y.; Armengol, J.A.; Pérez-Villegas, E.M.; Duque-Feria, P.; Flores, G.; Rodríguez-Moreno, A. Adenosine Receptor-Mediated Developmental Loss of Spike Timing-Dependent Depression in the Hippocampus. Cereb. Cortex 2019, 29, 3266–3281. [Google Scholar] [CrossRef] [Green Version]
- Bender, V.A.; Bender, K.J.; Brasier, D.J.; Feldman, D.E. Two coincidence detectors for spike timing-dependent plasticity in somatosensory cortex. J. Neurosci. 2006, 26, 4166–4177. [Google Scholar] [CrossRef]
- Brasier, D.J.; Feldman, D.E. Synapse-Specific Expression of Functional Presynaptic NMDA Receptors in Rat Somatosensory Cortex. J. Neurosci. 2008, 28, 2199–2211. [Google Scholar] [CrossRef]
- Rodríguez-Moreno, A.; Paulsen, O. Spike timing-dependent long-term depression requires presynaptic NMDA receptors. Nat. Neurosci. 2008, 11, 744–745. [Google Scholar] [CrossRef]
- Jung, S.J.; Kim, S.J.; Park, Y.K.; Oh, S.B.; Cho, K.; Kim, J. Group I mGluR regulates the polarity of spike-timing dependent plasticity in substantia gelatinosa neurons. Biochem. Biophys. Res. Commun. 2006, 347, 509–516. [Google Scholar] [CrossRef]
- Jung, S.-Y.; Kim, J.; Kwon, O. Bin; Jung, J.H.; An, K.; Jeong, A.Y.; Lee, C.J.; Choi, Y.-B.; Bailey, C.H.; Kandel, E.R.; et al. Input-specific synaptic plasticity in the amygdala is regulated by neuroligin-1 via postsynaptic NMDA receptors. Proc. Natl. Acad. Sci. USA 2010, 107, 4710–4715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.T.; Li, C.Y.; Zhao, J.P.; Poo, M.M.; Zhang, X.H. Spike-timing-dependent plasticity of neocortical excitatory synapses on inhibitory interneurons depends on target cell type. J. Neurosci. 2007, 27, 9711–9720. [Google Scholar] [CrossRef] [PubMed]
- Shipman, M.L.; Madasu, S.C.; Morielli, A.D.; Green, J.T. Intracerebellar infusion of an mGluR1/5 agonist enhances eyeblink conditioning. Behav. Neurosci. 2021, 135, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Kwag, J.; Paulsen, O. Gating of NMDA receptor-mediated hippocampal spike timing-dependent potentiation by mGluR5. Neuropharmacology 2012, 63, 701–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, H.H.W.; Rannio, S.; Jones, V.; Thomazeau, A.; Sjöström, P.J. NMDA receptors in axons: There’s no coincidence. J. Physiol. 2021, 599, 367–387. [Google Scholar] [CrossRef]
- Bi, G.-Q.; Rubin, J. Timing in synaptic plasticity: from detection to integration. Trends Neurosci. 2005, 28, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Froemke, R.C.; Poo, M.-M.; Dan, Y. Spike-timing-dependent synaptic plasticity depends on dendritic location. Nature 2005, 434, 221–225. [Google Scholar] [CrossRef]
- Nishiyama, M.; Hong, K.; Mikoshiba, K.; Poo, M.M.; Kato, K. Calcium stores regulate the polarity and input specificity of synaptic modification. Nature 2000, 408, 584–588. [Google Scholar] [CrossRef]
- Shouval, H.Z.; Bear, M.F.; Cooper, L.N. A unified model of NMDA receptor-dependent bidirectional synaptic plasticity. Proc. Natl. Acad. Sci. USA 2002, 99, 10831–10836. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Moreno, A.; Kohl, M.M.; Reeve, J.E.; Eaton, T.R.; Collins, H.A.; Anderson, H.L.; Paulsen, O.; Rodríguez-Moreno, A.; Kohl, M.M.; Reeve, J.E.; et al. Presynaptic induction and expression of timing-dependent long-term depression demonstrated by compartment-specific photorelease of a use-dependent NMDA receptor antagonist. J. Neurosci. 2011, 31, 8564–8569. [Google Scholar] [CrossRef]
- Egger, V.; Feldmeyer, D.; Sakmann, B. Coincidence detection and changes of synaptic efficacy in spiny stellate neurons in rat barrel cortex. Nat. Neurosci. 1999, 2, 1098–1105. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, G.; Larsen, R.S.; Rodríguez-Moreno, A.; Paulsen, O.; Sjöström, P.J. Towards resolving the presynaptic NMDA receptor debate. Curr. Opin. Neurobiol. 2018, 51, 1–7. [Google Scholar] [CrossRef]
- Pérez-Otaño, I.; Rodríguez-Moreno, A. Presynaptic NMDARs and astrocytes ally to control circuit-specific information flow. Proc. Natl. Acad. Sci. USA 2019, 116, 13166–13168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hlavackova, V.; Zabel, U.; Frankova, D.; Bätz, J.; Hoffmann, C.; Prezeau, L.; Pin, J.-P.; Blahos, J.; Lohse, M.J. Sequential inter- and intrasubunit rearrangements during activation of dimeric metabotropic glutamate receptor 1. Sci. Signal. 2012, 5, ra59. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. Neuronal calcium signaling. Neuron 1998, 21, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Inglebert, Y.; Debanne, D. Calcium and Spike Timing-Dependent Plasticity. Front. Cell. Neurosci. 2021, 15, 727336. [Google Scholar] [CrossRef]
- Nevian, T.; Sakmann, B. Spine Ca2+ Signaling in Spike-Timing-Dependent Plasticity. J. Neurosci. 2006, 26, 11001–11013. [Google Scholar] [CrossRef] [Green Version]
- Mateos-Aparicio, P.; Rodríguez-Moreno, A. Calcium Dynamics and Synaptic Plasticity. In Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2020; pp. 965–984. ISBN 978-3-030-12457-1. [Google Scholar]
- Cepeda-Prado, E.A.; Khodaie, B.; Quiceno, G.D.; Beythien, S.; Edelmann, E.; Lessmann, V. Calcium-Permeable AMPA Receptors Mediate Timing-Dependent LTP Elicited by Low Repeat Coincident Pre- and Postsynaptic Activity at Schaffer Collateral-CA1 Synapses. Cereb. Cortex 2021, 32, 1682–1703. [Google Scholar] [CrossRef]
- Hashimotodani, Y.; Ohno-Shosaku, T.; Tsubokawa, H.; Ogata, H.; Emoto, K.; Maejima, T.; Araishi, K.; Shin, H.-S.; Kano, M. Phospholipase Cbeta serves as a coincidence detector through its Ca2+ dependency for triggering retrograde endocannabinoid signal. Neuron 2005, 45, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Sabatini, B.L.; Maravall, M.; Svoboda, K. Ca(2+) signaling in dendritic spines. Curr. Opin. Neurobiol. 2001, 11, 349–356. [Google Scholar] [CrossRef]
- Bloodgood, B.L.; Sabatini, B.L. Ca(2+) signaling in dendritic spines. Curr. Opin. Neurobiol. 2007, 17, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Kreitzer, A.C.; Malenka, R.C. Dopamine modulation of state-dependent endocannabinoid release and long-term depression in the striatum. J. Neurosci. 2005, 25, 10537–10545. [Google Scholar] [CrossRef] [PubMed]
- Lorenzon, P.; Zacchetti, D.; Codazzi, F.; Fumagalli, G.; Meldolesi, J.; Grohovaz, F. Ca2+ waves in PC12 neurites: a bidirectional, receptor-oriented form of Ca2+ signaling. J. Cell Biol. 1995, 129, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Moreno, A.; Sistiaga, A.; Lerma, J.; Sánchez-Prieto, J. Switch from facilitation to inhibition of excitatory synaptic transmission by group I mGluR desensitization. Neuron 1998, 21, 1477–1486. [Google Scholar] [CrossRef]
- Porter, J.T.; McCarthy, K.D. Hippocampal astrocytes in situ respond to glutamate released from synaptic terminals. J. Neurosci. 1996, 16, 5073–5081. [Google Scholar] [CrossRef] [Green Version]
- Perea, G.; Araque, A. Astrocytes Potentiate Transmitter Release at Single Hippocampal Synapses. Science 2007, 317, 1083–1086. [Google Scholar] [CrossRef]
- Buzsáki, G.; Draguhn, A. Neuronal oscillations in cortical networks. Science 2004, 304, 1926–1929. [Google Scholar] [CrossRef] [Green Version]
- McBain, C.J.; Fisahn, A. Interneurons unbound. Nat. Rev. Neurosci. 2001, 2, 11–23. [Google Scholar] [CrossRef]
- Gray, E.E.; Murphy, J.G.; Liu, Y.; Trang, I.; Tabor, G.T.; Lin, L.; Hoffman, D.A. Disruption of GpI mGluR-Dependent Cav2.3 Translation in a Mouse Model of Fragile X Syndrome. J. Neurosci. 2019, 39, 7453–7464. [Google Scholar] [CrossRef]
- Crupi, R.; Impellizzeri, D.; Cuzzocrea, S. Role of Metabotropic Glutamate Receptors in Neurological Disorders. Front. Mol. Neurosci. 2019, 12, 20. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Gallego, I.; Rodríguez-Moreno, A.; Andrade-Talavera, Y. Role of Group I Metabotropic Glutamate Receptors in Spike Timing-Dependent Plasticity. Int. J. Mol. Sci. 2022, 23, 7807. https://doi.org/10.3390/ijms23147807
Martínez-Gallego I, Rodríguez-Moreno A, Andrade-Talavera Y. Role of Group I Metabotropic Glutamate Receptors in Spike Timing-Dependent Plasticity. International Journal of Molecular Sciences. 2022; 23(14):7807. https://doi.org/10.3390/ijms23147807
Chicago/Turabian StyleMartínez-Gallego, Irene, Antonio Rodríguez-Moreno, and Yuniesky Andrade-Talavera. 2022. "Role of Group I Metabotropic Glutamate Receptors in Spike Timing-Dependent Plasticity" International Journal of Molecular Sciences 23, no. 14: 7807. https://doi.org/10.3390/ijms23147807
APA StyleMartínez-Gallego, I., Rodríguez-Moreno, A., & Andrade-Talavera, Y. (2022). Role of Group I Metabotropic Glutamate Receptors in Spike Timing-Dependent Plasticity. International Journal of Molecular Sciences, 23(14), 7807. https://doi.org/10.3390/ijms23147807