
Low Doses of PFOA Promote Prostate and Breast Cancer Cells Growth through Different Pathways

 and
and 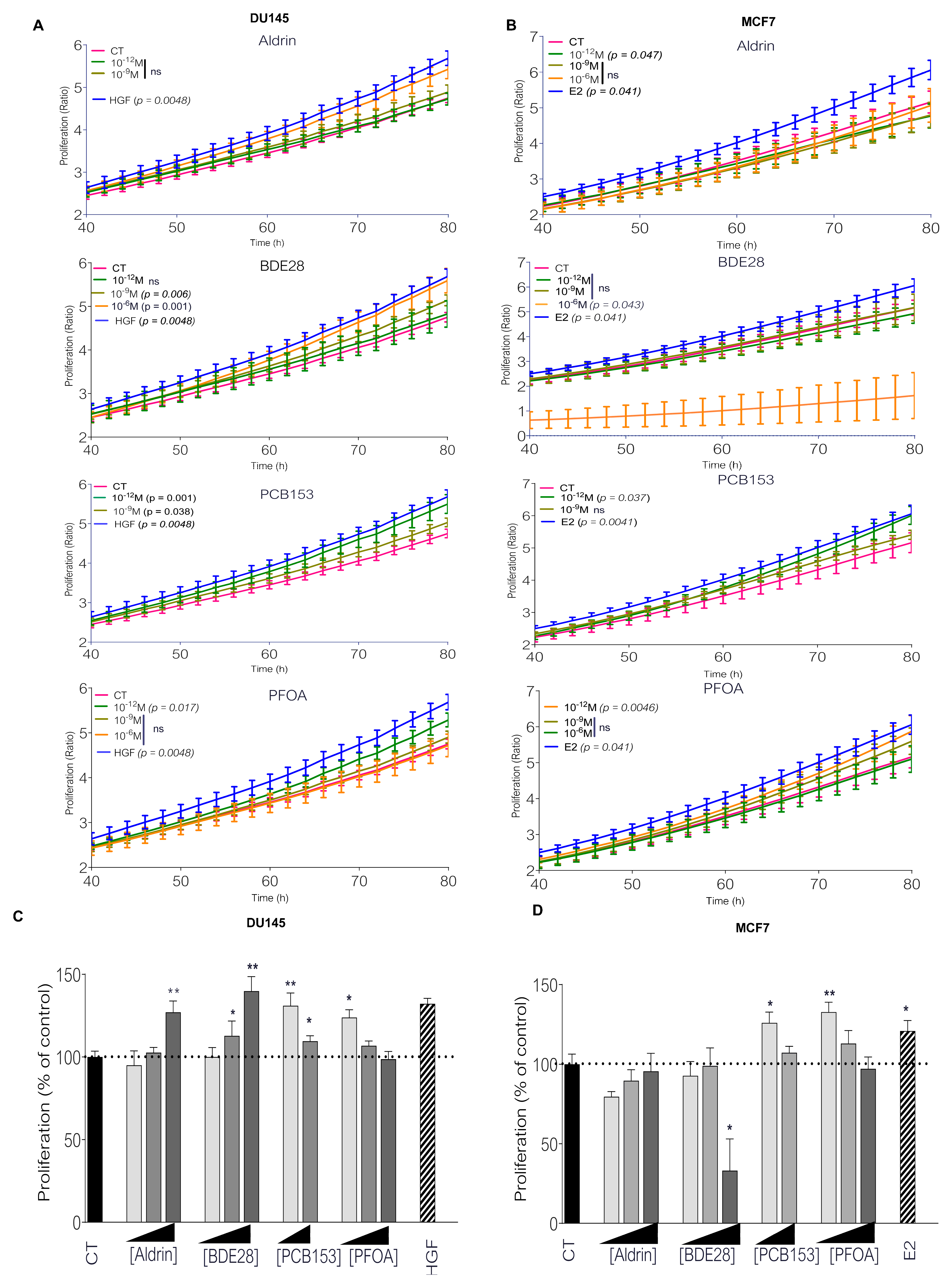{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
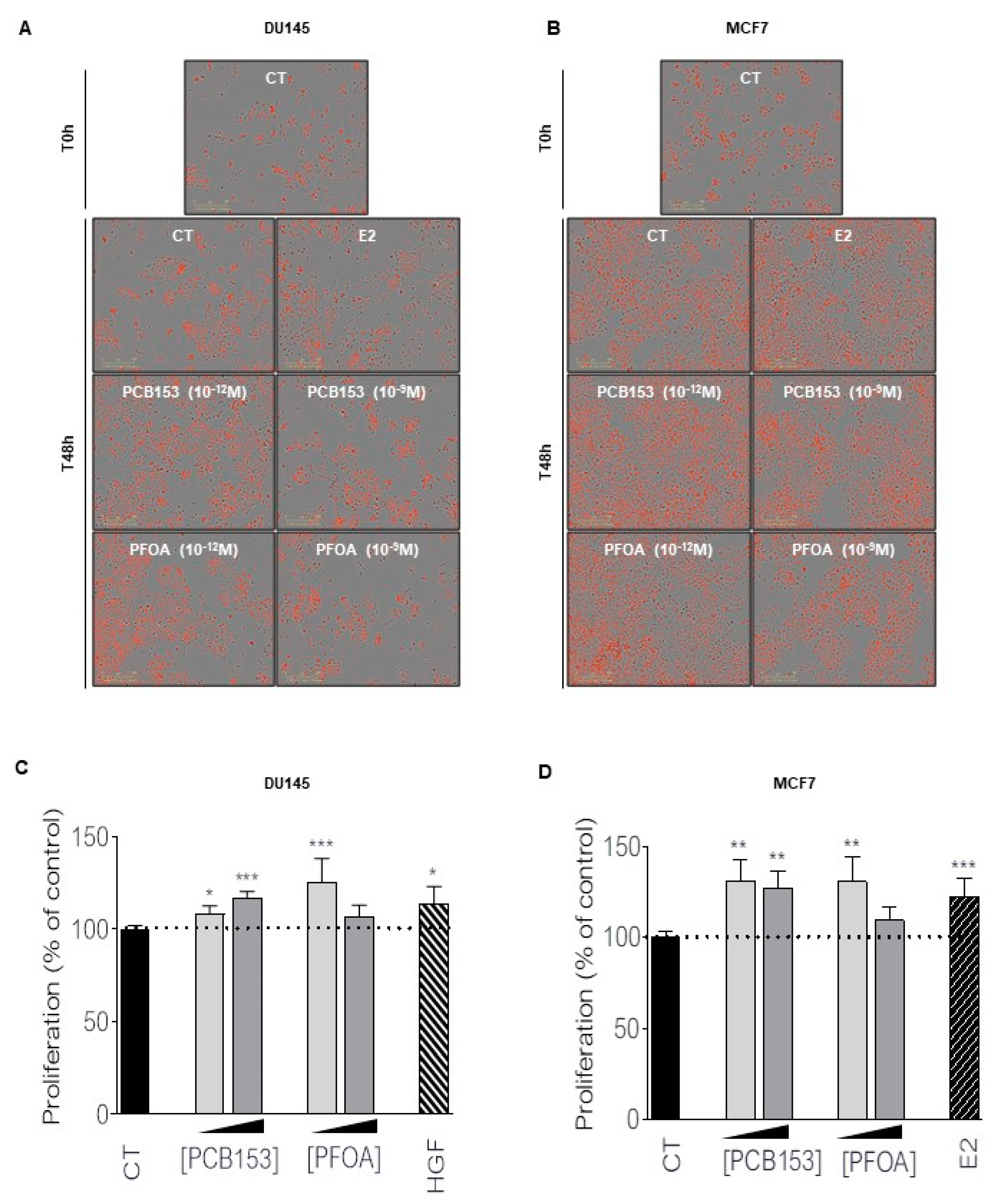
2.1. Picomolar Concentrations of PFOA and PCB153 Increase Prostate and Breast Cancer Cell Proliferation
2.2. The Proliferation Induced by PFOA Is Mediated by Plexin D1 in Prostate Cancer Cells
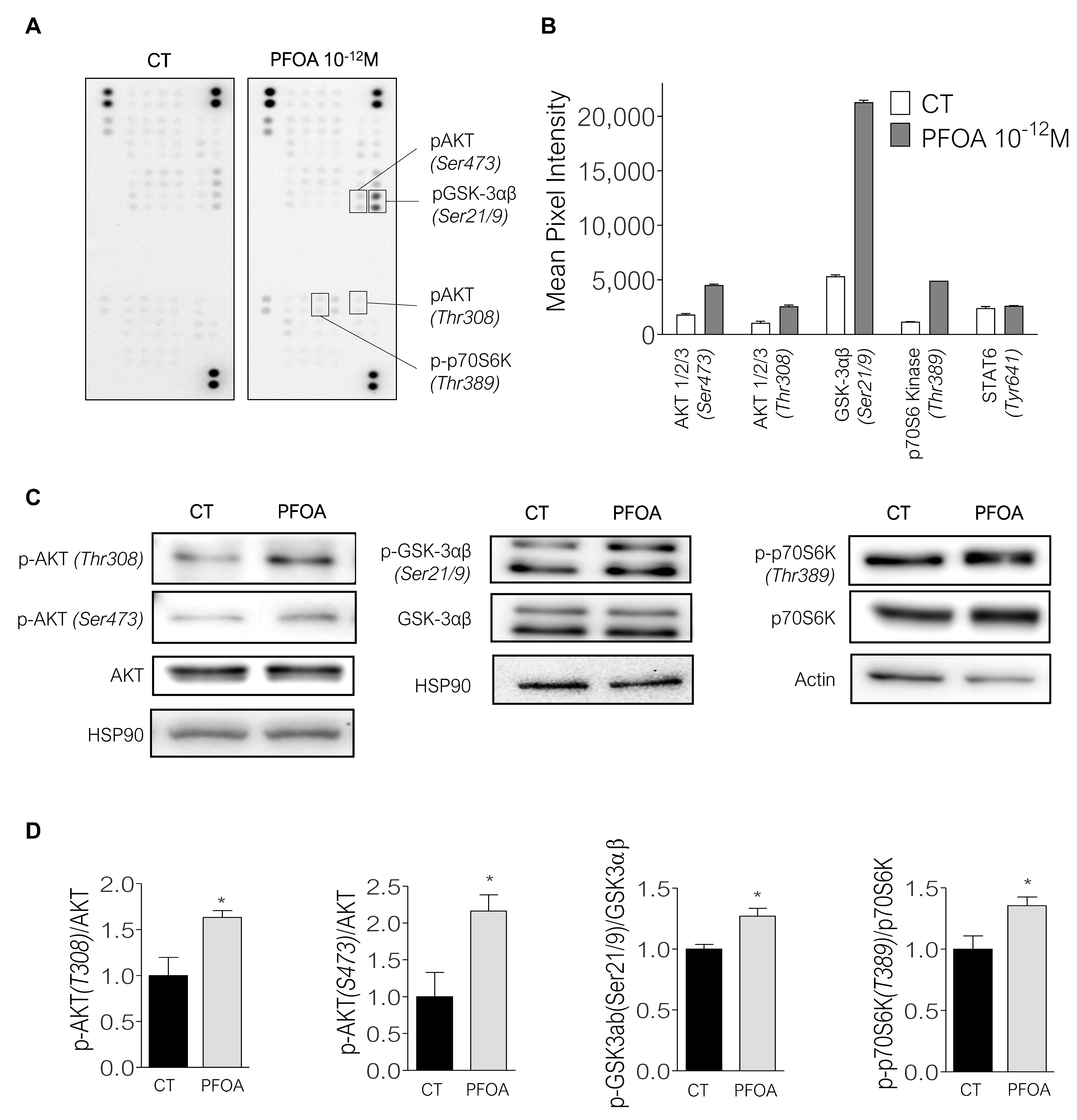
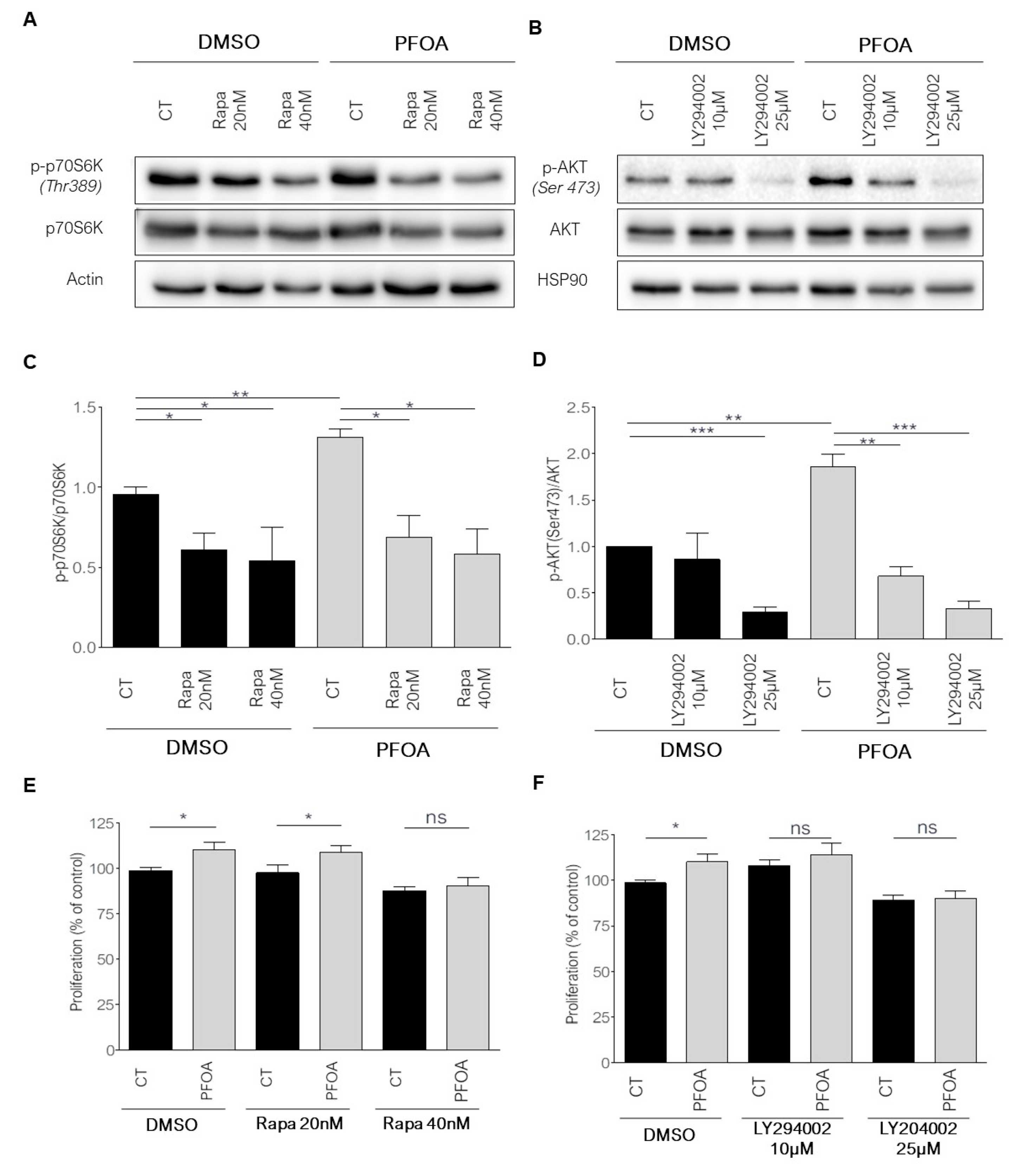
2.3. Breast Cancer Cells Proliferation Induced by Picomolar Concentrations of PFOA Is Dependent on the PI3K/Akt and mTORC1 Pathways
3. Discussion
4. Materials and Methods
4.1. Material
4.2. Cell Culture
4.3. Transfection
4.4. IncuCyte® Assay
4.5. BrdU Cell Proliferation Assay
4.6. Human Phospho-Kinase Array
4.7. Western Blot
4.8. Microarray Experiment
4.9. RNA Extraction and RT-qPCR
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Dyba, T.; Randi, G.; Bettio, M.; Gavin, A.; Visser, O.; Bray, F. Cancer incidence and mortality patterns in Europe: Estimates for 40 countries and 25 major cancers in 2018. Eur. J. Cancer 2018, 103, 356–387. [Google Scholar] [CrossRef] [PubMed]
- Hanin, L.; Pavlova, L. A quantitative insight into metastatic relapse of breast cancer. J. Theor. Biol. 2016, 394, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Holmes, D. Breast cancer: Increased risk with concurrent dietary and EDC exposures. Nat. Rev. Endocrinol. 2017, 13, 378. [Google Scholar] [CrossRef] [PubMed]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef] [PubMed]
- Koual, M.; Cano-Sancho, G.; Bats, A.S.; Tomkiewicz, C.; Kaddouch-Amar, Y.; Douay-Hauser, N.; Ngo, C.; Bonsang, H.; Delomenie, M.; Lecuru, F.; et al. Associations between persistent organic pollutants and risk of breast cancer metastasis. Environ. Int. 2019, 132, 105028. [Google Scholar] [CrossRef]
- Moore, R.W.; Fritz, W.A.; Schneider, A.J.; Lin, T.M.; Branam, A.M.; Safe, S.; Peterson, R.E. 2,3,7,8-Tetrachlorodibenzo-p-dioxin has both pro-carcinogenic and anti-carcinogenic effects on neuroendocrine prostate carcinoma formation in TRAMP mice. Toxicol. Appl. Pharmacol. 2016, 305, 242–249. [Google Scholar] [CrossRef] [Green Version]
- Nedellec, V.; Rabl, A.; Dab, W. Public health and chronic low chlordecone exposure in Guadeloupe, Part 1: Hazards, exposure-response functions, and exposures. Environ. Health 2016, 15, 75. [Google Scholar] [CrossRef] [Green Version]
- Burks, H.; Pashos, N.; Martin, E.; McLachlan, J.; Bunnell, B.; Burow, M. Endocrine disruptors and the tumor microenvironment: A new paradigm in breast cancer biology. Mol. Cell. Endocrinol. 2017, 457, 13–19. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Giari, L.; Guerranti, C.; Tognon, M.; Castaldelli, G.; Fano, E.A.; Martini, F. Environmental doses of perfluorooctanoic acid change the expression of genes in target tissues of common carp. Environ. Toxicol. Chem. 2018, 37, 942–948. [Google Scholar] [CrossRef]
- Dirinck, E.; Dirtu, A.C.; Jorens, P.G.; Malarvannan, G.; Covaci, A.; Van Gaal, L.F. Pivotal Role for the Visceral Fat Compartment in the Release of Persistent Organic Pollutants During Weight Loss. J. Clin. Endocrinol. Metab. 2015, 100, 4463–4471. [Google Scholar] [CrossRef] [Green Version]
- Fénichel, P.; Coquillard, P.; Brucker-Davis, F.; Marchand, P.; Cano-Sancho, G.; Boda, M.; Antignac, J.P.; Iannelli, A.; Gugenheim, J.; Le Bizec, B.; et al. Sustained bloodstream release of persistent organic pollutants induced by extensive weight loss after bariatric surgery: Implications for women of childbearing age. Environ. Int. 2021, 151, 106400. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Marchand, P.; Henegar, C.; Antignac, J.P.; Alili, R.; Poitou, C.; Bouillot, J.L.; Basdevant, A.; Le Bizec, B.; Barouki, R.; et al. Fate and complex pathogenic effects of dioxins and polychlorinated biphenyls in obese subjects before and after drastic weight loss. Environ. Health Perspect. 2011, 119, 377–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaPensee, E.W.; LaPensee, C.R.; Fox, S.; Schwemberger, S.; Afton, S.; Ben-Jonathan, N. Bisphenol A and estradiol are equipotent in antagonizing cisplatin-induced cytotoxicity in breast cancer cells. Cancer Lett. 2010, 290, 167–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R., Jr.; Lee, D.H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef]
- Goodson, W.H., 3rd; Lowe, L.; Carpenter, D.O.; Gilbertson, M.; Manaf Ali, A.; Lopez de Cerain Salsamendi, A.; Lasfar, A.; Carnero, A.; Azqueta, A.; Amedei, A.; et al. Assessing the carcinogenic potential of low-dose exposures to chemical mixtures in the environment: The challenge ahead. Carcinogenesis 2015, 36, S254–S296. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Santucci-Pereira, J.; Dang, N.M.; Kanefsky, J.; Rahulkannan, V.; Hillegass, M.; Joshi, S.; Gurdogan, H.; Chen, Z.; Bessonneau, V.; et al. Effects of Pubertal Exposure to Butyl Benzyl Phthalate, Perfluorooctanoic Acid, and Zeranol on Mammary Gland Development and Tumorigenesis in Rats. Int. J. Mol. Sci. 2022, 23, 1398. [Google Scholar] [CrossRef]
- Rignall, B.; Grote, K.; Gavrilov, A.; Weimer, M.; Kopp-Schneider, A.; Krause, E.; Appel, K.E.; Buchmann, A.; Robertson, L.W.; Lehmler, H.J.; et al. Biological and tumor-promoting effects of dioxin-like and non-dioxin-like polychlorinated biphenyls in mouse liver after single or combined treatment. Toxicol. Sci. 2013, 133, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Ali, E.S.; Sahu, U.; Villa, E.; O’Hara, B.P.; Gao, P.; Beaudet, C.; Wood, A.W.; Asara, J.M.; Ben-Sahra, I. ERK2 Phosphorylates PFAS to Mediate Posttranslational Control of De Novo Purine Synthesis. Mol. Cell 2020, 78, 1178–1191. [Google Scholar] [CrossRef]
- Mancini, F.R.; Cano-Sancho, G.; Mohamed, O.; Cervenka, I.; Omichessan, H.; Marchand, P.; Boutron-Ruault, M.C.; Arveux, P.; Severi, G.; Antignac, J.P.; et al. Plasma concentration of brominated flame retardants and postmenopausal breast cancer risk: A nested case-control study in the French E3N cohort. Environ. Health 2020, 19, 54. [Google Scholar] [CrossRef]
- Ramos, J.J.; Huetos, O.; Gonzalez, S.; Esteban, M.; Calvo, E.; Perez-Gomez, B.; Castano, A.; Bioambient es. Organochlorinated pesticides levels in a representative sample of the Spanish adult population: The Bioambient. es project. Int. J. Hyg. Environ. Health 2017, 220, 217–226. [Google Scholar] [CrossRef]
- Steffan, J.J.; Williams, B.C.; Welbourne, T.; Cardelli, J.A. HGF-induced invasion by prostate tumor cells requires anterograde lysosome trafficking and activity of Na+-H+ exchangers. J. Cell Sci. 2010, 123, 1151–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Migliaccio, A.; Di Domenico, M.; Castoria, G.; de Falco, A.; Bontempo, P.; Nola, E.; Auricchio, F. Tyrosine kinase/p21ras/MAP-kinase pathway activation by estradiol-receptor complex in MCF-7 cells. EMBO J. 1996, 15, 1292–1300. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.; Gurrapu, S.; Cagnoni, G.; Capparuccia, L.; Tamagnone, L. PlexinD1 Is a Novel Transcriptional Target and Effector of Notch Signaling in Cancer Cells. PLoS ONE 2016, 11, e0164660. [Google Scholar] [CrossRef] [Green Version]
- Steenland, K.; Fletcher, T.; Savitz, D.A. Epidemiologic evidence on the health effects of perfluorooctanoic acid (PFOA). Environ. Health Perspect. 2010, 118, 1100–1108. [Google Scholar] [CrossRef]
- Vieira, V.M.; Hoffman, K.; Shin, H.M.; Weinberg, J.M.; Webster, T.F.; Fletcher, T. Perfluorooctanoic acid exposure and cancer outcomes in a contaminated community: A geographic analysis. Environ. Health Perspect. 2013, 121, 318–323. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.-H.; Jacobs, D.R., Jr. Methodological issues in human studies of endocrine disrupting chemicals. Rev. Endocr. Metab. Disord. 2015, 16, 289–297. [Google Scholar] [CrossRef]
- Howsam, M.; Grimalt, J.O.; Guinó, E.; Navarro, M.; Martí-Ragué, J.; Peinado, M.A.; Capellá, G.; Moreno, V.; Bellvitge Colorectal Cancer, G. Organochlorine exposure and colorectal cancer risk. Environ. Health Perspect. 2004, 112, 1460–1466. [Google Scholar] [CrossRef] [Green Version]
- Casazza, A.; Finisguerra, V.; Capparuccia, L.; Camperi, A.; Swiercz, J.M.; Rizzolio, S.; Rolny, C.; Christensen, C.; Bertotti, A.; Sarotto, I.; et al. Sema3E-Plexin D1 signaling drives human cancer cell invasiveness and metastatic spreading in mice. J. Clin. Investig. 2010, 120, 2684–2698. [Google Scholar] [CrossRef] [Green Version]
- Mancini, F.R.; Cano-Sancho, G.; Gambaretti, J.; Marchand, P.; Boutron-Ruault, M.C.; Severi, G.; Arveux, P.; Antignac, J.P.; Kvaskoff, M. Perfluorinated alkylated substances serum concentration and breast cancer risk: Evidence from a nested case-control study in the French E3N cohort. Int. J. Cancer 2019, 22, 32357. [Google Scholar] [CrossRef]
- Miao, C.; Ma, J.; Zhang, Y.; Chu, Y.; Li, J.; Kuai, R.; Wang, S.; Peng, H. Perfluorooctanoic acid enhances colorectal cancer DLD-1 cells invasiveness through activating NF-kappaB mediated matrix metalloproteinase-2/-9 expression. Int. J. Clin. Exp. Pathol. 2015, 8, 10512–10522. [Google Scholar]
- Li, X.; Bao, C.; Ma, Z.; Xu, B.; Ying, X.; Liu, X.; Zhang, X. Perfluorooctanoic acid stimulates ovarian cancer cell migration, invasion via ERK/NF-kappaB/MMP-2/-9 pathway. Toxicol. Lett. 2018, 294, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, F.; Xu, P.; Miao, C.; Zeng, X.; Cui, X.; Lu, C.; Xie, H.; Yin, H.; Chen, F.; et al. Perfluorooctanoic acid stimulates breast cancer cells invasion and up-regulates matrix metalloproteinase-2/-9 expression mediated by activating NF-kappaB. Toxicol. Lett. 2014, 229, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Bunay, J.; Larriba, E.; Moreno, R.D.; Del Mazo, J. Chronic low-dose exposure to a mixture of environmental endocrine disruptors induces microRNAs/isomiRs deregulation in mouse concomitant with intratesticular estradiol reduction. Sci. Rep. 2017, 7, 3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnick, R.; Lucier, G.; Wolfe, M.; Hall, R.; Stancel, G.; Prins, G.; Gallo, M.; Reuhl, K.; Ho, S.M.; Brown, T.; et al. Summary of the National Toxicology Program’s report of the endocrine disruptors low-dose peer review. Environ. Health Perspect. 2002, 110, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.Y.; Lu, R.; Hu, D.P.; Imir, O.B.; Zuo, Q.; Moline, D.; Afradiasbagharani, P.; Liu, L.; Lowe, S.; Birch, L.; et al. Per- and polyfluoroalkyl substances target and alter human prostate stem-progenitor cells. Biochem. Pharm. 2022, 197, 114902. [Google Scholar] [CrossRef]
- Kim, J.; Oh, W.J.; Gaiano, N.; Yoshida, Y.; Gu, C. Semaphorin 3E-Plexin-D1 signaling regulates VEGF function in developmental angiogenesis via a feedback mechanism. Genes Dev. 2011, 25, 1399–1411. [Google Scholar] [CrossRef] [Green Version]
- Blanc, V.; Nariculam, J.; Munson, P.; Freeman, A.; Klocker, H.; Masters, J.; Williamson, M. A role for class 3 semaphorins in prostate cancer. Prostate 2011, 71, 649–658. [Google Scholar] [CrossRef]
- Li, K.; Gao, P.; Xiang, P.; Zhang, X.; Cui, X.; Ma, L.Q. Molecular mechanisms of PFOA-induced toxicity in animals and humans: Implications for health risks. Environ. Int. 2017, 99, 43–54. [Google Scholar] [CrossRef]
- Yan, S.; Zhang, H.; Zheng, F.; Sheng, N.; Guo, X.; Dai, J. Perfluorooctanoic acid exposure for 28 days affects glucose homeostasis and induces insulin hypersensitivity in mice. Sci. Rep. 2015, 5, 11029. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Liu, X.; Li, F.; Wang, Y.; Xu, Y.; Zhang, M.; Zhang, X.; Ying, X.; Zhang, X. Perfluorooctanoic acid induces human Ishikawa endometrial cancer cell migration and invasion through activation of ERK/mTOR signaling. Oncotarget 2016, 7, 66558–66568. [Google Scholar] [CrossRef] [Green Version]
- Costa, R.L.B.; Han, H.S.; Gradishar, W.J. Targeting the PI3K/AKT/mTOR pathway in triple-negative breast cancer: A review. Breast Cancer Res. Treat. 2018, 169, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Steenland, K.; Winquist, A. PFAS and cancer, a scoping review of the epidemiologic evidence. Environ. Res. 2021, 194, 110690. [Google Scholar] [CrossRef] [PubMed]
- Torrino, S.; Tiroille, V.; Dolfi, B.; Dufies, M.; Hinault, C.; Bonesso, L.; Dagnino, S.; Uhler, J.; Irondelle, M.; Gay, A.S.; et al. UBTD1 regulates ceramide balance and endolysosomal positioning to coordinate EGFR signaling. eLife 2021, 10, e68348. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, J.Y.; Sun, L.S.; Li, X.F.; Bai, J.Y.; Zhang, H.Y.; Li, T.J. miR-762 Promotes Malignant Development of Head and Neck Squamous Cell Carcinoma by Targeting PHLPP2 and FOXO4. OncoTargets 2019, 12, 11425–11436. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Charazac, A.; Hinault, C.; Dolfi, B.; Hautier, S.; Decondé Le Butor, C.; Bost, F.; Chevalier, N. Low Doses of PFOA Promote Prostate and Breast Cancer Cells Growth through Different Pathways. Int. J. Mol. Sci. 2022, 23, 7900. https://doi.org/10.3390/ijms23147900
Charazac A, Hinault C, Dolfi B, Hautier S, Decondé Le Butor C, Bost F, Chevalier N. Low Doses of PFOA Promote Prostate and Breast Cancer Cells Growth through Different Pathways. International Journal of Molecular Sciences. 2022; 23(14):7900. https://doi.org/10.3390/ijms23147900
Chicago/Turabian StyleCharazac, Aurélie, Charlotte Hinault, Bastien Dolfi, Solène Hautier, Célia Decondé Le Butor, Frédéric Bost, and Nicolas Chevalier. 2022. "Low Doses of PFOA Promote Prostate and Breast Cancer Cells Growth through Different Pathways" International Journal of Molecular Sciences 23, no. 14: 7900. https://doi.org/10.3390/ijms23147900
APA StyleCharazac, A., Hinault, C., Dolfi, B., Hautier, S., Decondé Le Butor, C., Bost, F., & Chevalier, N. (2022). Low Doses of PFOA Promote Prostate and Breast Cancer Cells Growth through Different Pathways. International Journal of Molecular Sciences, 23(14), 7900. https://doi.org/10.3390/ijms23147900