
Remodeling of the Lamina Cribrosa: Mechanisms and Potential Therapeutic Approaches for Glaucoma


Abstract
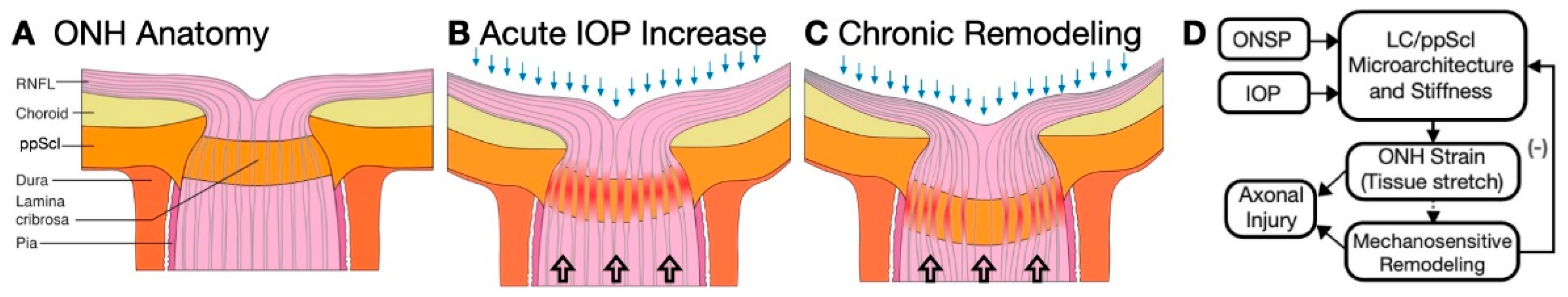
:1. Introduction
2. Remodeling Response in Aging and Glaucoma
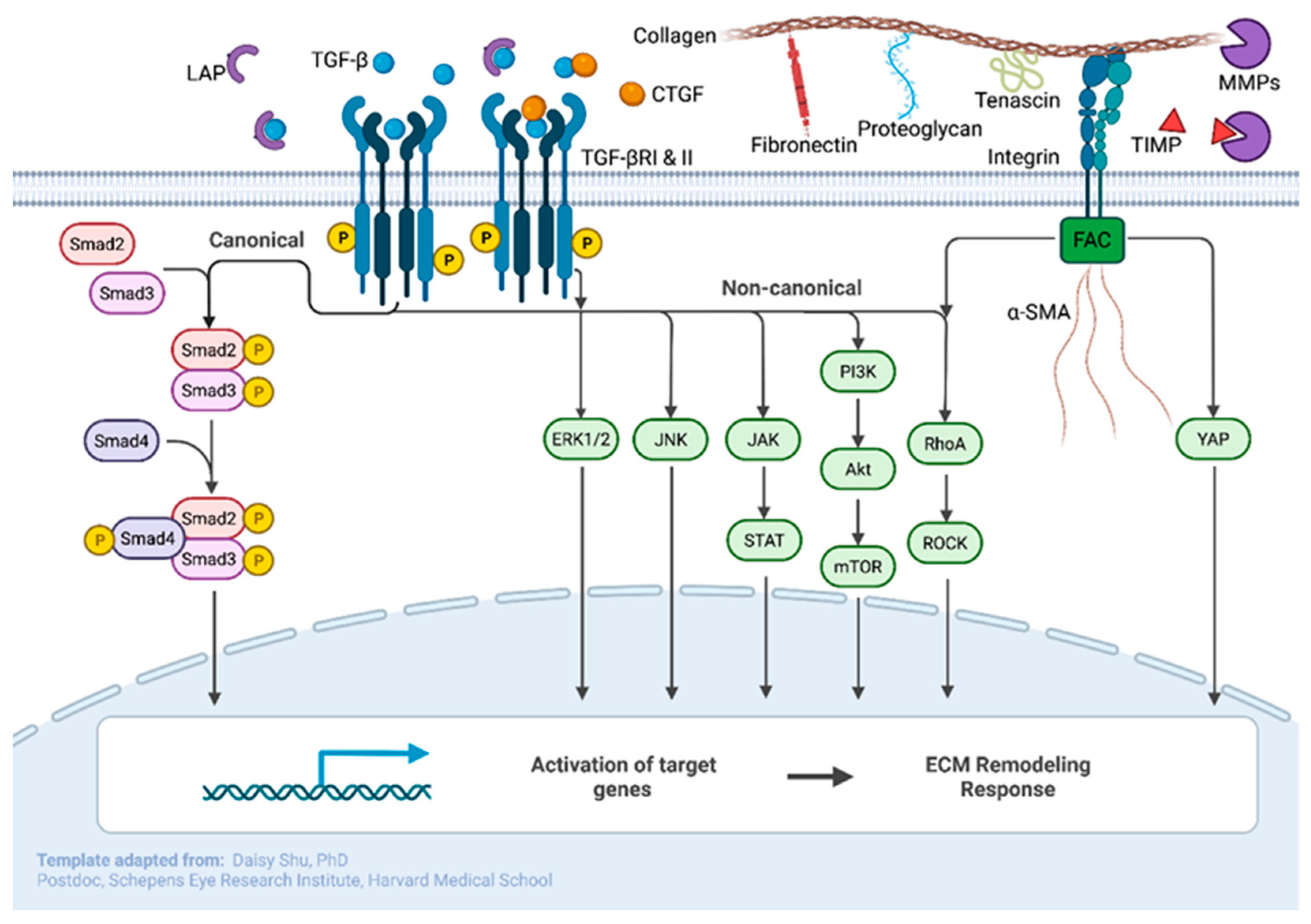
3. Lamina Cribrosa Cells
4. Glial Cells
5. Scleral Fibroblasts
6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tham, Y.-C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.-Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Quigley, H.A.; Broman, A.T. The number of people with glaucoma worldwide in 2010 and 2020. Br. J. Ophthalmol. 2006, 90, 262–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Downs, J.C.; Girkin, C.A. Lamina cribrosa in glaucoma. Curr. Opin. Ophthalmol. 2017, 28, 113–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigal, I.A. Interactions between geometry and mechanical properties on the optic nerve head. Investig. Ophthalmol. Vis. Sci. 2009, 50, 2785–2795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grytz, R.; Fazio, M.A.; Libertiaux, V.; Bruno, L.; Gardiner, S.; Girkin, C.A.; Downs, J.C. Age- and race-related differences in human scleral material properties. Investig. Ophthalmol. Vis. Sci. 2014, 55, 8163–8172. [Google Scholar] [CrossRef] [Green Version]
- Coudrillier, B.; Tian, J.; Alexander, S.; Myers, K.M.; Quigley, H.A.; Nguyen, T.D. Biomechanics of the human posterior sclera: Age- and glaucoma-related changes measured using inflation testing. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1714–1728. [Google Scholar] [CrossRef]
- Safa, B.N.; Wong, C.A.; Ha, J.; Ethier, C.R. Glaucoma and biomechanics. Curr. Opin. Ophthalmol. 2022, 33, 80–90. [Google Scholar] [CrossRef]
- Downs, J.C. Optic nerve head biomechanics in aging and disease. Exp. Eye Res. 2015, 133, 19–29. [Google Scholar] [CrossRef] [Green Version]
- AGIS Investigators. The Advanced Glaucoma Intervention Study (AGIS): 12. Baseline risk factors for sustained loss of visual field and visual acuity in patients with advanced glaucoma. Am. J. Ophthalmol. 2002, 134, 499–512. [Google Scholar] [CrossRef]
- Leske, M.C.; Heijl, A.; Hussein, M.; Bengtsson, B.; Hyman, L.; Komaroff, E.; Early Manifest Glaucoma Trial Group. Factors for glaucoma progression and the effect of treatment: The Early Manifest Glaucoma Trial. Arch. Ophthalmol. 2003, 121, 48–56. [Google Scholar] [CrossRef]
- Musch, D.C.; Gillespie, B.W.; Lichter, P.R.; Niziol, L.M.; Janz, N.K.; CIGTS Study Investigators. Visual field progression in the Collaborative Initial Glaucoma Treatment Study the impact of treatment and other baseline factors. Ophthalmology 2009, 116, 200–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drance, S.; Anderson, D.R.; Schulzer, M.; Collaborative Normal-Tension Glaucoma Study Group. Risk factors for progression of visual field abnormalities in normal-tension glaucoma. Am. J. Ophthalmol. 2001, 131, 699–708. [Google Scholar] [CrossRef]
- Gordon, M.O.; Beiser, J.A.; Brandt, J.D.; Heuer, D.K.; Higginbotham, E.J.; Johnson, C.A.; Keltner, J.L.; Miller, J.P.; Parrish, R.K.; Wilson, M.R.; et al. The Ocular Hypertension Treatment Study: Baseline factors that predict the onset of primary open-angle glaucoma. Arch. Ophthalmol. 2002, 120, 714–720, Discussion 829. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, M.R.; Andrzejewska, W.M.; Neufeld, A.H. Changes in the extracellular matrix of the human optic nerve head in primary open-angle glaucoma. Am. J. Ophthalmol. 1990, 109, 180–188. [Google Scholar] [CrossRef]
- Hernandez, M.R. Ultrastructural immunocytochemical analysis of elastin in the human lamina cribrosa. Changes in elastic fibers in primary open-angle glaucoma. Investig. Ophthalmol. Vis. Sci. 1992, 33, 2891–2903. [Google Scholar]
- Sawaguchi, S.; Yue, B.Y.; Fukuchi, T.; Abe, H.; Suda, K.; Kaiya, T.; Iwata, K. Collagen fibrillar network in the optic nerve head of normal monkey eyes and monkey eyes with laser-induced glaucoma—A scanning electron microscopic study. Curr. Eye Res. 1999, 18, 143–149. [Google Scholar] [CrossRef]
- Quigley, H.A. The contribution of the sclera and lamina cribrosa to the pathogenesis of glaucoma: Diagnostic and treatment implications. Prog. Brain Res. 2015, 220, 59–86. [Google Scholar] [CrossRef]
- Fazio, M.A.; Grytz, R.; Morris, J.S.; Bruno, L.; Gardiner, S.K.; Girkin, C.A.; Downs, J.C. Age-related changes in human peripapillary scleral strain. Biomech. Model. Mechanobiol. 2014, 13, 551–563. [Google Scholar] [CrossRef] [Green Version]
- Quigley, H.A.; Dorman-Pease, M.E.; Brown, A.E. Quantitative study of collagen and elastin of the optic nerve head and sclera in human and experimental monkey glaucoma. Curr. Eye Res. 1991, 10, 877–888. [Google Scholar] [CrossRef]
- Fukuchi, T.; Sawaguchi, S.; Hara, H.; Shirakashi, M.; Iwata, K. Extracellular matrix changes of the optic nerve lamina cribrosa in monkey eyes with experimentally chronic glaucoma. Graefe’s Arch. Clin. Exp. Ophthalmol. 1992, 230, 421–427. [Google Scholar] [CrossRef]
- Prendes, M.A.; Harris, A.; Wirostko, B.M.; Gerber, A.L.; Siesky, B. The role of transforming growth factor β in glaucoma and the therapeutic implications. Br. J. Ophthalmol. 2013, 97, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Zode, G.S.; Sethi, A.; Brun-Zinkernagel, A.-M.; Chang, I.-F.; Clark, A.F.; Wordinger, R.J. Transforming growth factor-β2 increases extracellular matrix proteins in optic nerve head cells via activation of the Smad signaling pathway. Mol. Vis. 2011, 17, 1745–1758. [Google Scholar] [PubMed]
- Kirwan, R.P.; Leonard, M.O.; Murphy, M.; Clark, A.F.; O’Brien, C.J. Transforming growth factor-beta-regulated gene transcription and protein expression in human GFAP-negative lamina cribrosa cells. Glia 2005, 52, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Kirwan, R.P.; Crean, J.K.; Fenerty, C.H.; Clark, A.F.; O’Brien, C.J. Effect of cyclical mechanical stretch and exogenous transforming growth factor-beta1 on matrix metalloproteinase-2 activity in lamina cribrosa cells from the human optic nerve head. J. Glaucoma 2004, 13, 327–334. [Google Scholar] [CrossRef]
- Annes, J.P.; Munger, J.S.; Rifkin, D.B. Making sense of latent TGFbeta activation. J. Cell Sci. 2003, 116, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Murphy-Ullrich, J.E.; Downs, J.C. The Thrombospondin1-TGF-β Pathway and Glaucoma. J. Ocul. Pharmacol. Ther. 2015, 31, 371–375. [Google Scholar] [CrossRef]
- Murphy, G.; Nagase, H. Progress in matrix metalloproteinase research. Mol. Aspects Med. 2008, 29, 290–308. [Google Scholar] [CrossRef] [Green Version]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef]
- Yuan, L.; Neufeld, A.H. Activated microglia in the human glaucomatous optic nerve head. J. Neurosci. Res. 2001, 64, 523–532. [Google Scholar] [CrossRef]
- Murphy-Ullrich, J.E.; Suto, M.J. Thrombospondin-1 regulation of latent TGF-β activation: A therapeutic target for fibrotic disease. Matrix Biol. 2018, 68–69, 28–43. [Google Scholar] [CrossRef]
- Haddadin, R.I.; Oh, D.-J.; Kang, M.H.; Villarreal, G.; Kang, J.-H.; Jin, R.; Gong, H.; Rhee, D.J. Thrombospondin-1 (TSP1)-null and TSP2-null mice exhibit lower intraocular pressures. Investig. Ophthalmol. Vis. Sci. 2012, 53, 6708–6717. [Google Scholar] [CrossRef] [PubMed]
- Iomdina, E.N.; Tikhomirova, N.K.; Bessmertny, A.M.; Serebryakova, M.V.; Baksheeva, V.E.; Zalevsky, A.O.; Kotelin, V.I.; Kiseleva, O.A.; Kosakyan, S.M.; Zamyatnin, A.A.; et al. Alterations in proteome of human sclera associated with primary open-angle glaucoma involve proteins participating in regulation of the extracellular matrix. Mol. Vis. 2020, 26, 623–640. [Google Scholar] [PubMed]
- Morrison, J.C. Integrins in the optic nerve head: Potential roles in glaucomatous optic neuropathy (an American Ophthalmological Society thesis). Trans. Am. Ophthalmol. Soc. 2006, 104, 453–477. [Google Scholar]
- Wallace, D.M.; O’Brien, C.J. The role of lamina cribrosa cells in optic nerve head fibrosis in glaucoma. Exp. Eye Res. 2016, 142, 102–109. [Google Scholar] [CrossRef]
- Hernandez, M.R.; Igoe, F.; Neufeld, A.H. Cell culture of the human lamina cribrosa. Investig. Ophthalmol. Vis. Sci. 1988, 29, 78–89. [Google Scholar]
- Kirwan, R.P.; Fenerty, C.H.; Crean, J.; Wordinger, R.J.; Clark, A.F.; O’Brien, C.J. Influence of cyclical mechanical strain on extracellular matrix gene expression in human lamina cribrosa cells in vitro. Mol. Vis. 2005, 11, 798–810. [Google Scholar] [PubMed]
- Wallace, D.M.; Clark, A.F.; Lipson, K.E.; Andrews, D.; Crean, J.K.; O’Brien, C.J. Anti-connective tissue growth factor antibody treatment reduces extracellular matrix production in trabecular meshwork and lamina cribrosa cells. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7836–7848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirwan, R.P.; Felice, L.; Clark, A.F.; O’Brien, C.J.; Leonard, M.O. Hypoxia regulated gene transcription in human optic nerve lamina cribrosa cells in culture. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2243–2255. [Google Scholar] [CrossRef] [Green Version]
- Tezel, G.; Wax, M.B. Hypoxia-inducible factor 1alpha in the glaucomatous retina and optic nerve head. Arch. Ophthalmol. 2004, 122, 1348–1356. [Google Scholar] [CrossRef] [Green Version]
- McElnea, E.M.; Quill, B.; Docherty, N.G.; Irnaten, M.; Siah, W.F.; Clark, A.F.; O’Brien, C.J.; Wallace, D.M. Oxidative stress, mitochondrial dysfunction and calcium overload in human lamina cribrosa cells from glaucoma donors. Mol. Vis. 2011, 17, 1182–1191. [Google Scholar]
- Yan, X.; Tezel, G.; Wax, M.B.; Edward, D.P. Matrix metalloproteinases and tumor necrosis factor alpha in glaucomatous optic nerve head. Arch. Ophthalmol. 2000, 118, 666–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Williams, G.; Downs, J.C.; Sigal, I.A.; Roberts, M.D.; Thompson, H.; Burgoyne, C.F. Posterior (outward) migration of the lamina cribrosa and early cupping in monkey experimental glaucoma. Investig. Ophthalmol. Vis. Sci. 2011, 52, 7109–7121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Thompson, H.; Roberts, M.D.; Sigal, I.A.; Downs, J.C.; Burgoyne, C.F. Deformation of the early glaucomatous monkey optic nerve head connective tissue after acute IOP elevation in 3-D histomorphometric reconstructions. Investig. Ophthalmol. Vis. Sci. 2011, 52, 345–363. [Google Scholar] [CrossRef] [PubMed]
- Liou, J.-J.; Geest, J.P.V. Effect of transforming growth factor beta 2 on matrix metalloproteinase activity in porcine lamina cribrosa cells. Investig. Ophthalmol. Vis. Sci. 2020, 61, 902. [Google Scholar]
- Burgoyne, C.F. The non-human primate experimental glaucoma model. Exp. Eye Res. 2015, 141, 57–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irnaten, M.; Barry, R.C.; Wallace, D.M.; Docherty, N.G.; Quill, B.; Clark, A.F.; O’Brien, C.J. Elevated maxi-K(+) ion channel current in glaucomatous lamina cribrosa cells. Exp. Eye Res. 2013, 115, 224–229. [Google Scholar] [CrossRef]
- Irnaten, M.; Zhdanov, A.; Brennan, D.; Crotty, T.; Clark, A.; Papkovsky, D.; O’Brien, C. Activation of the NFAT-Calcium Signaling Pathway in Human Lamina Cribrosa Cells in Glaucoma. Investig. Ophthalmol. Vis. Sci. 2018, 59, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Tidu, F.; De Zuani, M.; Jose, S.S.; Bendíčková, K.; Kubala, L.; Caruso, F.; Cavalieri, F.; Forte, G.; Frič, J. NFAT signaling in human mesenchymal stromal cells affects extracellular matrix remodeling and antifungal immune responses. iScience 2021, 24, 102683. [Google Scholar] [CrossRef]
- Asghar, M.Y.; Törnquist, K. Transient receptor potential canonical (TRPC) channels as modulators of migration and invasion. Int. J. Mol. Sci. 2020, 21, 1739. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Hopkins, C.R. Review of transient receptor potential canonical (TRPC5) channel modulators and diseases. J. Med. Chem. 2019, 62, 7589–7602. [Google Scholar] [CrossRef]
- Irnaten, M.; O’Malley, G.; Clark, A.F.; O’Brien, C.J. Transient receptor potential channels TRPC1/TRPC6 regulate lamina cribrosa cell extracellular matrix gene transcription and proliferation. Exp. Eye Res. 2020, 193, 107980. [Google Scholar] [CrossRef] [PubMed]
- Chigurupati, S.; Venkataraman, R.; Barrera, D.; Naganathan, A.; Madan, M.; Paul, L.; Pattisapu, J.V.; Kyriazis, G.A.; Sugaya, K.; Bushnev, S.; et al. Receptor channel TRPC6 is a key mediator of Notch-driven glioblastoma growth and invasiveness. Cancer Res. 2010, 70, 418–427. [Google Scholar] [CrossRef] [Green Version]
- Bomben, V.C.; Turner, K.L.; Barclay, T.-T.C.; Sontheimer, H. Transient receptor potential canonical channels are essential for chemotactic migration of human malignant gliomas. J. Cell. Physiol. 2011, 226, 1879–1888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, M.R. The optic nerve head in glaucoma: Role of astrocytes in tissue remodeling. Prog. Retin. Eye Res. 2000, 19, 297–321. [Google Scholar] [CrossRef]
- Bowman, C.L.; Ding, J.P.; Sachs, F.; Sokabe, M. Mechanotransducing ion channels in astrocytes. Brain Res. 1992, 584, 272–286. [Google Scholar] [CrossRef]
- Wang, R.; Seifert, P.; Jakobs, T.C. Astrocytes in the optic nerve head of glaucomatous mice display a characteristic reactive phenotype. Investig. Ophthalmol. Vis. Sci. 2017, 58, 924–932. [Google Scholar] [CrossRef]
- Pena, J.D.; Agapova, O.; Gabelt, B.T.; Levin, L.A.; Lucarelli, M.J.; Kaufman, P.L.; Hernandez, M.R. Increased elastin expression in astrocytes of the lamina cribrosa in response to elevated intraocular pressure. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2303–2314. [Google Scholar]
- Rogers, R.S.; Dharsee, M.; Ackloo, S.; Sivak, J.M.; Flanagan, J.G. Proteomics analyses of human optic nerve head astrocytes following biomechanical strain. Mol. Cell. Proteom. 2012, 11, M111.012302. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, M.R.; Wang, N.; Hanley, N.M.; Neufeld, A.H. Localization of collagen types I and IV mRNAs in human optic nerve head by in situ hybridization. Investig. Ophthalmol. Vis. Sci. 1991, 32, 2169–2177. [Google Scholar]
- Ye, H.; Yang, J.; Hernandez, M.R. Localization of collagen type III mRNA in normal human optic nerve heads. Exp. Eye Res. 1994, 58, 53–63. [Google Scholar] [CrossRef]
- Pena, J.D.; Roy, S.; Hernandez, M.R. Tropoelastin gene expression in optic nerve heads of normal and glaucomatous subjects. Matrix Biol. 1996, 15, 323–330. [Google Scholar] [CrossRef]
- Tehrani, S.; Davis, L.; Cepurna, W.O.; Choe, T.E.; Lozano, D.C.; Monfared, A.; Cooper, L.; Cheng, J.; Johnson, E.C.; Morrison, J.C. Astrocyte Structural and Molecular Response to Elevated Intraocular Pressure Occurs Rapidly and Precedes Axonal Tubulin Rearrangement within the Optic Nerve Head in a Rat Model. PLoS ONE 2016, 11, e0167364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Qu, J.; Jakobs, T.C. Reversible reactivity by optic nerve astrocytes. Glia 2013, 61, 1218–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, C.; Yu, A.; Welge-Lüssen, U.; Lütjen-Drecoll, E.; Birke, M. The effect of TGF-beta2 on elastin, type VI collagen, and components of the proteolytic degradation system in human optic nerve astrocytes. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1464–1472. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-L.; Sung, K.R.; Kwon, J.; Shin, J.A. Statins Suppress TGF-β2-Mediated MMP-2 and MMP-9 Expression and Activation Through RhoA/ROCK Inhibition in Astrocytes of the Human Optic Nerve Head. Investig. Ophthalmol. Vis. Sci. 2020, 61, 29. [Google Scholar] [CrossRef]
- Nakerakanti, S.S.; Bujor, A.M.; Trojanowska, M. CCN2 is required for the TGF-β induced activation of Smad1-Erk1/2 signaling network. PLoS ONE 2011, 6, e21911. [Google Scholar] [CrossRef]
- Junglas, B.; Kuespert, S.; Seleem, A.A.; Struller, T.; Ullmann, S.; Bösl, M.; Bosserhoff, A.; Köstler, J.; Wagner, R.; Tamm, E.R.; et al. Connective tissue growth factor causes glaucoma by modifying the actin cytoskeleton of the trabecular meshwork. Am. J. Pathol. 2012, 180, 2386–2403. [Google Scholar] [CrossRef]
- Dillinger, A.E.; Weber, G.R.; Mayer, M.; Schneider, M.; Göppner, C.; Ohlmann, A.; Shamonin, M.; Monkman, G.J.; Fuchshofer, R. CCN2/CTGF-A Modulator of the Optic Nerve Head Astrocyte. Front. Cell Dev. Biol. 2022, 10, 864433. [Google Scholar] [CrossRef]
- Dillinger, A.E.; Kuespert, S.; Froemel, F.; Tamm, E.R.; Fuchshofer, R. CCN2/CTGF promotor activity in the developing and adult mouse eye. Cell Tissue Res. 2021, 384, 625–641. [Google Scholar] [CrossRef]
- Miao, H.; Crabb, A.W.; Hernandez, M.R.; Lukas, T.J. Modulation of factors affecting optic nerve head astrocyte migration. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4096–4103. [Google Scholar] [CrossRef]
- Tezel, G.; Hernandez, M.R.; Wax, M.B. In vitro evaluation of reactive astrocyte migration, a component of tissue remodeling in glaucomatous optic nerve head. Glia 2001, 34, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.J.; Sun, D.; Jakobs, T.C. Astrocytes in the optic nerve head express putative mechanosensitive channels. Mol. Vis. 2015, 21, 749–766. [Google Scholar] [PubMed]
- Lee, P.; Chandel, N.S.; Simon, M.C. Cellular adaptation to hypoxia through hypoxia inducible factors and beyond. Nat. Rev. Mol. Cell Biol. 2020, 21, 268–283. [Google Scholar] [CrossRef] [PubMed]
- Savagian, C.A.; Dubielzig, R.R.; Nork, T.M. Comparison of the distribution of glial fibrillary acidic protein, heat shock protein 60, and hypoxia-inducible factor-1alpha in retinas from glaucomatous and normal canine eyes. Am. J. Vet. Res. 2008, 69, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Chidlow, G.; Wood, J.P.M.; Casson, R.J. Investigations into Hypoxia and Oxidative Stress at the Optic Nerve Head in a Rat Model of Glaucoma. Front. Neurosci. 2017, 11, 478. [Google Scholar] [CrossRef] [PubMed]
- Ergorul, C.; Ray, A.; Huang, W.; Wang, D.Y.; Ben, Y.; Cantuti-Castelvetri, I.; Grosskreutz, C.L. Hypoxia inducible factor-1α (HIF-1α) and some HIF-1 target genes are elevated in experimental glaucoma. J. Mol. Neurosci. 2010, 42, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Egger, M.; Schgoer, W.; Beer, A.G.E.; Jeschke, J.; Leierer, J.; Theurl, M.; Frauscher, S.; Tepper, O.M.; Niederwanger, A.; Ritsch, A.; et al. Hypoxia up-regulates the angiogenic cytokine secretoneurin via an HIF-1alpha- and basic FGF-dependent pathway in muscle cells. FASEB J. 2007, 21, 2906–2917. [Google Scholar] [CrossRef] [Green Version]
- Fuller, J.A.; Brun-Zinkernagel, A.-M.; Clark, A.F.; Wordinger, R.J. Subtilisin-like proprotein convertase expression, localization, and activity in the human retina and optic nerve head. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5759–5768. [Google Scholar] [CrossRef] [Green Version]
- Nour, N.; Mayer, G.; Mort, J.S.; Salvas, A.; Mbikay, M.; Morrison, C.J.; Overall, C.M.; Seidah, N.G. The cysteine-rich domain of the secreted proprotein convertases PC5A and PACE4 functions as a cell surface anchor and interacts with tissue inhibitors of metalloproteinases. Mol. Biol. Cell 2005, 16, 5215–5226. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, J.V.; Soto, I.; Kim, K.-Y.; Bushong, E.A.; Oglesby, E.; Valiente-Soriano, F.J.; Yang, Z.; Davis, C.O.; Bedont, J.L.; Son, J.L.; et al. Myelination transition zone astrocytes are constitutively phagocytic and have synuclein dependent reactivity in glaucoma. Proc. Natl. Acad. Sci. USA 2011, 108, 1176–1181. [Google Scholar] [CrossRef] [Green Version]
- Cooper, M.L.; Crish, S.D.; Inman, D.M.; Horner, P.J.; Calkins, D.J. Early astrocyte redistribution in the optic nerve precedes axonopathy in the DBA/2J mouse model of glaucoma. Exp. Eye Res. 2016, 150, 22–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, C.O.; Kim, K.-Y.; Bushong, E.A.; Mills, E.A.; Boassa, D.; Shih, T.; Kinebuchi, M.; Phan, S.; Zhou, Y.; Bihlmeyer, N.A.; et al. Transcellular degradation of axonal mitochondria. Proc. Natl. Acad. Sci. USA 2014, 111, 9633–9638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muench, N.A.; Patel, S.; Maes, M.E.; Donahue, R.J.; Ikeda, A.; Nickells, R.W. The influence of mitochondrial dynamics and function on retinal ganglion cell susceptibility in optic nerve disease. Cells 2021, 10, 1593. [Google Scholar] [CrossRef] [PubMed]
- Neufeld, A.H. Microglia in the optic nerve head and the region of parapapillary chorioretinal atrophy in glaucoma. Arch. Ophthalmol. 1999, 117, 1050–1056. [Google Scholar] [CrossRef] [Green Version]
- Crapser, J.D.; Arreola, M.A.; Tsourmas, K.I.; Green, K.N. Microglia as hackers of the matrix: Sculpting synapses and the extracellular space. Cell. Mol. Immunol. 2021, 18, 2472–2488. [Google Scholar] [CrossRef] [PubMed]
- Nayak, D.; Roth, T.L.; McGavern, D.B. Microglia development and function. Annu. Rev. Immunol. 2014, 32, 367–402. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, S.; Bähr, M. Immunocytochemical characterization of reactive optic nerve astrocytes and meningeal cells. Glia 1999, 26, 36–46. [Google Scholar] [CrossRef]
- Pena, J.D.; Varela, H.J.; Ricard, C.S.; Hernandez, M.R. Enhanced tenascin expression associated with reactive astrocytes in human optic nerve heads with primary open angle glaucoma. Exp. Eye Res. 1999, 68, 29–40. [Google Scholar] [CrossRef]
- Pena, J.D.; Taylor, A.W.; Ricard, C.S.; Vidal, I.; Hernandez, M.R. Transforming growth factor beta isoforms in human optic nerve heads. Br. J. Ophthalmol. 1999, 83, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Reinehr, S.; Reinhard, J.; Wiemann, S.; Stute, G.; Kuehn, S.; Woestmann, J.; Dick, H.B.; Faissner, A.; Joachim, S.C. Early remodelling of the extracellular matrix proteins tenascin-C and phosphacan in retina and optic nerve of an experimental autoimmune glaucoma model. J. Cell. Mol. Med. 2016, 20, 2122–2137. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.; Bao, X.; Zang, C.; Yang, H.; Sun, F.; Che, Y.; Wu, X.; Li, S.; Zhang, D.; Wang, Q. Integrin CD11b mediates α-synuclein-induced activation of NADPH oxidase through a Rho-dependent pathway. Redox Biol. 2018, 14, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, U.-K.; Kettenmann, H. Microglia: Active sensor and versatile effector cells in the normal and pathologic brain. Nat. Neurosci. 2007, 10, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- García-Bermúdez, M.Y.; Freude, K.K.; Mouhammad, Z.A.; van Wijngaarden, P.; Martin, K.K.; Kolko, M. Glial cells in glaucoma: Friends, foes, and potential therapeutic targets. Front. Neurol. 2021, 12, 624983. [Google Scholar] [CrossRef] [PubMed]
- Rolle, T.; Ponzetto, A.; Malinverni, L. The role of neuroinflammation in glaucoma: An update on molecular mechanisms and new therapeutic options. Front. Neurol. 2020, 11, 612422. [Google Scholar] [CrossRef]
- Tezel, G. Molecular regulation of neuroinflammation in glaucoma: Current knowledge and the ongoing search for new treatment targets. Prog. Retin. Eye Res. 2022, 87, 100998. [Google Scholar] [CrossRef]
- Mac Nair, C.E.; Nickells, R.W. Neuroinflammation in glaucoma and optic nerve damage. Prog. Mol. Biol. Transl. Sci. 2015, 134, 343–363. [Google Scholar] [CrossRef]
- Soto, I.; Howell, G.R. The complex role of neuroinflammation in glaucoma. Cold Spring Harb. Perspect. Med. 2014, 4, a017269. [Google Scholar] [CrossRef]
- Williams, P.A.; Marsh-Armstrong, N.; Howell, G.R.; Lasker/IRRF Initiative on Astrocytes and Glaucomatous Neurodegeneration Participants. Neuroinflammation in glaucoma: A new opportunity. Exp. Eye Res. 2017, 157, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Watson, P.G.; Young, R.D. Scleral structure, organisation and disease. A review. Exp. Eye Res. 2004, 78, 609–623. [Google Scholar] [CrossRef]
- Cone-Kimball, E.; Nguyen, C.; Oglesby, E.N.; Pease, M.E.; Steinhart, M.R.; Quigley, H.A. Scleral structural alterations associated with chronic experimental intraocular pressure elevation in mice. Mol. Vis. 2013, 19, 2023–2039. [Google Scholar]
- Hinz, B.; Phan, S.H.; Thannickal, V.J.; Prunotto, M.; Desmoulière, A.; Varga, J.; De Wever, O.; Mareel, M.; Gabbiani, G. Recent developments in myofibroblast biology: Paradigms for connective tissue remodeling. Am. J. Pathol. 2012, 180, 1340–1355. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B.; Phan, S.H.; Thannickal, V.J.; Galli, A.; Bochaton-Piallat, M.-L.; Gabbiani, G. The myofibroblast: One function, multiple origins. Am. J. Pathol. 2007, 170, 1807–1816. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B. Myofibroblasts. Exp. Eye Res. 2016, 142, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Oglesby, E.N.; Tezel, G.; Cone-Kimball, E.; Steinhart, M.R.; Jefferys, J.; Pease, M.E.; Quigley, H.A. Scleral fibroblast response to experimental glaucoma in mice. Mol. Vis. 2016, 22, 82–99. [Google Scholar]
- Shelton, L.; Rada, J.S. Effects of cyclic mechanical stretch on extracellular matrix synthesis by human scleral fibroblasts. Exp. Eye Res. 2007, 84, 314–322. [Google Scholar] [CrossRef] [Green Version]
- Fujikura, H.; Seko, Y.; Tokoro, T.; Mochizuki, M.; Shimokawa, H. Involvement of mechanical stretch in the gelatinolytic activity of the fibrous sclera of chicks, in vitro. JPN J. Ophthalmol. 2002, 46, 24–30. [Google Scholar] [CrossRef]
- Yamaoka, A.; Matsuo, T.; Shiraga, F.; Ohtsuki, H. TIMP-1 production by human scleral fibroblast decreases in response to cyclic mechanical stretching. Ophthalmic Res. 2001, 33, 98–101. [Google Scholar] [CrossRef]
- Chow, A.; McCrea, L.; Kimball, E.; Schaub, J.; Quigley, H.; Pitha, I. Dasatinib inhibits peripapillary scleral myofibroblast differentiation. Exp. Eye Res. 2020, 194, 107999. [Google Scholar] [CrossRef]
- Cui, W.; Bryant, M.R.; Sweet, P.M.; McDonnell, P.J. Changes in gene expression in response to mechanical strain in human scleral fibroblasts. Exp. Eye Res. 2004, 78, 275–284. [Google Scholar] [CrossRef]
- Coudrillier, B.; Pijanka, J.; Jefferys, J.; Sorensen, T.; Quigley, H.A.; Boote, C.; Nguyen, T.D. Collagen structure and mechanical properties of the human sclera: Analysis for the effects of age. J. Biomech. Eng. 2015, 137, 041006. [Google Scholar] [CrossRef] [Green Version]
- Voorhees, A.P.; Jan, N.-J.; Hua, Y.; Yang, B.; Sigal, I.A. Peripapillary sclera architecture revisited: A tangential fiber model and its biomechanical implications. Acta Biomater. 2018, 79, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Szeto, J.; Chow, A.; McCrea, L.; Mozzer, A.; Nguyen, T.D.; Quigley, H.A.; Pitha, I. Regional differences and physiologic behaviors in peripapillary scleral fibroblasts. Investig. Ophthalmol. Vis. Sci. 2021, 62, 27. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, W.; Zhao, F.; Zhou, Q.; Reinach, P.S.; Deng, L.; Ma, L.; Luo, S.; Srinivasalu, N.; Pan, M.; et al. Scleral hypoxia is a target for myopia control. Proc. Natl. Acad. Sci. USA 2018, 115, E7091–E7100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.; Jiang, J.; Ding, B.; Xue, K.; Sun, X.; Qian, S. Mechanical strain regulates myofibroblast differentiation of human scleral fibroblasts by YAP. Front. Physiol. 2021, 12, 712509. [Google Scholar] [CrossRef]
- Pitha, I.; Oglesby, E.; Chow, A.; Kimball, E.; Pease, M.E.; Schaub, J.; Quigley, H. Rho-Kinase Inhibition Reduces Myofibroblast Differentiation and Proliferation of Scleral Fibroblasts Induced by Transforming Growth Factor β and Experimental Glaucoma. Transl. Vis. Sci. Technol. 2018, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Qu, J.; Chen, H.; Zhu, L.; Ambalavanan, N.; Girkin, C.A.; Murphy-Ullrich, J.E.; Downs, J.C.; Zhou, Y. High-Magnitude and/or High-Frequency Mechanical Strain Promotes Peripapillary Scleral Myofibroblast Differentiation. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7821–7830. [Google Scholar] [CrossRef] [Green Version]
- Skonier, J.; Neubauer, M.; Madisen, L.; Bennett, K.; Plowman, G.D.; Purchio, A.F. cDNA cloning and sequence analysis of beta ig-h3, a novel gene induced in a human adenocarcinoma cell line after treatment with transforming growth factor-beta. DNA Cell Biol. 1992, 11, 511–522. [Google Scholar] [CrossRef]
- Shelton, L.; Troilo, D.; Lerner, M.R.; Gusev, Y.; Brackett, D.J.; Rada, J.S. Microarray analysis of choroid/RPE gene expression in marmoset eyes undergoing changes in ocular growth and refraction. Mol. Vis. 2008, 14, 1465–1479. [Google Scholar]
- Shelton, L.; Rada, J.A.S. Inhibition of human scleral fibroblast cell attachment to collagen type I by TGFBIp. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3542–3552. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Cui, D.; Yang, X.; Hu, J.; Wan, W.; Zeng, J. The crucial role of collagen-binding integrins in maintaining the mechanical properties of human scleral fibroblasts-seeded collagen matrix. Mol. Vis. 2011, 17, 1334–1342. [Google Scholar]
- Kimball, E.C.; Nguyen, C.; Steinhart, M.R.; Nguyen, T.D.; Pease, M.E.; Oglesby, E.N.; Oveson, B.C.; Quigley, H.A. Experimental scleral cross-linking increases glaucoma damage in a mouse model. Exp. Eye Res. 2014, 128, 129–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannon, B.G.; Schwaner, S.A.; Boazak, E.M.; Gerberich, B.G.; Winger, E.J.; Prausnitz, M.R.; Ethier, C.R. Sustained scleral stiffening in rats after a single genipin treatment. J. R. Soc. Interface 2019, 16, 20190427. [Google Scholar] [CrossRef] [PubMed]
- Coudrillier, B.; Campbell, I.C.; Read, A.T.; Geraldes, D.M.; Vo, N.T.; Feola, A.; Mulvihill, J.; Albon, J.; Abel, R.L.; Ethier, C.R. Effects of peripapillary scleral stiffening on the deformation of the lamina cribrosa. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2666–2677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamdaoui, M.E.; Levy, A.M.; Stuber, A.B.; Girkin, C.A.; Kraft, T.W.; Samuels, B.C.; Grytz, R. Scleral crosslinking using genipin can compromise retinal structure and function in tree shrews. Exp. Eye Res. 2022, 219, 109039. [Google Scholar] [CrossRef]
- Murienne, B.J.; Jefferys, J.L.; Quigley, H.A.; Nguyen, T.D. The effects of glycosaminoglycan degradation on the mechanical behavior of the posterior porcine sclera. Acta Biomater. 2015, 12, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Spoerl, E.; Boehm, A.G.; Pillunat, L.E. The influence of various substances on the biomechanical behavior of lamina cribrosa and peripapillary sclera. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1286–1290. [Google Scholar] [CrossRef] [Green Version]
- Hatami-Marbini, H.; Pachenari, M. Tensile Viscoelastic Properties of the Sclera after Glycosaminoglycan Depletion. Curr. Eye Res. 2021, 46, 1299–1308. [Google Scholar] [CrossRef]
- Tribble, J.R.; Williams, P.A.; Caterson, B.; Sengpiel, F.; Morgan, J.E. Digestion of the glycosaminoglycan extracellular matrix by chondroitinase ABC supports retinal ganglion cell dendritic preservation in a rodent model of experimental glaucoma. Mol. Brain 2018, 11, 69. [Google Scholar] [CrossRef]
- Ehanire, T.; Ren, L.; Bond, J.; Medina, M.; Li, G.; Bashirov, L.; Chen, L.; Kokosis, G.; Ibrahim, M.; Selim, A.; et al. Angiotensin II stimulates canonical TGF-β signaling pathway through angiotensin type 1 receptor to induce granulation tissue contraction. J. Mol. Med. 2015, 93, 289–302. [Google Scholar] [CrossRef] [Green Version]
- Quigley, H.A.; Pitha, I.F.; Welsbie, D.S.; Nguyen, C.; Steinhart, M.R.; Nguyen, T.D.; Pease, M.E.; Oglesby, E.N.; Berlinicke, C.A.; Mitchell, K.L.; et al. Losartan treatment protects retinal ganglion cells and alters scleral remodeling in experimental glaucoma. PLoS ONE 2015, 10, e0141137. [Google Scholar] [CrossRef] [Green Version]
- Ota, T.; Murata, H.; Sugimoto, E.; Aihara, M.; Araie, M. Prostaglandin analogues and mouse intraocular pressure: Effects of tafluprost, latanoprost, travoprost, and unoprostone, considering 24-hour variation. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2006–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.Y.L.; Kim, J.H.; Lee, D.E.; Lee, J.H.; Park, C.K. Changes of the Retina and Intrinsic Survival Signals in a Rat Model of Glaucoma following Brinzolamide and Travoprost Treatments. Ophthalmic Res. 2011, 46, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Kurashima, H.; Watabe, H.; Sato, N.; Abe, S.; Ishida, N.; Yoshitomi, T. Effects of prostaglandin F(2α) analogues on endothelin-1-induced impairment of rabbit ocular blood flow: Comparison among tafluprost, travoprost, and latanoprost. Exp. Eye Res. 2010, 91, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Gagliuso, D.J.; Wang, R.-F.; Mittag, T.W.; Podos, S.M. Additivity of bimatoprost or travoprost to latanoprost in glaucomatous monkey eyes. Arch. Ophthalmol. 2004, 122, 1342–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netland, P.A.; Landry, T.; Sullivan, E.K.; Andrew, R.; Silver, L.; Weiner, A.; Mallick, S.; Dickerson, J.; Bergamini, M.V.; Robertson, S.M.; et al. Travoprost compared with latanoprost and timolol in patients with open-angle glaucoma or ocular hypertension. Am. J. Ophthalmol. 2001, 132, 472–484. [Google Scholar] [CrossRef]
- Hellberg, M.R.; Sallee, V.L.; McLaughlin, M.A.; Sharif, N.A.; Desantis, L.; Dean, T.R.; Zinke, P.W. Preclinical efficacy of travoprost, a potent and selective FP prostaglandin receptor agonist. J. Ocul. Pharmacol. Ther. 2001, 17, 421–432. [Google Scholar] [CrossRef]
- Carvalho, A.B.; Laus, J.L.; Costa, V.P.; Barros, P.S.M.; Silveira, P.R. Effects of travoprost 0.004% compared with latanoprost 0.005% on the intraocular pressure of normal dogs. Vet. Ophthalmol. 2006, 9, 121–125. [Google Scholar] [CrossRef]
- Schnichels, S.; Hurst, J.; de Vries, J.W.; Ullah, S.; Gruszka, A.; Kwak, M.; Löscher, M.; Dammeier, S.; Bartz-Schmidt, K.-U.; Spitzer, M.S.; et al. Self-assembled DNA nanoparticles loaded with travoprost for glaucoma-treatment. Nanomedicine 2020, 29, 102260. [Google Scholar] [CrossRef]
- Hernández, M.; Urcola, J.H.; Vecino, E. Retinal ganglion cell neuroprotection in a rat model of glaucoma following brimonidine, latanoprost or combined treatments. Exp. Eye Res. 2008, 86, 798–806. [Google Scholar] [CrossRef]
- McDonald, J.E.; Kiland, J.A.; Kaufman, P.L.; Bentley, E.; Ellinwood, N.M.; McLellan, G.J. Effect of topical latanoprost 0.005% on intraocular pressure and pupil diameter in normal and glaucomatous cats. Vet. Ophthalmol. 2016, 19, 13–23. [Google Scholar] [CrossRef]
- El-Nimri, N.W.; Wildsoet, C.F. Effects of topical latanoprost on intraocular pressure and myopia progression in young guinea pigs. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2644–2651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, Y.; Hu, Z.; Meng, Q.; Fang, P.; Liu, H. Bimatoprost increases mechanosensitivity of trigeminal ganglion neurons innervating the inner walls of rat anterior chambers via activation of TRPA1. Investig. Ophthalmol. Vis. Sci. 2016, 57, 567–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowston, J.G.; Lindsey, J.D.; Morris, C.A.; Wheeler, L.; Medeiros, F.A.; Weinreb, R.N. Effect of bimatoprost on intraocular pressure in prostaglandin FP receptor knockout mice. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4571–4577. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Dinh, T.; Woodward, D.F.; Holland, M.; Yuan, Y.-D.; Lin, T.-H.; Wheeler, L.A. Bimatoprost: Mechanism of ocular surface hyperemia associated with topical therapy. Cardiovasc. Drug Rev. 2005, 23, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Stamer, W.D.; Piwnica, D.; Jolas, T.; Carling, R.W.; Cornell, C.L.; Fliri, H.; Martos, J.; Pettit, S.N.; Wang, J.W.; Woodward, D.F. Cellular basis for bimatoprost effects on human conventional outflow. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5176–5181. [Google Scholar] [CrossRef]
- Bartoe, J.T.; Davidson, H.J.; Horton, M.T.; Jung, Y.; Brightman, A.H. The effects of bimatoprost and unoprostone isopropyl on the intraocular pressure of normal cats. Vet. Ophthalmol. 2005, 8, 247–252. [Google Scholar] [CrossRef]
- Ogundele, A.B.; Earnest, D.; McLaughlin, M.A. In vivo comparative study of ocular vasodilation, a relative indicator of hyperemia, in guinea pigs following treatment with bimatoprost ophthalmic solutions 0.01% and 0.03%. Clin. Ophthalmol. 2010, 4, 649–652. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.S.; Burke, J.; Shen, J.; Almazan, A.; Orilla, W.; Hughes, P.; Zhang, J.; Li, H.; Struble, C.; Miller, P.E.; et al. Bimatoprost sustained-release intracameral implant reduces episcleral venous pressure in dogs. Vet. Ophthalmol. 2018, 21, 376–381. [Google Scholar] [CrossRef] [Green Version]
- Fukano, Y.; Kawazu, K. Disposition and metabolism of a novel prostanoid antiglaucoma medication, tafluprost, following ocular administration to rats. Drug Metab. Dispos. 2009, 37, 1622–1634. [Google Scholar] [CrossRef] [Green Version]
- Kanamori, A.; Naka, M.; Fukuda, M.; Nakamura, M.; Negi, A. Tafluprost protects rat retinal ganglion cells from apoptosis in vitro and in vivo. Graefes Arch. Clin. Exp. Ophthalmol. 2009, 247, 1353–1360. [Google Scholar] [CrossRef]
- Mayama, C.; Ishii, K.; Saeki, T.; Ota, T.; Tomidokoro, A.; Araie, M. Effects of topical phenylephrine and tafluprost on optic nerve head circulation in monkeys with unilateral experimental glaucoma. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4117–4124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumi, N.; Nagaoka, T.; Sato, E.; Mori, F.; Takahashi, A.; Sogawa, K.; Yoshida, A. Short-term effects of topical tafluprost on retinal blood flow in cats. J. Ocul. Pharmacol. Ther. 2008, 24, 521–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Mao, W. Tafluprost once daily for treatment of elevated intraocular pressure in patients with open-angle glaucoma. Clin. Ophthalmol. 2013, 7, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Kwak, J.; Kang, S.; Lee, E.R.; Park, S.; Park, S.; Park, E.; Lim, J.; Seo, K. Effect of preservative-free tafluprost on intraocular pressure, pupil diameter, and anterior segment structures in normal canine eyes. Vet. Ophthalmol. 2017, 20, 34–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arfaee, F.; Armin, A. A comparison between the effect of topical tafluprost and latanoprost on intraocular pressure in healthy male guinea pigs. J. Exotic Pet Med. 2021, 39, 91–95. [Google Scholar] [CrossRef]
- Fuchshofer, R.; Kuespert, S.; Junglas, B.; Tamm, E.R. The prostaglandin f2α analog fluprostenol attenuates the fibrotic effects of connective tissue growth factor on human trabecular meshwork cells. J. Ocul. Pharmacol. Ther. 2014, 30, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Saeki, T.; Tsuruga, H.; Aihara, M.; Araie, M.; Rittenhouse, K. Dose-Response Profile of PF-03187207 (PF-207) and Peak IOP Lowering Response Following Single Topical Administration to FP Receptor Knockout Mice vs. Wild Type Mice. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4064. [Google Scholar]
- Krauss, A.H.P.; Impagnatiello, F.; Toris, C.B.; Gale, D.C.; Prasanna, G.; Borghi, V.; Chiroli, V.; Chong, W.K.M.; Carreiro, S.T.; Ongini, E. Ocular hypotensive activity of BOL-303259-X, a nitric oxide donating prostaglandin F2α agonist, in preclinical models. Exp. Eye Res. 2011, 93, 250–255. [Google Scholar] [CrossRef]
- Mehran, N.A.; Sinha, S.; Razeghinejad, R. New glaucoma medications: Latanoprostene bunod, netarsudil, and fixed combination netarsudil-latanoprost. Eye 2020, 34, 72–88. [Google Scholar] [CrossRef]
- Liu, H.K.; Chiou, G.C.; Garg, L.C. Ocular hypotensive effects of timolol in cat eyes. Arch. Ophthalmol. 1980, 98, 1467–1469. [Google Scholar] [CrossRef]
- Watanabe, K.; Chiou, G.C. Action mechanism of timolol to lower the intraocular pressure in rabbits. Ophthalmic Res. 1983, 15, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Schuettauf, F.; Quinto, K.; Naskar, R.; Zurakowski, D. Effects of anti-glaucoma medications on ganglion cell survival: The DBA/2J mouse model. Vision Res. 2002, 42, 2333–2337. [Google Scholar] [CrossRef] [Green Version]
- Goto, W.; Ota, T.; Morikawa, N.; Otori, Y.; Hara, H.; Kawazu, K.; Miyawaki, N.; Tano, Y. Protective effects of timolol against the neuronal damage induced by glutamate and ischemia in the rat retina. Brain Res. 2002, 958, 10–19. [Google Scholar] [CrossRef]
- Watson, P.; Stjernschantz, J. A six-month, randomized, double-masked study comparing latanoprost with timolol in open-angle glaucoma and ocular hypertension. The Latanoprost Study Group. Ophthalmology 1996, 103, 126–137. [Google Scholar] [CrossRef]
- Bartels, S.P. Aqueous humor flow measured with fluorophotometry in timolol-treated primates. Investig. Ophthalmol. Vis. Sci. 1988, 29, 1498–1504. [Google Scholar]
- Yu, D.Y.; Su, E.N.; Cringle, S.J.; Alder, V.A.; Yu, P.K.; Desantis, L. Effect of betaxolol, timolol and nimodipine on human and pig retinal arterioles. Exp. Eye Res. 1998, 67, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.N.; Miller, P.E.; Felchle, L.M. Effects of topical administration of latanoprost, timolol, or a combination of latanoprost and timolol on intraocular pressure, pupil size, and heart rate in clinically normal dogs. Am. J. Vet. Res. 2010, 71, 1055–1061. [Google Scholar] [CrossRef]
- Millar, J.C.; Clark, A.F.; Pang, I.-H. Assessment of aqueous humor dynamics in the mouse by a novel method of constant-flow infusion. Investig. Ophthalmol. Vis. Sci. 2011, 52, 685–694. [Google Scholar] [CrossRef]
- Wood, J.P.; DeSantis, L.; Chao, H.M.; Osborne, N.N. Topically applied betaxolol attenuates ischaemia-induced effects to the rat retina and stimulates BDNF mRNA. Exp. Eye Res. 2001, 72, 79–86. [Google Scholar] [CrossRef]
- Osborne, N.N.; DeSantis, L.; Bae, J.H.; Ugarte, M.; Wood, J.P.; Nash, M.S.; Chidlow, G. Topically applied betaxolol attenuates NMDA-induced toxicity to ganglion cells and the effects of ischaemia to the retina. Exp. Eye Res. 1999, 69, 331–342. [Google Scholar] [CrossRef]
- Uji, Y.; Kuze, M.; Matubara, H.; Doi, M.; Sasoh, M. Effects of the beta1-selective adrenergic antagonist betaxolol on electroretinography in the perfused cat eye. Doc. Ophthalmol. 2003, 106, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Holló, G.; Whitson, J.T.; Faulkner, R.; McCue, B.; Curtis, M.; Wieland, H.; Chastain, J.; Sanders, M.; DeSantis, L.; Przydryga, J.; et al. Concentrations of betaxolol in ocular tissues of patients with glaucoma and normal monkeys after 1 month of topical ocular administration. Investig. Ophthalmol. Vis. Sci. 2006, 47, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, Y.; Araie, M.; Tomita, K.; Nagahara, M. Effect of topical betaxolol on tissue circulation in the human optic nerve head. J. Ocul. Pharmacol. Ther. 1999, 15, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.E.; Schmidt, G.M.; Vainisi, S.J.; Swanson, J.F.; Herrmann, M.K. The efficacy of topical prophylactic antiglaucoma therapy in primary closed angle glaucoma in dogs: A multicenter clinical trial. J. Am. Anim. Hosp. Assoc. 2000, 36, 431–438. [Google Scholar] [CrossRef]
- Goldenberg-Cohen, N.; Dadon-Bar-El, S.; Hasanreisoglu, M.; Avraham-Lubin, B.C.R.; Dratviman-Storobinsky, O.; Cohen, Y.; Weinberger, D. Possible neuroprotective effect of brimonidine in a mouse model of ischaemic optic neuropathy. Clin. Exp. Ophthalmol. 2009, 37, 718–729. [Google Scholar] [CrossRef]
- Acheampong, A.A.; Small, D.; Baumgarten, V.; Welty, D.; Tang-Liu, D. Formulation effects on ocular absorption of brimonidine in rabbit eyes. J. Ocul. Pharmacol. Ther. 2002, 18, 325–337. [Google Scholar] [CrossRef]
- Gelatt, K.N.; MacKay, E.O. Effect of single and multiple doses of 0.2% brimonidine tartrate in the glaucomatous Beagle. Vet. Ophthalmol. 2002, 5, 253–262. [Google Scholar] [CrossRef]
- Burke, J.; Schwartz, M. Preclinical evaluation of brimonidine. Surv. Ophthalmol. 1996, 41, S9–S18. [Google Scholar] [CrossRef]
- Schnichels, S.; Hurst, J.; de Vries, J.W.; Ullah, S.; Frößl, K.; Gruszka, A.; Löscher, M.; Bartz-Schmidt, K.-U.; Spitzer, M.S.; Herrmann, A. Improved Treatment Options for Glaucoma with Brimonidine-Loaded Lipid DNA Nanoparticles. ACS Appl. Mater. Interfaces 2021, 13, 9445–9456. [Google Scholar] [CrossRef]
- Tamhane, M.; Luu, K.T.; Attar, M. Ocular pharmacokinetics of brimonidine drug delivery system in monkeys and translational modeling for selection of dose and frequency in clinical trials. J. Pharmacol. Exp. Ther. 2021, 378, 207–214. [Google Scholar] [CrossRef]
- Toris, C.B.; Gleason, M.L.; Camras, C.B.; Yablonski, M.E. Effects of brimonidine on aqueous humor dynamics in human eyes. Arch. Ophthalmol. 1995, 113, 1514–1517. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Y.; Lv, H.; Jiang, X.; Zhang, M.; Li, X. α-adrenergic agonist brimonidine control of experimentally induced myopia in guinea pigs: A pilot study. Mol. Vis. 2017, 23, 785–798. [Google Scholar] [PubMed]
- Morrison, J.C.; Nylander, K.B.; Lauer, A.K.; Cepurna, W.O.; Johnson, E. Glaucoma drops control intraocular pressure and protect optic nerves in a rat model of glaucoma. Investig. Ophthalmol. Vis. Sci. 1998, 39, 526–531. [Google Scholar]
- Gabelt, B.T.; Robinson, J.C.; Hubbard, W.C.; Peterson, C.M.; Debink, N.; Wadhwa, A.; Kaufman, P.L. Apraclonidine and brimonidine effects on anterior ocular and cardiovascular physiology in normal and sympathectomized monkeys. Exp. Eye Res. 1994, 59, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Toris, C.B.; Tafoya, M.E.; Camras, C.B.; Yablonski, M.E. Effects of apraclonidine on aqueous humor dynamics in human eyes. Ophthalmology 1995, 102, 456–461. [Google Scholar] [CrossRef]
- Orgül, S.; Bacon, D.R.; Van Buskirk, E.M.; Cioffi, G.A. Optic nerve vasomotor effects of topical apraclonidine hydrochloride. Br. J. Ophthalmol. 1996, 80, 82–84. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.E.; Nelson, M.J.; Rhaesa, S.L. Effects of topical administration of 0.5% apraclonidine on intraocular pressure, pupil size, and heart rate in clinically normal dogs. Am. J. Vet. Res. 1996, 57, 79–82. [Google Scholar]
- Miller, P.E.; Rhaesa, S.L. Effects of topical administration of 0.5% apraclonidine on intraocular pressure, pupil size, and heart rate in clinically normal cats. Am. J. Vet. Res. 1996, 57, 83–86. [Google Scholar]
- Li, T.; Wang, Y.; Chen, J.; Gao, X.; Pan, S.; Su, Y.; Zhou, X. Co-delivery of brinzolamide and miRNA-124 by biodegradable nanoparticles as a strategy for glaucoma therapy. Drug Deliv. 2020, 27, 410–421. [Google Scholar] [CrossRef] [Green Version]
- Desantis, L. Preclinical overview of brinzolamide1. Surv. Ophthalmol. 2000, 44, S119–S129. [Google Scholar] [CrossRef]
- Li, N.; Shi, H.-M.; Cong, L.; Lu, Z.-Z.; Ye, W.; Zhang, Y.-Y. Outflow facility efficacy of five drugs in enucleated porcine eyes by a method of constant-pressure perfusion. Int. J. Clin. Exp. Med. 2015, 8, 7184–7191. [Google Scholar] [PubMed]
- Di, Y.; Luo, X.-M.; Qiao, T.; Lu, N. Intraocular pressure with rebound tonometry and effects of topical intraocular pressure reducing medications in guinea pigs. Int. J. Ophthalmol. 2017, 10, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Toris, C.B.; Zhan, G.-L.; McLaughlin, M.A. Effects of brinzolamide on aqueous humor dynamics in monkeys and rabbits. J. Ocul. Pharmacol. Ther. 2003, 19, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Cvetkovic, R.S.; Perry, C.M. Brinzolamide: A review of its use in the management of primary open-angle glaucoma and ocular hypertension. Drugs Aging 2003, 20, 919–947. [Google Scholar] [CrossRef] [PubMed]
- Ingram, C.J.; Brubaker, R.F. Effect of brinzolamide and dorzolamide on aqueous humor flow in human eyes. Am. J. Ophthalmol. 1999, 128, 292–296. [Google Scholar] [CrossRef]
- Chandra, S.; Muir, E.R.; Deo, K.; Kiel, J.W.; Duong, T.Q. Effects of dorzolamide on retinal and choroidal blood flow in the DBA/2J mouse model of glaucoma. Investig. Ophthalmol. Vis. Sci. 2016, 57, 826–831. [Google Scholar] [CrossRef] [Green Version]
- Pitha, I.; Kimball, E.C.; Oglesby, E.N.; Pease, M.E.; Fu, J.; Schaub, J.; Kim, Y.-C.; Hu, Q.; Hanes, J.; Quigley, H.A. Sustained Dorzolamide Release Prevents Axonal and Retinal Ganglion Cell Loss in a Rat Model of IOP-Glaucoma. Transl. Vis. Sci. Technol. 2018, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Percicot, C.L.; Schnell, C.R.; Debon, C.; Hariton, C. Continuous intraocular pressure measurement by telemetry in alpha-chymotrypsin-induced glaucoma model in the rabbit: Effects of timolol, dorzolamide, and epinephrine. J. Pharmacol. Toxicol. Methods 1996, 36, 223–228. [Google Scholar] [CrossRef]
- Stefánsson, E.; Jensen, P.K.; Eysteinsson, T.; Bang, K.; Kiilgaard, J.F.; Dollerup, J.; Scherfig, E.; la Cour, M. Optic Nerve Oxygen Tension in Pigs and the Effect of Carbonic Anhydrase Inhibitors. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2756–2761. [Google Scholar]
- Dietrich, U.M.; Chandler, M.J.; Cooper, T.; Vidyashankar, A.; Chen, G. Effects of topical 2% dorzolamide hydrochloride alone and in combination with 0.5% timolol maleate on intraocular pressure in normal feline eyes. Vet. Ophthalmol. 2007, 10, 95–100. [Google Scholar] [CrossRef]
- Gelatt, K.N.; MacKay, E.O. Changes in intraocular pressure associated with topical dorzolamide and oral methazolamide in glaucomatous dogs. Vet. Ophthalmol. 2001, 4, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.F.; Serle, J.B.; Gagliuso, D.J.; Podos, S.M. Comparison of the ocular hypotensive effect of brimonidine, dorzolamide, latanoprost, or artificial tears added to timolol in glaucomatous monkey eyes. J. Glaucoma 2000, 9, 458–462. [Google Scholar] [CrossRef]
- Larsson, L.I.; Alm, A. Aqueous humor flow in human eyes treated with dorzolamide and different doses of acetazolamide. Arch. Ophthalmol. 1998, 116, 19–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avila, M.Y.; Stone, R.A.; Civan, M.M. Knockout of A3 Adenosine Receptors Reduces Mouse Intraocular Pressure. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3021–3026. [Google Scholar]
- Findl, O.; Hansen, R.M.; Fulton, A.B. The effects of acetazolamide on the electroretinographic responses in rats. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1019–1026. [Google Scholar]
- Kaur, I.P.; Singh, M.; Kanwar, M. Formulation and evaluation of ophthalmic preparations of acetazolamide. Int. J. Pharm. 2000, 199, 119–127. [Google Scholar] [CrossRef]
- Maren, T.H. Ion secretion into the posterior aqueous humor of dogs and monkeys. Exp. Eye Res. 1977, 25, 245–247. [Google Scholar] [CrossRef]
- Macri, F.J.; Dixon, R.L.; Rall, D.P. Aqueous humor turnover rates in the cat. I. Effect of acetazolamide. Investig. Ophthalmol. 1965, 4, 927–934. [Google Scholar]
- Fridriksdóttir, H.; Loftsson, T.; Stefánsson, E. Formulation and testing of methazolamide cyclodextrin eye drop solutions. J. Control. Release 1997, 44, 95–99. [Google Scholar] [CrossRef]
- Guđmundsdóttir, E.; Stefánsson, E.; Bjarnadóttir, G.; Sigurjónsdóttir, J.F.; Guđmundsdóttir, G.; Masson, M.; Loftsson, T. Methazolamide 1% in Cyclodextrin Solution Lowers IOP in Human Ocular Hypertension. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3552–3554. [Google Scholar]
- Li, G.; Mukherjee, D.; Navarro, I.; Ashpole, N.E.; Sherwood, J.M.; Chang, J.; Overby, D.R.; Yuan, F.; Gonzalez, P.; Kopczynski, C.C.; et al. Visualization of conventional outflow tissue responses to netarsudil in living mouse eyes. Eur. J. Pharmacol. 2016, 787, 20–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, R.; Li, G.; Le, T.D.; Kopczynski, C.; Stamer, W.D.; Gong, H. Netarsudil increases outflow facility in human eyes through multiple mechanisms. Investig. Ophthalmol. Vis. Sci. 2016, 57, 6197–6209. [Google Scholar] [CrossRef] [PubMed]
- Leary, K.A.; Lin, K.-T.; Steibel, J.P.; Harman, C.D.; Komáromy, A.M. Safety and efficacy of topically administered netarsudil (RhopressaTM) in normal and glaucomatous dogs with ADAMTS10-open-angle glaucoma (ADAMTS10-OAG). Vet. Ophthalmol. 2021, 24, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Kitaoka, Y.; Sase, K.; Tsukahara, C.; Fujita, N.; Arizono, I.; Kogo, J.; Tokuda, N.; Takagi, H. Axonal Protection by Netarsudil, a ROCK Inhibitor, Is Linked to an AMPK-Autophagy Pathway in TNF-Induced Optic Nerve Degeneration. Investig. Ophthalmol. Vis. Sci. 2022, 63, 4. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-W.; Sherman, B.; Moore, L.A.; Laethem, C.L.; Lu, D.-W.; Pattabiraman, P.P.; Rao, P.V.; deLong, M.A.; Kopczynski, C.C. Discovery and preclinical development of netarsudil, a novel ocular hypotensive agent for the treatment of glaucoma. J. Ocul. Pharmacol. Ther. 2018, 34, 40–51. [Google Scholar] [CrossRef]
- Isobe, T.; Kasai, T.; Kawai, H. Ocular penetration and pharmacokinetics of ripasudil following topical administration to rabbits. J. Ocul. Pharmacol. Ther. 2016, 32, 405–414. [Google Scholar] [CrossRef]
- Kamiya, T.; Omae, T.; Nakabayashi, S.; Takahashi, K.; Tanner, A.; Yoshida, A. Effect of Rho Kinase Inhibitor Ripasudil (K-115) on Isolated Porcine Retinal Arterioles. J. Ocul. Pharmacol. Ther. 2021, 37, 104–111. [Google Scholar] [CrossRef]
- Nakabayashi, S.; Kawai, M.; Yoshioka, T.; Song, Y.-S.; Tani, T.; Yoshida, A.; Nagaoka, T. Effect of intravitreal Rho kinase inhibitor ripasudil (K-115) on feline retinal microcirculation. Exp. Eye Res. 2015, 139, 132–135. [Google Scholar] [CrossRef]
- Nishijima, E.; Namekata, K.; Kimura, A.; Guo, X.; Harada, C.; Noro, T.; Nakano, T.; Harada, T. Topical ripasudil stimulates neuroprotection and axon regeneration in adult mice following optic nerve injury. Sci. Rep. 2020, 10, 15709. [Google Scholar] [CrossRef]
- Wada, Y.; Higashide, T.; Nagata, A.; Sugiyama, K. Effects of ripasudil, a rho kinase inhibitor, on blood flow in the optic nerve head of normal rats. Graefe’s Arch. Clin. Exp. Ophthalmol. 2019, 257, 303–311. [Google Scholar] [CrossRef]
- Inoue, T.; Tanihara, H. Ripasudil hydrochloride hydrate: Targeting Rho kinase in the treatment of glaucoma. Expert Opin. Pharmacother. 2017, 18, 1669–1673. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Maruyama, K.; Himori, N.; Omodaka, K.; Yokoyama, Y.; Shiga, Y.; Morin, R.; Nakazawa, T. The novel Rho kinase (ROCK) inhibitor K-115: A new candidate drug for neuroprotective treatment in glaucoma. Investig. Ophthalmol. Vis. Sci. 2014, 55, 7126–7136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.; Gao, D. Fasudil, a Rho-associated protein kinase inhibitor, attenuates retinal ischemia and reperfusion injury in rats. Int. J. Mol. Med. 2011, 28, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Khallaf, A.M.; El-Moslemany, R.M.; Ahmed, M.F.; Morsi, M.H.; Khalafallah, N.M. Exploring a Novel Fasudil-Phospholipid Complex Formulated as Liposomal Thermosensitive in situ Gel for Glaucoma. Int. J. Nanomed. 2022, 17, 163–181. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, M.; Yoshida, J.; Saito, K.; Sagawa, H.; Tokita, Y.; Watanabe, M. Differential effects of two ROCK inhibitors, Fasudil and Y-27632, on optic nerve regeneration in adult cats. Brain Res. 2008, 1201, 23–33. [Google Scholar] [CrossRef]
- Pakravan, M.; Beni, A.N.; Ghahari, E.; Varshochian, R.; Yazdani, S.; Esfandiari, H.; Ahmadieh, H. The Ocular Hypotensive Efficacy of Topical Fasudil, a Rho-Associated Protein Kinase Inhibitor, in Patients with End-Stage Glaucoma. Am. J. Ther. 2016, 24, e676–e680. [Google Scholar] [CrossRef]
- Da, B.; Cao, Y.; Wei, H.; Chen, Z.; Shui, Y.; Li, Z. Antagonistic effects of tranilast on proliferation and collagen synthesis induced by TGF-beta2 in cultured human trabecular meshwork cells. J. Huazhong Univ. Sci. Technol. Med. Sci. 2004, 24, 490–496. [Google Scholar] [CrossRef]
- Cao, Y.; Hu, Y.; Li, J.; Shui, Y.; Da, B.; Wei, H. Effect of Tranilast on Collagen Synthesis and TGF–Beta2 Expression of Cultured Human Lamina Cribrosa Astrocytes. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1544. [Google Scholar]
- Spitzer, M.S.; Sat, M.; Schramm, C.; Schnichels, S.; Schultheiss, M.; Yoeruek, E.; Dzhelebov, D.; Szurman, P. Biocompatibility and antifibrotic effect of UV-cross-linked hyaluronate as a release-system for tranilast after trabeculectomy in a rabbit model—A pilot study. Curr. Eye Res. 2012, 37, 463–470. [Google Scholar] [CrossRef]
- Pfeiffer, N.; Voykov, B.; Renieri, G.; Bell, K.; Richter, P.; Weigel, M.; Thieme, H.; Wilhelm, B.; Lorenz, K.; Feindor, M.; et al. First-in-human phase I study of ISTH0036, an antisense oligonucleotide selectively targeting transforming growth factor beta 2 (TGF-β2), in subjects with open-angle glaucoma undergoing glaucoma filtration surgery. PLoS ONE 2017, 12, e0188899. [Google Scholar] [CrossRef] [Green Version]
- Hasenbach, K.; Van Bergen, T.; Vandewalle, E.; De Groef, L.; Van Hove, I.; Moons, L.; Stalmans, I.; Fettes, P.; Leo, E.; Wosikowski, K.; et al. Potent and selective antisense oligonucleotides targeting the transforming growth factor beta (TGF-β) isoforms in advanced glaucoma: A preclinical evaluation. MAIO 2016, 1, 20–28. [Google Scholar] [CrossRef]
- Nakamura, H.; Siddiqui, S.S.; Shen, X.; Malik, A.B.; Pulido, J.S.; Kumar, N.M.; Yue, B.Y.J.T. RNA interference targeting transforming growth factor-beta type II receptor suppresses ocular inflammation and fibrosis. Mol. Vis. 2004, 10, 703–711. [Google Scholar] [PubMed]
- Mead, A.L.; Wong, T.T.L.; Cordeiro, M.F.; Anderson, I.K.; Khaw, P.T. Evaluation of Anti-TGF-β2 Antibody as a New Postoperative Anti-scarring Agent in Glaucoma Surgery. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3394. [Google Scholar] [CrossRef] [Green Version]
- CAT-152 0102 Trabeculectomy Study Group; Khaw, P.; Grehn, F.; Holló, G.; Overton, B.; Wilson, R.; Vogel, R.; Smith, Z. A phase III study of subconjunctival human anti-transforming growth factor beta(2) monoclonal antibody (CAT-152) to prevent scarring after first-time trabeculectomy. Ophthalmology 2007, 114, 1822–1830. [Google Scholar] [CrossRef] [PubMed]
- Shan, S.-W.; Do, C.-W.; Lam, T.C.; Li, H.-L.; Stamer, W.D.; To, C.-H. Thrombospondin-1 mediates Rho-kinase inhibitor-induced increase in outflow-facility. J. Cell. Physiol. 2021, 236, 8226–8238. [Google Scholar] [CrossRef]
- Chen, W.-S.; Cao, Z.; Krishnan, C.; Panjwani, N. Verteporfin without light stimulation inhibits YAP activation in trabecular meshwork cells: Implications for glaucoma treatment. Biochem. Biophys. Res. Commun. 2015, 466, 221–225. [Google Scholar] [CrossRef]
- Matsubara, A.; Nakazawa, T.; Husain, D.; Iliaki, E.; Connolly, E.; Michaud, N.A.; Gragoudas, E.S.; Miller, J.W. Investigating the effect of ciliary body photodynamic therapy in a glaucoma mouse model. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2498–2507. [Google Scholar] [CrossRef]
- Parodi, M.B.; Iacono, P. Photodynamic therapy with verteporfin for anterior segment neovascularizations in neovascular glaucoma. Am. J. Ophthalmol. 2004, 138, 157–158. [Google Scholar] [CrossRef]
- Ko, M.-L.; Chen, C.-F.; Peng, P.-H.; Peng, Y.-H. Simvastatin upregulates Bcl-2 expression and protects retinal neurons from early ischemia/reperfusion injury in the rat retina. Exp. Eye Res. 2011, 93, 580–585. [Google Scholar] [CrossRef]
- Krempler, K.; Schmeer, C.W.; Isenmann, S.; Witte, O.W.; Löwel, S. Simvastatin improves retinal ganglion cell survival and spatial vision after acute retinal ischemia/reperfusion in mice. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2606–2618. [Google Scholar] [CrossRef]
- Nagaoka, T.; Takahashi, A.; Sato, E.; Izumi, N.; Hein, T.W.; Kuo, L.; Yoshida, A. Effect of systemic administration of simvastatin on retinal circulation. Arch. Ophthalmol. 2006, 124, 665–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaoka, T.; Hein, T.W.; Yoshida, A.; Kuo, L. Simvastatin elicits dilation of isolated porcine retinal arterioles: Role of nitric oxide and mevalonate-rho kinase pathways. Investig. Ophthalmol. Vis. Sci. 2007, 48, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-L.; Sung, K.R.; Shin, J.A.; Young Yoon, J.; Jang, J. Statins reduce TGF-beta2-modulation of the extracellular matrix in cultured astrocytes of the human optic nerve head. Exp. Eye Res. 2017, 164, 55–63. [Google Scholar] [CrossRef]
- Kim, M.; Shin, J.; Sung, K. Statins regulate MMP-2 and MMP-9 secretion and activation in human ONH astrocytes. Investig. Ophthalmol. Vis. Sci. 2018, 59, 6145. [Google Scholar]
- Villarreal, G.; Chatterjee, A.; Oh, S.S.; Oh, D.-J.; Rhee, D.J. Pharmacological regulation of SPARC by lovastatin in human trabecular meshwork cells. Investig. Ophthalmol. Vis. Sci. 2014, 55, 1657–1665. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-H.; Yoo, C.; Kim, Y.Y. Effect of Lovastatin on Wound-Healing Modulation After Glaucoma Filtration Surgery in a Rabbit Model. Investig. Ophthalmol. Vis. Sci. 2016, 57, 1871–1877. [Google Scholar] [CrossRef] [Green Version]
- Song, X.-Y.; Chen, Y.-Y.; Liu, W.-T.; Cong, L.; Zhang, J.-L.; Zhang, Y.; Zhang, Y.-Y. Atorvastatin reduces IOP in ocular hypertension in vivo and suppresses ECM in trabecular meshwork perhaps via FGD4. Int. J. Mol. Med. 2022, 49, 76. [Google Scholar] [CrossRef]
- Kim, M.-L.; Sung, K.R.; Kwon, J.; Choi, G.W.; Shin, J.A. Neuroprotective effect of statins in a rat model of chronic ocular hypertension. Int. J. Mol. Sci. 2021, 22, 12500. [Google Scholar] [CrossRef]
- Cong, L.; Fu, S.; Zhang, J.; Zhao, J.; Zhang, Y. Effects of atorvastatin on porcine aqueous humour outflow and trabecular meshwork cells. Exp. Ther. Med. 2018, 15, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, R.; Krasilnikova, A.; Mohamed, S.N.L. Topical losartan reduces IOP by altering TM morphology in rats with steroid-induced ocular hypertension. Indian J. Physiol. 2018, 62, 238–248. [Google Scholar]
- Shah, G.B.; Sharma, S.; Mehta, A.A.; Goyal, R.K. Oculohypotensive effect of angiotensin-converting enzyme inhibitors in acute and chronic models of glaucoma. J. Cardiovasc. Pharmacol. 2000, 36, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Costagliola, C.; Verolino, M.; De Rosa, M.L.; Iaccarino, G.; Ciancaglini, M.; Mastropasqua, L. Effect of oral losartan potassium administration on intraocular pressure in normotensive and glaucomatous human subjects. Exp. Eye Res. 2000, 71, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Sawaguchi, S.; Yue, B.Y.; Yeh, P.; Tso, M.O. Effects of intracameral injection of chondroitinase ABC in vivo. Arch. Ophthalmol. 1992, 110, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Murienne, B.J.; Chen, M.L.; Quigley, H.A.; Nguyen, T.D. The contribution of glycosaminoglycans to the mechanical behaviour of the posterior human sclera. J. R. Soc. Interface 2016, 13, 20160367. [Google Scholar] [CrossRef]
- Kirihara, T.; Shimazaki, A.; Nakamura, M.; Miyawaki, N. Ocular hypotensive efficacy of Src-family tyrosine kinase inhibitors via different cellular actions from Rock inhibitors. Exp. Eye Res. 2014, 119, 97–105. [Google Scholar] [CrossRef]
- Belmadani, S.; Bernal, J.; Wei, C.-C.; Pallero, M.A.; Dell’italia, L.; Murphy-Ullrich, J.E.; Berecek, K.H. A thrombospondin-1 antagonist of transforming growth factor-beta activation blocks cardiomyopathy in rats with diabetes and elevated angiotensin II. Am. J. Pathol. 2007, 171, 777–789. [Google Scholar] [CrossRef] [Green Version]
- Lu, A.; Miao, M.; Schoeb, T.R.; Agarwal, A.; Murphy-Ullrich, J.E. Blockade of TSP1-dependent TGF-β activity reduces renal injury and proteinuria in a murine model of diabetic nephropathy. Am. J. Pathol. 2011, 178, 2573–2586. [Google Scholar] [CrossRef] [Green Version]
- Leask, A. Breathe, breathe in the air: The anti-CCN2 antibody pamrevlumab (FG-3019) completes a successful phase II clinical trial for idiopathic pulmonary fibrosis. J. Cell Commun. Signal. 2019, 13, 441–442. [Google Scholar] [CrossRef]
- Wang, J.; Harris, A.; Prendes, M.A.; Alshawa, L.; Gross, J.C.; Wentz, S.M.; Rao, A.B.; Kim, N.J.; Synder, A.; Siesky, B. Targeting Transforming Growth Factor-β Signaling in Primary Open-Angle Glaucoma. J. Glaucoma 2017, 26, 390–395. [Google Scholar] [CrossRef]
- Webber, H.C.; Bermudez, J.Y.; Sethi, A.; Clark, A.F.; Mao, W. Crosstalk between TGFβ and Wnt signaling pathways in the human trabecular meshwork. Exp. Eye Res. 2016, 148, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Wordinger, R.J.; Sharma, T.; Clark, A.F. The role of TGF-β2 and bone morphogenetic proteins in the trabecular meshwork and glaucoma. J. Ocul. Pharmacol. Ther. 2014, 30, 154–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanihara, H.; Inoue, T.; Yamamoto, T.; Kuwayama, Y.; Abe, H.; Suganami, H.; Araie, M.; K-115 Clinical Study Group. Intra-ocular pressure-lowering effects of a Rho kinase inhibitor, ripasudil (K-115), over 24 hours in primary open-angle glaucoma and ocular hypertension: A randomized, open-label, crossover study. Acta Ophthalmol. 2015, 93, e254–e260. [Google Scholar] [CrossRef]
- Tanihara, H.; Inoue, T.; Yamamoto, T.; Kuwayama, Y.; Abe, H.; Araie, M.; K-115 Clinical Study Group. Phase 2 randomized clinical study of a Rho kinase inhibitor, K-115, in primary open-angle glaucoma and ocular hypertension. Am. J. Ophthalmol. 2013, 156, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Hirooka, K.; Nitta, E.; Ukegawa, K.; Tsujikawa, A. Additive intraocular pressure lowering effects of the rho kinase inhibitor, ripasudil in glaucoma patients not able to obtain adequate control after other maximal tolerated medical therapy. Adv. Ther. 2016, 33, 1628–1634. [Google Scholar] [CrossRef]
- Pokrovskaya, O.; Wallace, D.; O’Brien, C. The emerging role of statins in glaucoma pathological mechanisms and therapeutics. Open J. Ophthalmol. 2014, 4, 124–138. [Google Scholar] [CrossRef] [Green Version]
- McCann, P.; Hogg, R.E.; Fallis, R.; Azuara-Blanco, A. The Effect of Statins on Intraocular Pressure and on the Incidence and Progression of Glaucoma: A Systematic Review and Meta-Analysis. Investig. Ophthalmol. Vis. Sci. 2016, 57, 2729–2748. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Xiong, R.; Wu, Y.; Ha, J.; Wang, W.; Han, X.; He, M. Associations of statin use with the onset and progression of open-angle glaucoma: A systematic review and meta-analysis. EClinicalMedicine 2022, 46, 101364. [Google Scholar] [CrossRef]
- McGwin, G.; McNeal, S.; Owsley, C.; Girkin, C.; Epstein, D.; Lee, P.P. Statins and other cholesterol-lowering medications and the presence of glaucoma. Arch. Ophthalmol. 2004, 122, 822–826. [Google Scholar] [CrossRef] [Green Version]
- Weinreb, R.N. Enhancement of scleral macromolecular permeability with prostaglandins. Trans. Am. Ophthalmol. Soc. 2001, 99, 319–343. [Google Scholar]



{kind=link}
{kind=link}
| Mechanism of Action | Drug(s) | Impact on Optic Nerve Remodeling | Models Tested | References |
|---|---|---|---|---|
| Prostaglandin F receptor agonist | Bimatoprost, Latanoprost, Fluprostenol, Tafluprost, Travoprost | Upregulation of MMP-1, -3, -9 | Mouse, Rat, Rabbit, Guinea Pig, Cat, Dog, Pig, Primate, Human | [131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148,149,150,151,152,153,154,155,156] |
| Hybrid prostaglandin F receptor agonist and nitric oxide donator | Latanoprostene bunod | Upregulation of MMPs and decrease cell contractility | Mouse, Rabbit, Dog, Primate, Human | [157,158,159] |
| β-adrenoceptor antagonist | Betaxolol, Timolol | Increased blood flow velocity | Mouse, Rat, Rabbit, Cat, Dog, Pig, Primate, Human | [160,161,162,163,164,165,166,167,168,169,170,171,172,173,174] |
| α2-adrenergic agonist | Apraclonidine, Brimonidine | Anti-apoptotic; RGC survival signal | Mouse, Rat, Guinea Pig, Rabbit, Cat, Dog, Pig, Primate, Human | [139,175,176,177,178,179,180,181,182,183,184,185,186,187,188] |
| Carbonic anhydrase inhibitor | Acetazolamide, Brinzolamide, Dorzolamide, Methazolamide | Increased blood flow and oxygen tension | Mouse, Rat, Guinea Pig, Rabbit, Dog, Pig, Primate, Human | [189,190,191,192,193,194,195,196,197,198,199,200,201,202,203,204,205,206,207,208,209,210] |
| ROCK Inhibitor | Fasudil, Netarsudil, Ripasudil | Inhibits contractility and migration of fibroblasts; inhibits production of ECM; inhibits cell death pathways | Mouse, Rat, Rabbit, Dog, Primate, Human | [211,212,213,214,215,216,217,218,219,220,221,222,223,224,225,226] |
| Inhibits secretion of TGF-β | Tranilast | Prevents TGF-β mediated fibrotic responses by nearby cells | Rabbit, Human culture | [227,228,229] |
| Inhibit transcription of TGF-β | ISTH0036, TbetaRII (RNAi) | Decreased levels of TGF-β expression | Mouse, Human Culture, Human | [230,231,232] |
| Direct immunosuppression of TGF-β | Lerdelimumab | Targeted inactivation of TGF-β to prevent receptor binding | Rabbit, Human | [233,234] |
| Inhibit TSP1 binding to LAP | LSKL | Inhibits TSP1 mediated activation of latent TGF-β | Mouse | [235] |
| Direct immunosuppression of CTGF | Pamrevlumab | Inhibits CTGF interaction with TGF-β | Human Culture | [37] |
| Reduce YAP and CTGF expression | Verteporfin (without light activation) | Reduces cell contractility via YAP; reduces CTGF interaction with TGF-β | Mouse, Human Culture, Human | [236,237,238] |
| Increased nitric oxide production | Atorvastatin, Lovastatin, Simvastatin | Inhibit RhoA/ROCK pathway and reduce levels of MMP-2 and -9, decrease cell contractility | Mouse, Rat, Rabbit, Dog, Pig, Human Culture | [65,239,240,241,242,243,244,245,246,247,248,249] |
| Angiotensin 1 receptor (AT1R) inhibitor | Losartan | Inhibits Smad2 phosphorylation | Mice, Rat, Rabbit, Human | [130,250,251,252] |
| Glycosaminoglycan degrading enzyme | Chondroitinase ABC | Weakens ECM (reduces stiffness) | Rat, Pig, Primate, Human Culture | [125,128,253,254] |
| Inhibit myosin light chain phosphorylation | Src-family tyrosine kinase (SFK) inhibitors (PP2) | Alters cell adhesion, reduces cell contractility, and permeability of cell layers | Rabbit, Human Culture | [108,255] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strickland, R.G.; Garner, M.A.; Gross, A.K.; Girkin, C.A. Remodeling of the Lamina Cribrosa: Mechanisms and Potential Therapeutic Approaches for Glaucoma. Int. J. Mol. Sci. 2022, 23, 8068. https://doi.org/10.3390/ijms23158068
Strickland RG, Garner MA, Gross AK, Girkin CA. Remodeling of the Lamina Cribrosa: Mechanisms and Potential Therapeutic Approaches for Glaucoma. International Journal of Molecular Sciences. 2022; 23(15):8068. https://doi.org/10.3390/ijms23158068
Chicago/Turabian StyleStrickland, Ryan G., Mary Anne Garner, Alecia K. Gross, and Christopher A. Girkin. 2022. "Remodeling of the Lamina Cribrosa: Mechanisms and Potential Therapeutic Approaches for Glaucoma" International Journal of Molecular Sciences 23, no. 15: 8068. https://doi.org/10.3390/ijms23158068
APA StyleStrickland, R. G., Garner, M. A., Gross, A. K., & Girkin, C. A. (2022). Remodeling of the Lamina Cribrosa: Mechanisms and Potential Therapeutic Approaches for Glaucoma. International Journal of Molecular Sciences, 23(15), 8068. https://doi.org/10.3390/ijms23158068