
Impact of Multiple Sclerosis Risk Polymorphism rs7665090 on MANBA Activity, Lysosomal Endocytosis, and Lymphocyte Activation

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population/Patients and Controls
2.2. Cell and DNA Extraction
2.3. Genotyping
2.4. Gene Expression
2.5. Assessment of β-Mannosidose Activity
2.6. Cell Activation
2.7. Endocytic Activity
2.8. Cell Metabolism
2.9. Statistical Analysis
3. Results
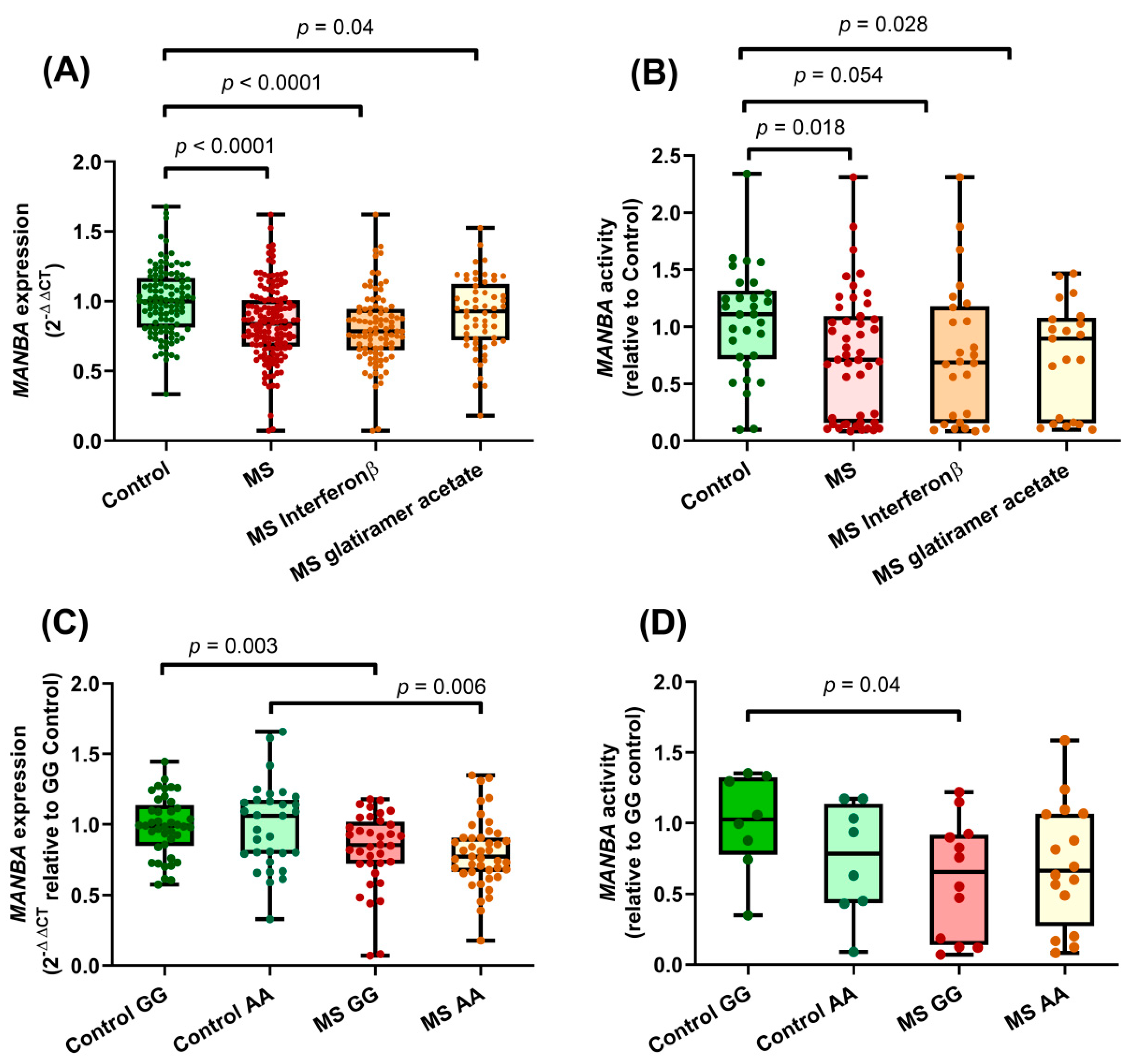
3.1. Decreased MANBA Expression and Enzymatic Activity in Lymphocytes from MS Patients Compared to Those from Healthy Controls
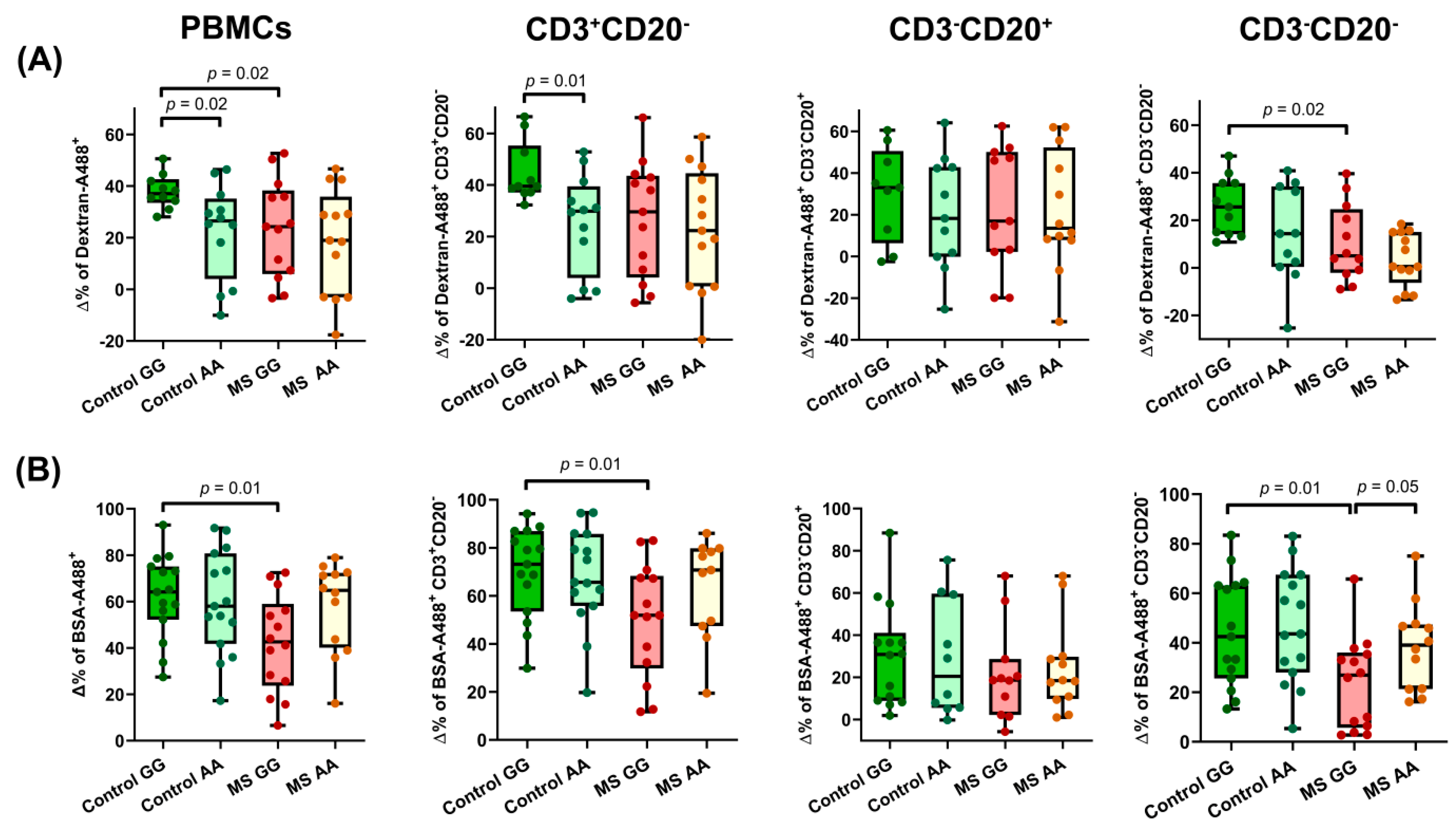
3.2. Endocytic Activity Is Modified by rs7665090
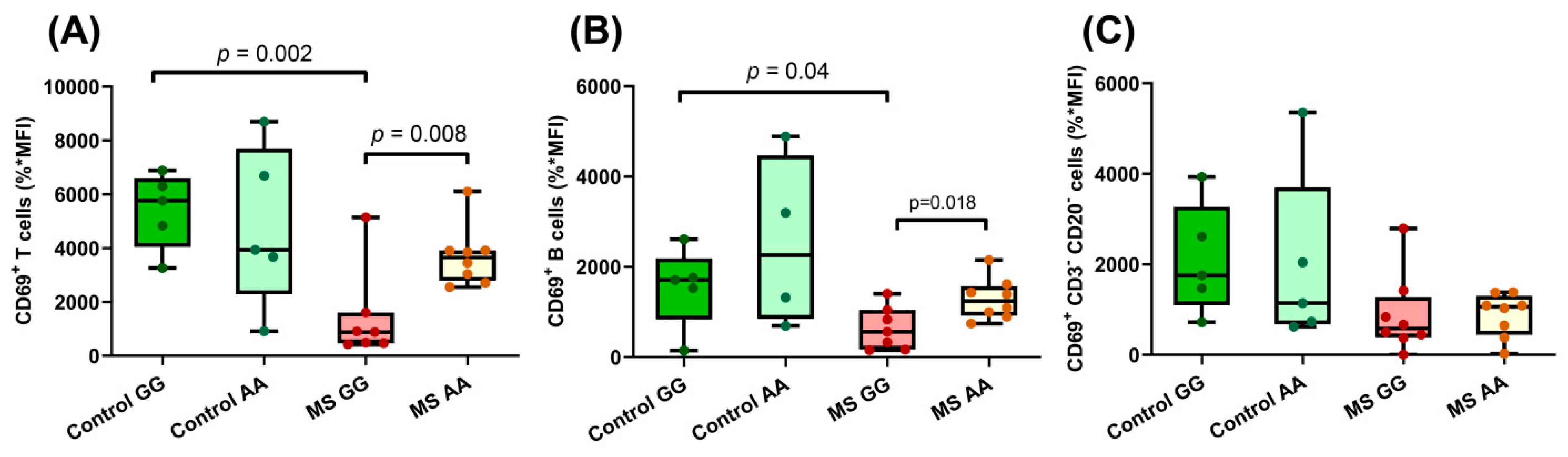
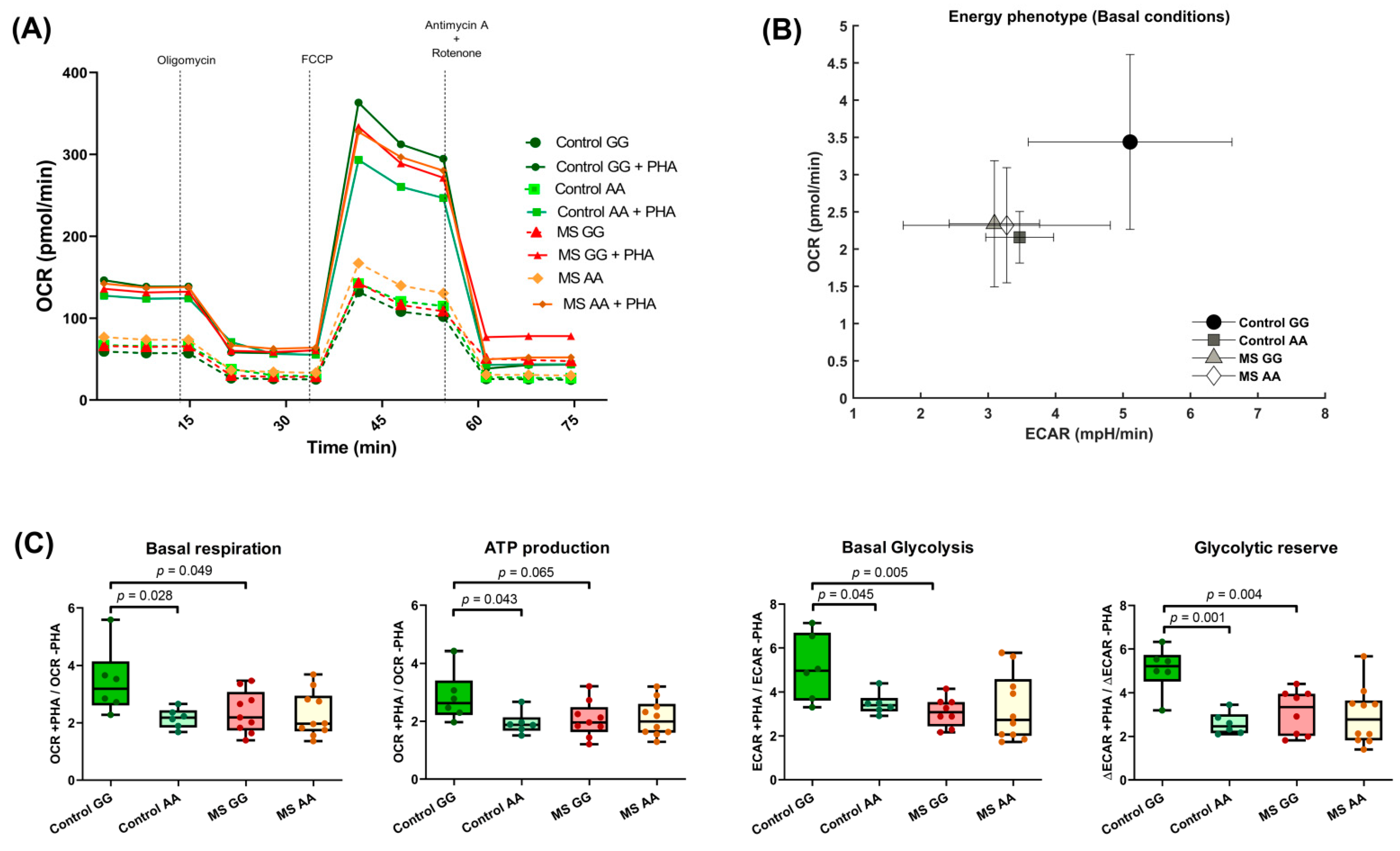
3.3. The SNP rs7665090 Modulates Lymphocyte Activation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reich, D.S.; Lucchinetti, C.F.; Calabresi, P.A. Multiple Sclerosis. N. Engl. J. Med. 2018, 378, 169–180. [Google Scholar] [CrossRef]
- Olsson, T.; Barcellos, L.F.; Alfredsson, L. Interactions between genetic, lifestyle and environmental risk factors for multiple sclerosis. Nat. Rev. Neurol. 2017, 13, 25–36. [Google Scholar] [CrossRef] [PubMed]
- IMSGC. Multiple sclerosis genomic map implicates peripheral immune cells and microglia in susceptibility. Science 2019, 365, eaav7188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IMSGC. A systems biology approach uncovers cell-specific gene regulatory effects of genetic associations in multiple sclerosis. Nat. Commun. 2019, 10, 2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mountjoy, E.; Schmidt, E.M.; Carmona, M.; Schwartzentruber, J.; Peat, G.; Miranda, A.; Fumis, L.; Hayhurst, J.; Buniello, A.; Karim, M.A.; et al. An open approach to systematically prioritize causal variants and genes at all published human GWAS trait-associated loci. Nat. Genet. 2021, 53, 1527–1533. [Google Scholar] [CrossRef]
- Roostaei, T.; Klein, H.U.; Ma, Y.; Felsky, D.; Kivisäkk, P.; Connor, S.M.; Kroshilina, A.; Yung, C.; Kaskow, B.J.; Shao, X.; et al. Proximal and distal effects of genetic susceptibility to multiple sclerosis on the T cell epigenome. Nat. Commun. 2021, 12, 7078. [Google Scholar] [CrossRef]
- Sun, B.B.; Kurki, M.I.; Foley, C.N.; Mechakra, A.; Chen, C.Y.; Marshall, E.; Wilk, J.B.; Chahine, M.; Chevalier, P.; Christé, G.; et al. Genetic associations of protein-coding variants in human disease. Nature 2022, 603, 95–102. [Google Scholar] [CrossRef]
- Yazar, S.; Alquicira-Hernandez, J.; Wing, K.; Senabouth, A.; Gordon, M.G.; Andersen, S.; Lu, Q.; Rowson, A.; Taylor, T.R.P.; Clarke, L.; et al. Single-cell eQTL mapping identifies cell type-specific genetic control of autoimmune disease. Science 2022, 376, eabf3041. [Google Scholar] [CrossRef]
- Gregory, A.P.; Dendrou, C.A.; Attfield, K.E.; Haghikia, A.; Xifara, D.K.; Butter, F.; Poschmann, G.; Kaur, G.; Lambert, L.; Leach, O.A.; et al. TNF receptor 1 genetic risk mirrors outcome of anti-TNF therapy in multiple sclerosis. Nature 2012, 488, 508–511. [Google Scholar] [CrossRef] [Green Version]
- Orrù, V.; Steri, M.; Sidore, C.; Marongiu, M.; Serra, V.; Olla, S.; Sole, G.; Lai, S.; Dei, M.; Mulas, A.; et al. Complex genetic signatures in immune cells underlie autoimmunity and inform therapy. Nat. Genet. 2020, 52, 1036–1045. [Google Scholar] [CrossRef]
- Galarza-Muñoz, G.; Briggs, F.B.S.; Evsyukova, I.; Schott-Lerner, G.; Kennedy, E.M.; Nyanhete, T.; Wang, L.; Bergamaschi, L.; Widen, S.G.; Tomaras, G.D.; et al. Human Epistatic Interaction Controls IL7R Splicing and Increases Multiple Sclerosis Risk. Cell 2017, 169, 72–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mena, J.; Alloza, I.; Tulloch Navarro, R.; Aldekoa, A.; Díez García, J.; Villanueva Etxebarria, A.; Lindskog, C.; Antigüedad, A.; Boyero, S.; Mendibe-Bilbao, M.D.M.; et al. Genomic Multiple Sclerosis Risk Variants Modulate the Expression of the ANKRD55-IL6ST Gene Region in Immature Dendritic Cells. Front. Immunol. 2021, 12, 816930. [Google Scholar] [CrossRef] [PubMed]
- Beecham, A.H.; Patsopoulos, N.A.; Xifara, D.K.; Davis, M.F.; Kemppinen, A.; Cotsapas, C.; Shah, T.S.; Spencer, C.; Booth, D.; Goris, A.; et al. Analysis of immune-related loci identifies 48 new susceptibility variants for multiple sclerosis. Nat. Genet. 2013, 45, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Lill, C.M.; Schjeide, B.M.; Graetz, C.; Ban, M.; Alcina, A.; Ortiz, M.A.; Pérez, J.; Damotte, V.; Booth, D.; Lopez de Lapuente, A.; et al. MANBA, CXCR5, SOX8, RPS6KB1 and ZBTB46 are genetic risk loci for multiple sclerosis. Brain 2013, 136, 1778–1782. [Google Scholar] [CrossRef] [Green Version]
- Winchester, B. Lysosomal metabolism of glycoproteins. Glycobiology 2005, 15, 1r–15r. [Google Scholar] [CrossRef] [Green Version]
- Cooper, A.; Sardharwalla, I.B.; Roberts, M.M. Human beta-mannosidase deficiency. N. Engl. J. Med. 1986, 315, 1231. [Google Scholar]
- Wenger, D.A.; Sujansky, E.; Fennessey, P.V.; Thompson, J.N. Human beta-mannosidase deficiency. N. Engl. J. Med. 1986, 315, 1201–1205. [Google Scholar] [CrossRef]
- Dorland, L.; Duran, M.; Hoefnagels, F.E.; Breg, J.N.; Fabery de Jonge, H.; Cransberg, K.; van Sprang, F.J.; van Diggelen, O.P. Beta-mannosidosis in two brothers with hearing loss. J. Inherit. Metab. Dis. 1988, 11 (Suppl. 2), 255–258. [Google Scholar] [CrossRef]
- Kleijer, W.J.; Hu, P.; Thoomes, R.; Boer, M.; Huijmans, J.G.; Blom, W.; Van Diggelen, O.P.; Seemanova, E.; Macek, M. Beta-mannosidase deficiency: Heterogeneous manifestation in the first female patient and her brother. J. Inherit. Metab. Dis. 1990, 13, 867–872. [Google Scholar] [CrossRef] [Green Version]
- Wijburg, H.; de Jong, J.; Wevers, R.; Bakkeren, J.; Trijbels, F.; Sengers, R. Beta-mannosidosis and ethanolaminuria in a female patient. Eur. J. Pediatrics 1992, 151, 311. [Google Scholar] [CrossRef]
- Yu, P.; Cui, Y.; Cai, W.; Wu, H.; Xiao, X.; Shao, Q.; Ma, L.; Guo, S.; Wu, N.; Jin, Z.B.; et al. Lysosomal storage disease in the brain: Mutations of the β-mannosidase gene identified in autosomal dominant nystagmus. Genet. Med. 2015, 17, 971–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Yao, T.; Cai, J.; Zhang, Q.; Li, S.; Li, H.; Fu, X.; Wu, J. A novel genetic variant potentially altering the expression of MANBA in the cerebellum associated with attention deficit hyperactivity disorder in Han Chinese children. World J. Biol. Psychiatry 2022, 1–12, On-line ahead or print. [Google Scholar] [CrossRef] [PubMed]
- Rørvig, S.; Østergaard, O.; Heegaard, N.H.; Borregaard, N. Proteome profiling of human neutrophil granule subsets, secretory vesicles, and cell membrane: Correlation with transcriptome profiling of neutrophil precursors. J. Leukoc. Biol. 2013, 94, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Thaysen-Andersen, M.; Venkatakrishnan, V.; Loke, I.; Laurini, C.; Diestel, S.; Parker, B.L.; Packer, N.H. Human neutrophils secrete bioactive paucimannosidic proteins from azurophilic granules into pathogen-infected sputum. J. Biol. Chem. 2015, 290, 8789–8802. [Google Scholar] [CrossRef] [Green Version]
- Ban, M.; Liao, W.; Baker, A.; Compston, A.; Thorpe, J.; Molyneux, P.; Fraser, M.; Khadake, J.; Jones, J.; Coles, A.; et al. Transcript specific regulation of expression influences susceptibility to multiple sclerosis. Eur. J. Hum. Genet. 2020, 28, 826–834. [Google Scholar] [CrossRef]
- Hauser, S.L.; Waubant, E.; Arnold, D.L.; Vollmer, T.; Antel, J.; Fox, R.J.; Bar-Or, A.; Panzara, M.; Sarkar, N.; Agarwal, S.; et al. B-cell depletion with rituximab in relapsing-remitting multiple sclerosis. N. Engl. J. Med. 2008, 358, 676–688. [Google Scholar] [CrossRef] [Green Version]
- Attfield, K.E.; Jensen, L.T.; Kaufmann, M.; Friese, M.A.; Fugger, L. The immunology of multiple sclerosis. Nat. Rev. Immunol. 2022. On-line ahead or print. [Google Scholar] [CrossRef]
- Polman, C.H.; Reingold, S.C.; Banwell, B.; Clanet, M.; Cohen, J.A.; Filippi, M.; Fujihara, K.; Havrdova, E.; Hutchinson, M.; Kappos, L.; et al. Diagnostic criteria for multiple sclerosis: 2010 revisions to the McDonald criteria. Ann. Neurol. 2011, 69, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; Abecasis, G.R. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wan, T.; Wan, M.; Liu, B.; Cheng, R.; Zhang, R. The effect of the size of fluorescent dextran on its endocytic pathway. Cell Biol. Int. 2015, 39, 531–539. [Google Scholar] [CrossRef]
- Cui, S.; Verroust, P.J.; Moestrup, S.K.; Christensen, E.I. Megalin/gp330 mediates uptake of albumin in renal proximal tubule. Am. J. Physiol. 1996, 271, F900–F907. [Google Scholar] [CrossRef] [PubMed]
- Desdín-Micó, G.; Soto-Heredero, G.; Mittelbrunn, M. Mitochondrial activity in T cells. Mitochondrion 2018, 41, 51–57. [Google Scholar] [CrossRef]
- Abell, N.S.; DeGorter, M.K.; Gloudemans, M.J.; Greenwald, E.; Smith, K.S.; He, Z.; Montgomery, S.B. Multiple causal variants underlie genetic associations in humans. Science 2022, 375, 1247–1254. [Google Scholar] [CrossRef] [PubMed]
- Hitomi, Y.; Nakatani, K.; Kojima, K.; Nishida, N.; Kawai, Y.; Kawashima, M.; Aiba, Y.; Nagasaki, M.; Nakamura, M.; Tokunaga, K. NFKB1 and MANBA Confer Disease Susceptibility to Primary Biliary Cholangitis via Independent Putative Primary Functional Variants. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 515–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyle, A.P.; Hong, E.L.; Hariharan, M.; Cheng, Y.; Schaub, M.A.; Kasowski, M.; Karczewski, K.J.; Park, J.; Hitz, B.C.; Weng, S.; et al. Annotation of functional variation in personal genomes using RegulomeDB. Genome Res. 2012, 22, 1790–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Local, A.; Huang, H.; Albuquerque, C.P.; Singh, N.; Lee, A.Y.; Wang, W.; Wang, C.; Hsia, J.E.; Shiau, A.K.; Ge, K.; et al. Identification of H3K4me1-associated proteins at mammalian enhancers. Nat. Genet. 2018, 50, 73–82. [Google Scholar] [CrossRef]
- Rada-Iglesias, A. Is H3K4me1 at enhancers correlative or causative? Nat. Genet. 2018, 50, 4–5. [Google Scholar] [CrossRef]
- Creyghton, M.P.; Cheng, A.W.; Welstead, G.G.; Kooistra, T.; Carey, B.W.; Steine, E.J.; Hanna, J.; Lodato, M.A.; Frampton, G.M.; Sharp, P.A.; et al. Histone H3K27ac separates active from poised enhancers and predicts developmental state. Proc. Natl. Acad. Sci. USA 2010, 107, 21931–21936. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Vicente-García, C.; Seruggia, D.; Moltó, E.; Fernandez-Miñán, A.; Neto, A.; Lee, E.; Gómez-Skarmeta, J.L.; Montoliu, L.; Lunyak, V.V.; et al. MIR retrotransposon sequences provide insulators to the human genome. Proc. Natl. Acad. Sci. USA 2015, 112, E4428–E4437. [Google Scholar] [CrossRef] [Green Version]
- Ricaño-Ponce, I.; Zhernakova, D.V.; Deelen, P.; Luo, O.; Li, X.; Isaacs, A.; Karjalainen, J.; Di Tommaso, J.; Borek, Z.A.; Zorro, M.M.; et al. Refined mapping of autoimmune disease associated genetic variants with gene expression suggests an important role for non-coding RNAs. J. Autoimmun. 2016, 68, 62–74. [Google Scholar] [CrossRef] [Green Version]
- van Pelt, J.; Hokke, C.H.; Dorland, L.; Duran, M.; Kamerling, J.P.; Vliegenthart, J.F. Accumulation of mannosyl-beta(1→4)-N-acetylglucosamine in fibroblasts and leukocytes of patients with a deficiency of beta-mannosidase. Clin. Chim. Acta Int. J. Clin. Chem. 1990, 187, 55–60. [Google Scholar] [CrossRef] [Green Version]
- Fermaintt, C.S.; Sano, K.; Liu, Z.; Ishii, N.; Seino, J.; Dobbs, N.; Suzuki, T.; Fu, Y.X.; Lehrman, M.A.; Matsuo, I.; et al. A bioactive mammalian disaccharide associated with autoimmunity activates STING-TBK1-dependent immune response. Nat. Commun. 2019, 10, 2377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuin, J.; Roth, G.; Zhan, Y.; Cramard, J.; Redolfi, J.; Piskadlo, E.; Mach, P.; Kryzhanovska, M.; Tihanyi, G.; Kohler, H.; et al. Nonlinear control of transcription through enhancer-promoter interactions. Nature 2022, 604, 571–577. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP | Location | Alleles | Distance to rs7665090 (bp) | D′ | R2 | Correlated Alleles | RegulomeDB | |
|---|---|---|---|---|---|---|---|---|
| Rank | Score | |||||||
| rs7665090 | chr4:103551603 | (A/G) | 0 | 1.0 | 1.0 | A = A, G = G | 1f | 0.880 |
| rs4013 | chr4:103552813 | (C/T) | 1210 | 1.0 | 1.0 | A = C, G = T | 1f | 0.553 |
| rs735403 | chr4:103553543 | (C/T) | 1940 | 1.0 | 1.0 | A = C, G = T | 1f | 0.360 |
| rs735404 | chr4:103553665 | (G/A) | 2062 | 0.996 | 0.988 | A = G, G = A | 1f | 0.223 |
| rs2125211 | chr4:103559876 | (A/G) | 8273 | 1.0 | 1.0 | A = A, G = G | 1f | 0.223 |
| rs227361 | chr4:103586977 | (C/T) | 35,374 | 0.93 | 0.812 | A = T, G = C | 1b | 0.995 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Jiménez, A.; López-Cotarelo, P.; Agudo-Jiménez, T.; Casanova, I.; Silanes, C.L.d.; Martín-Requero, Á.; Matesanz, F.; Urcelay, E.; Espino-Paisán, L. Impact of Multiple Sclerosis Risk Polymorphism rs7665090 on MANBA Activity, Lysosomal Endocytosis, and Lymphocyte Activation. Int. J. Mol. Sci. 2022, 23, 8116. https://doi.org/10.3390/ijms23158116
González-Jiménez A, López-Cotarelo P, Agudo-Jiménez T, Casanova I, Silanes CLd, Martín-Requero Á, Matesanz F, Urcelay E, Espino-Paisán L. Impact of Multiple Sclerosis Risk Polymorphism rs7665090 on MANBA Activity, Lysosomal Endocytosis, and Lymphocyte Activation. International Journal of Molecular Sciences. 2022; 23(15):8116. https://doi.org/10.3390/ijms23158116
Chicago/Turabian StyleGonzález-Jiménez, Adela, Pilar López-Cotarelo, Teresa Agudo-Jiménez, Ignacio Casanova, Carlos López de Silanes, Ángeles Martín-Requero, Fuencisla Matesanz, Elena Urcelay, and Laura Espino-Paisán. 2022. "Impact of Multiple Sclerosis Risk Polymorphism rs7665090 on MANBA Activity, Lysosomal Endocytosis, and Lymphocyte Activation" International Journal of Molecular Sciences 23, no. 15: 8116. https://doi.org/10.3390/ijms23158116
APA StyleGonzález-Jiménez, A., López-Cotarelo, P., Agudo-Jiménez, T., Casanova, I., Silanes, C. L. d., Martín-Requero, Á., Matesanz, F., Urcelay, E., & Espino-Paisán, L. (2022). Impact of Multiple Sclerosis Risk Polymorphism rs7665090 on MANBA Activity, Lysosomal Endocytosis, and Lymphocyte Activation. International Journal of Molecular Sciences, 23(15), 8116. https://doi.org/10.3390/ijms23158116