
The Multiple Roles of CD147 in the Development and Progression of Oral Squamous Cell Carcinoma: An Overview

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. CD147 and Invasive OSCC Development
3. Reciprocal Interaction between CD147 and OPMD/OSCC-Associated Inflammation
4. CD147 and Mobile Phenotype of Oral Epithelial Cells
5. CD147 and the Growth of OSCC
6. CD147 and OSCC Resistance to Therapy
7. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AKT | protein kinase B |
| AP | activator protein |
| CAF | cancer-associated fibroblast |
| CD | cluster of differentiation |
| COX | Cyclooxygenase |
| ECM | extracellular matrix |
| EGF | epidermal growth factor |
| EGFR | epidermal growth factor receptor |
| EMT | epithelial-to-mesenchymal transition |
| ERK | extracellular-regulated kinase |
| GLUT | glucose transporter |
| HIF | Hypoxia-Inducible Factor |
| IL | Interleukin |
| MAPK | mitogen-activated protein kinase |
| MCT | monocarboxylate transporter |
| MMP | matrix metalloproteinase |
| MT-MP | membrane type-matrix metalloproteinase |
| NF-kB | Nuclear Factor-kappa B |
| OPMD | oral potentially malignant disorder |
| PI3K | phosphoinositide 3 kinase |
| SCC | squamous cell carcinoma |
| SNAI | zinc finger snail homolog |
| Sp | Specificity protein |
| TGF | transforming growth factor |
| TIMP | tissue inhibitor of matrix metalloproteinase |
| TNF | tumor necrosis factor |
| TWIST | basic helix-loop-helix twist homolog |
| uPA | urokinase-type plasminogen activator |
| uPAR | urokinase-type plasminogen activator receptor |
| VEGF | vascular endothelial growth factor |
| ZEB | zinc finger E-box-binding homeobox |
References
- Panarese, I.; Aquino, G.; Ronchi, A.; Longo, F.; Montella, M.; Cozzolino, I.; Roccuzzo, G.; Colella, G.; Caraglia, M.; Franco, R. Oral and Oropharyngeal squamous cell carcinoma: Prognostic and predictive parameters in the etiopathogenetic route. Expert Rev. Anticancer 2019, 19, 105–119. [Google Scholar] [CrossRef]
- Blatt, S.; Krüger, M.; Ziebart, T.; Sagheb, K.; Schiegnitz, E.; Goetze, E.; Al-Nawas, B.; Pabst, A.M. Biomarkers in diagnosis and therapy of oral squamous cell carcinoma: A review of the literature. J. Craniomaxillofac. Surg. 2017, 45, 722–730. [Google Scholar] [CrossRef]
- Ahmad, W.M.A.W.; Yaqoob, M.A.; Noor, N.F.M.; Ghazali, F.M.M.; Rahman, N.A.; Tang, L.; Aleng, N.A.; Alam, M.K. The Predictive Model of Oral Squamous Cell Survival Carcinoma: A Methodology of Validation. Biomed. Res. Int. 2021, 2021, 5436894. [Google Scholar] [CrossRef] [PubMed]
- Gondivkar, S.M.; Gadbail, A.R.; Sarode, S.C.; Hedaoo, A.; Dasgupta, S.; Sharma, B.; Sharma, A.; Gondivkar, R.S.; Yuwanati, M.; Patil, S.; et al. Oral and general health-related quality of life in oral squamous cell carcinoma patients- comparative analysis of different treatment regims. J. Oral Biol. Craniofac. Res. 2021, 11, 125–131. [Google Scholar] [CrossRef]
- Magnes, T.; Wagner, S.; Kiem, D.; Weiss, L.; Rinnerthaler, G.; Greil, R.; Melchardt, T. Prognostic and Predictive Factors in Advanced Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 4981. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Liu, F.; Xiao, S.; He, L.; Wu, W.; Zhao, Q. miR-29a-3p enhances the radiosensitivity of oral squamous cell carcinoma cells by inhibiting ADAM12. Eur. J. Histochem. 2021, 65, 3295. [Google Scholar] [CrossRef]
- Ma, H.; Shujaat, S.; Bila, M.; Nanhekhan, L.; Vranckx, J.; Politis, C.; Jacobs, R. Survival analysis of segmental mandibulectomy with immediate vascularized fibula flap reconstruction in stage IV oral squamous cell carcinoma patients. J. Stomatol. Oral Maxillofac. Surg. 2022, 123, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Warnakulasuriya, S. Oral potentially malignant disorders: A comprehensive review on clinical aspects and management. Oral Oncol. 2020, 102, 104550. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Hu, Y.; Zhou, X.; Liu, S.; Han, Q.; Cheng, L. Role of Oral Bacteria in the Development of Oral Squamous Cell Carcinoma. Cancers 2020, 12, 2797. [Google Scholar] [CrossRef]
- Venkatesh, A.; Elengkumaran, S.; Ravindran, C.; Malathi, N. Association of Human Papilloma Virus in Oral Squamous Cell Carcinoma: An Alarming Need for Human Papillomavirus 16 Screening in Cancer Patients. J. Pharm. Bioallied. Sci. 2021, 13, S1224–S1227. [Google Scholar] [CrossRef]
- Neville, B.W.; Day, T.A. Oral cancer and precancerous lesions. CA Cancer J. Clin. 2002, 52, 195–215. [Google Scholar] [CrossRef] [PubMed]
- Kabzinski, J.; Maczynska, M.; Majsterek, I. MicroRNA as a Novel Biomarker in the Diagnosis of Head and Neck Cancer. Biomolecules 2021, 11, 844. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Wang, K.; Chen, Y.; Zhang, Y. Identification of Differentially Expressed Long Noncoding RNAs as Functional Biomarkers and Construction of Function Enrichment Network in Oral Squamous Cell Carcinoma. Evid. Based Complement. Altern. Med. 2022, 2022, 1572249. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.C.; Dong, S.S.; Li, Z.J.; Li, C.X. Clinicopathological Significance of AKT1 and PLK1 Expression in Oral Squamous Cell Carcinoma. Dis. Markers 2022, 2022, 7300593. [Google Scholar] [CrossRef]
- Segura, I.G.; Secchi, D.G.; Galíndez, M.F.; Carrica, A.; Bologna-Molina, R.; Brunotto, M.; Centeno, V.A. Connexin 43, Bcl-2, Bax, Ki67, and E-cadherin patterns in oral squamous cell carcinoma and its relationship with GJA1 rs12197797 C/G. Med. Oral Patol. Oral Cir. Bucal. 2022, 27, e366–e374. [Google Scholar] [CrossRef]
- Vaidya, M.; Dmello, C.; Mogre, S. Utility of Keratins as Biomarkers for Human Oral Precancer and Cancer. Life 2022, 12, 343. [Google Scholar] [CrossRef]
- Vanini, J.V.; Koyama, L.K.S.; de Matos, L.L.; Junior, J.M.F.; Cernea, C.R.; Nagano, C.P.; Coutinho-Camillo, C.M.; Hsieh, R.; Lourenço, S.V. Epithelial-mesenchymal transition related to bone invasion in oral squamous cell carcinoma. J. Bone Oncol. 2022, 33, 100418. [Google Scholar] [CrossRef]
- Cao, Z.; Xiang, J.; Li, C. Expression of extracellular matrix metalloproteinase inducer and enhancement of the production of matrix metalloproteinase-1 in tongue squamous cell carcinoma. Int. J. Oral Maxillofac. Surg. 2009, 38, 880–885. [Google Scholar] [CrossRef]
- Zhang, C.; Man, D.P.; Ma, S.M.; Cao, S.W.; Li, D.W. [Expressions and significances of CD147, OPN and MMP-2 in oral squamous cell carcinoma]. Sichuan Da Xue Xue Bao Yi Xue Ban 2012, 43, 683–686. [Google Scholar]
- Berndt, A.; Richter, P.; Kosmehl, H.; Franz, M. Tenascin-C and carcinoma cell invasion in oral and urinary bladder cancer. Cell Adhes. Migr. 2015, 9, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Gautam, S.S.; Singh, R.P.; Karsauliya, K.; Sonker, A.K.; Reddy, P.J.; Mehrotra, D.; Gupta, S.; Singh, S.; Kumar, R.; Singh, S.P. Label-free plasma proteomics for the identification of the putative biomarkers of oral squamous cell carcinoma. J. Proteom. 2022, 259, 104541. [Google Scholar] [CrossRef]
- Menderico Junior, G.M.; Theodoro, T.R.; Pasini, F.S.; de Menezes Ishikawa, M.; Santos, N.S.S.; de Mello, E.S.; da Silva Pinhal, M.A.; Moyses, R.A.; Kulcsar, M.A.V.; Dedivitis, R.A.; et al. MicroRNA-mediated extracellular matrix remodeling in squamous cell carcinoma of the oral cavity. Head Neck 2021, 43, 2364–2376. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Wang, S.; Li, Z.; Hu, X.; Yang, X.; Song, Y.; Jing, Y.; Hu, Q.; Ni, Y. Identification of Metabolism-Associated Biomarkers for Early and Precise Diagnosis of Oral Squamous Cell Carcinoma. Biomolecules 2022, 12, 400. [Google Scholar] [CrossRef]
- Cho, U.; Sung, Y.E.; Kim, M.S.; Lee, Y.S. Prognostic Role of Systemic Inflammatory Markers in Patients Undergoing Surgical Resection for Oral Squamous Cell Carcinoma. Biomedicines 2022, 10, 1268. [Google Scholar] [CrossRef]
- Elmahgoub, F. Could salivary biomarkers be useful in the early detection of oral cancer and oral potentially malignant disorders, and is there a relationship between these biomarkers and risk factors? Evid. Based Dent. 2022, 23, 30–31. [Google Scholar] [CrossRef]
- Guindolet, D.; Gabison, E.E. Role of CD147 (EMMPRIN/Basigin) in Tissue Remodeling. Anat. Rec. 2020, 303, 1584–1589. [Google Scholar] [CrossRef]
- Feldman, M.; La, V.D.; Lombardo Bedran, T.B.; Palomari Spolidorio, D.M.; Grenier, D. Porphyromonas gingivalis-mediated shedding of extracellular matrix metalloproteinase inducer (EMMPRIN) by oral epithelial cells: A potential role in inflammatory periodontal disease. Microbes Infect. 2011, 13, 1261–1269. [Google Scholar] [CrossRef]
- Mauris, J.; Woodward, A.M.; Cao, Z.; Panjwani, N.; Argüeso, P. Molecular basis for MMP9 induction and disruption of epithelial cell-cell contacts by galectin-3. J. Cell Sci. 2014, 127, 3141–3148. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Xiang, J.; Li, C.; Cao, Z.; Huang, Z. Increased expression of extracellular matrix metalloproteinase inducer is associated with matrix metalloproteinase-1 and -2 in gingival tissues from patients with periodontitis. J. Periodontal Res. 2009, 44, 125–132. [Google Scholar] [CrossRef]
- Bordador, L.C.; Li, X.; Toole, B.; Chen, B.; Regezi, J.; Zardi, L.; Hu, Y.; Ramos, D.M. Expression of emmprin by oral squamous cell carcinoma. Int. J. Cancer 2000, 85, 347–352. [Google Scholar] [CrossRef]
- de Carvalho Fraga, C.A.; Farias, L.C.; de Oliveira, M.V.; Domingos, P.L.; Pereira, C.S.; Silva, T.F.; Roy, A.; Gomez, R.S.; de Paula, A.M.; Guimarães, A.L. Increased VEGFR2 and MMP9 protein levels are associated with epithelial dysplasia grading. Pathol. Res. Pract. 2014, 210, 959–964. [Google Scholar] [CrossRef]
- Bai, Y.; Huang, W.; Ma, L.T.; Jiang, J.L.; Chen, Z.N. Importance of N-glycosylation on CD147 for its biological functions. Int. J. Mol. Sci. 2014, 15, 6356–6377. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, M.; Suzuki, S.; Ishikawa, K. Cyclophilin A-EMMPRIN interaction induces invasion of head and neck squamous cell carcinoma. Oncol. Rep. 2012, 27, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Rosa, A.; Butt, E.; Hopper, C.P.; Loroch, S.; Bender, M.; Schulze, H.; Sickmann, A.; Vorlova, S.; Seizer, P.; Heinzmann, D.; et al. Cyclophilin A Is Not Acetylated at Lysine-82 and Lysine-125 in Resting and Stimulated Platelets. Int. J. Mol. Sci. 2022, 23, 1469. [Google Scholar] [CrossRef]
- Wu, Q.; Zhang, C.; He, J.; Wang, C.; Hu, X.; Li, N.; Zou, H.; Qin, J.; Yuan, M.; Wang, Y. Downregulation of caveolin-1 promotes murine breast cancer cell line progression by highly glycosylated CD147. Anticancer Drugs 2021, 32, 626–634. [Google Scholar] [CrossRef]
- Ramos, D.M.; Dang, D. EMMPRIN expression in oral SCC is regulated by FYN kinase. Anticancer Res. 2011, 31, 1205–1209. [Google Scholar]
- Wang, Q.; Xu, B.; Fan, K.; Wu, J.; Wang, T. CypB-CD147 Signaling Is Involved in Crosstalk between Cartilage and FLS in Collagen-Induced Arthritis. Mediat. Inflamm. 2020, 2020, 6473858. [Google Scholar] [CrossRef]
- Omi, Y.; Shibata, N.; Okamoto, T.; Obara, T.; Kobayashi, M. The role of CD147 in the invasiveness of follicular thyroid carcinoma cells. Thyroid 2012, 22, 383–394. [Google Scholar] [CrossRef]
- Wang, W.; Xiong, H.; Hu, Z.; Zhao, R.; Hu, Y.; Chen, W.; Han, Y.; Yang, L.; Hu, X.; Wang, C.; et al. Experimental study on TGF-β1-mediated CD147 expression in oral submucous fibrosis. Oral Dis. 2018, 24, 993–1000. [Google Scholar] [CrossRef]
- Berditchevski, F.; Chang, S.; Bodorova, J.; Hemler, M.E. Generation of monoclonal antibodies to integrin-associated proteins. Evidence that alpha3beta1 complexes with EMMPRIN/basigin/OX47/M6. J. Biol. Chem. 1997, 272, 29174–29180. [Google Scholar] [CrossRef] [Green Version]
- Richard, V.; Pillai, M.R. The stem cell code in oral epithelial tumorigenesis: ‘the cancer stem cell shift hypothesis’. Biochim. Biophys. Acta 2010, 1806, 146–162. [Google Scholar] [CrossRef]
- Luo, Z.; Dong, X.; Ke, Q.; Duan, Q.; Shen, L. Downregulation of CD147 by chitooligosaccharide inhibits MMP-2 expression and suppresses the metastatic potential of human gastric cancer. Oncol. Lett. 2014, 8, 361–366. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Toyoma, S.; Kawasaki, Y.; Nanjo, H.; Yamada, T. CD147 promotes invasion and MMP-9 expression through MEK signaling and predicts poor prognosis in hypopharyngeal squamous cell carcinoma. Adv. Clin. Exp. Med. 2021, 30, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Mitre, G.P.; Balbinot, K.M.; Ribeiro, A.L.R.; da Silva Kataoka, M.S.; de Melo Alves Júnior, S.; de Jesus Viana Pinheiro, J. Key proteins of invadopodia are overexpressed in oral squamous cell carcinoma suggesting an important role of MT1-MMP in the tumoral progression. Diagn. Pathol. 2021, 16, 33. [Google Scholar] [CrossRef]
- Jia, L.; Wei, W.; Cao, J.; Xu, H.; Miao, X.; Zhang, J. Silencing CD147 inhibits tumor progression and increases chemosensitivity in murine lymphoid neoplasm P388D1 cells. Ann. Hematol. 2009, 88, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wu, L.; Xiao, T.; Tang, L.; Jia, X.; Guo, Y.; Zhang, J.; Li, J.; He, Y.; Su, J.; et al. TRAF6 regulates EGF-induced cell transformation and cSCC malignant phenotype through CD147/EGFR. Oncogenesis 2018, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Maghsood, F.; Mirshafiey, A.; Farahani, M.M.; Modarressi, M.H.; Jafari, P.; Motevaseli, E. Dual Effects of Cell Free Supernatants from Lactobacillus acidophilus and Lactobacillus rhamnosus GG in Regulation of MMP-9 by Up-Regulating TIMP-1 and Down-Regulating CD147 in PMADifferentiated THP-1 Cells. Cell J. 2018, 19, 559–568. [Google Scholar] [CrossRef]
- Lescaille, G.; Menashi, S.; Cavelier-Balloy, B.; Khayati, F.; Quemener, C.; Podgorniak, M.P.; Naïmi, B.; Calvo, F.; Lebbe, C.; Mourah, S. EMMPRIN/CD147 up-regulates urokinase-type plasminogen activator: Implications in oral tumor progression. BMC Cancer 2012, 12, 115. [Google Scholar] [CrossRef]
- Dang, D.; Atakilit, A.; Ramos, D.M. EMMPRIN modulates migration and deposition of TN-C in oral squamous carcinoma. Anticancer Res. 2008, 28, 2049–2054. [Google Scholar]
- Yu, B.; Zhang, Y.; Wu, K.; Wang, L.; Jiang, Y.; Chen, W.; Yan, M. CD147 promotes progression of head and neck squamous cell carcinoma via NF-kappa B signaling. J. Cell Mol. Med. 2019, 23, 954–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muramatsu, T. Basigin (CD147), a multifunctional transmembrane glycoprotein with various binding partners. J. Biochem. 2016, 159, 481–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Lu, M.; Li, Y.; Shang, Y.K.; Wang, S.J.; Meng, Y.; Wang, Z.; Li, Z.S.; Chen, H.; Chen, Z.N.; et al. Regulation of a TGF-β1-CD147 self-sustaining network in the differentiation plasticity of hepatocellular carcinoma cells. Oncogene 2016, 35, 5468–5479. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Nakada, M.T.; Rafferty, P.; Laraio, J.; McCabe, F.L.; Millar, H.; Cunningham, M.; Snyder, L.A.; Bugelski, P.; Yan, L. Regulation of vascular endothelial growth factor expression by EMMPRIN via the PI3K-Akt signaling pathway. Mol. Cancer Res. 2006, 4, 371–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siu, A.; Chang, J.; Lee, C.; Lee, S.; Lee, C.; Ramos, D.M. Expression of EMMPRIN modulates mediators of tumor invasion in oral squamous cell carcinoma. J. Calif. Dent. Assoc. 2013, 41, 831–838. [Google Scholar] [PubMed]
- Almeida, L.M.C.A.; Silva, R.; Cavadas, B.; Lima, J.; Pereira, L.; Soares, P.; Sobrinho-Simões, M.; Lopes, J.M.; Máximo, V. GLUT1, MCT1/4 and CD147 overexpression supports the metabolic reprogramming in papillary renal cell carcinoma. Histol. Histopathol. 2017, 32, 1029–1040. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.; Wilson, M.C.; Heddle, C.; Brown, M.H.; Barclay, A.N.; Halestrap, A.P. CD147 is tightly associated with lactate transporters MCT1 and MCT4 and facilitates their cell surface expression. EMBO J. 2000, 19, 3896–3904. [Google Scholar] [CrossRef]
- Wang, C.H.; Yao, H.; Chen, L.N.; Jia, J.F.; Wang, L.; Dai, J.Y.; Zheng, Z.H.; Chen, Z.N.; Zhu, P. CD147 induces angiogenesis through a vascular endothelial growth factor and hypoxia-inducible transcription factor 1α-mediated pathway in rheumatoid arthritis. Arthritis Rheum. 2012, 64, 1818–1827. [Google Scholar] [CrossRef] [Green Version]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [PubMed] [Green Version]
- Itoh, Y. Membrane-Type matrix metalloproteinases: Their functions and regulations. Matrix Biol. 2015, 44, 207–223. [Google Scholar] [CrossRef]
- Li, L.; Luo, D.; Liao, Y.; Peng, K.; Zeng, Y. Mycoplasma genitalium Protein of Adhesion Induces Inflammatory Cytokines via Cyclophilin A-CD147 Activating the ERK-NF-κB Pathway in Human Urothelial Cells. Front. Immunol. 2020, 11, 2052. [Google Scholar] [CrossRef]
- Boulos, S.; Meloni, B.P.; Arthur, P.G.; Majda, B.; Bojarski, C.; Knuckey, N.W. Evidence that intracellular cyclophilin A and cyclophilin A/CD147 receptor-mediated ERK1/2 signalling can protect neurons against in vitro oxidative and ischemic injury. Neurobiol. Dis. 2007, 25, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Ding, L.; Ning, H.; Wu, C.; Fu, K.; Wang, Y.; Zhang, Y.; Liu, Y.; Zhou, L. CD147 knockdown improves the antitumor efficacy of trastuzumab in HER2-positive breast cancer cells. Oncotarget 2016, 7, 57737–57751. [Google Scholar] [CrossRef] [Green Version]
- Dana, P.; Saisomboon, S.; Kariya, R.; Okada, S.; Obchoei, S.; Sawanyawisuth, K.; Wongkham, C.; Pairojkul, C.; Wongkham, S.; Vaeteewoottacharn, K. CD147 augmented monocarboxylate transporter-1/4 expression through modulation of the Akt-FoxO3-NF-κB pathway promotes cholangiocarcinoma migration and invasion. Cell Oncol. 2020, 43, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, M.; Miyagaki, T.; Kamijo, H.; Oka, T.; Boki, H.; Takahashi-Shishido, N.; Suga, H.; Sugaya, M.; Sato, S. CD147-Cyclophilin a Interactions Promote Proliferation and Survival of Cutaneous T-Cell Lymphoma. Int. J. Mol. Sci. 2021, 22, 7889. [Google Scholar] [CrossRef] [PubMed]
- Seizer, P.; Ungern-Sternberg, S.N.; Schönberger, T.; Borst, O.; Münzer, P.; Schmidt, E.M.; Mack, A.F.; Heinzmann, D.; Chatterjee, M.; Langer, H.; et al. Extracellular cyclophilin A activates platelets via EMMPRIN (CD147) and PI3K/Akt signaling, which promotes platelet adhesion and thrombus formation in vitro and in vivo. Arter. Thromb. Vasc. Biol. 2015, 35, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.W.; Sun, X.; Shen, X.F.; Lu, Y.; Wang, J.Q.; Sun, Z.R.; Miao, C.H.; Chen, J. Propofol attenuates TNF-α-induced MMP-9 expression in human cerebral microvascular endothelial cells by inhibiting Ca2+/CAMK II/ERK/NF-κB signaling pathway. Acta Pharm. Sin. 2019, 40, 1303–1313. [Google Scholar] [CrossRef]
- Chen, Y.J.; Lee, Y.C.; Huang, C.H.; Chang, L.S. Gallic acid-capped gold nanoparticles inhibit EGF-induced MMP-9 expression through suppression of p300 stabilization and NFκB/c-Jun activation in breast cancer MDA-MB-231 cells. Toxicol. Appl. Pharm. 2016, 310, 98–107. [Google Scholar] [CrossRef]
- Lin, H.Y.; Chen, Y.S.; Wang, K.; Chien, H.W.; Hsieh, Y.H.; Yang, S.F. Fisetin inhibits epidermal growth factor-induced migration of ARPE-19 cells by suppression of AKT activation and Sp1-dependent MMP-9 expression. Mol. Vis. 2017, 23, 900–910. [Google Scholar]
- Muscella, A.; Vetrugno, C.; Cossa, L.G.; Marsigliante, S. TGF-β1 activates RSC96 Schwann cells migration and invasion through MMP-2 and MMP-9 activities. J. Neurochem. 2020, 153, 525–538. [Google Scholar] [CrossRef]
- Nazir, S.U.; Kumar, R.; Singh, A.; Khan, A.; Tanwar, P.; Tripathi, R.; Mehrotra, R.; Hussain, S. Breast cancer invasion and progression by MMP-9 through Ets-1 transcription factor. Gene 2019, 711, 143952. [Google Scholar] [CrossRef]
- Yang, C.C.; Hsiao, L.D.; Yang, C.M. Galangin Inhibits LPS-Induced MMP-9 Expression via Suppressing Protein Kinase-Dependent AP-1 and FoxO1 Activation in Rat Brain Astrocytes. J. Inflamm. Res. 2020, 13, 945–960. [Google Scholar] [CrossRef]
- Huet, E.; Gabison, E.; Vallee, B.; Mougenot, N.; Linguet, G.; Riou, B.; Jarosz, C.; Menashi, S.; Besse, S. Deletion of extracellular matrix metalloproteinase inducer/CD147 induces altered cardiac extracellular matrix remodeling in aging mice. J. Physiol. Pharm. 2015, 66, 355–366. [Google Scholar]
- Lee, C.L.; Lam, M.P.; Lam, K.K.; Leung, C.O.; Pang, R.T.; Chu, I.K.; Wan, T.H.; Chai, J.; Yeung, W.S.; Chiu, P.C. Identification of CD147 (basigin) as a mediator of trophoblast functions. Hum. Reprod. 2013, 28, 2920–2929. [Google Scholar] [CrossRef] [Green Version]
- Lottin, M.; Soudet, S.; Fercot, J.; Racine, F.; Demagny, J.; Bettoni, J.; Chatelain, D.; Sevestre, M.A.; Mammeri, Y.; Lamuraglia, M.; et al. Molecular Landscape of the Coagulome of Oral Squamous Cell Carcinoma. Cancers 2022, 14, 460. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Moon, A. A curcumin analog CA-5f inhibits urokinase-type plasminogen activator and invasive phenotype of triple-negative breast cancer cells. Toxicol. Res. 2021, 38, 19–26. [Google Scholar] [CrossRef]
- Ismail, A.A.; Shaker, B.T.; Bajou, K. The Plasminogen-Activator Plasmin System in Physiological and Pathophysiological Angiogenesis. Int. J. Mol. Sci. 2021, 23, 337. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Jiang, Y.W.; Kuo, C.L.; Way, T.D.; Chou, Y.C.; Chang, Y.S.; Chung, J.G. Chrysin inhibit human melanoma A375.S2 cell migration and invasion via affecting MAPK signaling and NF-κB signaling pathway in vitro. Environ. Toxicol. 2019, 34, 434–442. [Google Scholar] [CrossRef]
- Amos, S.; Redpath, G.T.; Dipierro, C.G.; Carpenter, J.E.; Hussaini, I.M. Epidermal growth factor receptor-mediated regulation of urokinase plasminogen activator expression and glioblastoma invasion via C-SRC/MAPK/AP-1 signaling pathways. J. Neuropathol. Exp. Neurol. 2010, 69, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.H.; Lu, K.H.; Yang, J.S.; Hsieh, Y.H.; Lin, C.W.; Yang, S.F. Dihydromyricetin suppresses cell metastasis in human osteosarcoma through SP-1- and NF-κB-modulated urokinase plasminogen activator inhibition. Phytomedicine 2021, 90, 153642. [Google Scholar] [CrossRef] [PubMed]
- Watabe, T.; Yoshida, K.; Shindoh, M.; Kaya, M.; Fujikawa, K.; Sato, H.; Seiki, M.; Ishii, S.; Fujinaga, K. The Ets-1 and Ets-2 transcription factors activate the promoters for invasion-associated urokinase and collagenase genes in response to epidermal growth factor. Int. J. Cancer 1998, 77, 128–137. [Google Scholar] [CrossRef]
- Huang, C.; Sun, Z.; Sun, Y.; Chen, X.; Zhu, X.; Fan, C.; Liu, B.; Zhao, Y.; Zhang, W. Association of increased ligand cyclophilin A and receptor CD147 with hypoxia, angiogenesis, metastasis and prognosis of tongue squamous cell carcinoma. Histopathology 2012, 60, 793–803. [Google Scholar] [CrossRef]
- Pătru, A.; Şurlin, V.; Mărgăritescu, C.; Ciucă, E.M.; Matei, M.; Dumitrescu, D.; Camen, A. Immunohistochemical evaluation of D2-40, Galectin-3, Maspin and MCM7 expression in palate squamous cell carcinomas. Rom. J. Morphol. Embryol. 2021, 62, 133–149. [Google Scholar] [CrossRef] [PubMed]
- Radulescu, R.; Totan, A.R.; Imre, M.M.; Miricescu, D.; Didilescu, A.; Greabu, M. Mediators of extracellular matrix degradation and inflammation: A new team of possible biomarkers for oral squamous cell carcinoma stage. Exp. Ther. Med. 2021, 22, 877. [Google Scholar] [CrossRef]
- de Vicente, J.C.; Lequerica-Fernández, P.; Santamaría, J.; Fresno, M.F. Expression of MMP-7 and MT1-MMP in oral squamous cell carcinoma as predictive indicator for tumor invasion and prognosis. J. Oral Pathol. Med. 2007, 36, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Lawal, A.O.; Adisa, A.O.; Kolude, B.; Adeyemi, B.F. Immunohistochemical expression of MMP-2 and MMP-8 in oral squamous cell carcinoma. J. Clin. Exp. Dent. 2015, 7, e203–e207. [Google Scholar] [CrossRef] [PubMed]
- Miguel, A.F.P.; Mello, F.W.; Melo, G.; Rivero, E.R.C. Association between immunohistochemical expression of matrix metalloproteinases and metastasis in oral squamous cell carcinoma: Systematic review and meta-analysis. Head Neck 2020, 42, 569–584. [Google Scholar] [CrossRef] [PubMed]
- Patil, R.; Mahajan, A.; Pradeep, G.L.; Prakash, N.; Patil, S.; Khan, S.M. Expression of matrix metalloproteinase-9 in histological grades of oral squamous cell carcinoma: An immunohistochemical study. J. Oral Maxillofac. Pathol. 2021, 25, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Soares, A.B.; Eymael, D.; Magalhaes, M. Expression of invadopodia markers can identify oral lesions with a high risk of malignant transformation. J. Pathol. Clin. Res. 2021, 7, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Grass, G.D.; Tolliver, L.B.; Bratoeva, M.; Toole, B.P. CD147, CD44, and the epidermal growth factor receptor (EGFR) signaling pathway cooperate to regulate breast epithelial cell invasiveness. J. Biol. Chem. 2013, 288, 26089–26104. [Google Scholar] [CrossRef] [Green Version]
- Hwang, Y.S.; Park, K.K.; Chung, W.Y. Invadopodia formation in oral squamous cell carcinoma: The role of epidermal growth factor receptor signalling. Arch. Oral Biol. 2012, 57, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Yamahana, H.; Terashima, M.; Takatsuka, R.; Asada, C.; Suzuki, T.; Uto, Y.; Takino, T. TGF-β1 facilitates MT1-MMP-mediated proMMP-9 activation and invasion in oral squamous cell carcinoma cells. Biochem. Biophys. Rep. 2021, 27, 101072. [Google Scholar] [CrossRef]
- Ghosh, S.; Koblinski, J.; Johnson, J.; Liu, Y.; Ericsson, A.; Davis, J.W.; Shi, Z.; Ravosa, M.J.; Crawford, S.; Frazier, S.; et al. Urinary-type plasminogen activator receptor/alpha 3 beta 1 integrin signaling, altered gene expression, and oral tumor progression. Mol. Cancer Res. 2010, 8, 145–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinohara, M.; Nakamura, S.; Sasaki, M.; Kurahara, S.; Ikebe, T.; Harada, T.; Shirasuna, K. Expression of integrins in squamous cell carcinoma of the oral cavity. Correlations with tumor invasion and metastasis. Am. J. Clin. Pathol. 1999, 111, 75–88. [Google Scholar] [CrossRef] [Green Version]
- Düzlü, M.; Karamert, R.; Tutar, H.; Şahin, M.; Türkcan, A.; Yılmaz, M. Diagnostic role of neutrophil-lymphocyte ratio in oral cavity cancers. Niger. J. Clin. Pract. 2018, 21, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Schernberg, A.; Canova, C.; Blanchard, P.; Gorphe, P.; Breuskin, I.; Mirghani, H.; Moya-Plana, A.; Janot, F.; Bidault, F.; Chargari, C.; et al. Prognostic factors in patients with soft palate squamous cell carcinoma. Head Neck 2019, 41, 1441–1449. [Google Scholar] [CrossRef] [PubMed]
- Melaiu, O.; Lucarini, V.; Cifaldi, L.; Fruci, D. Influence of the Tumor Microenvironment on NK Cell Function in Solid Tumors. Front. Immunol. 2020, 10, 3038. [Google Scholar] [CrossRef] [PubMed]
- Lucarini, V.; Melaiu, O.; Tempora, P.; D’Amico, S.; Locatelli, F.; Fruci, D. Dendritic Cells: Behind the Scenes of T-Cell Infiltration into the Tumor Microenvironment. Cancers 2021, 13, 433. [Google Scholar] [CrossRef] [PubMed]
- Weiße, J.; Rosemann, J.; Krauspe, V.; Kappler, M.; Eckert, A.W.; Haemmerle, M.; Gutschner, T. RNA-Binding Proteins as Regulators of Migration, Invasion and Metastasis in Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2020, 21, 6835. [Google Scholar] [CrossRef] [PubMed]
- Kamarajan, P.; Alhazzazi, T.Y.; Danciu, T.; D’silva, N.J.; Verdin, E.; Kapila, Y.L. Receptor-interacting protein (RIP) and Sirtuin-3 (SIRT3) are on opposite sides of anoikis and tumorigenesis. Cancer 2012, 118, 5800–5810. [Google Scholar] [CrossRef]
- Lu, Y.; Lin, J.; Duan, M.; Rui, Y.; Zheng, H.; Zhu, L.; Zhu, X.; Wei, J. Anlotinib Suppresses Oral Squamous Cell Carcinoma Growth and Metastasis by Targeting the RAS Protein to Inhibit the PI3K/Akt Signalling Pathway. Anal. Cell Pathol. 2021, 2021, 5228713. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, J. Icariin inhibits oral squamous cell carcinoma cell proliferation and induces apoptosis via inhibiting the NF-κB and PI3K/AKT pathways. Exp. Ther. Med. 2021, 22, 942. [Google Scholar] [CrossRef]
- Jiang, L.; Xiao, J. 2-phenylethynesulfonamide inhibits growth of oral squamous cell carcinoma cells by blocking the function of heat shock protein 70. Biosci. Rep. 2020, 40, BSR20200079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Shen, X.; Tian, J. The effects of periodontitis associated microbiota on the development of oral squamous cell carcinoma. Biochem. Biophys. Res. Commun. 2021, 576, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Shen, X.; Zhou, M.; Tang, B. Periodontal Pathogens Promote Oral Squamous Cell Carcinoma by Regulating ATR and NLRP3 Inflammasome. Front. Oncol. 2021, 11, 722797. [Google Scholar] [CrossRef]
- Miguel, A.F.P.; Embaló, B.; Alves Dias, H.B.; Rivero, E.R.C. Immunohistochemical Expression of MMP-9, TIMP-1, and Vimentin and its Correlation With Inflammatory Reaction and Clinical Parameters in Oral Epithelial Dysplasia. Appl. Immunohistochem. Mol. Morphol. 2021, 29, 382–389. [Google Scholar] [CrossRef]
- Gegunde, S.; Alfonso, A.; Alvariño, R.; Alonso, E.; Botana, L.M. Cyclophilins A, B, and C Role in Human T Lymphocytes Upon Inflammatory Conditions. Front. Immunol. 2021, 12, 609196. [Google Scholar] [CrossRef]
- Wang, C.H.; Dai, J.Y.; Wang, L.; Jia, J.F.; Zheng, Z.H.; Ding, J.; Chen, Z.N.; Zhu, P. Expression of CD147 (EMMPRIN) on neutrophils in rheumatoid arthritis enhances chemotaxis, matrix metalloproteinase production and invasiveness of synoviocytes. J. Cell Mol. Med. 2011, 15, 850–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babiuch, K.; Kuśnierz-Cabala, B.; Kęsek, B.; Okoń, K.; Darczuk, D.; Chomyszyn-Gajewska, M. Evaluation of Proinflammatory, NF-kappaB Dependent Cytokines: IL-1α, IL-6, IL-8, and TNF-α in Tissue Specimens and Saliva of Patients with Oral Squamous Cell Carcinoma and Oral Potentially Malignant Disorders. J. Clin. Med. 2020, 9, 867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.C.; Chen, C.C.; Lin, C.C.; Chen, C.H.; Lin, T.S.; Shieh, T.Y. Interleukin-1 beta in oral submucous fibrosis, verrucous hyperplasia and squamous cell carcinoma tissues. Kaohsiung J. Med. Sci. 1999, 15, 513–519. [Google Scholar]
- Chadwick, J.W.; Macdonald, R.; Ali, A.A.; Glogauer, M.; Magalhaes, M.A. TNFα Signaling Is Increased in Progressing Oral Potentially Malignant Disorders and Regulates Malignant Transformation in an Oral Carcinogenesis Model. Front. Oncol. 2021, 11, 741013. [Google Scholar] [CrossRef]
- Oo, M.W.; Kawai, H.; Takabatake, K.; Shan, Q.; Eain, H.S.; Sukegawa, S.; Nakano, K.; Nagatsuka, H. Cancer-Associated Stromal Cells Promote the Contribution of MMP2-Positive Bone Marrow-Derived Cells to Oral Squamous Cell Carcinoma Invasion. Cancers 2021, 14, 137. [Google Scholar] [CrossRef]
- Antunes, D.M.; Rodrigues, M.F.S.D.; Guimarães, D.M.; Duarte, C.M.E.; Miguita, L.; Corrêa, L.; DEOliveira, A.P.L.; Fernandes, K.P.S.; Nunes, F.D. Nonsteroidal Anti-inflammatory Drugs Modulate Gene Expression of Inflammatory Mediators in Oral Squamous Cell Carcinoma. Anticancer Res. 2019, 39, 2385–2394. [Google Scholar] [CrossRef]
- Bae, J.Y.; Kim, E.K.; Yang, D.H.; Zhang, X.; Park, Y.J.; Lee, D.Y.; Che, C.M.; Kim, J. Reciprocal interaction between carcinoma-associated fibroblasts and squamous carcinoma cells through interleukin-1α induces cancer progression. Neoplasia 2014, 16, 928–938. [Google Scholar] [CrossRef] [Green Version]
- Iulia Irimie, A.; Braicu, C.; Zanoaga, O.; Pileczki, V.; Soritau, O.; Berindan-Neagoe, I.; Septimiu Campian, R. Inhibition of tumor necrosis factor alpha using RNA interference in oral squamous cell carcinoma. J. BUON 2015, 20, 1107–1114. [Google Scholar]
- Rao, S.K.; Pavicevic, Z.; Du, Z.; Kim, J.G.; Fan, M.; Jiao, Y.; Rosebush, M.; Samant, S.; Gu, W.; Pfeffer, L.M.; et al. Pro-inflammatory genes as biomarkers and therapeutic targets in oral squamous cell carcinoma. J. Biol. Chem. 2010, 285, 32512–32521. [Google Scholar] [CrossRef] [Green Version]
- Luger, T.A. Epidermal cytokines. Acta Derm. Venereol. Suppl. 1989, 151, 61–76, discussion 106–110. [Google Scholar] [CrossRef]
- Formanek, M.; Knerer, B.; Temmel, A.; Thurnher, D.; Millesi, W.; Kornfehl, J. Oral keratinocytes derived from the peritonsillar mucosa express the proinflammatory cytokine IL-6 without prior stimulation. J. Oral Pathol. Med. 1998, 27, 202–206. [Google Scholar] [CrossRef]
- Ferrari, E.; Pezzi, M.E.; Cassi, D.; Pertinhez, T.A.; Spisni, A.; Meleti, M. Salivary Cytokines as Biomarkers for Oral Squamous Cell Carcinoma: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 6795. [Google Scholar] [CrossRef]
- Rhodus, N.L.; Ho, V.; Miller, C.S.; Myers, S.; Ondrey, F. NF-kappaB dependent cytokine levels in saliva of patients with oral preneoplastic lesions and oral squamous cell carcinoma. Cancer Detect. Prev. 2005, 29, 42–45. [Google Scholar] [CrossRef]
- Dana, P.; Kariya, R.; Lert-Itthiporn, W.; Seubwai, W.; Saisomboon, S.; Wongkham, C.; Okada, S.; Wongkham, S.; Vaeteewoottacharn, K. Homophilic Interaction of CD147 Promotes IL-6-Mediated Cholangiocarcinoma Invasion via the NF-κB-Dependent Pathway. Int. J. Mol. Sci. 2021, 22, 13496. [Google Scholar] [CrossRef]
- Schmidt, R.; Bültmann, A.; Fischel, S.; Gillitzer, A.; Cullen, P.; Walch, A.; Jost, P.; Ungerer, M.; Tolley, N.D.; Lindemann, S.; et al. Extracellular matrix metalloproteinase inducer (CD147) is a novel receptor on platelets, activates platelets, and augments nuclear factor kappaB-dependent inflammation in monocytes. Circ. Res. 2008, 102, 302–309. [Google Scholar] [CrossRef]
- Saputra, W.D.; Shono, H.; Ohsaki, Y.; Sultana, H.; Komai, M.; Shirakawa, H. Geranylgeraniol Inhibits Lipopolysaccharide-Induced Inflammation in Mouse-Derived MG6 Microglial Cells via NF-κB Signaling Modulation. Int. J. Mol. Sci. 2021, 22, 10543. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.A.; Shin, J.Y.; Hong, S.S.; Cho, Y.R.; Park, J.H.; Seo, D.W.; Oh, J.S.; Kang, J.S.; Lee, J.H.; Ahn, E.K. Tetracera loureiri Extract Regulates Lipopolysaccharide-Induced Inflammatory Response Via Nuclear Factor-κB and Mitogen Activated Protein Kinase Signaling Pathways. Plants 2022, 11, 284. [Google Scholar] [CrossRef] [PubMed]
- Yoon, W.J.; Lee, N.H.; Hyun, C.G. Limonene suppresses lipopolysaccharide-induced production of nitric oxide, prostaglandin E2, and pro-inflammatory cytokines in RAW 264.7 macrophages. J. Oleo Sci. 2010, 59, 415–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhao, C.; Meng, J.; Li, N.; Xu, Z.; Liu, X.; Hou, S. Galectin-3 regulates microglial activation and promotes inflammation through TLR4/MyD88/NF-kB in experimental autoimmune uveitis. Clin. Immunol. 2022, 236, 108939. [Google Scholar] [CrossRef] [PubMed]
- Chou, W.C.; Tsai, K.L.; Hsieh, P.L.; Wu, C.H.; Jou, I.M.; Tu, Y.K.; Ma, C.H. Galectin-3 facilitates inflammation and apoptosis in chondrocytes through upregulation of the TLR-4-mediated oxidative stress pathway in TC28a2 human chondrocyte cells. Environ. Toxicol. 2022, 37, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, W. Resveratrol Alleviates Ischemic Brain Injury by Inhibiting the Activation of Pro-Inflammatory Microglia Via the CD147/MMP-9 Pathway. J. Stroke Cerebrovasc. Dis. 2022, 31, 106307. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Yuan, X.; Lv, X.; Liu, Q.; Tao, L.; Meng, J. Caveolin-1 negatively regulates inflammation and fibrosis in silicosis. J. Cell Mol. Med. 2022, 26, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Abdulkareem, A.A.; Shelton, R.M.; Landini, G.; Cooper, P.R.; Milward, M.R. Periodontal pathogens promote epithelial-mesenchymal transition in oral squamous carcinoma cells in vitro. Cell Adhes. Migr. 2018, 12, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.W.; Zhou, J.P.; Bi, Y.L.; Wang, J.Y.; Yu, R.; Deng, C.; Wang, W.K.; Li, X.Z.; Huang, R.; Zhang, J.; et al. The role of MAPK signaling pathway in formation of EMT in oral squamous carcinoma cells induced by TNF-α. Mol. Biol. Rep. 2019, 46, 3149–3156. [Google Scholar] [CrossRef]
- Zhou, J.P.; Gao, Z.L.; Zhou, M.L.; He, M.Y.; Xu, X.H.; Tao, D.T.; Yang, C.C.; Liu, L.K. Snail interacts with Id2 in the regulation of TNF-α-induced cancer cell invasion and migration in OSCC. Am. J. Cancer Res. 2015, 5, 1680–1691. [Google Scholar]
- Angelini, D.J.; Hyun, S.W.; Grigoryev, D.N.; Garg, P.; Gong, P.; Singh, I.S.; Passaniti, A.; Hasday, J.D.; Goldblum, S.E. TNF-alpha increases tyrosine phosphorylation of vascular endothelial cadherin and opens the paracellular pathway through fyn activation in human lung endothelia. Am. J. Physiol. Lung Cell Mol. Physiol. 2006, 291, L1232–L1245. [Google Scholar] [CrossRef] [PubMed]
- Hallek, M.; Neumann, C.; Schäffer, M.; Danhauser-Riedl, S.; von Bubnoff, N.; de Vos, G.; Druker, B.J.; Yasukawa, K.; Griffin, J.D.; Emmerich, B. Signal transduction of interleukin-6 involves tyrosine phosphorylation of multiple cytosolic proteins and activation of Src-family kinases Fyn, Hck, and Lyn in multiple myeloma cell lines. Exp. Hematol. 1997, 25, 1367–1377. [Google Scholar] [PubMed]
- Razani-Boroujerdi, S.; Langley, R.J.; Singh, S.P.; Pena-Philippides, J.C.; Rir-sima-ah, J.; Gundavarapu, S.; Mishra, N.C.; Sopori, M.L. The role of IL-1β in nicotine-induced immunosuppression and neuroimmune communication. J. Neuroimmune Pharm. 2011, 6, 585–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Yang, X.; Zhang, H.; Liu, X.; Pan, S.; Li, C. The role of extracellular matrix metalloproteinase inducer glycosylation in regulating matrix metalloproteinases in periodontitis. J. Periodontal Res. 2018, 53, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, X.; Li, C.; He, C.; Ren, B.; Deng, Q.; Gao, W.; Wang, B. Aurora-A modulates MMP-2 expression via AKT/NF-κB pathway in esophageal squamous cell carcinoma cells. Acta Biochim. Biophys. Sin. 2016, 48, 520–527. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.Y.; Hong, O.Y.; Youn, H.J.; Kim, M.G.; Kim, C.H.; Jung, S.H.; Kim, J.S. 15d-PGJ2 inhibits NF-κB and AP-1-mediated MMP-9 expression and invasion of breast cancer cell by means of a heme oxygenase-1-dependent mechanism. BMB Rep. 2020, 53, 212–217. [Google Scholar] [CrossRef]
- Petruzzi, M.N.; Cherubini, K.; Salum, F.G.; de Figueiredo, M.A. Role of tumour-associated macrophages in oral squamous cells carcinoma progression: An update on current knowledge. Diagn. Pathol. 2017, 12, 32. [Google Scholar] [CrossRef] [Green Version]
- Chanmee, T.; Ontong, P.; Konno, K.; Itano, N. Tumor-associated macrophages as major players in the tumor microenvironment. Cancers 2014, 6, 1670–1690. [Google Scholar] [CrossRef] [Green Version]
- Haque, A.S.M.R.; Moriyama, M.; Kubota, K.; Ishiguro, N.; Sakamoto, M.; Chinju, A.; Mochizuki, K.; Sakamoto, T.; Kaneko, N.; Munemura, R.; et al. CD206+ tumor-associated macrophages promote proliferation and invasion in oral squamous cell carcinoma via EGF production. Sci. Rep. 2019, 9, 14611. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Guinea, M.C.; Slomiany, M.G.; Bratoeva, M.; Grass, G.D.; Tolliver, L.B.; Maria, B.L.; Toole, B.P. CD147-dependent heterogeneity in malignant and chemoresistant properties of cancer cells. Am. J. Pathol. 2013, 182, 577–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, I.; Higa-Nakamine, S.; Uehara, A.; Sugahara, K.; Kakinohana, M.; Yamamoto, H. Regulation of epidermal growth factor expression and morphology of lung epithelial cells by interleukin-1β. J. Biochem. 2020, 168, 113–123. [Google Scholar] [CrossRef]
- Schmiegel, W.; Roeder, C.; Schmielau, J.; Rodeck, U.; Kalthoff, H. Tumor necrosis factor alpha induces the expression of transforming growth factor alpha and the epidermal growth factor receptor in human pancreatic cancer cells. Proc. Natl. Acad. Sci. USA 1993, 90, 863–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schraufstatter, I.U.; Trieu, K.; Zhao, M.; Rose, D.M.; Terkeltaub, R.A.; Burger, M. IL-8-mediated cell migration in endothelial cells depends on cathepsin B activity and transactivation of the epidermal growth factor receptor. J. Immunol. 2003, 171, 6714–6722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wehbe, H.; Henson, R.; Meng, F.; Mize-Berge, J.; Patel, T. Interleukin-6 contributes to growth in cholangiocarcinoma cells by aberrant promoter methylation and gene expression. Cancer Res. 2006, 66, 10517–10524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Diamond, M.E.; Ottaviano, A.J.; Joseph, M.J.; Ananthanarayan, V.; Munshi, H.G. Transforming growth factor-beta 1 promotes matrix metalloproteinase-9-mediated oral cancer invasion through snail expression. Mol. Cancer Res. 2008, 6, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Costea, D.E.; Hills, A.; Osman, A.H.; Thurlow, J.; Kalna, G.; Huang, X.; Pena Murillo, C.; Parajuli, H.; Suliman, S.; Kulasekara, K.K.; et al. Identification of two distinct carcinoma-associated fibroblast subtypes with differential tumor-promoting abilities in oral squamous cell carcinoma. Cancer Res. 2013, 73, 3888–3901. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Xu, H. Regulation of transforming growth factor-beta1 by circANKS1B/miR-515-5p affects the metastatic potential and cisplatin resistance in oral squamous cell carcinoma. Bioengineered 2021, 12, 12420–12430. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Tang, Q.; Chen, X.; Fan, W.; Zhou, Z.; Huang, W.; Liang, F. TGF-β1 and IL-17A comediate the protumor phenotype of neutrophils to regulate the epithelial-mesenchymal transition in oral squamous cell carcinoma. J. Oral Pathol. Med. 2021, 50, 353–361. [Google Scholar] [CrossRef]
- Thomas, G.J.; Hart, I.R.; Speight, P.M.; Marshall, J.F. Binding of TGF-beta1 latency-associated peptide (LAP) to alpha(v)beta6 integrin modulates behaviour of squamous carcinoma cells. Br. J. Cancer 2002, 87, 859–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Y.H.; Feng, Q.Y.; Liu, Q.; Li, H.Y.; Song, X.L.; Hu, Z.X.; Xu, Z.Y.; Li, J.H.; Li, M.J.; Zheng, W.L.; et al. Posttranscriptional regulation of MMP-9 by HuR contributes to IL-1β-induced pterygium fibroblast migration and invasion. J. Cell Physiol. 2020, 235, 5130–5140. [Google Scholar] [CrossRef] [PubMed]
- Gabasa, M.; Arshakyan, M.; Llorente, A.; Chuliá-Peris, L.; Pavelescu, I.; Xaubet, A.; Pereda, J.; Alcaraz, J. Interleukin-1β Modulation of the Mechanobiology of Primary Human Pulmonary Fibroblasts: Potential Implications in Lung Repair. Int. J. Mol. Sci. 2020, 21, 8417. [Google Scholar] [CrossRef]
- Li, Y.; Su, G.; Zhong, Y.; Xiong, Z.; Huang, T.; Quan, J.; Huang, J.; Wen, X.; Luo, C.; Zheng, W.; et al. HB-EGF-induced IL-8 secretion from airway epithelium leads to lung fibroblast proliferation and migration. BMC Pulm. Med. 2021, 21, 347. [Google Scholar] [CrossRef] [PubMed]
- Nishikai-Yan Shen, T.; Kanazawa, S.; Kado, M.; Okada, K.; Luo, L.; Hayashi, A.; Mizuno, H.; Tanaka, R. Interleukin-6 stimulates Akt and p38 MAPK phosphorylation and fibroblast migration in non-diabetic but not diabetic mice. PLoS ONE 2017, 12, e0178232. [Google Scholar] [CrossRef]
- Oryan, A.; Alemzadeh, E.; Eskandari, M.H. Kefir Accelerates Burn Wound Healing Through Inducing Fibroblast Cell Migration In Vitro and Modulating the Expression of IL-1ß, TGF-ß1, and bFGF Genes In Vivo. Probiotics Antimicrob Proteins 2019, 11, 874–886. [Google Scholar] [CrossRef]
- Porter, K.E.; Turner, N.A. Cardiac fibroblasts: At the heart of myocardial remodeling. Pharmacol. Ther. 2009, 123, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Saini, S.; Liu, T.; Yoo, J. TNF-α stimulates colonic myofibroblast migration via COX-2 and Hsp27. J. Surg. Res. 2016, 204, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Bouchard, G.; Garcia-Marques, F.J.; Karacosta, L.G.; Zhang, W.; Bermudez, A.; Riley, N.M.; Varma, S.; Mehl, L.C.; Benson, J.A.; Shrager, J.B.; et al. Multiomics Analysis of Spatially Distinct Stromal Cells Reveals Tumor-Induced O-Glycosylation of the CDK4-pRB Axis in Fibroblasts at the Invasive Tumor Edge. Cancer Res. 2022, 82, 648–664. [Google Scholar] [CrossRef] [PubMed]
- Hata, S.; Okamura, K.; Hatta, M.; Ishikawa, H.; Yamazaki, J. Proteolytic and non-proteolytic activation of keratinocyte-derived latent TGF-β1 induces fibroblast differentiation in a wound-healing model using rat skin. J. Pharmacol. Sci. 2014, 124, 230–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garlick, D.S.; Li, J.; Sansoucy, B.; Wang, T.; Griffith, L.; Fitzgerald, T.; Butterfield, J.; Charbonneau, B.; Violette, S.M.; Weinreb, P.H.; et al. α(V)β(6) integrin expression is induced in the POET and Pten(pc-/-) mouse models of prostatic inflammation and prostatic adenocarcinoma. Am. J. Transl. Res. 2012, 4, 165–174. [Google Scholar] [PubMed]
- Paterson, I.C.; Matthews, J.B.; Huntley, S.; Robinson, C.M.; Fahey, M.; Parkinson, E.K.; Prime, S.S. Decreased expression of TGF-beta cell surface receptors during progression of human oral squamous cell carcinoma. J. Pathol. 2001, 193, 458–467. [Google Scholar] [CrossRef]
- Räsänen, K.; Vaheri, A. TGF-beta1 causes epithelial-mesenchymal transition in HaCaT derivatives, but induces expression of COX-2 and migration only in benign, not in malignant keratinocytes. J. Dermatol. Sci. 2010, 58, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Woo, K.M.; Lee, G.; Kook, J.K.; Min, B.M. Conversion of normal human oral keratinocytes to tumorigenic cells is associated with the acquisition of resistance to TGF-beta. Int. J. Oncol. 1998, 12, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Chung, T.W.; Choi, H.J.; Kwak, C.H.; Song, K.H.; Suh, S.J.; Kwon, K.M.; Chang, Y.C.; Park, Y.G.; Chang, H.W.; et al. Ganglioside GM3 participates in the TGF-β1-induced epithelial-mesenchymal transition of human lens epithelial cells. Biochem. J. 2013, 449, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Fang, S.; Liu, Q.; Gao, J.; Wang, X.; Zhu, H.; Zhu, Z.; Ji, F.; Wu, J.; Ma, Y.; et al. TGF-β1/p65/MAT2A pathway regulates liver fibrogenesis via intracellular SAM. EBioMedicine 2019, 42, 458–469. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Yin, L.; Cai, Y.; Hu, Q.; Huang, J.; Ji, Q.; Hu, Y.; Huang, W.; Liu, F.; Shi, S.; et al. Epigenetic regulation of integrin β6 transcription induced by TGF-β1 in human oral squamous cell carcinoma cells. J. Cell Biochem. 2018, 119, 4193–4204. [Google Scholar] [CrossRef]
- Ly, T.D.; Kleine, A.; Plümers, R.; Fischer, B.; Schmidt, V.; Hendig, D.; Distler, J.H.W.; Kuhn, J.; Knabbe, C.; Faust, I. Cytokine-mediated induction of human xylosyltransferase-I in systemic sclerosis skin fibroblasts. Biochem. Biophys. Res. Commun. 2021, 549, 34–39. [Google Scholar] [CrossRef]
- Shi, Y.; Tao, M.; Ni, J.; Tang, L.; Liu, F.; Chen, H.; Ma, X.; Hu, Y.; Zhou, X.; Qiu, A.; et al. Requirement of Histone Deacetylase 6 for Interleukin-6 Induced Epithelial-Mesenchymal Transition, Proliferation, and Migration of Peritoneal Mesothelial Cells. Front. Pharm. 2021, 12, 722638. [Google Scholar] [CrossRef] [PubMed]
- Cabral-Dias, R.; Lucarelli, S.; Zak, K.; Rahmani, S.; Judge, G.; Abousawan, J.; DiGiovanni, L.F.; Vural, D.; Anderson, K.E.; Sugiyama, M.G.; et al. Fyn and TOM1L1 are recruited to clathrin-coated pits and regulate Akt signaling. J. Cell Biol. 2022, 221, e201808181. [Google Scholar] [CrossRef]
- Malik, A.; Khatri, R.; Gupta, S.K. Interdependence of JAK-STAT and MAPK signaling pathways during EGF-mediated HTR-8/SVneo cell invasion. PLoS ONE 2017, 12, e0178269. [Google Scholar] [CrossRef]
- Zhou, M.Y.; Cheng, M.L.; Huang, T.; Hu, R.H.; Zou, G.L.; Li, H.; Zhang, B.F.; Zhu, J.J.; Liu, Y.M.; Liu, Y.; et al. Transforming growth factor beta-1 upregulates glucose transporter 1 and glycolysis through canonical and noncanonical pathways in hepatic stellate cells. World J. Gastroenterol. 2021, 27, 6908–6926. [Google Scholar] [CrossRef] [PubMed]
- Niture, S.K.; Khatri, R.; Jaiswal, A.K. Regulation of Nrf2-an update. Free Radic. Biol. Med. 2014, 66, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Li, H.Y.; Ju, D.; Zhang, D.W.; Li, H.; Kong, L.M.; Guo, Y.; Li, C.; Wang, X.L.; Chen, Z.N.; Bian, H. Activation of TGF-β1-CD147 positive feedback loop in hepatic stellate cells promotes liver fibrosis. Sci. Rep. 2015, 5, 16552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins-Port, C.E.; Higgins, P.J. Regulation of extracellular matrix remodeling following transforming growth factor-beta1/epidermal growth factor-stimulated epithelial-mesenchymal transition in human premalignant keratinocytes. Cells Tissues Organs 2007, 185, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Janecka-Widła, A.; Majchrzyk, K.; Mucha-Małecka, A.; Słonina, D.; Biesaga, B. Prognostic potential of Akt, pAkt(Ser473) and pAkt(Thr308) immunoreactivity in relation to HPV prevalence in head and neck squamous cell carcinoma patients. Pathol. Res. Pract. 2022, 229, 153684. [Google Scholar] [CrossRef] [PubMed]
- Massarelli, E.; Liu, D.D.; Lee, J.J.; El-Naggar, A.K.; Lo Muzio, L.; Staibano, S.; De Placido, S.; Myers, J.N.; Papadimitrakopoulou, V.A. Akt activation correlates with adverse outcome in tongue cancer. Cancer 2005, 104, 2430–2436. [Google Scholar] [CrossRef] [PubMed]
- Rong, C.; Muller, M.F.; Xiang, F.; Jensen, A.; Weichert, W.; Major, G.; Plinkert, P.K.; Hess, J.; Affolter, A. Adaptive ERK signalling activation in response to therapy and in silico prognostic evaluation of EGFR-MAPK in HNSCC. Br. J. Cancer 2020, 123, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Degen, M.; Natarajan, E.; Barron, P.; Widlund, H.R.; Rheinwald, J.G. MAPK/ERK-dependent translation factor hyperactivation and dysregulated laminin γ2 expression in oral dysplasia and squamous cell carcinoma. Am. J. Pathol. 2012, 180, 2462–2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.A.; Lee, K.H.; Lee, D.H.; Lee, J.K.; Lim, S.C.; Joo, Y.E.; Chung, I.J.; Noh, M.G.; Yoon, T.M. Receptor tyrosine kinase, RON, promotes tumor progression by regulating EMT and the MAPK signaling pathway in human oral squamous cell carcinoma. Int. J. Oncol. 2019, 55, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Sha, J.; Bai, Y.; Ngo, H.X.; Okui, T.; Kanno, T. Overview of Evidence-Based Chemotherapy for Oral Cancer: Focus on Drug Resistance Related to the Epithelial-Mesenchymal Transition. Biomolecules 2021, 11, 893. [Google Scholar] [CrossRef]
- Hong, K.O.; Kim, J.H.; Hong, J.S.; Yoon, H.J.; Lee, J.I.; Hong, S.P.; Hong, S.D. Inhibition of Akt activity induces the mesenchymal-to-epithelial reverting transition with restoring E-cadherin expression in KB and KOSCC-25B oral squamous cell carcinoma cells. J. Exp. Clin. Cancer Res. 2009, 28, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, S.Z.; Blair, K.J.; Rahimy, E.; Kiang, A.; Abhold, E.; Fan, J.B.; Wang-Rodriguez, J.; Altuna, X.; Ongkeko, W.M. Salinomycin induces cell death and differentiation in head and neck squamous cell carcinoma stem cells despite activation of epithelial-mesenchymal transition and Akt. BMC Cancer 2012, 12, 556. [Google Scholar] [CrossRef] [Green Version]
- Li, N.Y.; Weber, C.E.; Wai, P.Y.; Cuevas, B.D.; Zhang, J.; Kuo, P.C.; Mi, Z. An MAPK-dependent pathway induces epithelial-mesenchymal transition via Twist activation in human breast cancer cell lines. Surgery 2013, 154, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Massi, D.; Hemmings, B.A.; Mandalà, M.; Hu, Z.; Wicki, A.; Xue, G. AKT-ions with a TWIST between EMT and MET. Oncotarget 2016, 7, 62767–62777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuccarini, M.; Giuliani, P.; Buccella, S.; Di Liberto, V.; Mudò, G.; Belluardo, N.; Carluccio, M.; Rossini, M.; Condorelli, D.F.; Rathbone, M.P.; et al. Modulation of the TGF-β1-induced epithelial to mesenchymal transition (EMT) mediated by P1 and P2 purine receptors in MDCK cells. Purinergic. Signal. 2017, 13, 429–442. [Google Scholar] [CrossRef]
- Haensel, D.; Dai, X. Epithelial-to-mesenchymal transition in cutaneous wound healing: Where we are and where we are heading. Dev. Dyn. 2018, 247, 473–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Angadi, P.V.; Patil, P.V.; Angadi, V.; Mane, D.; Shekar, S.; Hallikerimath, S.; Kale, A.D.; Kardesai, S.G. Immunoexpression of Epithelial Mesenchymal Transition Proteins E-Cadherin, β-Catenin, and N-Cadherin in Oral Squamous Cell Carcinoma. Int. J. Surg. Pathol. 2016, 24, 696–703. [Google Scholar] [CrossRef]
- Ghuwalewala, S.; Ghatak, D.; Das, P.; Dey, S.; Sarkar, S.; Alam, N.; Panda, C.K.; Roychoudhury, S. CD44(high)CD24(low) molecular signature determines the Cancer Stem Cell and EMT phenotype in Oral Squamous Cell Carcinoma. Stem Cell Res. 2016, 16, 405–417. [Google Scholar] [CrossRef] [Green Version]
- Min, A.; Xiong, H.; Wang, W.; Hu, X.; Wang, C.; Mao, T.; Yang, L.; Huang, D.; Xia, K.; Su, T. CD147 promotes proliferation and migration of oral cancer cells by inhibiting junctions between E-cadherin and β-catenin. J. Oral Pathol. Med. 2020, 49, 1019–1029. [Google Scholar] [CrossRef]
- Cui, J.; Huang, W.; Wu, B.; Jin, J.; Jing, L.; Shi, W.P.; Liu, Z.Y.; Yuan, L.; Luo, D.; Li, L.; et al. N-glycosylation by N-acetylglucosaminyltransferase V enhances the interaction of CD147/basigin with integrin β1 and promotes HCC metastasis. J. Pathol. 2018, 245, 41–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dana, P.; Kariya, R.; Vaeteewoottacharn, K.; Sawanyawisuth, K.; Seubwai, W.; Matsuda, K.; Okada, S.; Wongkham, S. Upregulation of CD147 Promotes Metastasis of Cholangiocarcinoma by Modulating the Epithelial-to-Mesenchymal Transitional Process. Oncol. Res. 2017, 25, 1047–1059. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Liu, Y.; Wang, Z.; Yang, Y.; Li, M.; Yuan, D.; Zhang, X.; Li, Y. CD147 Promoted Epithelial Mesenchymal Transition in Airway Epithelial Cells Induced by Cigarette Smoke via Oxidative Stress Signaling Pathway. COPD 2020, 17, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.O.; Lee, J.I.; Hong, S.P.; Hong, S.D. Thymosin β4 induces proliferation, invasion, and epithelial-to-mesenchymal transition of oral squamous cell carcinoma. Amino Acids 2016, 48, 117–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.Y.; Cho, K.H.; Jeong, B.Y.; Park, C.G.; Lee, H.Y. Zeb1 for RCP-induced oral cancer cell invasion and its suppression by resveratrol. Exp. Mol. Med. 2020, 52, 1152–1163. [Google Scholar] [CrossRef]
- Lu, K.; Dong, J.L.; Fan, W.J. Twist1/2 activates MMP2 expression via binding to its promoter in colorectal cancer. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8210–8219. [Google Scholar] [CrossRef]
- Kang, S.U.; Choi, J.W.; Chang, J.W.; Kim, K.I.; Kim, Y.S.; Park, J.K.; Kim, Y.E.; Lee, Y.S.; Yang, S.S.; Kim, C.H. N2 non-thermal atmospheric pressure plasma promotes wound healing in vitro and in vivo: Potential modulation of adhesion molecules and matrix metalloproteinase-9. Exp. Dermatol. 2017, 26, 163–170. [Google Scholar] [CrossRef]
- Fang, F.; Li, Q.; Wu, M.; Nie, C.; Xu, H.; Wang, L. CD147 promotes epithelial-mesenchymal transition of prostate cancer cells via the Wnt/β-catenin pathway. Exp. Ther. Med. 2020, 20, 3154–3160. [Google Scholar] [CrossRef]
- Lai, K.C.; Liu, C.J.; Lin, T.J.; Mar, A.C.; Wang, H.H.; Chen, C.W.; Hong, Z.X.; Lee, T.C. Blocking TNF-α inhibits angiogenesis and growth of IFIT2-depleted metastatic oral squamous cell carcinoma cells. Cancer Lett. 2016, 370, 207–215. [Google Scholar] [CrossRef]
- Lee, C.H.; Chang, J.S.; Syu, S.H.; Wong, T.S.; Chan, J.Y.; Tang, Y.C.; Yang, Z.P.; Yang, W.C.; Chen, C.T.; Lu, S.C.; et al. IL-1β promotes malignant transformation and tumor aggressiveness in oral cancer. J. Cell Physiol. 2015, 230, 875–884. [Google Scholar] [CrossRef]
- Xiao, L.; Li, X.; Cao, P.; Fei, W.; Zhou, H.; Tang, N.; Liu, Y. Interleukin-6 mediated inflammasome activation promotes oral squamous cell carcinoma progression via JAK2/STAT3/Sox4/NLRP3 signaling pathway. J. Exp. Clin. Cancer Res. 2022, 41, 166. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Kawabata-Iwakawa, R.; Ida, S.; Mito, I.; Tada, H.; Chikamatsu, K. Upregulated glycolysis correlates with tumor progression and immune evasion in head and neck squamous cell carcinoma. Sci. Rep. 2021, 11, 17789. [Google Scholar] [CrossRef] [PubMed]
- Botha, H.; Farah, C.S.; Koo, K.; Cirillo, N.; McCullough, M.; Paolini, R.; Celentano, A. The Role of Glucose Transporters in Oral Squamous Cell Carcinoma. Biomolecules 2021, 11, 1070. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Peng, Y.; Zhao, J.; Xie, Z.; Lei, X.; Tang, G. Discovery and development of tumor glycolysis rate-limiting enzyme inhibitors. Bioorg. Chem. 2021, 112, 104891. [Google Scholar] [CrossRef] [PubMed]
- Chandel, V.; Maru, S.; Kumar, A.; Kumar, A.; Sharma, A.; Rathi, B.; Kumar, D. Role of monocarboxylate transporters in head and neck squamous cell carcinoma. Life Sci. 2021, 279, 119709. [Google Scholar] [CrossRef] [PubMed]
- Pezzuto, A.; Carico, E. Role of HIF-1 in Cancer Progression: Novel Insights. A Review. Curr. Mol. Med. 2018, 18, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Bharti, A.; Urs, A.B.; Kumar, P. Significance of HIF-1α Expression and LOXL-2 Localization in Progression of Oral Squamous Cell Carcinoma. Asian Pac. J. Cancer Prev. 2021, 22, 341–347. [Google Scholar] [CrossRef]
- Jensen, D.H.; Therkildsen, M.H.; Dabelsteen, E. A reverse Warburg metabolism in oral squamous cell carcinoma is not dependent upon myofibroblasts. J. Oral Pathol. Med. 2015, 44, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.A.; Kamal, M.A.; Akhtar, S. Tumor Angiogenesis and VEGFR-2, Mechanism, Pathways and Current Biological Therapeutic Interventions. Curr. Drug Metab. 2021, 22, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, N.; Uma Rao, K.D.; Ranganathan, K.; Joshua, E.; Thavarajah, R. Study of expression of endoglin (CD105) in oral squamous cell carcinoma. J. Oral Maxillofac. Pathol. 2021, 25, 552. [Google Scholar] [CrossRef]
- Essa, A.A.M.; Deraz, E.M. Expression of CD44 (NKI-P1) in oral squamous cell carcinoma associated vascular endothelial cells: A relationship to tumor angiogenesis. Saudi Dent. J. 2022, 34, 21–26. [Google Scholar] [CrossRef]
- Toffoli, S.; Feron, O.; Raes, M.; Michiels, C. Intermittent hypoxia changes HIF-1alpha phosphorylation pattern in endothelial cells: Unravelling of a new PKA-dependent regulation of HIF-1alpha. Biochim. Biophys. Acta 2007, 1773, 1558–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valle, I.B.; Schuch, L.F.; da Silva, J.M.; Gala-García, A.; Diniz, I.M.A.; Birbrair, A.; Abreu, L.G.; Silva, T.A. Pericyte in Oral Squamous Cell Carcinoma: A Systematic Review. Head Neck Pathol. 2020, 14, 1080–1091. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, R.; Ghosh, B.; Mandal, M.; Nawn, D.; Banerjee, S.; Pal, M.; Paul, R.R.; Banerjee, S.; Chatterjee, J. Pathophysiological relationship between hypoxia associated oxidative stress, Epithelial-mesenchymal transition, stemness acquisition and alteration of Shh/Gli-1 axis during oral sub-mucous fibrosis and oral squamous cell carcinoma. Eur. J. Cell Biol. 2021, 100, 151146. [Google Scholar] [CrossRef]
- Mohanta, S.; Siddappa, G.; Valiyaveedan, S.G.; Dodda Thimmasandra Ramanjanappa, R.; Das, D.; Pandian, R.; Khora, S.S.; Kuriakose, M.A.; Suresh, A. Cancer stem cell markers in patterning differentiation and in prognosis of oral squamous cell carcinoma. Tumour. Biol. 2017, 39, 1010428317703656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, R.C.; Lopes, N.M.; Amôr, N.G.; Ponce, J.B.; Schmerling, C.K.; Lara, V.S.; Moyses, R.A.; Rodini, C.O. CD44 and ALDH1 immunoexpression as prognostic indicators of invasion and metastasis in oral squamous cell carcinoma. J. Oral Pathol. Med. 2018, 47, 740–747. [Google Scholar] [CrossRef]
- Cirillo, N.; Wu, C.; Prime, S.S. Heterogeneity of Cancer Stem Cells in Tumorigenesis, Metastasis, and Resistance to Antineoplastic Treatment of Head and Neck Tumours. Cells 2021, 10, 3068. [Google Scholar] [CrossRef]
- de Freitas Filho, S.A.J.; Coutinho-Camillo, C.M.; Oliveira, K.K.; Bettim, B.B.; Pinto, C.A.L.; Kowalski, L.P.; Oliveira, D.T. Prognostic Implications of ALDH1 and Notch1 in Different Subtypes of Oral Cancer. J. Oncol. 2021, 2021, 6663720. [Google Scholar] [CrossRef]
- Singh, P.; Augustine, D.; Rao, R.S.; Patil, S.; Awan, K.H.; Sowmya, S.V.; Haragannavar, V.C.; Prasad, K. Role of cancer stem cells in head-and-neck squamous cell carcinoma—A systematic review. J. Carcinog. 2021, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Emich, H.; Chapireau, D.; Hutchison, I.; Mackenzie, I. The potential of CD44 as a diagnostic and prognostic tool in oral cancer. J. Oral Pathol. Med. 2015, 44, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Lee, H.Y. Targeting Cancer Stem Cell Markers or Pathways: A Potential Therapeutic Strategy for Oral Cancer Treatment. Int. J. Stem Cells 2021, 14, 386–399. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Jiang, J.; Chen, H.N.; Zhou, L.; Huang, Z.; Qin, S.; Jin, P.; Luo, M.; Li, B.; Shi, J.; et al. Redox-sensitive cyclophilin A elicits chemoresistance through realigning cellular oxidative status in colorectal cancer. Cell Rep. 2021, 37, 110069. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.J.; Kim, H.P.; Lee, K.S.; Yoo, Y.D.; Kwon, Y.T.; Kim, K.M.; Kim, T.Y.; Yi, E.C. Proteomic analysis reveals that CD147/EMMPRIN confers chemoresistance in cancer stem cell-like cells. Proteomics 2013, 13, 1714–1725. [Google Scholar] [CrossRef]
- Wu, J.; Li, Y.; Dang, Y.Z.; Gao, H.X.; Jiang, J.L.; Chen, Z.N. HAb18G/CD147 promotes radioresistance in hepatocellular carcinoma cells: A potential role for integrin β1 signaling. Mol. Cancer Ther. 2015, 14, 553–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezayatmand, H.; Razmkhah, M.; Razeghian-Jahromi, I. Drug resistance in cancer therapy: The Pandora’s Box of cancer stem cells. Stem Cell Res. 2022, 13, 181. [Google Scholar] [CrossRef]
- Lin, S.C.; Wang, C.P.; Chen, Y.M.; Lu, S.Y.; Fann, M.J.; Liu, C.J.; Kao, S.Y.; Chang, K.W. Regulation of IGFBP-5 expression during tumourigenesis and differentiation of oral keratinocytes. J. Pathol. 2002, 198, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.K.; Chen, W.; Park, N.H. Regulation of Epithelial Cell Proliferation, Differentiation, and Plasticity by Grainyhead-Like 2 During Oral Carcinogenesis. Crit. Rev. Oncog. 2018, 23, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, D.; Bruland, O.; Parajuli, H.; Osman, T.A.; Teh, M.T.; Johannessen, A.C.; Costea, D.E. S100A16 promotes differentiation and contributes to a less aggressive tumor phenotype in oral squamous cell carcinoma. BMC Cancer 2015, 15, 631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkataswamy, P.; Samudrala Venkatesiah, S.; Rao, R.S.; Banavar, S.R.; Patil, S.; Augustine, D.; Haragannavar, V.C. Immunohistochemical expression of Tazarotene-induced Gene 3 in oral squamous cell carcinoma. J. Oral Pathol. Med. 2021, 50, 403–409. [Google Scholar] [CrossRef]
- Samman, M.; Wood, H.M.; Conway, C.; Stead, L.; Daly, C.; Chalkley, R.; Berri, S.; Senguven, B.; Ross, L.; Egan, P.; et al. A novel genomic signature reclassifies an oral cancer subtype. Int. J. Cancer 2015, 137, 2364–2373. [Google Scholar] [CrossRef] [Green Version]
- Nagler, R.; Weizman, A.; Gavish, A. Cigarette smoke, saliva, the translocator protein 18 kDa (TSPO), and oral cancer. Oral Dis. 2019, 25, 1843–1849. [Google Scholar] [CrossRef] [Green Version]
- Ding, P.; Zhang, X.; Jin, S.; Duan, B.; Chu, P.; Zhang, Y.; Chen, Z.N.; Xia, B.; Song, F. CD147 functions as the signaling receptor for extracellular divalent copper in hepatocellular carcinoma cells. Oncotarget 2017, 8, 51151–51163. [Google Scholar] [CrossRef] [Green Version]
- Ke, X.; Li, L.; Dong, H.L.; Chen, Z.N. Acquisition of anoikis resistance through CD147 upregulation: A new mechanism underlying metastasis of hepatocellular carcinoma cells. Oncol. Lett. 2012, 3, 1249–1254. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, J.; Xu, Y.; Li, L.; Zheng, M. CD147 promotes glucose metabolism, invasion and metastasis via PI3K/AKT pathway in oral squamous cell carcinomas. Transl. Cancer Res. 2019, 8, 1486–1496. [Google Scholar] [CrossRef] [PubMed]
- Fei, F.; Li, X.; Xu, L.; Li, D.; Zhang, Z.; Guo, X.; Yang, H.; Chen, Z.; Xing, J. CD147-CD98hc complex contributes to poor prognosis of non-small cell lung cancer patients through promoting cell proliferation via the PI3K/Akt signaling pathway. Ann. Surg. Oncol. 2014, 21, 4359–4368. [Google Scholar] [CrossRef]
- Kuang, Y.H.; Chen, X.; Su, J.; Wu, L.S.; Liao, L.Q.; Li, D.; Chen, Z.S.; Kanekura, T. RNA interference targeting the CD147 induces apoptosis of multi-drug resistant cancer cells related to XIAP depletion. Cancer Lett. 2009, 276, 189–195. [Google Scholar] [CrossRef]
- Hahn, J.N.; Kaushik, D.K.; Mishra, M.K.; Wang, J.; Silva, C.; Yong, V.W. Impact of Minocycline on Extracellular Matrix Metalloproteinase Inducer, a Factor Implicated in Multiple Sclerosis Immunopathogenesis. J. Immunol. 2016, 197, 3850–3860. [Google Scholar] [CrossRef] [Green Version]
- Meng, Q.; Pu, L.; Lu, Q.; Wang, B.; Li, S.; Liu, B.; Li, F. Morin hydrate inhibits atherosclerosis and LPS-induced endothelial cells inflammatory responses by modulating the NFκB signaling-mediated autophagy. Int. Immunopharmacol. 2021, 100, 108096. [Google Scholar] [CrossRef]
- Liu, R.; Tan, Q.; Luo, Q. Decreased expression level and DNA-binding activity of specificity protein 1 via cyclooxygenase-2 inhibition antagonizes radiation resistance, cell migration and invasion in radiation-resistant lung cancer cells. Oncol. Lett. 2018, 16, 3029–3037. [Google Scholar] [CrossRef]
- Arora, R.; Bharti, V.; Gaur, P.; Aggarwal, S.; Mittal, M.; Das, S.N. Operculina turpethum extract inhibits growth and proliferation by inhibiting NF-κB, COX-2 and cyclin D1 and induces apoptosis by up regulating P53 in oral cancer cells. Arch. Oral Biol. 2017, 80, 1–9. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| CD147 Stimulator | Action | Reference |
|---|---|---|
| Fyn tyrosin kinase | Triggering CD147 expression | Ramos DM et al. [36] |
| Interleukin-1 | Induction of CD147 expression | Wang Q et al. [37] |
| Epidermal Growth Factor | Upregulation of CD147 expression | Omi Y et al. [38] |
| Transforming Growth Factor-β1 | Upregulation of CD147 expression | Wang W et al. [39] |
| Glycosyltransferases | CD147 activation | Bai Y et al. [32] |
| CD147 anchored to the surface of (or released by) neighboring cells | CD147 activation | Guindolet D et al. [26] |
| Galectin 3 | CD147 activation | Mauris J et al. [28] |
| Cyclophilin A | CD147 activation | Takahashi M et al. [33] |
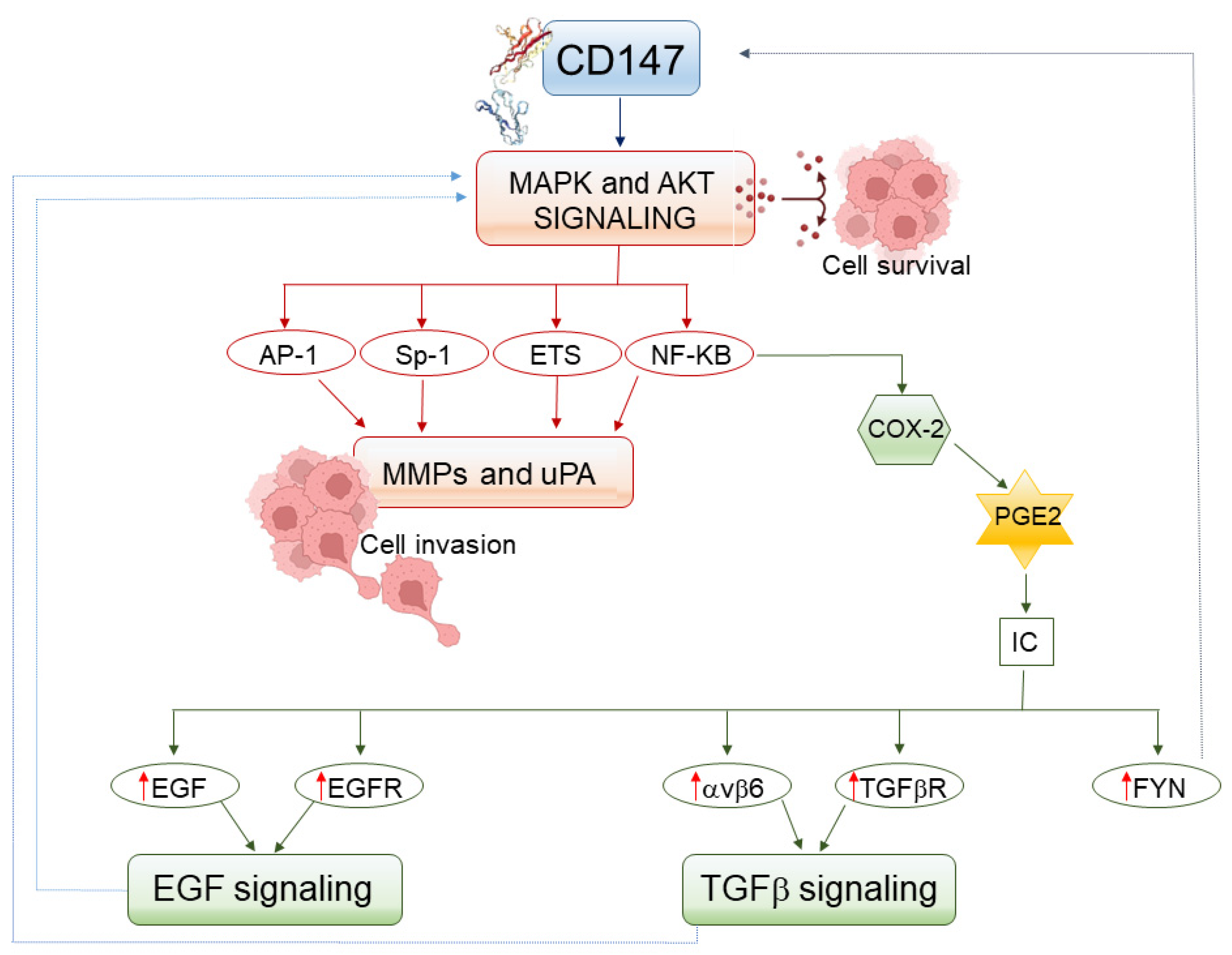
| CD147-Targeted Molecule | Effect of the Action Carried Out by CD147 | References |
|---|---|---|
| α3β1, α6β1 | basal epithelial cells’ adhesion to the basement membrane | Richard V et al. [41] |
| MMP-1 | disruption of intercellular adhesion, epithelial cell locomotion and growth | Cao Z et al. [18] |
| MT1-MMP | ECM degradation, MMP-2 or -9 activation, cellular invasion | Mitre GP et al. [44] |
| MMP-2 | ECM degradation, cellular invasion | Luo Z et al. [42] |
| MMP-9 | ECM degradation, cellular invasion | Suzuki S et al. [43] |
| TIMPs | increase in MMPs activity | Maghsood F et al. [47] |
| uPA | ECM degradation, plasminogen or MMPs activation, cellular invasion | Lescaille G et al. [48] |
| Tenascin | facilitation of OSCC cell migration | Dang D et al. [49] |
| NF-kB | induction of COX-2, inflammatory cytokines, and MMPs expression | Yu B et al. [50] |
| Endothelial selectin | leukocytes extravasation | Muramatsu T [51] |
| EGF and TGF-β1 | EMT and cell invasion | Wu J et al. [52] |
| VEGF | Angiogenesis | Tang Y et al. [53] |
| ZEB, SNAI, TWIST | EMT, MMPs expression | Siu A et al. [54] |
| GLUTs | increase in glucose uptake by OSCC cells | Almeida LMCA et al. [55] |
| MCTs | lactate export from OSCC cells, functional impairment of CD8+ T cells, HIF-1 activation | Kirk P et al. [56] |
| HIF-1 | MMPs or VEGF expression, cell invasion, angiogenesis | Wang CH et al. [57] |
| CD44 | survival, anchorage-independent growth, and drug resistance of OSCC cells | Richard V et al. [41] |
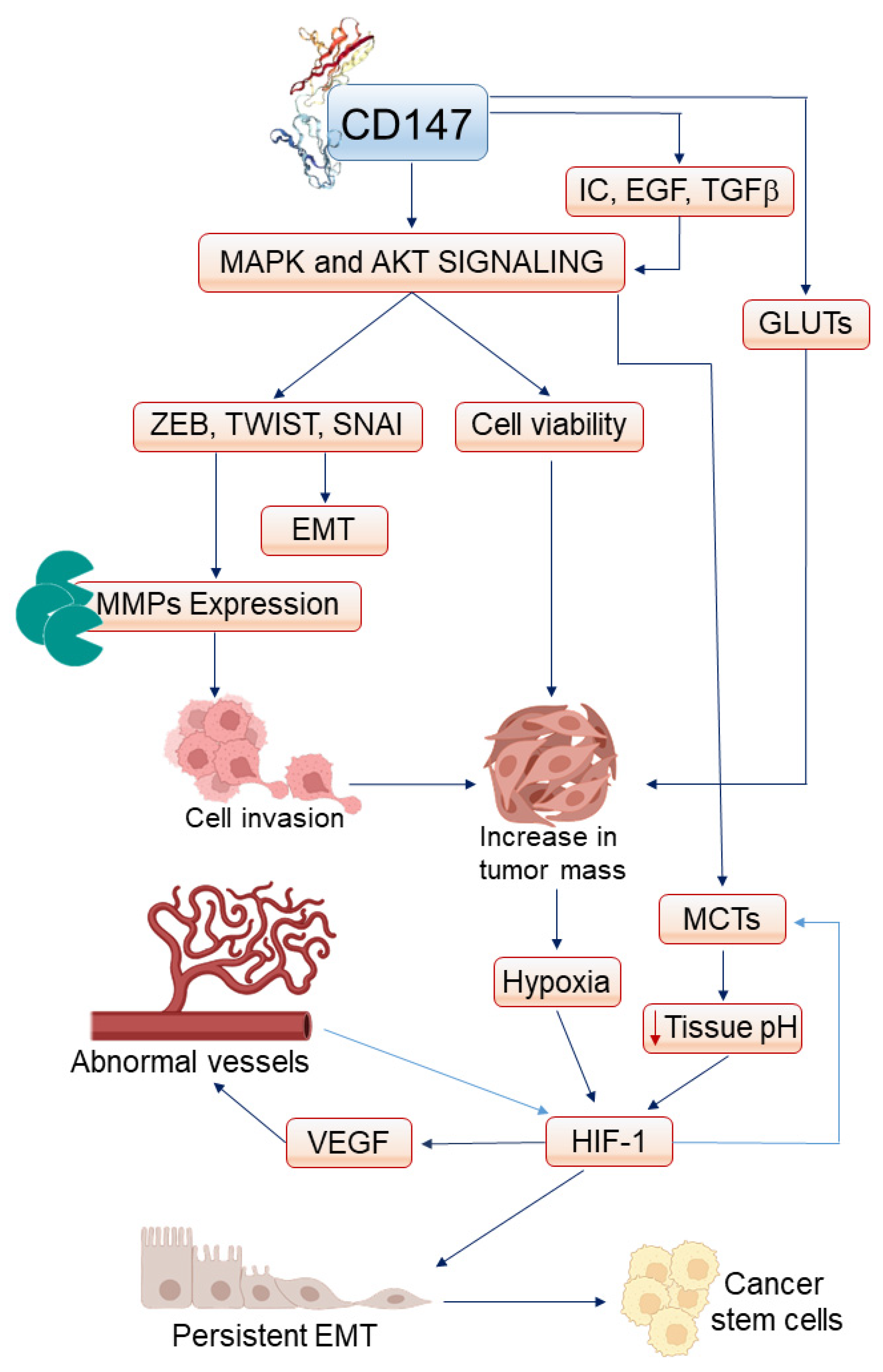
| Effect | Consequence | Reference |
|---|---|---|
| Synthesis of MMPs | Cell invasion | Ding P et al. [232] |
| Activation of the NF-kB/COX-2 axis | Inflammatory cytokines expression, EMT | Dana P et al. [120] |
| Upregulation of MCTs | Lowering of tissue pH, activation of HIF-1 | Dana P et al. [63] |
| Synthesis of VEGF | Angiogenesis | Tang Y et al. [53] |
| Cell survival—I | Circulating cancer cells escape anoikis | Ke X et al. [233] |
| Cell survival—II | Cancer cells resist chemotherapy | Kang MJ et al. [223] |
| Cell survival—III | Cancer cells resist radiotherapy | Wu J et al. [224] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barillari, G.; Melaiu, O.; Gargari, M.; Pomella, S.; Bei, R.; Campanella, V. The Multiple Roles of CD147 in the Development and Progression of Oral Squamous Cell Carcinoma: An Overview. Int. J. Mol. Sci. 2022, 23, 8336. https://doi.org/10.3390/ijms23158336
Barillari G, Melaiu O, Gargari M, Pomella S, Bei R, Campanella V. The Multiple Roles of CD147 in the Development and Progression of Oral Squamous Cell Carcinoma: An Overview. International Journal of Molecular Sciences. 2022; 23(15):8336. https://doi.org/10.3390/ijms23158336
Chicago/Turabian StyleBarillari, Giovanni, Ombretta Melaiu, Marco Gargari, Silvia Pomella, Roberto Bei, and Vincenzo Campanella. 2022. "The Multiple Roles of CD147 in the Development and Progression of Oral Squamous Cell Carcinoma: An Overview" International Journal of Molecular Sciences 23, no. 15: 8336. https://doi.org/10.3390/ijms23158336
APA StyleBarillari, G., Melaiu, O., Gargari, M., Pomella, S., Bei, R., & Campanella, V. (2022). The Multiple Roles of CD147 in the Development and Progression of Oral Squamous Cell Carcinoma: An Overview. International Journal of Molecular Sciences, 23(15), 8336. https://doi.org/10.3390/ijms23158336