
ATP-Binding Cassette G Transporters and Their Multiple Roles Especially for Male Fertility in Arabidopsis, Rice and Maize

 ,
, 
Abstract
:1. Introduction
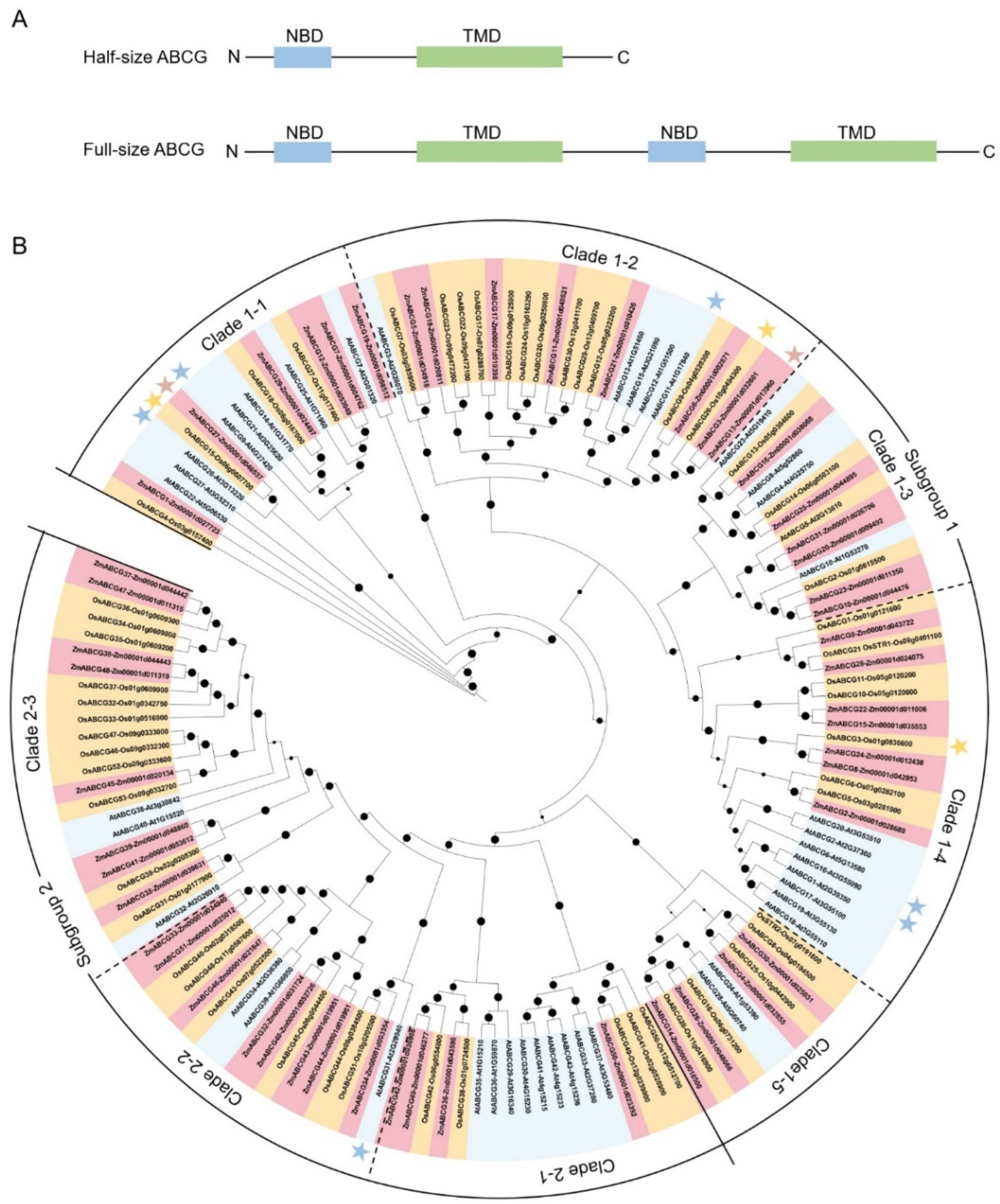
2. The ABCG Transporters in Maize and Their Phylogenetic Analysis
2.1. The Nomenclature and Characterization of the Maize ABCG Subfamily
2.2. Phylogenetic Analysis and Classification of ABCGs
3. Multiple Functions of ABCG Transporters in Arabidopsis, Rice and Maize
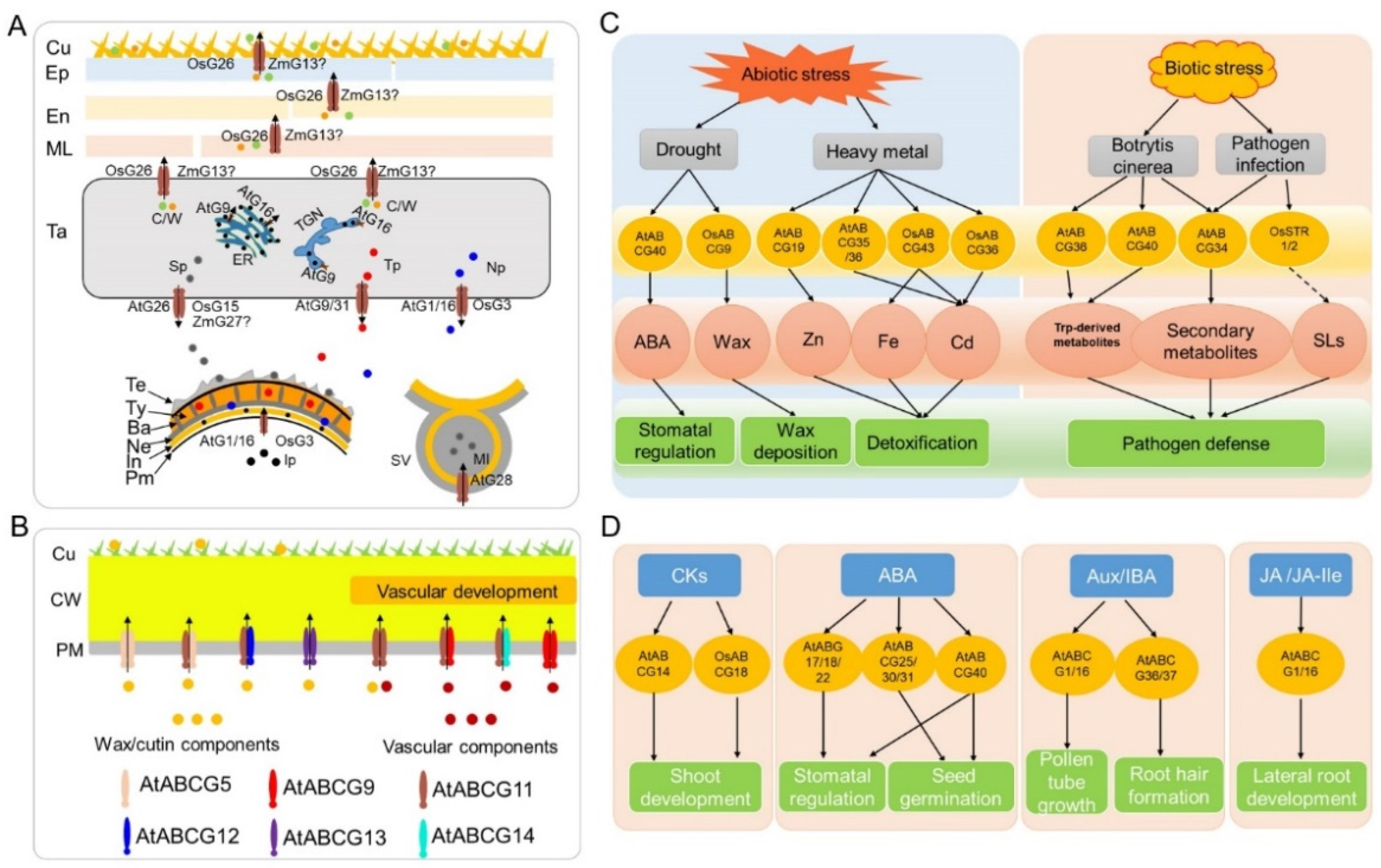
3.1. Anther and Pollen Development
3.2. Vegetative and Female Organ Development
3.3. Biotic and Abiotic Stress Response
3.4. Hormone Transport and Signaling
4. Functional Predictions of ABCG Genes in Maize
4.1. Functional Prediction of ABCGs Based on Homologous Analysis
4.2. Functional Prediction of ABCGs Based on Bioinformatic Analysis
5. Substrate Identification of Plant ABCG Transporters
6. Conclusions and Perspective
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Higgins, C.F. ABC transporters: From microorganisms to man. Annu. Rev. Cell. Biol. 1992, 8, 67–113. [Google Scholar] [CrossRef] [PubMed]
- Lefevre, F.; Boutry, M. Towards identification of the substrates of ATP-binding cassette transporters. Plant Physiol. 2018, 178, 18–39. [Google Scholar] [CrossRef] [PubMed]
- Verrier, P.J.; Bird, D.; Burla, B.; Dassa, E.; Forestier, C.; Geisler, M.; Klein, M.; Kolukisaoglu, U.; Lee, Y.; Martinoia, E.; et al. Plant ABC proteins-a unified nomenclature and updated inventory. Trends Plant Sci. 2008, 13, 151–159. [Google Scholar] [CrossRef]
- Hwang, J.U.; Song, W.Y.; Hong, D.; Ko, D.; Yamaoka, Y.; Jang, S.; Yim, S.; Lee, E.; Khare, D.; Kim, K.; et al. Plant ABC transporters enable many unique aspects of a terrestrial plant’s lifestyle. Mol. Plant 2016, 9, 338–355. [Google Scholar] [CrossRef] [PubMed]
- Do, T.H.T.; Martinoia, E.; Lee, Y.; Hwang, J.U. 2021 update on ATP-binding cassette (ABC) transporters: How they meet the needs of plants. Plant Physiol. 2021, 187, 1876–1892. [Google Scholar] [CrossRef]
- Pang, K.; Li, Y.; Liu, M.; Meng, Z.; Yu, Y. Inventory and general analysis of the ATP-binding cassette (ABC) gene superfamily in maize (Zea mays L.). Gene 2013, 526, 411–428. [Google Scholar] [CrossRef]
- Fang, C.; Wu, S.; Niu, C.; Hou, Q.; An, X.; Wei, X.; Zhao, L.; Jiang, Y.; Liu, X.; Wan, X. Triphasic regulation of ZmMs13 encoding an ABCG transporter is sequentially required for callose dissolution, pollen exine and anther cuticle formation in maize. J. Adv. Res. 2022, 38, JARE-S-22-01434. [Google Scholar]
- Jiang, Y.; Li, Z.; Liu, X.; Zhu, T.; Xie, K.; Hou, Q.; Yan, T.; Niu, C.; Zhang, S.; Yang, M.; et al. ZmFAR1 and ZmABCG26 regulated by microRNA are essential for lipid metabolism in maize anther. Int. J. Mol. Sci. 2021, 22, 7916. [Google Scholar] [CrossRef]
- Li, L.; Li, D.; Liu, S.; Ma, X.; Dietrich, C.R.; Hu, H.C.; Zhang, G.; Liu, Z.; Zheng, J.; Wang, G.; et al. The maize glossy13 gene, cloned via BSR-Seq and Seq-walking encodes a putative ABC transporter required for the normal accumulation of epicuticular waxes. PLoS ONE 2013, 8, e82333. [Google Scholar] [CrossRef]
- Xu, Q.; Yang, L.; Kang, D.; Ren, Z.; Liu, Y. Maize MS2 encodes an ATP-binding cassette transporter that is essential for anther development. Crop J. 2021, 9, 1301–1308. [Google Scholar] [CrossRef]
- Choi, H.; Ohyama, K.; Kim, Y.Y.; Jin, J.Y.; Lee, S.B.; Yamaoka, Y.; Muranaka, T.; Suh, M.C.; Fujioka, S.; Lee, Y. The role of Arabidopsis ABCG9 and ABCG31 ATP binding cassette transporters in pollen fitness and the deposition of steryl glycosides on the pollen coat. Plant Cell 2014, 26, 310–324. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Jin, J.-Y.; Choi, S.; Hwang, J.-U.; Kim, Y.-Y.; Suh, M.C.; Lee, Y. An ABCG/WBC-type ABC transporter is essential for transport of sporopollenin precursors for exine formation in developing pollen. Plant J. 2011, 65, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Quilichini, T.D.; Friedmann, M.C.; Samuels, A.L.; Douglas, C.J. ATP-binding cassette transporter G26 is required for male fertility and pollen exine formation in arabidopsis. Plant Physiol. 2010, 154, 678–690. [Google Scholar] [CrossRef] [PubMed]
- Quilichini, T.D.; Samuels, A.L.; Douglas, C.J. ABCG26-mediated polyketide trafficking and hydroxycinnamoyl spermidines contribute to pollen wall exine formation in Arabidopsis. Plant Cell 2014, 26, 4483–4498. [Google Scholar] [CrossRef]
- Niu, B.X.; He, F.R.; He, M.; Ren, D.; Chen, L.T.; Liu, Y.G. The ATP-binding cassette transporter OsABCG15 is required for anther development and pollen fertility in rice. J. Integr. Plant Biol. 2013, 55, 710–720. [Google Scholar] [CrossRef]
- Qin, P.; Tu, B.; Wang, Y.; Deng, L.; Quilichini, T.D.; Li, T.; Wang, H.; Ma, B.; Li, S. ABCG15 encodes an ABC transporter protein, and is essential for post-meiotic anther and pollen exine development in rice. Plant Cell Physiol. 2013, 54, 138–154. [Google Scholar] [CrossRef]
- Panikashvili, D.; Shi, J.X.; Bocobza, S.; Franke, R.B.; Schreiber, L.; Aharoni, A. The Arabidopsis DSO/ABCG11 transporter affects cutin metabolism in reproductive organs and suberin in roots. Mol. Plant 2010, 3, 563–575. [Google Scholar] [CrossRef]
- Chang, Z.; Chen, Z.; Yan, W.; Xie, G.; Lu, J.; Wang, N.; Lu, Q.; Yao, N.; Yang, G.; Xia, J.; et al. An ABC transporter, OsABCG26, is required for anther cuticle and pollen exine formation and pollen-pistil interactions in rice. Plant Sci. 2016, 253, 21–30. [Google Scholar] [CrossRef]
- Zhao, G.; Shi, J.; Liang, W.; Xue, F.; Luo, Q.; Zhu, L.; Qu, G.; Chen, M.; Schreiber, L.; Zhang, D. Two ATP binding cassette g transporters, rice ATP binding cassette G26 and ATP binding cassette G15, collaboratively regulate rice male reproduction. Plant Physiol. 2015, 169, 2064–2079. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, L.; Chen, P.; Cai, H.; Hou, Z.; Jin, X.; Aslam, M.; Chai, M.; Lai, L.; He, Q.; et al. ATP binding cassette transporters ABCG1 and ABCG16 affect reproductive development via auxin signalling in Arabidopsis. Plant J. 2020, 102, 1172–1186. [Google Scholar] [CrossRef]
- Yadav, V.; Molina, I.; Ranathunge, K.; Castillo, I.Q.; Rothstein, S.J.; Reed, J.W. ABCG transporters are required for suberin and pollen wall extracellular barriers in Arabidopsis. Plant Cell 2014, 26, 3569–3588. [Google Scholar] [CrossRef] [PubMed]
- Yim, S.; Khare, D.; Kang, J.; Hwang, J.U.; Liang, W.; Martinoia, E.; Zhang, D.; Kang, B.; Lee, Y. Postmeiotic development of pollen surface layers requires two Arabidopsis ABC-type transporters. Plant Cell Rep. 2016, 35, 1863–1873. [Google Scholar] [CrossRef]
- Chang, Z.; Jin, M.; Yan, W.; Chen, H.; Qiu, S.; Fu, S.; Xia, J.; Liu, Y.; Chen, Z.; Wu, J.; et al. The ATP-binding cassette (ABC) transporter OsABCG3 is essential for pollen development in rice. Rice 2018, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Zou, T.; Yuan, G.; He, Z.; Li, W.; Tao, Y.; Liu, M.; Zhou, D.; Zhao, H.; Zhu, J.; et al. Less and shrunken pollen 1 (LSP1) encodes a member of the ABC transporter family required for pollen wall development in rice (Oryza sativa L.). Crop J. 2020, 8, 492–504. [Google Scholar] [CrossRef]
- Le Hir, R.; Sorin, C.; Chakraborti, D.; Moritz, T.; Schaller, H.; Tellier, F.; Robert, S.; Morin, H.; Bako, L.; Bellini, C. ABCG9, ABCG11 and ABCG14 ABC transporters are required for vascular development in Arabidopsis. Plant J. 2013, 76, 811–824. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, H.E.; Shin, J.J.; Bird, D.A.; Samuels, A.L. Arabidopsis ABCG transporters, which are required for export of diverse cuticular lipids, dimerize in different combinations. Plant Cell 2010, 22, 3066–3075. [Google Scholar] [CrossRef]
- Shanmugarajah, K.; Linka, N.; Grafe, K.; Smits, S.H.J.; Weber, A.P.M.; Zeier, J.; Schmitt, L. ABCG1 contributes to suberin formation in Arabidopsis thaliana roots. Sci. Rep. 2019, 9, 11381. [Google Scholar] [CrossRef]
- Li, Q.; Zheng, J.; Li, S.; Huang, G.; Skilling, S.J.; Wang, L.; Li, L.; Li, M.; Yuan, L.; Liu, P. Transporter-mediated nuclear entry of jasmonoyl-isoleucine is essential for jasmonate signaling. Mol. Plant 2017, 10, 695–708. [Google Scholar] [CrossRef]
- Wang, F.; Yu, G.; Liu, P. Transporter-mediated subcellular distribution in the metabolism and signaling of jasmonates. Front. Plant Sci. 2019, 10, 390. [Google Scholar] [CrossRef]
- Kang, J.; Yim, S.; Choi, H.; Kim, A.; Lee, K.P.; Lopez-Molina, L.; Martinoia, E.; Lee, Y. Abscisic acid transporters cooperate to control seed germination. Nat. Commun. 2015, 6, 8113. [Google Scholar] [CrossRef]
- Do, T.H.T.; Choi, H.; Palmgren, M.; Martinoia, E.; Hwang, J.U.; Lee, Y. Arabidopsis ABCG28 is required for the apical accumulation of reactive oxygen species in growing pollen tubes. Proc. Natl. Acad. Sci. USA 2019, 116, 12540–12549. [Google Scholar] [CrossRef] [PubMed]
- Fedi, F.; O’Neill, C.M.; Menard, G.; Trick, M.; Dechirico, S.; Corbineau, F.; Bailly, C.; Eastmond, P.J.; Penfield, S. Awake1, an ABC-type transporter, reveals an essential role for suberin in the control of seed dormancy. Plant Physiol. 2017, 174, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Kim, K.Y.; Zhang, J.; Yamaoka, Y.; Gao, P.; Kim, H.; Hwang, J.U.; Suh, M.C.; Kang, B.; Lee, Y. Arabidopsis seedling establishment under waterlogging requires ABCG5-mediated formation of a dense cuticle layer. New Phytol. 2021, 229, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Shiono, K.; Ando, M.; Nishiuchi, S.; Takahashi, H.; Watanabe, K.; Nakamura, M.; Matsuo, Y.; Yasuno, N.; Yamanouchi, U.; Fujimoto, M.; et al. RCN1/OsABCG5, an ATP-binding cassette (ABC) transporter, is required for hypodermal suberization of roots in rice (Oryza sativa). Plant J. 2014, 80, 40–51. [Google Scholar] [CrossRef]
- Yasuno, N.; Takamure, I.; Kidou, S.I.; Tokuji, Y.; Ureshi, A.N.; Funabiki, A.; Ashikaga, K.; Yamanouchi, U.; Yano, M.; Kato, K. Rice shoot branching requires an ATP-binding cassette subfamily G protein. New Phytol. 2009, 182, 91–101. [Google Scholar] [CrossRef]
- Pighin, J.A.; Zheng, H.; Balakshin, L.J.; Goodman, I.P.; Western, T.L.; Jetter, R.; Kunst, L.; Samuels, A.L. Plant cuticular lipid export requires an ABC transporter. Science 2004, 306, 702–704. [Google Scholar] [CrossRef]
- Panikashvili, D.; Shi, J.X.; Schreiber, L.; Aharoni, A. The Arabidopsis ABCG13 transporter is required for flower cuticle secretion and patterning of the petal epidermis. New Phytol. 2011, 190, 113–124. [Google Scholar] [CrossRef]
- Takeda, S.; Iwasaki, A.; Tatematsu, K.; Okada, K. The half-size ABC transporter FOLDED PETALS 2/ABCG13 is involved in petal elongation through narrow spaces in Arabidopsis thaliana floral buds. Plants 2014, 3, 348–358. [Google Scholar] [CrossRef]
- Kuromori, T.; Sugimoto, E.; Ohiraki, H.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Functional relationship of AtABCG21 and AtABCG 22 in stomatal regulation. Sci. Rep. 2017, 7, 12501. [Google Scholar] [CrossRef]
- Takeuchi, M.; Kegasa, T.; Watanabe, A.; Tamura, M.; Tsutsumi, Y. Expression analysis of transporter genes for screening candidate monolignol transporters using Arabidopsis thaliana cell suspensions during tracheary element differentiation. J. Plant Res. 2018, 131, 297–305. [Google Scholar] [CrossRef]
- Kim, K.; Choi, B.Y.; Kang, J.; Shim, D.; Martinoia, E.; Lee, Y. Arabidopsis ABCG27 plays an essential role in flower and leaf development by modulating abscisic acid content. Physiol. Plant 2022, 174, e13734. [Google Scholar] [CrossRef] [PubMed]
- Alejandro, S.; Lee, Y.; Tohge, T.; Sudre, D.; Osorio, S.; Park, J.; Bovet, L.; Lee, Y.; Geldner, N.; Fernie, A.R.; et al. AtABCG29 is a monolignol transporter involved in lignin biosynthesis. Curr. Biol. 2012, 22, 1207–1212. [Google Scholar] [CrossRef] [PubMed]
- Bessire, M.; Borel, S.; Fabre, G.; Carraca, L.; Efremova, N.; Yephremov, A.; Cao, Y.; Jetter, R.; Jacquat, A.C.; Metraux, J.P.; et al. A member of the pleiotropic drug resistance family of ATP binding cassette transporters is required for the formation of a functional cuticle in Arabidopsis. Plant Cell 2011, 23, 1958–1970. [Google Scholar] [CrossRef] [PubMed]
- Elejalde-Palmett, C.; Martinez San Segundo, I.; Garroum, I.; Charrier, L.; De Bellis, D.; Mucciolo, A.; Guerault, A.; Liu, J.; Zeisler-Diehl, V.; Aharoni, A.; et al. ABCG transporters export cutin precursors for the formation of the plant cuticle. Curr. Biol. 2021, 31, 2111–2123.e9. [Google Scholar] [CrossRef]
- Garroum, I.; Bidzinski, P.; Daraspe, J.; Mucciolo, A.; Humbel, B.M.; Morel, J.B.; Nawrath, C. Cuticular defects in Oryza sativa ATP-binding cassette transporter G31 mutant plants cause dwarfism, elevated defense responses and pathogen resistance. Plant Cell Physiol. 2016, 57, 1179–1188. [Google Scholar] [CrossRef]
- Kaneda, M.; Schuetz, M.; Lin, B.S.; Chanis, C.; Hamberger, B.; Western, T.L.; Ehlting, J.; Samuels, A.L. ABC transporters coordinately expressed during lignification of Arabidopsis stems include a set of ABCBs associated with auxin transport. J. Exp. Bot. 2011, 62, 2063–2077. [Google Scholar] [CrossRef]
- Khare, D.; Choi, H.; Huh, S.U.; Bassin, B.; Kim, J.; Martinoia, E.; Sohn, K.H.; Paek, K.H.; Lee, Y. Arabidopsis ABCG34 contributes to defense against necrotrophic pathogens by mediating the secretion of camalexin. Proc. Natl. Acad. Sci. USA 2017, 114, E5712–E5720. [Google Scholar] [CrossRef]
- Fourcroy, P.; Siso-Terraza, P.; Sudre, D.; Saviron, M.; Reyt, G.; Gaymard, F.; Abadia, A.; Abadia, J.; Alvarez-Fernandez, A.; Briat, J.F. Involvement of the ABCG37 transporter in secretion of scopoletin and derivatives by Arabidopsis roots in response to iron deficiency. New Phytol. 2014, 201, 155–167. [Google Scholar] [CrossRef]
- Ruzicka, K.; Strader, L.C.; Bailly, A.; Yang, H.; Blakeslee, J.; Langowski, L.; Nejedla, E.; Fujita, H.; Itoh, H.; Syono, K.; et al. Arabidopsis PIS1 encodes the ABCG37 transporter of auxinic compounds including the auxin precursor indole-3-butyric acid. Proc. Natl. Acad. Sci. USA 2010, 107, 10749–10753. [Google Scholar] [CrossRef]
- Matsuda, S.; Takano, S.; Sato, M.; Furukawa, K.; Nagasawa, H.; Yoshikawa, S.; Kasuga, J.; Tokuji, Y.; Yazaki, K.; Nakazono, M.; et al. Rice stomatal closure requires guard cell plasma membrane ATP-binding cassette transporter RCN1/OsABCG5. Mol. Plant 2016, 9, 417–427. [Google Scholar] [CrossRef]
- Mentewab, A.; Matheson, K.; Adebiyi, M.; Robinson, S.; Elston, B. RNA-seq analysis of the effect of kanamycin and the ABC transporter AtWBC19 on Arabidopsis thaliana seedlings reveals changes in metal content. PLoS ONE 2014, 9, e109310. [Google Scholar] [CrossRef] [PubMed]
- Mentewab, A.; Stewart, C.N., Jr. Overexpression of an Arabidopsis thaliana ABC transporter confers kanamycin resistance to transgenic plants. Nat. Biotechnol. 2005, 23, 1177–1180. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.N.T.; Lee, S.B.; Suh, M.C.; An, G.; Jung, K.-H. OsABCG9 is an important abc transporter of cuticular wax deposition in rice. Front. Plant Sci. 2018, 9, 960. [Google Scholar] [CrossRef] [PubMed]
- Oda, K.; Otani, M.; Uraguchi, S.; Akihiro, T.; Fujiwara, T. Rice ABCG43 is cd inducible and confers cd tolerance on yeast. Biosci. Biotechnol. Biochem. 2011, 75, 1211–1213. [Google Scholar] [CrossRef] [PubMed]
- Badri, D.V.; Loyola-Vargas, V.M.; Broeckling, C.D.; De-la-Pena, C.; Jasinski, M.; Santelia, D.; Martinoia, E.; Sumner, L.W.; Banta, L.M.; Stermitz, F.; et al. Altered profile of secondary metabolites in the root exudates of Arabidopsis ATP-binding cassette transporter mutants. Plant Physiol. 2008, 146, 762–771. [Google Scholar] [CrossRef]
- Kim, D.Y.; Bovet, L.; Maeshima, M.; Martinoia, E.; Lee, Y. The ABC transporter AtPDR8 is a cadmium extrusion pump conferring heavy metal resistance. Plant J. 2007, 50, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Jin, J.Y.; Alejandro, S.; Martinoia, E.; Lee, Y. Overexpression of AtABCG36 improves drought and salt stress resistance in Arabidopsis. Physiol. Plant. 2010, 139, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.; Dittgen, J.; Sanchez-Rodriguez, C.; Hou, B.H.; Molina, A.; Schulze-Lefert, P.; Lipka, V.; Somerville, S. Arabidopsis PEN3/PDR8, an ATP binding cassette transporter, contributes to nonhost resistance to inappropriate pathogens that enter by direct penetration. Plant Cell 2006, 18, 731–746. [Google Scholar] [CrossRef]
- Strader, L.C.; Bartel, B. The Arabidopsis PLEIOTROPIC DRUG RESISTANCE8/ABCG36 ATP binding cassette transporter modulates sensitivity to the auxin precursor indole-3-butyric acid. Plant Cell 2009, 21, 1992–2007. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Lu, Y.; Zhang, X.; Yang, G.; Chao, D.; Wang, Z.; Shi, M.; Chen, J.; Chao, D.Y.; Li, R.; et al. The ABC transporter ABCG36 is required for cadmium tolerance in rice. J. Exp. Bot. 2019, 70, 5909–5918. [Google Scholar] [CrossRef]
- Kang, J.; Hwang, J.U.; Lee, M.; Kim, Y.Y.; Assmann, S.M.; Martinoia, E.; Lee, Y. PDR-type ABC transporter mediates cellular uptake of the phytohormone abscisic acid. Proc. Natl. Acad. Sci. USA 2010, 107, 2355–2360. [Google Scholar] [CrossRef] [PubMed]
- Shimadzu, S.; Seo, M.; Terashima, I.; Yamori, W. Whole irradiated plant leaves showed faster photosynthetic induction than individually irradiated leaves via improved stomatal opening. Front. Plant Sci. 2019, 10, 1512. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xu, J.; Wang, X.; He, X.; Wang, Y.; Zhou, J.; Zhang, S.; Meng, X. The Arabidopsis pleiotropic drug resistance transporters PEN3 and PDR12 mediate camalexin secretion for resistance to Botrytis cinerea. Plant Cell 2019, 31, 2206–2222. [Google Scholar] [CrossRef]
- Gutjahr, C.; Radovanovic, D.; Geoffroy, J.; Zhang, Q.; Siegler, H.; Chiapello, M.; Casieri, L.; An, K.; An, G.; Guiderdoni, E.; et al. The half-size abc transporters STR1 and STR2 are indispensable for mycorrhizal arbuscule formation in rice. Plant J. 2012, 69, 906–920. [Google Scholar] [CrossRef]
- Ko, D.; Kang, J.; Kiba, T.; Park, J.; Kojima, M.; Do, J.; Kim, K.Y.; Kwon, M.; Endler, A.; Song, W.Y.; et al. Arabidopsis ABCCG14 is essential for the root-to-shoot translocation of cytokinin. Proc. Natl. Acad. Sci. USA 2014, 111, 7150–7155. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yu, N.; Ju, M.; Fan, B.; Zhang, Y.; Zhu, E.; Zhang, M.; Zhang, K. Abc transporter OsABCG18 controls the shootward transport of cytokinins and grain yield in rice. J. Exp. Bot. 2019, 70, 6277–6291. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Kilambi, H.V.; Liu, J.; Bar, H.; Lazary, S.; Egbaria, A.; Ripper, D.; Charrier, L.; Belew, Z.M.; Wulff, N.; et al. ABC homeostasis and long-distance translocation are redundantly regulated by ABCG ABC importers. Sci. Adv. 2021, 7, eabf6069. [Google Scholar] [CrossRef] [PubMed]
- Kuromori, T.; Sugimoto, E.; Shinozaki, K. Arabidopsis mutants of AtABCG22, an ABC transporter gene, increase water transpiration and drought susceptibility. Plant J. 2011, 67, 885–894. [Google Scholar] [CrossRef]
- Kuromori, T.; Miyaji, T.; Yabuuchi, H.; Shimizu, H.; Sugimoto, E.; Kamiya, A.; Moriyama, Y.; Shinozaki, K. ABC transporter AtABCG25 is involved in abscisic acid transport and responses. Proc. Natl. Acad. Sci. USA 2010, 107, 2361–2366. [Google Scholar] [CrossRef]
- Liu, C.; Li, Z.; Tian, D.; Xu, M.; Pan, J.; Wu, H.; Wang, C.; Otegui, M.S. AP1/2 beta-mediated exocytosis of tapetum-specific transporters is required for pollen development in Arabidopsis thaliana. Plant Cell 2022, koac192. [Google Scholar] [CrossRef]
- Borghi, L.; Kang, J.; Ko, D.; Lee, Y.; Martinoia, E. The role of ABCG-type ABC transporters in phytohormone transport. Biochem. Soc. Trans. 2015, 43, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, Z.; Wu, S.; Wan, X. The essential roles of sugar metabolism for pollen development and male fertility in plants. Crop J. 2021, 9, 1223–1236. [Google Scholar] [CrossRef]
- Wan, X.; Wu, S.; Li, Z.; An, X.; Tian, Y. Lipid metabolism: Critical roles in male fertility and other aspects of reproductive development in plants. Mol. Plant 2020, 13, 955–983. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Wu, S.; Li, Z.; Dong, Z.; An, X.; Ma, B.; Tian, Y.; Li, J. Maize genic male-sterility genes and their applications in hybrid breeding: Progress and perspectives. Mol. Plant 2019, 12, 321–342. [Google Scholar] [CrossRef]
- Jiang, Y.; An, X.; Li, Z.; Yan, T.; Zhu, T.; Xie, K.; Liu, S.; Hou, Q.; Zhao, L.; Wu, S.; et al. CRISPR/Cas9-based discovery of maize transcription factors regulating male sterility and their functional conservation in plants. Plant Biotechnol. J. 2021, 19, 1769–1784. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | New Name | Synonyms 1 | Gene Models (B73 V4) | Chr. | Length (aa) | Expression Organs 2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Root | Meristem | Leave | Internode | Tassel | Anther | Cob | Silk | Seed | Embryo | ||||||
| 1 | ZmABCG1 | ABC-2 | Zm00001d027723 | 1 | 753 | + | + | ||||||||
| 2 | ZmABCG2 | ABCG2 | Zm00001d028689 | 1 | 790 | + | + | ||||||||
| 3 | ZmABCG3 | ABCG11 | Zm00001d032601 | 1 | 736 | + | + | + | + | + | + | + | |||
| 4 | ZmABCG4 | ABCG24 | Zm00001d032855 | 1 | 883 | + | + | + | + | + | + | + | + | + | |
| 5 | ZmABCG5 | ABCG3 | Zm00001d034918 | 1 | 1078 | + | + | + | + | + | + | + | + | + | |
| 6 | ZmABCG6 | ABCG11 | Zm00001d002871 | 2 | 721 | + | + | + | + | + | + | ||||
| 7 | ZmABCG7 | ABCG25 | Zm00001d004762 | 2 | 632 | + | + | + | + | ||||||
| 8 | ZmABCG8 | ABCG6 | Zm00001d042953 | 3 | 761 | + | + | + | |||||||
| 9 | ZmABCG9 | ABCG25 | Zm00001d043722 | 3 | 738 | + | + | + | |||||||
| 10 | ZmABCG10 | ABCG10 | Zm00001d044476 | 3 | 609 | + | + | + | |||||||
| 11 | ZmABCG11 | ABCG11 | Zm00001d048621 | 4 | 720 | + | + | + | + | ||||||
| 12 | ZmABCG12 | - | Zm00001d053049 | 4 | 636 | + | + | + | + | + | + | + | |||
| 13 | ZmABCG13 | ABCG2 | Zm00001d013960 | 5 | 710 | + | + | + | + | + | + | + | + | ||
| 14 | ZmABCG14 | ABCG24 | Zm00001d018500 | 5 | 1112 | + | |||||||||
| 15 | ZmABCG15 | ABCG16 | Zm00001d035553 | 6 | 714 | + | + | + | + | + | + | + | + | ||
| 16 | ZmABCG16 | - | Zm00001d038068 | 6 | 694 | + | + | + | |||||||
| 17 | ZmABCG17 | - | Zm00001d019398 | 7 | 693 | + | + | + | + | ||||||
| 18 | ZmABCG18 | ABCG11 | Zm00001d020811 | 7 | 736 | + | + | + | |||||||
| 19 | ZmABCG19 | ABC-2 | Zm00001d008512 | 8 | 695 | + | + | + | + | + | + | + | + | ||
| 20 | ZmABCG20 | - | Zm00001d009492 | 8 | 275 | + | + | + | + | + | + | + | |||
| 21 | ZmABCG21 | - | Zm00001d010426 | 8 | 690 | + | + | + | + | + | + | + | |||
| 22 | ZmABCG22 | - | Zm00001d011006 | 8 | 712 | + | + | + | + | + | + | + | + | + | |
| 23 | ZmABCG23 | ABCG10 | Zm00001d011350 | 8 | 507 | + | + | ||||||||
| 24 | ZmABCG24 | ABCG16 | Zm00001d012438 | 8 | 743 | + | + | + | |||||||
| 25 | ZmABCG25 | - | Zm00001d044895 | 9 | 618 | + | + | + | + | + | + | + | + | + | |
| 26 | ZmABCG26 | ABCG28 | Zm00001d046068 | 9 | 1040 | + | |||||||||
| 27 | ZmABCG27 | ms2 | Zm00001d046537 | 9 | 676 | + | + | + | + | + | + | + | + | ||
| 28 | ZmABCG28 | - | Zm00001d024075 | 10 | 892 | + | + | + | |||||||
| 29 | ZmABCG29 | - | Zm00001d024497 | 10 | 727 | + | + | + | + | + | + | + | |||
| 30 | ZmABCG30 | ABCG24 | Zm00001d025031 | 10 | 1101 | + | + | + | + | + | + | ||||
| 31 | ZmABCG31 | - | Zm00001d026706 | 10 | 750 | + | + | + | + | + | + | + | + | + | |
| 32 | ZmABCG32 | - | Zm00001d031724 | 1 | 1357 | + | + | + | |||||||
| 33 | ZmABCG33 | - | Zm00001d034940 | 1 | 1490 | + | + | + | + | + | + | + | + | ||
| 34 | ZmABCG34 | ABCG51 | Zm00001d003354 | 2 | 1425 | + | + | + | + | ||||||
| 35 | ZmABCG35 | glossy13 | Zm00001d039631 | 3 | 1425 | + | + | + | + | + | + | ||||
| 36 | ZmABCG36 | ABCG38 | Zm00001d043598 | 3 | 1488 | + | + | + | |||||||
| 37 | ZmABCG37 | - | Zm00001d044442 | 3 | 1574 | + | + | + | + | + | + | ||||
| 38 | ZmABCG38 | ABCG37 | Zm00001d044443 | 3 | 1448 | + | + | + | |||||||
| 39 | ZmABCG39 | ABCG39 | Zm00001d048860 | 4 | 1446 | + | + | + | + | + | + | ||||
| 40 | ZmABCG40 | ABCG45 | Zm00001d052726 | 4 | 1237 | + | |||||||||
| 41 | ZmABCG41 | ABCG39 | Zm00001d053612 | 4 | 1443 | + | + | + | + | ||||||
| 42 | ZmABCG42 | ABCG42 | Zm00001d036986 | 6 | 1508 | + | + | + | + | ||||||
| 43 | ZmABCG43 | ABCG45 | Zm00001d019951 | 7 | 1224 | + | |||||||||
| 44 | ZmABCG44 | - | Zm00001d019961 | 7 | 1056 | + | + | + | |||||||
| 45 | ZmABCG45 | ABCG53 | Zm00001d020134 | 7 | 1451 | + | + | + | |||||||
| 46 | ZmABCG46 | ABCG43 | Zm00001d021647 | 7 | 1445 | + | + | + | |||||||
| 47 | ZmABCG47 | ABCG36 | Zm00001d011315 | 8 | 1439 | + | + | + | + | + | + | ||||
| 48 | ZmABCG48 | ABCG37 | Zm00001d011319 | 8 | 1449 | + | |||||||||
| 49 | ZmABCG49 | ABCG42 | Zm00001d046277 | 9 | 1540 | + | + | + | + | ||||||
| 50 | ZmABCG50 | ABCG41 | Zm00001d023392 | 10 | 1472 | + | |||||||||
| 51 | ZmABCG51 | ABCG48 | Zm00001d025012 | 10 | 1465 | + | + | + | + | ||||||
| No. | Gene Name | Gene ID | Maize Ortholog | Expression Organ | Biological Function | Reference |
|---|---|---|---|---|---|---|
| I. Anther development and male fertility | ||||||
| 1 | AtABCG11 | AT1G17840 | ZmABCG13 ZmABCG3 ZmABCG6 | Stems; Leaves; Roots; Inflorescences; Flowers; Seeds; Siliques | Flower cutin and root suberin formation; Vascular development. | [17,25,26] |
| 2 | OsABCG26 | Os10g0494300 | ZmABCG13 ZmABCG3 | Anthers; Pistils | Anther cuticle and pollen exine formation and pollen-pistil interaction | [18,19] |
| 3 | ZmMs13/ ZmABCG13 * | Zm00001d013960 | - | Anthers | Callose dissolution; Anther cuticle and pollen exine formation | [7] |
| 4 | AtABCG26 | AT3G13220 | ZmABCG27 | Anthers; Siliques; Leaves; Stems | Exine formation | [12,13,14] |
| 5 | OsABCG15 | Os06g0607700 | ZmABCG27 | Anthers | Exine formation and pollen development | [15,16,19] |
| 6 | ZmMs2/ ZmABCG27 ** | Zm00001d046537 | - | Anthers | Transport of anther cutin and wax components | [8,10] |
| 7 | AtABCG1 | At2G39350 | ZmABCG2 | Roots; Flowers; Stamens; Pistils; Seedlings; Leaves; Shoot meristems | Male gametophyte development; Pollen tube growth; Suberin formation in roots. | [20,21,22,27] |
| 8 | AtABCG16 | AT3G55090 | ZmABCG2 | Cotyledons; Roots; Seeds; Flowers; Leaves | Male gametophyte development and pollen tube growth; Anther filament; JA and ABA responses; Plant pathogen response. | [20,21,22,28,29] |
| 9 | OsABCG3/ LSP1 | Os01g0836600 | ZmABCG24 ZmABCG8 | Anthers | Normal pollen fertility and pollen wall formation | [23,24] |
| 10 | AtABCG9 | AT4G27420 | ZmABCG29 | Cotyledons; Leaves; Stamen; Siliques; Roots; Anthers | Sterol accumulated on pollen surface; Proper vascular development | [11,25] |
| 11 | AtABCG31 | AT2G29940 | ZmABCG34 | Inflorescences; Anthers; Rosette leaves; Stems; Seedlings; Siliques | Sterol accumulation on pollen surface; ABA export from endosperm; Seed germination; Disease response | [11,30] |
| 12 | AtABCG28 | AT5G60740 | ZmABCG26 ZmABCG14 | Pollen tubes; Pollen grains | Polyamine and ROS translocation at the growing tip of pollen tube | [31] |
| II. Vegetative and female organ development | ||||||
| 1 | AtABCG2 | AT2G37360 | ZmABCG2 | Roots; Seedlings; Anthers | Suberin barrier synthesis in roots and seed coats | [21] |
| 2 | AtABCG6 | AT5G13580 | ZmABCG2 | Roots; Seeds; Anthers | Suberin barrier synthesis in roots and seed coats | [21] |
| 3 | AtABCG20/ AtAwake1 | AT3G53510 | ZmABCG2 | Roots; Seedings and anthers | Suberin barrier synthesis in roots and seed coats; Seed dormancy | [21,32] |
| 4 | AtABCG5 | AT2G13610 | ZmABCG31 | Cotyledons; Roots; Shoots | Wax precursor transport; Shoot branching; Root suberization | [33,34,35] |
| 5 | AtABCG12 | AT1G51500 | ZmABCG21 | Stems; Leaves; Siliques; Flowers; Roots | Cuticle formation; Wax secretion; Abiotic stress response | [26,36] |
| 6 | AtABCG13 | AT1G51460 | ZmABCG21 | Flowers; Leaves; Stems; Siliques; Roots | Flower cuticular lipids transport | [37,38] |
| 7 | AtABCG21 | AT3G25620 | ZmABCG29 | Seedlings; Leaves | Stomatal regulation | [39] |
| 8 | AtABCG27 | AT3G52310 | ZmABCG1 | Flowers; Pistils; Leaves | Cellulose synthesis; Flower and leaf development | [40,41] |
| 9 | AtABCG29 | AT3G16340 | ZmABCG36 | Roots; Stems; Leaves; Anthers; Siliques; Seedings | Lignin biosynthesis | [40,42] |
| 10 | AtABCG32 | AT2G26910 | ZmABCG35 | Leaves; Stems; Flowers; Seedlings; Siliques | Cuticular layer of the cell wall formation | [43,44] |
| 11 | OsABCG31 | Os01g0177900 | ZmABCG35 | Leaves | Cuticle formation | [45] |
| 12 | AtABCG33 | AT2G37280 | ZmABCG50 | Stems; Roots | Cellulose synthesis; Lignification | [40,46] |
| 13 | AtABCG34 | AT2G36380 | ZmABCG46 ZmABCG33 ZmABCG51 | Roots; Leaves | Camalexin secretion to leaf surface and thereby prevents A. brassicicola infection; Root exudation | [47] |
| 14 | AtABCG37 | AT3G53480 | ZmABCG50 | Roots; Seedlings | IBA transported out of the cells; Secretion of scopoletin and derivatives | [48,49] |
| 15 | ZmGL13 | Zm00001d039631 | - | Leaves | Necrotic glossy leaf; Plants smaller than nonmutant sibs | [9] |
| III. Biotic and abiotic stress response | ||||||
| 1 | OsABCG5/ RCN1 | Os03g0281900 | ZmABCG2 | Roots; Tiller buds; Basal part of stem; Leaves. | Lateral shoot outgrowth; ABA accumulation in guard cells under drought stress; Root hypodermis suberization | [34,35,50] |
| 2 | AtABCG19 | AT3G55130 | ZmABCG2 | Leaves; Roots; Flowers; Seedings | Kanamycin resistance; Zinc homeostasis; Nicotianamine transport | [51,52] |
| 3 | OsABCG9 | Os04g0528300 | ZmABCG6 | Seminal roots; Young shoots; Anthers; Stems | Cuticular permeability and drought sensitivity | [53] |
| 4 | OsABCG43 | Os07g0522500 | ZmABCG46 | Roots | Cd tolerance in yeast | [54] |
| 5 | AtABCG35 | AT1G15210 | ZmABCG36 | Roots | Root exudation; Cadmium response | [55,56] |
| 6 | AtABCG36 | AT1G59870 | ZmABCG36 | Roots; Leaves; hydathodes; Stems; Inflorescences; Flowers; Siliques | An efflux pump of Cd2+ or Cd conjugates; Drought and salt resistance; IBA Transport; Root and cotyledon development | [57,58,59] |
| 7 | OsABCG36 | Os01g0609300 | ZmABCG36 | Roots; Shoots | Cadmium tolerance; Heavy metal stress | [60] |
| 8 | AtABCG40 | AT1G15520 | ZmABCG41/ ZmABCG39 | Inflorescences; Flowers; Roots; Leaves; Stems; Seeds; Siliques | Stomatal regulation and ABA importation; Seed germination; Pathogen and drought response | [30,61,62,63] |
| 9 | OsSTR1 | Os09g0401100 | ZmABCG28 | Roots; Panicles; Leaves; Stems; Embryos | Mycorrhizal arbuscule formation | [64] |
| 10 | OsSTR2 | Os07g0191600 | - | |||
| IV. Hormone transport and signaling | ||||||
| 1 | AtABCG14 | AT1G31770 | ZmABCG29 | Cotyledons; Rosette leaves; Flowers; Roots; Siliques | Vascular development; Cytokinin translocation in shoot | [25,65] |
| 2 | OsABCG18 | Os08g0167000 | ZmABCG29 | Roots; Stems; Leaves; Panicles | Long-distance transport of cytokinin in shoot and promote grain yield | [66] |
| 3 | AtABCG17 | AT3G55100 | ZmABCG2 | Roots; Leaves | ABA homeostasis and long-distance translocation | [67] |
| 4 | AtABCG18 | AT3G55110 | ||||
| 5 | AtABCG22 | AT5G06530 | ZmABCG1 | Seedlings; Roots; Stems; Leaves; Flowers | Stomatal regulation; ABA signaling; Lignification | [39,40,68] |
| 6 | AtABCG25 | AT1G71960 | ZmABCG12/ ZmABCG7 | Seedlings; Roots; Stems; Leaves; Flowers | Stomatal regulation; ABA export from endosperm; Seed germination | [30,62,69] |
| 7 | AtABCG30 | AT4G15230 | ZmABCG50 | Roots; Seeds | ABA import into the embryo; Seed germination; Monolignol transport | [30,55] |
| No. | Transporters | Types | Substrate(s) | Method of Substrate Identification | Reference |
|---|---|---|---|---|---|
| I. Isotope labelling experiment in vivo | |||||
| 1 | AtABCG11 | Half-size | 10,16-diOH C16:0-2-glycerol; ω-OH C16:0 | [3H]-10,16-diOH C16:0-2-glycerol; [14C]-u-OH C16:0 export assay using N. benthamiana protoplast system | [44] |
| 2 | AtABCG14 | Half-size | Cytokinins | WT and atabcg14 mutant cultivated in 14C-labeled trans-zeatin (tZ) medium | [65] |
| 3 | AtABCG16 | Half-size | Jasmonate | 3H-JA transport of yeast strain expressing AtABCG16; transport of 3H-JA and 3H-JA-Ile by the nuclei isolated from abcg16 plant | [28] |
| 4 | AtABCG25 | Half-size | ABA | 3H-ABA transport of yeast strains expressing AtABCG25 | [30,69] |
| 5 | AtABCG30 | Full-size | ABA | 3H-ABA transport of yeast strains expressing AtABCG30 | [30] |
| 6 | AtABCG31 | Full-size | ABA | 3H-ABA transport of yeast strains expressing AtABCG31 | [30] |
| 7 | AtABCG32 | Full-size | 10,16-diOH C16:0-2-glycerol; ω-OH C16:0; C16:0 DCA | [3H]-10,16-diOH C16:0-2-glycerol; [14C]-u-OH C16:0; [14C]-C16:0 DCA export assay using N. benthamiana protoplast system | [44] |
| 8 | AtABCG37 | Full-size | Auxin precursor indole-3-butyric acid (IBA) | 3H-IBA export from abcg37 leaf mesophyll protoplasts; 3H-IBA transport in yeast strains expressing AtABCG37; export of 3H-2,4-D and 3H-IBA in HeLa cells expressing AtABCG37 | [49] |
| 9 | AtABCG40 | Full-size | ABA | 3H-ABA uptake in yeast strain YMM12 and BY-2 cell lines expressing AtABCG40 | [30] |
| II. Substance analysis using the GC-MS system | |||||
| 1 | AtABCG2 | Half-size | Suberin precursors | Suberin monomers analysis by using GC-MS system | [21] |
| 2 | OsABCG5/ RCN1 | Half-size | ABA; suberin monomers | Phenotypic and expression analysis of RCN1 in WT and RCN1-RNAi plants treated with ABA; Histochemical staining and suberin contents analysis by GC-MS | [34,35,50] |
| 3 | AtABCG5 | Half-size | Cutin/wax precursors | Substance analysis by using GC-MS | [33] |
| 4 | AtABCG6 | Half-size | Fatty acids; Fatty alcohols | Suberin monomers analysis by using GC-MS | [21] |
| 5 | AtABCG9 | Half-size | Steryl glycosides | Substance analysis by using GC-MS | [11] |
| 6 | OsABCG9 | Half-size | Wax precursors | Substance analysis by using GC-MS | [53] |
| 7 | OsABCG26 | Half-size | Anther cuticular wax and cutin monomers | Substance analysis by using GC-MS | [18,19] |
| 8 | ZmMS13/ ZmABCG13 | Half-size | Anther cuticular wax and cutin monomers | Substance analysis by using GC-MS | [7] |
| 9 | AtABCG12 | Half-size | Lipids; Wax components | Substance analysis by using GC-MS | [26,36] |
| 10 | AtABCG13 | Half-size | Cuticular lipids | Substance analysis by using GC-MS | [37] |
| 11 | AtABCG20/ Awake1 | Half-size | Suberin precursors; Fatty acids | Substance analysis by using GC-MS | [21,32] |
| 12 | AtABCG26 | Half-size | Sporopollenin precursors; Polyketides | Substance analysis by using GC-MS | [12,14] |
| 13 | OsABCG15 | Half-size | Sporopollenin and cutin precursors | Substance analysis by using GC-MS | [15,16] |
| 14 | ZmMS2/ ABCG26 | Half-size | Sporopollenin and cutin precursors | Substance analysis by using GC-MS | [8,10] |
| 15 | OsABCG31 | Full-size | Cutin precursors | Substance analysis by using GC-MS | [45] |
| III. Other methods | |||||
| 1 | AtABCG1 | Half-size | Fatty alcohols; Fatty acids | AtABCG1 protein purified in Pichia pastoris to test its ATPase assay | [27] |
| 2 | ZmMS13/ ZmABCG13 | Half-size | Fatty alcohols; Fatty acids | ZmMS13 protein purified in E. coli and its ATPase assay in vitro | [7] |
| 3 | OsABCG18 | Half-size | Cytokinins | Export assay with OsABCG18 expressing N. benthamiana protoplasts | [66] |
| 4 | OsABCG3/ LSP1 | Half-size | Pollen wall and coat materials | Phenotypic and qRT-PCR analysis of WT and osabcg3 mutant | [23,24] |
| 5 | AtABCG19 | Half-size | Kanamycin | Kanamycin treatment of AtABCG19-overexpressing plants | [52] |
| 6 | AtABCG22 | Half-size | ABA; lignin precursors | Speculation according to the same phenotype with AtABCG21 | [40,68] |
| 7 | AtABCG28 | Half-size | Polyamine | Immunostaining of polyamines in pollen tubes | [31] |
| 8 | AtABCG29 | Full-size | p-coumaryl alcohol | p-coumaryl alcohol uptake using microsomes from S. cerevisiae expressing AtABCG29 | [42] |
| 9 | AtABCG34 | Full-size | Camalexin | Camalexin toxicity assay in Arabidopsis and BY2 cells expressing AtABCG34 | [47] |
| 10 | OsABCG43 | Full-size | Cd | Cd inducible and confers Cd tolerance on OsABCG43-expressing yeast cell. | [54] |
| 11 | AtABCG36 | Full-size | Lignin precursors; Cd; IBA | The atabcg36 mutants and WT were tested in IBA-containing agar medium under yellow filtered light conditions. | [57,59] |
| 12 | OsABCG36 | Full-size | Cd2+ | Efflux transport of Cd in OsABCG36-expressing yeast cell. | [60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Fang, C.; Li, Z.; Wang, Y.; Pan, S.; Wu, Y.; An, X.; Long, Y.; Wan, X. ATP-Binding Cassette G Transporters and Their Multiple Roles Especially for Male Fertility in Arabidopsis, Rice and Maize. Int. J. Mol. Sci. 2022, 23, 9304. https://doi.org/10.3390/ijms23169304
Wu S, Fang C, Li Z, Wang Y, Pan S, Wu Y, An X, Long Y, Wan X. ATP-Binding Cassette G Transporters and Their Multiple Roles Especially for Male Fertility in Arabidopsis, Rice and Maize. International Journal of Molecular Sciences. 2022; 23(16):9304. https://doi.org/10.3390/ijms23169304
Chicago/Turabian StyleWu, Suowei, Chaowei Fang, Ziwen Li, Yanbo Wang, Shuangshuang Pan, Yuru Wu, Xueli An, Yan Long, and Xiangyuan Wan. 2022. "ATP-Binding Cassette G Transporters and Their Multiple Roles Especially for Male Fertility in Arabidopsis, Rice and Maize" International Journal of Molecular Sciences 23, no. 16: 9304. https://doi.org/10.3390/ijms23169304
APA StyleWu, S., Fang, C., Li, Z., Wang, Y., Pan, S., Wu, Y., An, X., Long, Y., & Wan, X. (2022). ATP-Binding Cassette G Transporters and Their Multiple Roles Especially for Male Fertility in Arabidopsis, Rice and Maize. International Journal of Molecular Sciences, 23(16), 9304. https://doi.org/10.3390/ijms23169304