
Identification of a Novel Lipase with AHSMG Pentapeptide in Hypocreales and Glomerellales Filamentous Fungi

 , and
, and
Abstract
:1. Introduction
2. Results
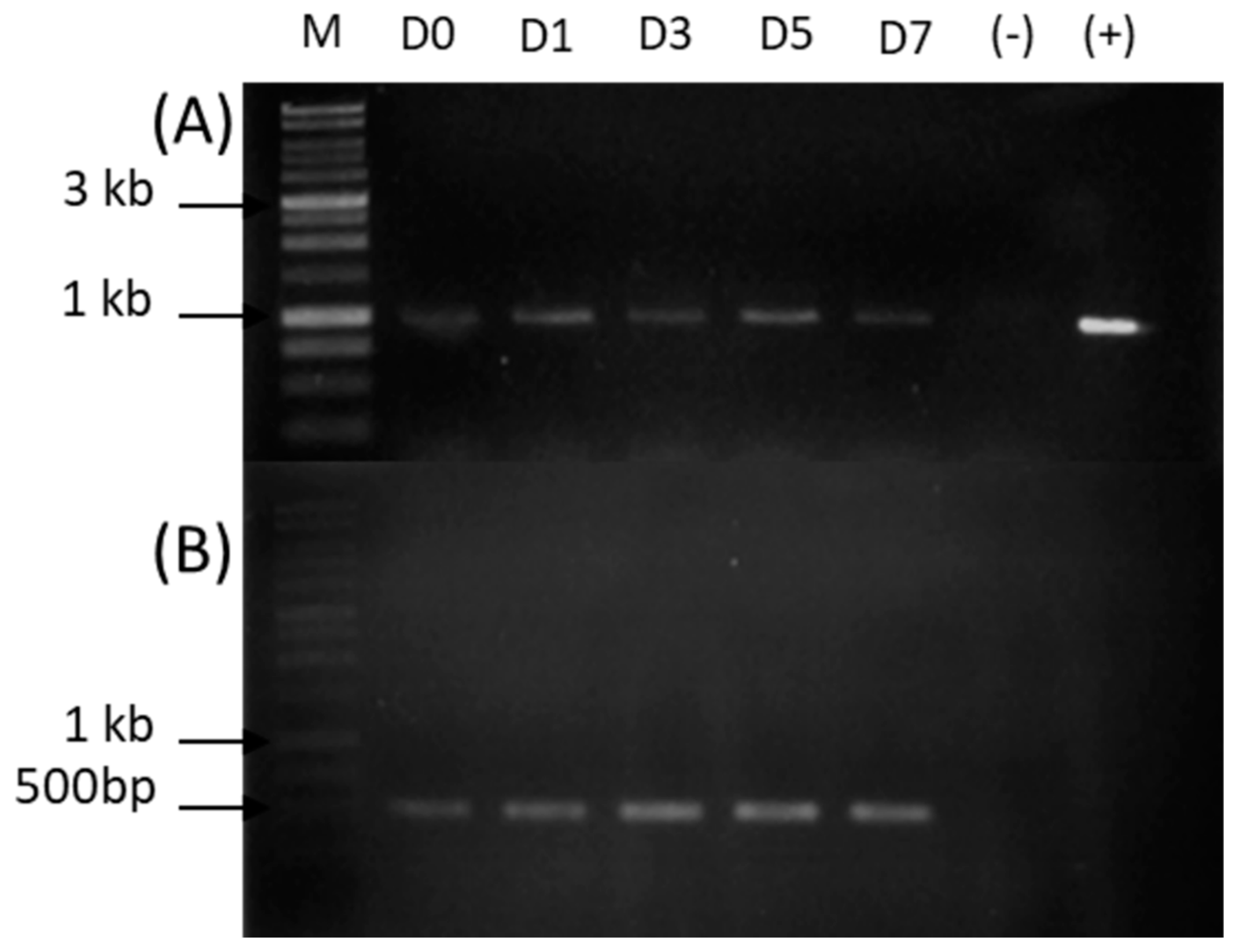
2.1. Expression of Lipase 135964 in T. harzianum
2.2. Sequencing
2.3. In Silico Characterization
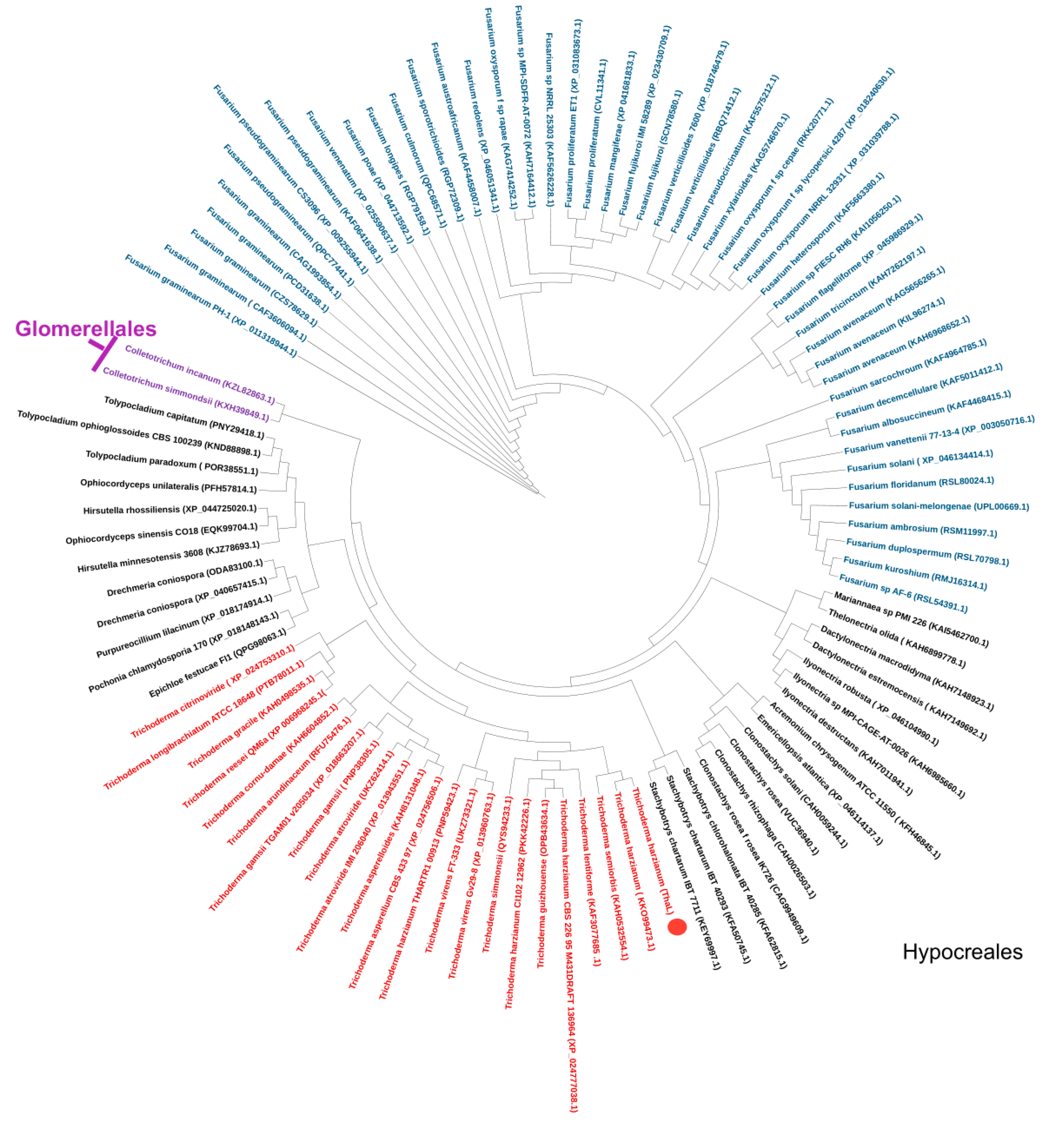
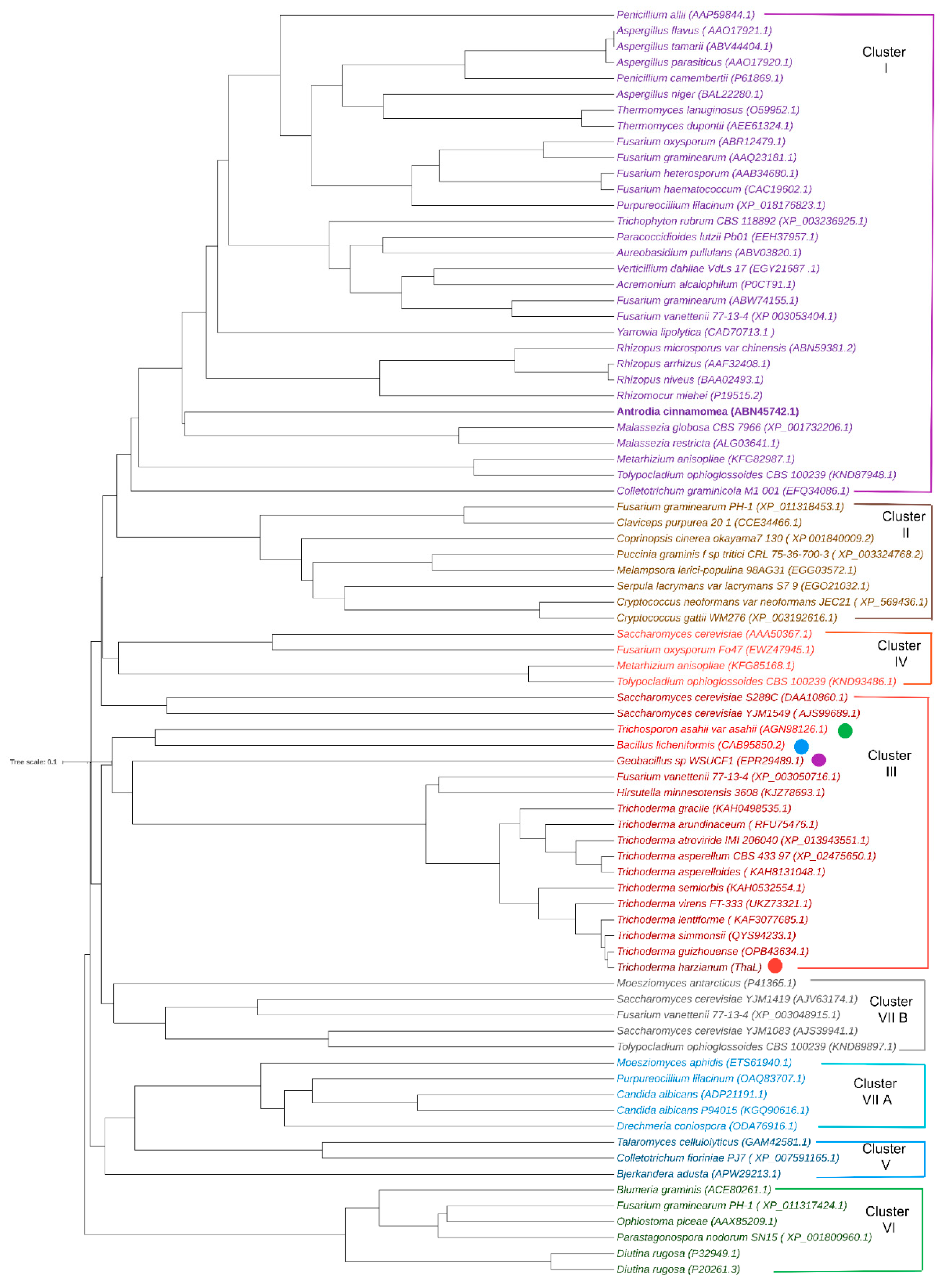
2.4. Orthologs and the Phylogenetic Tree
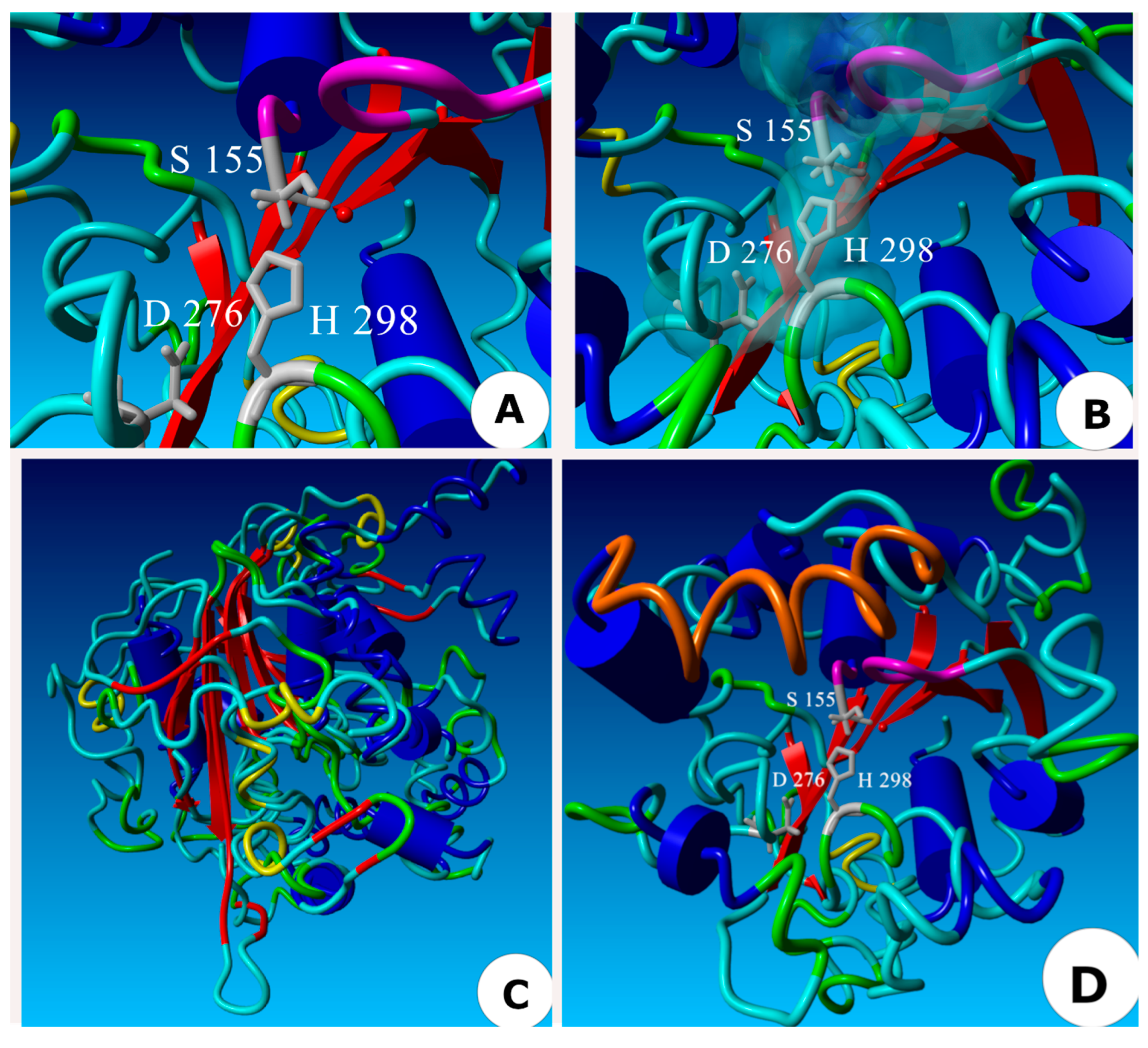
2.5. Three-Dimensional Model of the ThaL Lipase
2.6. Pathogenicity Induction by Starvation
3. Discussion
4. Materials and Methods
4.1. Biological Material
4.2. Pathogenicity Induction by Starvation
4.3. RNA Extraction and cDNA Synthesis
4.4. Cloning and Sequencing
4.5. Expression Analysis
4.6. Bioinformatics
4.6.1. In Silico Characterization
4.6.2. Ortholog Identification and Phylogenetic Analysis
4.6.3. 3D Modeling
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Casas-Godoy, L.; Gasteazoro, F.; Duquesne, S.; Bordes, F.; Marty, A.; Sandoval, G. Lipases: An Overview. In Lipases and Phospholipases; Sandoval, G., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2018; Volume 1835, pp. 3–38. [Google Scholar] [CrossRef]
- Negi, S. Lipases: A Promising Tool for Food Industry. In Green Bio-Processes; Parameswaran, B., Varjani, S., Raveendran, S., Eds.; Energy, Environment, and Sustainability; Springer: Singapore, 2019; pp. 181–198. [Google Scholar] [CrossRef]
- Gupta, R.; Kumari, A.; Syal, P.; Singh, Y. Molecular and Functional Diversity of Yeast and Fungal Lipases: Their Role in Biotechnology and Cellular Physiology. Prog. Lipid Res. 2015, 57, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Pérez, M.M.; Gonçalves, E.C.S.; Vici, A.C.; Salgado, J.C.S.; de Moraes Polizeli, M.D.L.T. Fungal Lipases: Versatile Tools for White Biotechnology. In Recent Advancement in White Biotechnology through Fungi; Yadav, A.N., Mishra, S., Singh, S., Gupta, A., Eds.; Fungal Biology; Springer International Publishing: Cham, Switzerland, 2019; pp. 361–404. [Google Scholar] [CrossRef]
- Hulo, N. The PROSITE Database. Nucleic Acids Res. 2006, 34, D227–D230. [Google Scholar] [CrossRef] [PubMed]
- Arpigny, J.L.; Jaeger, K.-E. Bacterial Lipolytic Enzymes: Classification and Properties. Biochem. J. 1999, 343, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Meghwanshi, G.K.; Kumar, R. Current Perspectives for Microbial Lipases from Extremophiles and Metagenomics. Biochimie 2021, 182, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Kovacic, F.; Babic, N.; Krauss, U.; Jaeger, K.-E. Classification of Lipolytic Enzymes from Bacteria. In Aerobic Utilization of Hydrocarbons, Oils, and Lipids; Rojo, F., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 255–289. [Google Scholar] [CrossRef]
- Hausmann, S.; Jaeger, K.-E. Lipolytic Enzymes from Bacteria. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1099–1126. [Google Scholar] [CrossRef]
- Fischer, M. The Lipase Engineering Database: A Navigation and Analysis Tool for Protein Families. Nucleic Acids Res. 2003, 31, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Bai, Q.; Cai, Y.; Yang, L.; Wu, Q.; Su, M.; Tang, L. Substitution of L133 with Methionine in GXSXG Domain Significantly Changed the Activity of Penicillium Expansum Lipase. Catal. Lett. 2022, 152, 2047–2055. [Google Scholar] [CrossRef]
- Hosseini, M.; Karkhane, A.A.; Yakhchali, B.; Shamsara, M.; Aminzadeh, S.; Morshedi, D.; Haghbeen, K.; Torktaz, I.; Karimi, E.; Safari, Z. In Silico and Experimental Characterization of Chimeric Bacillus Thermocatenulatus Lipase with the Complete Conserved Pentapeptide of Candida Rugosa Lipase. Appl. Biochem. Biotechnol. 2013, 169, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Kanjanavas, P.; Khuchareontaworn, S.; Khawsak, P.; Pakpitcharoen, A.; Pothivejkul, K.; Santiwatanakul, S.; Matsui, K.; Kajiwara, T.; Chansiri, K. Purification and Characterization of Organic Solvent and Detergent Tolerant Lipase from Thermotolerant Bacillus Sp. RN2. Int. J. Mol. Sci. 2010, 11, 3783–3792. [Google Scholar] [CrossRef]
- Sangeetha, R.; Arulpandi, I.; Geetha, A. Molecular Characterization of a Proteolysis-Resistant Lipase from Bacillus Pumilus SG2. Braz. J. Microbiol. 2014, 45, 389–393. [Google Scholar] [CrossRef]
- Handrick, R.; Reinhardt, S.; Focarete, M.L.; Scandola, M.; Adamus, G.; Kowalczuk, M.; Jendrossek, D. A New Type of Thermoalkalophilic Hydrolase of Paucimonas Lemoignei with High Specificity for Amorphous Polyesters of Short Chain-Length Hydroxyalkanoic Acids. J. Biol. Chem. 2001, 276, 36215–36224. [Google Scholar] [CrossRef]
- Masaki, K.; Kamini, N.R.; Ikeda, H.; Iefuji, H. Cutinase-Like Enzyme from the Yeast Cryptococcus Sp. Strain S-2 Hydrolyzes Polylactic Acid and Other Biodegradable Plastics. Appl. Environ. Microbiol. 2005, 71, 7548–7550. [Google Scholar] [CrossRef] [PubMed]
- Chakravorty, D.; Parameswaran, S.; Dubey, V.K.; Patra, S. In Silico Characterization of Thermostable Lipases. Extremophiles 2011, 15, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Gupta, R. Functional Characterisation of Novel Enantioselective Lipase TALipA from Trichosporon Asahii MSR54: Sequence Comparison Revealed New Signature Sequence AXSXG Among Yeast Lipases. Appl. Biochem. Biotechnol. 2015, 175, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Canseco-Pérez, M.; Castillo-Avila, G.; Chi-Manzanero, B.; Islas-Flores, I.; Apolinar-Hernández, M.; Rivera-Muñoz, G.; Gamboa-Angulo, M.; Sanchez-Teyer, F.; Couoh-Uicab, Y.; Canto-Canché, B. Fungal Screening on Olive Oil for Extracellular Triacylglycerol Lipases: Selection of a Trichoderma Harzianum Strain and Genome Wide Search for the Genes. Genes 2018, 9, 62. [Google Scholar] [CrossRef]
- Chakravarty, S.; Varadarajan, R. Elucidation of Determinants of Protein Stability through Genome Sequence Analysis. FEBS Lett. 2000, 470, 65–69. [Google Scholar] [CrossRef]
- Kumar, S.; Tsai, C.-J.; Nussinov, R. Factors Enhancing Protein Thermostability. Protein Eng. Des. Sel. 2000, 13, 179–191. [Google Scholar] [CrossRef]
- Trivedi, S.; Gehlot, H.S.; Rao, S.R. Protein Thermostability in Archaea and Eubacteria. Genet. Mol. Res. 2006, 5, 816–827. [Google Scholar]
- Russell, R.J.; Hough, D.W.; Danson, M.J.; Taylor, G.L. The Crystal Structure of Citrate Synthase from the Thermophilic Archaeon, Thermoplasma Acidophilum. Structure 1994, 2, 1157–1167. [Google Scholar] [CrossRef]
- Nur, M.; Wood, K.; Michelmore, R. EffectorO: Motif-Independent Prediction of Effectors in Oomycete Genomes Using Machine Learning and Lineage Specificity; preprint. bioRxiv 2021. [Google Scholar] [CrossRef]
- Wood, K.J.; Nur, M.; Gil, J.; Fletcher, K.; Lakeman, K.; Gann, D.; Gothberg, A.; Khuu, T.; Kopetzky, J.; Naqvi, S.; et al. Effector Prediction and Characterization in the Oomycete Pathogen Bremia Lactucae Reveal Host-Recognized WY Domain Proteins That Lack the Canonical RXLR Motif. PLoS Pathog. 2020, 16, e1009012. [Google Scholar] [CrossRef]
- Rozewicki, J.; Li, S.; Amada, K.M.; Standley, D.M.; Katoh, K. MAFFT-DASH: Integrated Protein Sequence and Structural Alignment. Nucleic Acids Res. 2019, 47, W5–W10. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (ITOL): An Online Tool for Phylogenetic Tree Display and Annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef]
- Nardini, M.; Lang, D.A.; Liebeton, K.; Jaeger, K.-E.; Dijkstra, B.W. Crystal Structure of Pseudomonas Aeruginosa Lipase in the Open Conformation. J. Biol. Chem. 2000, 275, 31219–31225. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Canul, Y.Y.; Canto-Canché, B.; Berezovski, M.V.; Mironov, G.; Loyola-Vargas, V.M.; Barba de Rosa, A.P.; Tzec-Simá, M.; Brito-Argáez, L.; Carrillo-Pech, M.; Grijalva-Arango, R.; et al. The Cell Wall Proteome from Two Strains of Pseudocercospora Fijiensis with Differences in Virulence. World J. Microbiol. Biotechnol. 2019, 35, 105. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Robertson, S.L.; Parker, J.; Muddiman, D.C.; Dean, R.A. Comparative Proteomic Analysis between Nitrogen Supplemented and Starved Conditions in Magnaporthe Oryzae. Proteome Sci. 2017, 15, 20. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, R.S.; Arias, M.J.D.; Bedoya, J.C.; Rueda, L.E.A.; Sánchez, C.Y.; Granada, G.S.D. Metabolites Produced by Antagonistic Microbes Inhibit the Principal Avocado Pathogens in Vitro. Agron. Colomb. 2015, 33, 58–63. [Google Scholar] [CrossRef]
- Ramesh, A.; Harani Devi, P.; Chattopadhyay, S.; Kavitha, M. Commercial Applications of Microbial Enzymes. In Microbial Enzymes: Roles and Applications in Industries; Arora, N.K., Mishra, J., Mishra, V., Eds.; Microorganisms for Sustainability; Springer: Singapore, 2020; Volume 11, pp. 137–184. [Google Scholar] [CrossRef]
- Chandra, P.; Enespa; Singh, R.; Arora, P.K. Microbial Lipases and Their Industrial Applications: A Comprehensive Review. Microb. Cell Fact 2020, 19, 169. [Google Scholar] [CrossRef]
- Darwish, A.M.G.; Abo Nahas, H.H.; Korra, Y.H.; Osman, A.A.; El-Kholy, W.M.; Reyes-Córdova, M.; Saied, E.M.; Abdel-Azeem, A.M. Fungal Lipases: Insights into Molecular Structures and Biotechnological Applications in Medicine and Dairy Industry. In Industrially Important Fungi for Sustainable Development; Abdel-Azeem, A.M., Yadav, A.N., Yadav, N., Sharma, M., Eds.; Fungal Biology; Springer International Publishing: Cham, Switzerland, 2021; pp. 461–514. [Google Scholar] [CrossRef]
- Geoffry, K.; Achur, R.N. Screening and Production of Lipase from Fungal Organisms. Biocatal. Agric. Biotechnol. 2018, 14, 241–253. [Google Scholar] [CrossRef]
- Eggert, T.; van Pouderoyen, G.; Pencreac’h, G.; Douchet, I.; Verger, R.; Dijkstra, B.W.; Jaeger, K.-E. Biochemical Properties and Three-Dimensional Structures of Two Extracellular Lipolytic Enzymes from Bacillus Subtilis. Colloids Surf. B Biointerfaces 2002, 26, 37–46. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, M.; Xing, S.; Wu, T.; He, H.; Bielicki, J.K.; Chen, J. Identification and Biochemical Characterization of a Novel Hormone-Sensitive Lipase Family Esterase Est19 from the Antarctic Bacterium Pseudomonas Sp. E2-15. Biomolecules 2021, 11, 1552. [Google Scholar] [CrossRef]
- Duarte-Oliveira, C.; Rodrigues, F.; Gonçalves, S.M.; Goldman, G.H.; Carvalho, A.; Cunha, C. The Cell Biology of the Trichosporon-Host Interaction. Front. Cell. Infect. Microbiol. 2017, 7, 118. [Google Scholar] [CrossRef] [PubMed]
- Heidarzadeh, N.; Baghaee-Ravari, S. Application of Bacillus pumilus as a Potential Biocontrol Agent of Fusarium Wilt of Tomato. Arch. Phytopathol. Plant Prot. 2015, 48, 841–849. [Google Scholar] [CrossRef]
- Khan, N.; Maymon, M.; Hirsch, A. Combating Fusarium Infection Using Bacillus-Based Antimicrobials. Microorganisms 2017, 5, 75. [Google Scholar] [CrossRef] [PubMed]
- Bosqueiro, A.S.; Bizarria Júnior, R.; Rosa-Magri, M.M. Potential of Trichosporon Asahii against Alternaria Sp. and Mechanisms of Actions. Summa Phytopathol. 2020, 46, 20–25. [Google Scholar] [CrossRef]
- Park, M.; Do, E.; Jung, W.H. Lipolytic Enzymes Involved in the Virulence of Human Pathogenic Fungi. Mycobiology 2013, 41, 67–72. [Google Scholar] [CrossRef]
- Stehr, F.; Kretschmar, M.; Kröger, C.; Hube, B.; Schäfer, W. Microbial Lipases as Virulence Factors. J. Mol. Catal. B Enzym. 2003, 22, 347–355. [Google Scholar] [CrossRef]
- Voigt, C.A.; Schäfer, W.; Salomon, S. A Secreted Lipase of Fusarium Graminearum Is a Virulence Factor Required for Infection of Cereals: Lipase as a Virulence Factor. Plant J. 2005, 42, 364–375. [Google Scholar] [CrossRef]
- Blümke, A.; Falter, C.; Herrfurth, C.; Sode, B.; Bode, R.; Schäfer, W.; Feussner, I.; Voigt, C.A. Secreted Fungal Effector Lipase Releases Free Fatty Acids to Inhibit Innate Immunity-Related Callose Formation during Wheat Head Infection. Plant Physiol. 2014, 165, 346–358. [Google Scholar] [CrossRef]
- Schuster, A.; Schmoll, M. Biology and Biotechnology of Trichoderma. Appl. Microbiol. Biotechnol. 2010, 87, 787–799. [Google Scholar] [CrossRef]
- Pupin, A.M.; Messias, C.L.; Piedrabuena, A.E.; Roberts, D.W. Total Lipids and Fatty Acids of Strains of Metarhizium Anisopliae. Braz. J. Microbiol. 2000, 31, 121–128. [Google Scholar] [CrossRef]
- Keyhani, N.O. Lipid Biology in Fungal Stress and Virulence: Entomopathogenic Fungi. Fungal Biol. 2018, 122, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Berto, P.; Comménil, P.; Belingheri, L.; Dehorter, B. Occurrence of a Lipase in Spores of Alternaria brassicicola with a Crucial Role in the Infection of Cauliflower Leaves. FEMS Microbiol. Lett. 1999, 180, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Nasser Eddine, A.; Hannemann, F.; Schäfer, W. Cloning and Expression Analysis of NhL1, a Gene Encoding an Extracellular Lipase from the Fungal Pea Pathogen Nectria Haematococca MP VI (Fusarium solani f. Sp. Pisi) That Is Expressed in Planta. Mol. Gen. Genom. 2001, 265, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Adetunji, A.I.; Olaniran, A.O. Production Strategies and Biotechnological Relevance of Microbial Lipases: A Review. Braz. J. Microbiol. 2021, 52, 1257–1269. [Google Scholar] [CrossRef] [PubMed]
- Akao, T.; Gomi, K.; Goto, K.; Okazaki, N.; Akita, O. Subtractive Cloning of CDNA from Aspergillus Oryzae Differentially Regulated between Solid-State Culture and Liquid (Submerged) Culture. Curr. Genet. 2002, 41, 275–281. [Google Scholar] [CrossRef]
- Teufel, F.; Armenteros, J.J.A.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef]
- Carreón-Anguiano, K.G.; Islas-Flores, I.; Vega-Arreguín, J.; Sáenz-Carbonell, L.; Canto-Canché, B. EffHunter: A Tool for Prediction of Effector Protein Candidates in Fungal Proteomic Databases. Biomolecules 2020, 10, 712. [Google Scholar] [CrossRef]
- Navarrete, F.; Grujic, N.; Stirnberg, A.; Saado, I.; Aleksza, D.; Gallei, M.; Adi, H.; Alcântara, A.; Khan, M.; Bindics, J.; et al. The Pleiades Are a Cluster of Fungal Effectors That Inhibit Host Defenses. PLoS Pathog. 2021, 17, e1009641. [Google Scholar] [CrossRef]
- Tzelepis, G.; Dölfors, F.; Holmquist, L.; Dixelius, C. Plant Mitochondria and Chloroplasts Are Targeted by the Rhizoctonia Solani RsCRP1 Effector. Biochem. Biophys. Res. Commun. 2021, 544, 86–90. [Google Scholar] [CrossRef]
- Garcia-Ceron, D.; Lowe, R.G.T.; McKenna, J.A.; Brain, L.M.; Dawson, C.S.; Clark, B.; Berkowitz, O.; Faou, P.; Whelan, J.; Bleackley, M.R.; et al. Extracellular Vesicles from Fusarium Graminearum Contain Protein Effectors Expressed during Infection of Corn. J. Fungi 2021, 7, 977. [Google Scholar] [CrossRef]
- Liebana-Jordan, M.; Brotons, B.; Falcon-Perez, J.M.; Gonzalez, E. Extracellular Vesicles in the Fungi Kingdom. Int. J. Mol. Sci. 2021, 22, 7221. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Dodds, P.N.; Gardiner, D.M.; Singh, K.B.; Taylor, J.M. Improved Prediction of Fungal Effector Proteins from Secretomes with EffectorP 2.0: Prediction of Fungal Effectors with EffectorP 2.0. Mol. Plant Pathol. 2018, 19, 2094–2110. [Google Scholar] [CrossRef] [PubMed]
- Puglisi, I.; Faedda, R.; Sanzaro, V.; Lo Piero, A.R.; Petrone, G.; Cacciola, S.O. Identification of Differentially Expressed Genes in Response to Mercury I and II Stress in Trichoderma Harzianum. Gene 2012, 506, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar] [CrossRef]
- Hallgren, J.; Tsirigos, K.D.; Pedersen, M.D.; Almagro Armenteros, J.J.; Marcatili, P.; Nielsen, H.; Krogh, A.; Winther, O. DeepTMHMM Predicts Alpha and Beta Transmembrane Proteins Using Deep Neural Networks; preprint. Bioinformatics 2022. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, 585–587. [Google Scholar] [CrossRef]
- Mészáros, B.; Erdős, G.; Dosztányi, Z. IUPred2A: Context-Dependent Prediction of Protein Disorder as a Function of Redox State and Protein Binding. Nucleic Acids Res. 2018, 46, W329–W337. [Google Scholar] [CrossRef]
- Gupta, R.; Brunak, S. Prediction of Glycosylation across the Human Proteome and the Correlation to Protein Function. Pac. Symp. Biocomput. 2002, 7, 310–322. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X Version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering Key Features in Protein Structures with the New ENDscript Server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef]
- Matsumura, H.; Yamamoto, T.; Leow, T.C.; Mori, T.; Salleh, A.B.; Basri, M.; Inoue, T.; Kai, Y.; Rahman, R.N.Z.R.A. Novel Cation-π Interaction Revealed by Crystal Structure of Thermoalkalophilic Lipase. Proteins 2007, 70, 592–598. [Google Scholar] [CrossRef]
- Yadav, S.; Dubei, A.; Yadav, S.; Bisht, D.; Darmwal, N.; Yadav, D. Amino Acid Sequences Based Phylogenetic and Motif Assessment of Lipases from Different Organisms. Online J. Bioinform. 2012, 13, 400–417. [Google Scholar]
- Konagurthu, A.S.; Whisstock, J.C.; Stuckey, P.J.; Lesk, A.M. MUSTANG: A Multiple Structural Alignment Algorithm. Proteins 2006, 64, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Krieger, E.; Joo, K.; Lee, J.; Lee, J.; Raman, S.; Thompson, J.; Tyka, M.; Baker, D.; Karplus, K. Improving Physical Realism, Stereochemistry, and Side-Chain Accuracy in Homology Modeling: Four Approaches That Performed Well in CASP8: High-Resolution Homology Modeling. Proteins 2009, 77, 114–122. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Source | T. harzianum Strain | Length (Amino Acids) | % Identity with 135964 | % Identity with KKO99473.1 | % Identity with ThaL |
|---|---|---|---|---|---|---|
| 135964 | CBS 226.95 v1.0, JGI | CBS 226.95 | 339 | -------- | 98.23 | 97.65 |
| KKO99473.1 | GenBank | T6776 | 340 | 98.23 | ------------- | 99.71 |
| ThaL | From this study | B13-1 | 340 | 97.65 | 99.71 | ---------- |
| Order | Family | Genus | Species |
|---|---|---|---|
| Hypocreales | |||
| Hypocreacea | |||
| Trichoderma | harzianum | ||
| lentiforme | |||
| simmonsii | |||
| guizhouense | |||
| virens | |||
| asperellum | |||
| arundinaceum | |||
| asperelloides | |||
| semiorbis | |||
| atroviride | |||
| gracile | |||
| reesei | |||
| longibrachiatum | |||
| gamsii | |||
| citrinoviride | |||
| cornu-damae | |||
| Ophiocordycipitaceae | |||
| Hirsutella | minnesotensis | ||
| rhossiliensis | |||
| Drechmeria | coniospora | ||
| Tolypocladium | capitatum | ||
| ophioglossoides CBS 100239 | |||
| paradoxum | |||
| Ophiocordyceps | sinensis CO18 | ||
| unilateralis | |||
| Purpureocillium | lilacinum | ||
| Nectriaceae | Fusarium | AF-6(solani species complex) | |
| solani | |||
| kuroshium | |||
| AF-8 | |||
| vanettenii | |||
| floridanum | |||
| euwallaceae | |||
| ambrosium | |||
| albosuccineum | |||
| decemcellulare | |||
| sarcochroum | |||
| flagelliforme | |||
| FIESC-RH6 | |||
| sporotrichioides | |||
| avenaceum | |||
| heterosporum | |||
| poae | |||
| venenatum | |||
| pseudograminearum | |||
| graminearum | |||
| MPI-SDFR-AT-0072 | |||
| oxyporum f. sp. rapae | |||
| graminearum PH-1 | |||
| austroamericanum | |||
| culmorum | |||
| redolens | |||
| longipes | |||
| tricinctum | |||
| austroafricanum | |||
| pseudograminearum CS3096 | |||
| mangiferae | |||
| xylarioides | |||
| proliferatum | |||
| proliferatum ET1 | |||
| oxysporum NRRL 32931 | |||
| fujikuroi | |||
| fujikuroi IMI58289 | |||
| pseudocircinatum | |||
| verticillioides | |||
| oxysporum f. sp. lycopersici 4287 | |||
| oxysporum f. sp. melonis 26406 | |||
| oxysporum f. sp. cepae | |||
| oxysporum | |||
| pseudoanthophilum | |||
| Ilyonectria | destructans | ||
| MPI-CAGE-AT-0026 | |||
| robusta | |||
| Thelonectria | olida | ||
| Dactylonectria | estremocensis | ||
| macrididyma | |||
| Stachybotryaceae | Stachybotrys | chartarum | |
| chlorohalonata | |||
| Bionectriaceae | Clonostachys | solani | |
| rosea | |||
| rhizophaga | |||
| Clavicipitaceae | Pochonia | chlamydosporia 170 | |
| Hypocreales incertae sedis | Emericellopsis | atlantica | |
| Acremonium | chrysogenum | ||
| Clavicipitaceae | Epichloe | festucae | |
| Glomerellales | |||
| Glomerellaceae | |||
| Colletotrichum | spaethianum | ||
| simmondsii | |||
| sojae |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Domínguez, D.E.; Chí-Manzanero, B.; Rodríguez-Argüello, M.M.; Todd, J.N.A.; Islas-Flores, I.; Canseco-Pérez, M.Á.; Canto-Canché, B. Identification of a Novel Lipase with AHSMG Pentapeptide in Hypocreales and Glomerellales Filamentous Fungi. Int. J. Mol. Sci. 2022, 23, 9367. https://doi.org/10.3390/ijms23169367
Gutiérrez-Domínguez DE, Chí-Manzanero B, Rodríguez-Argüello MM, Todd JNA, Islas-Flores I, Canseco-Pérez MÁ, Canto-Canché B. Identification of a Novel Lipase with AHSMG Pentapeptide in Hypocreales and Glomerellales Filamentous Fungi. International Journal of Molecular Sciences. 2022; 23(16):9367. https://doi.org/10.3390/ijms23169367
Chicago/Turabian StyleGutiérrez-Domínguez, Denise Esther, Bartolomé Chí-Manzanero, María Mercedes Rodríguez-Argüello, Jewel Nicole Anna Todd, Ignacio Islas-Flores, Miguel Ángel Canseco-Pérez, and Blondy Canto-Canché. 2022. "Identification of a Novel Lipase with AHSMG Pentapeptide in Hypocreales and Glomerellales Filamentous Fungi" International Journal of Molecular Sciences 23, no. 16: 9367. https://doi.org/10.3390/ijms23169367
APA StyleGutiérrez-Domínguez, D. E., Chí-Manzanero, B., Rodríguez-Argüello, M. M., Todd, J. N. A., Islas-Flores, I., Canseco-Pérez, M. Á., & Canto-Canché, B. (2022). Identification of a Novel Lipase with AHSMG Pentapeptide in Hypocreales and Glomerellales Filamentous Fungi. International Journal of Molecular Sciences, 23(16), 9367. https://doi.org/10.3390/ijms23169367