

Characterization of a Novel Fe2+ Activated Non-Blue Laccase from Methylobacterium extorquens


Abstract
:1. Introduction
2. Results
2.1. Recombinant Expression and Purification of Melac13220
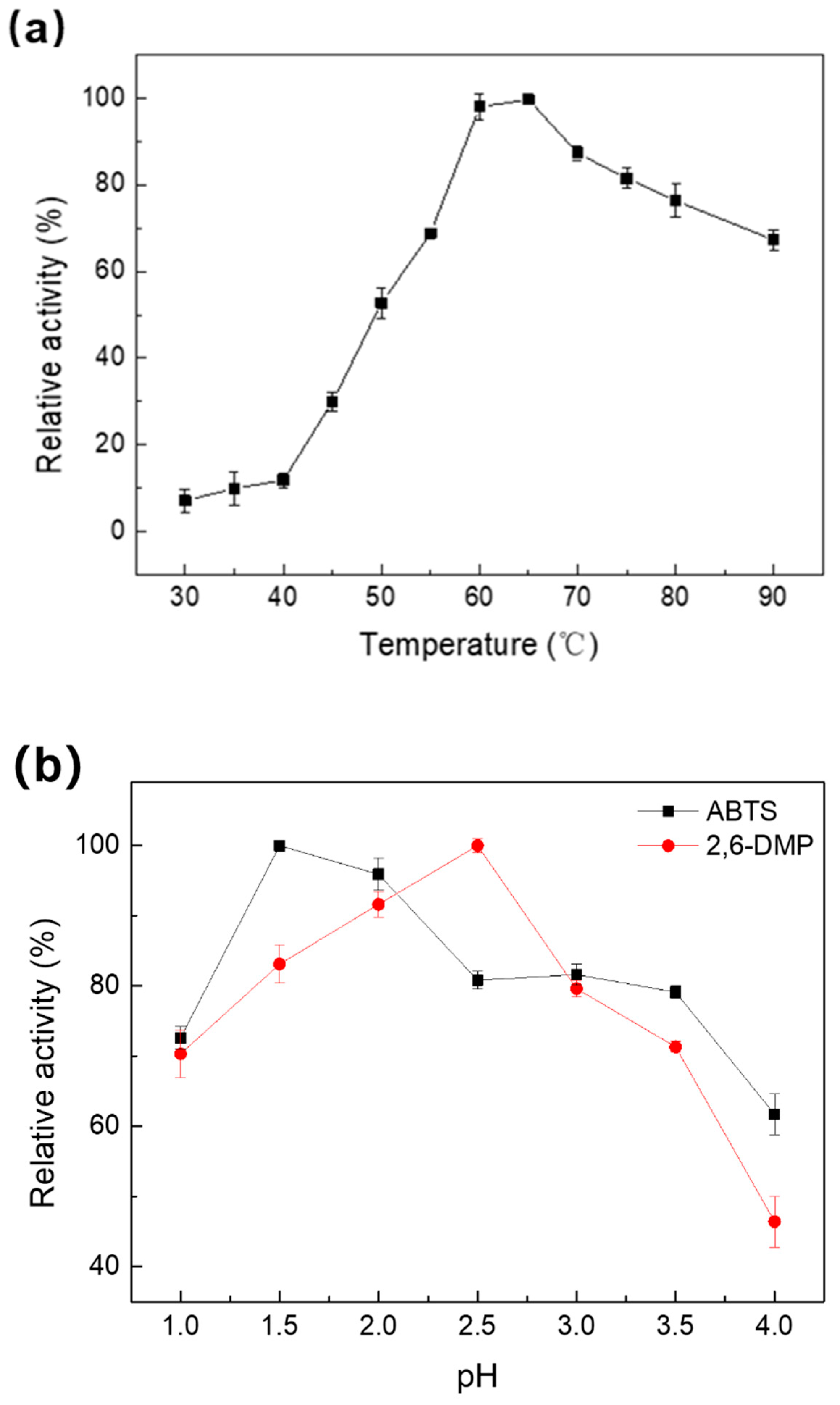
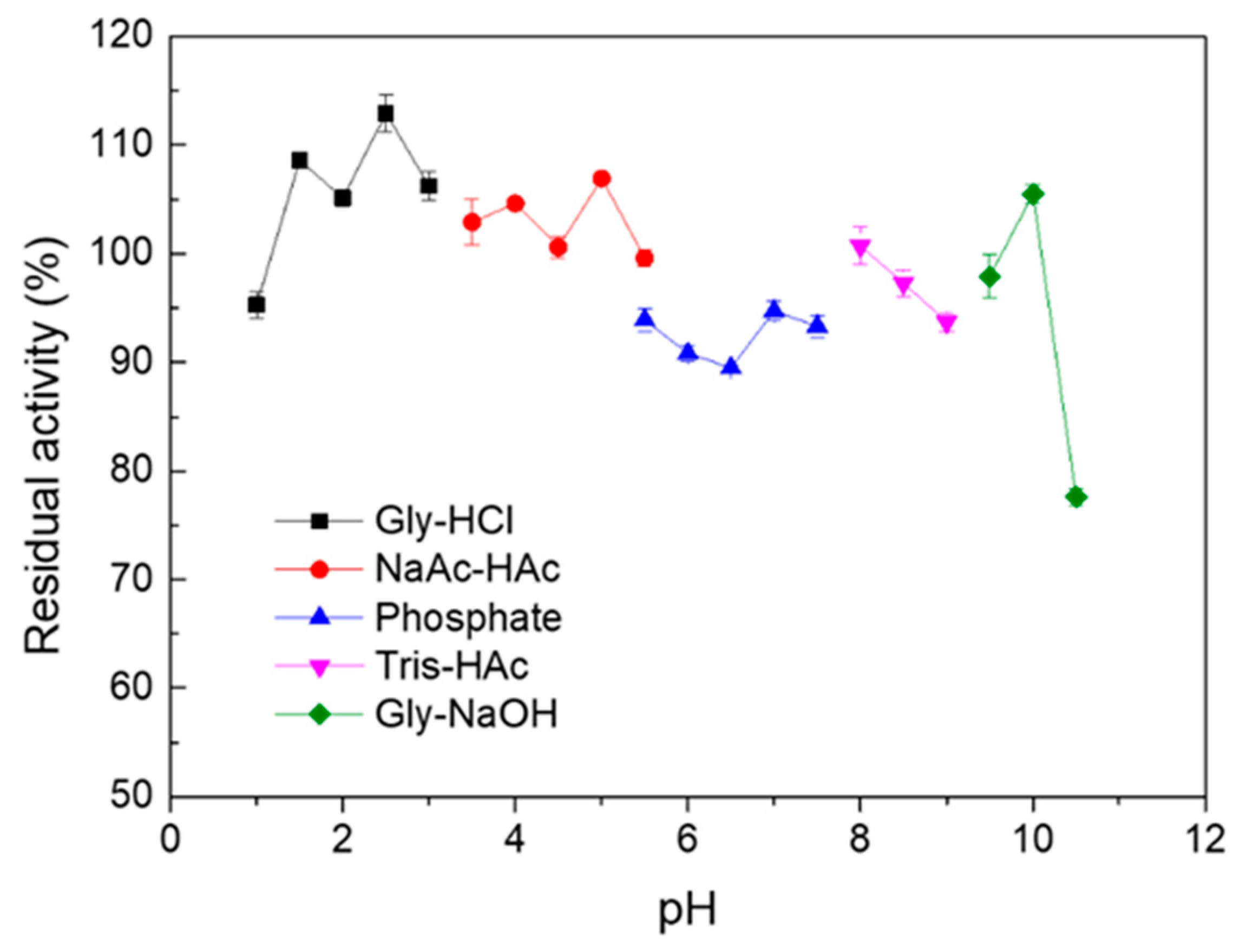
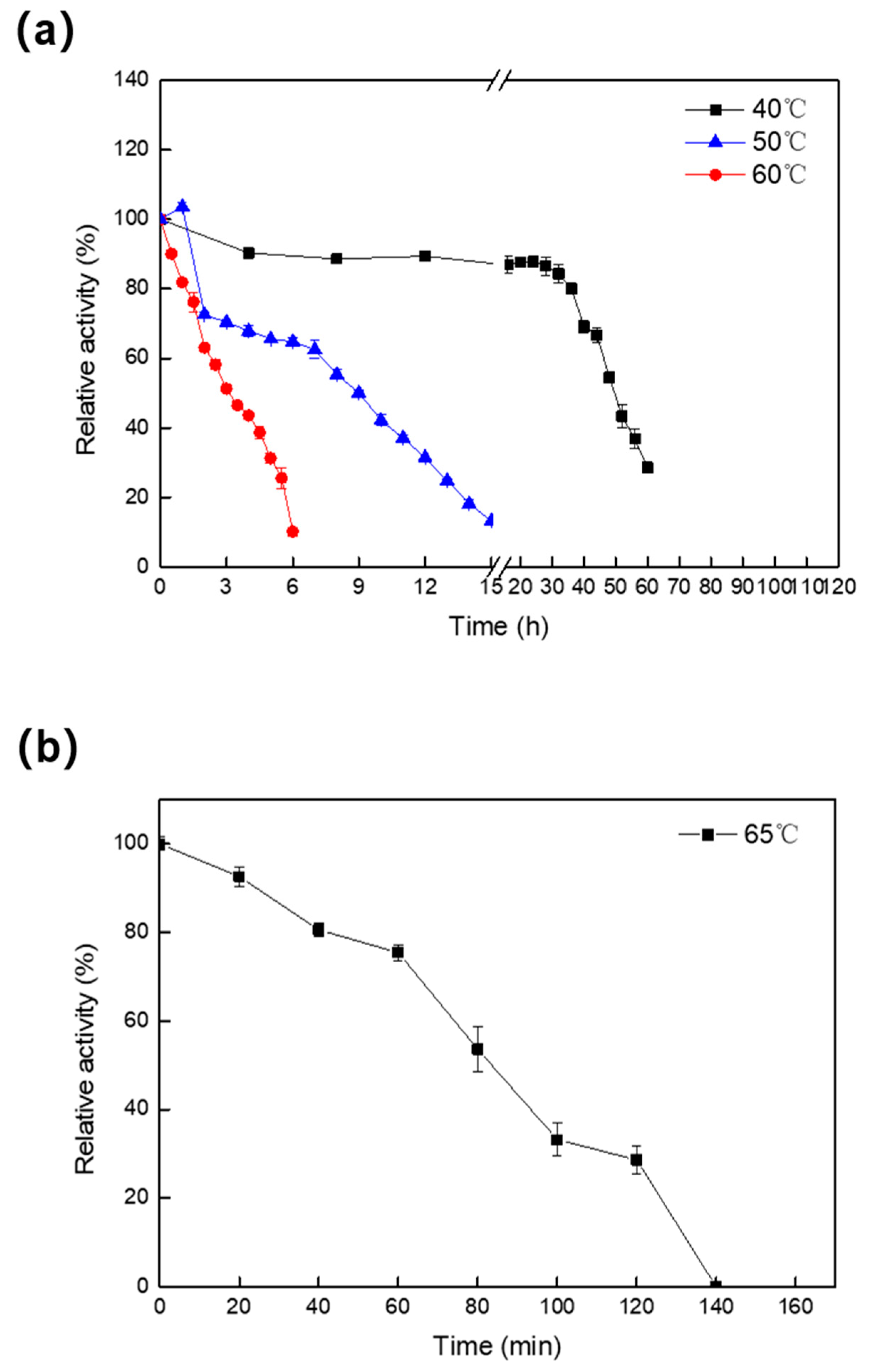
2.2. Effect of Temperature and pH on the Activity and Stability of Melac13220
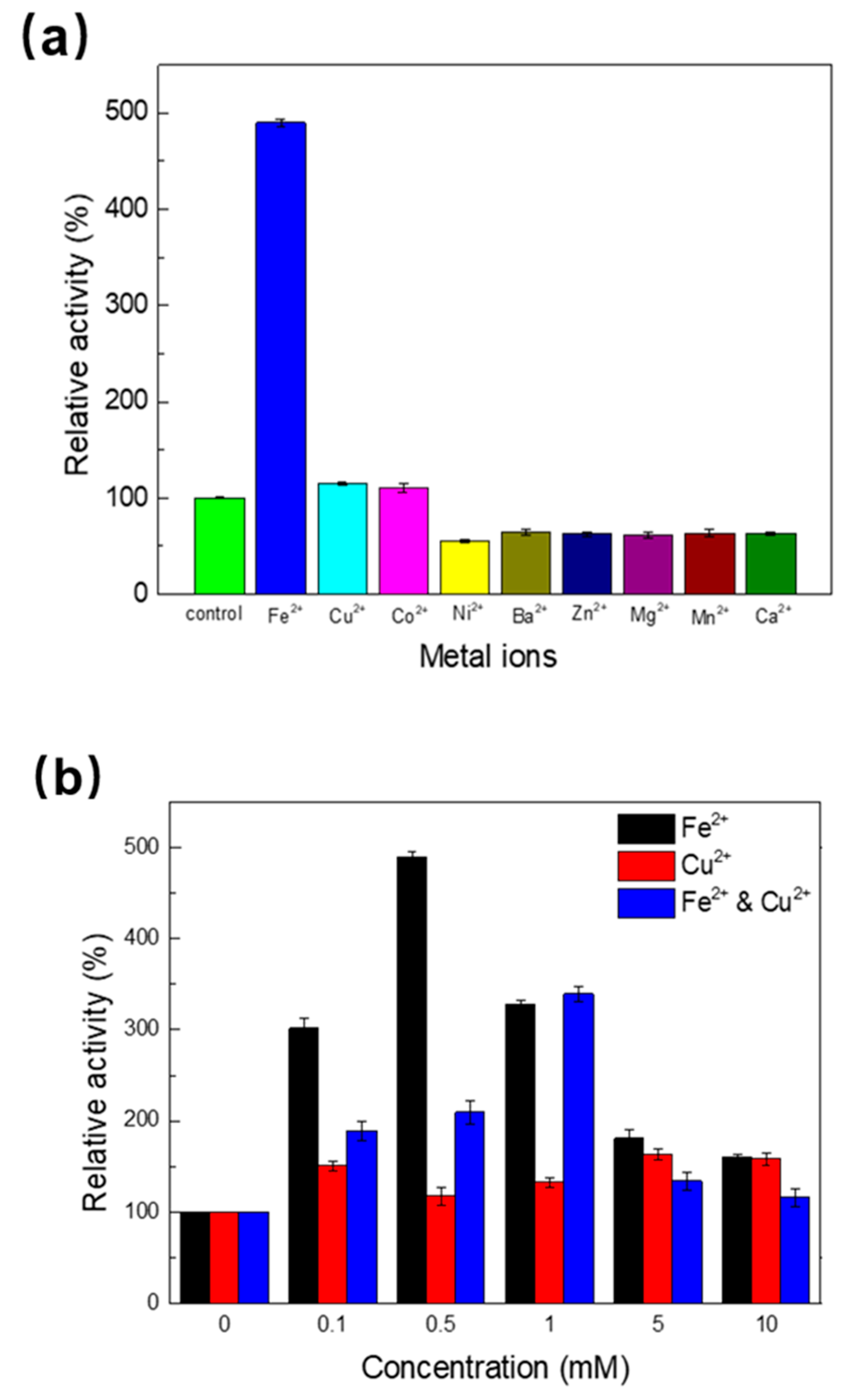
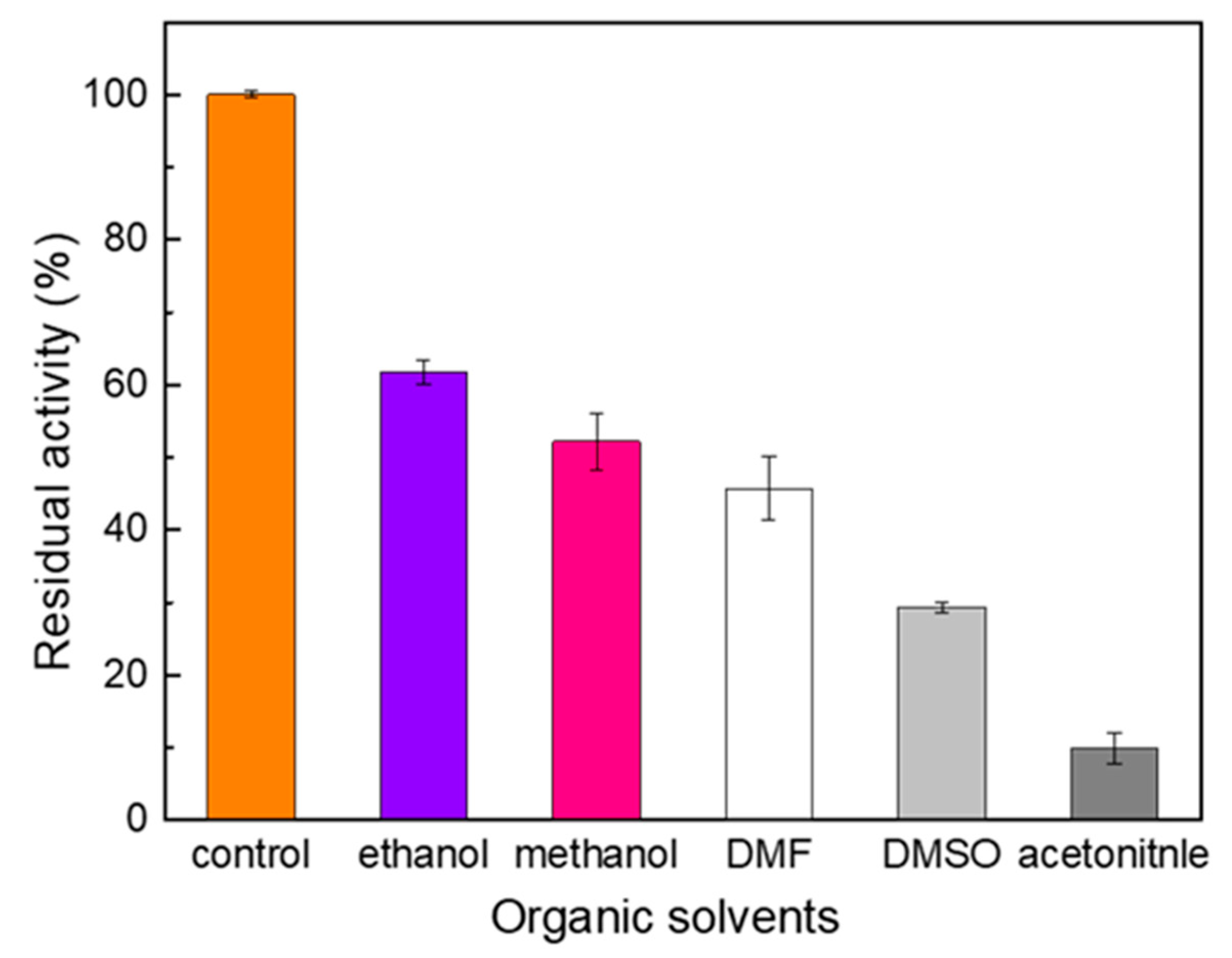
2.3. Effect of Metal Ions and Organic Solvent on the Activity of Melac13220
2.4. Enzyme Kinetic Properties
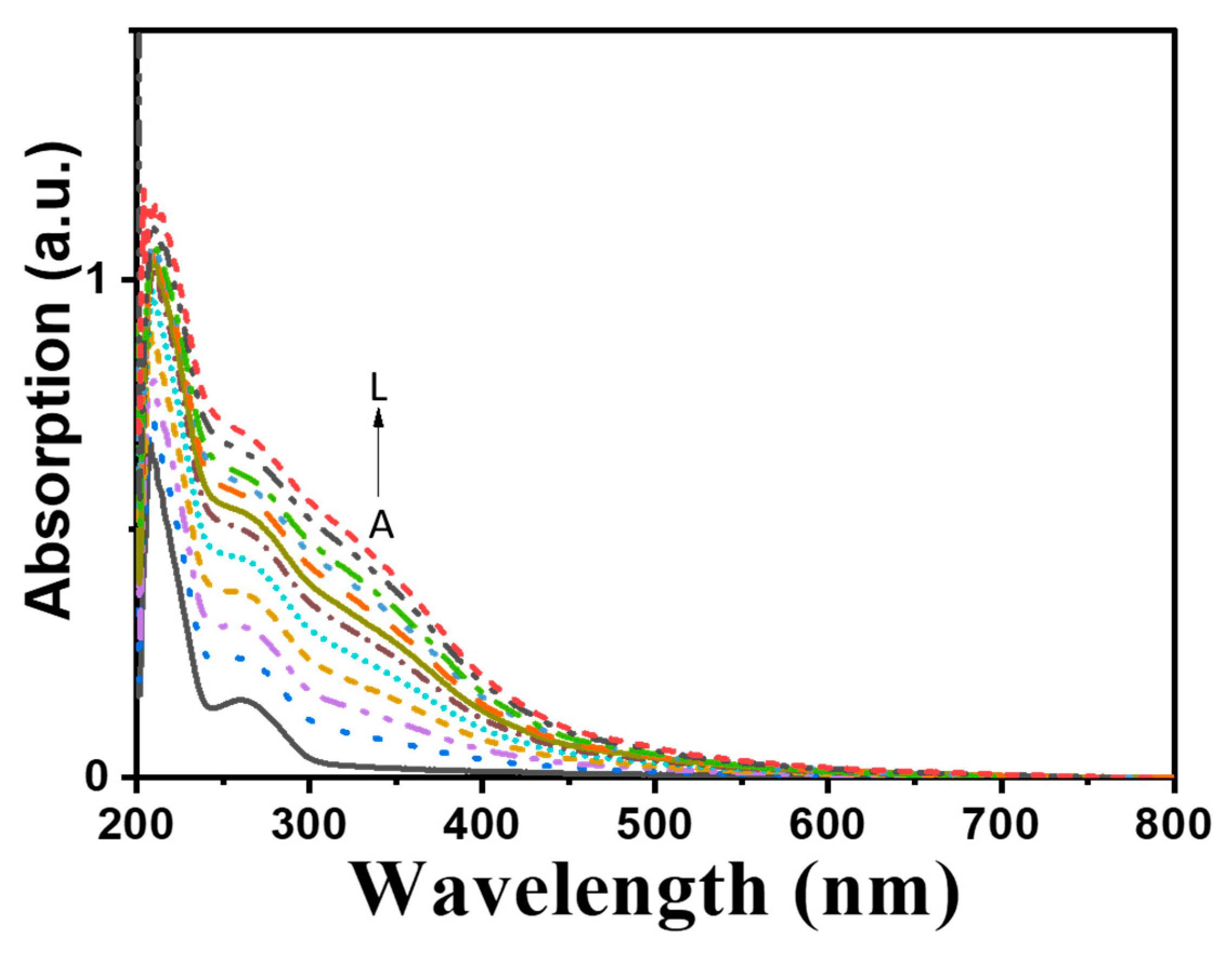
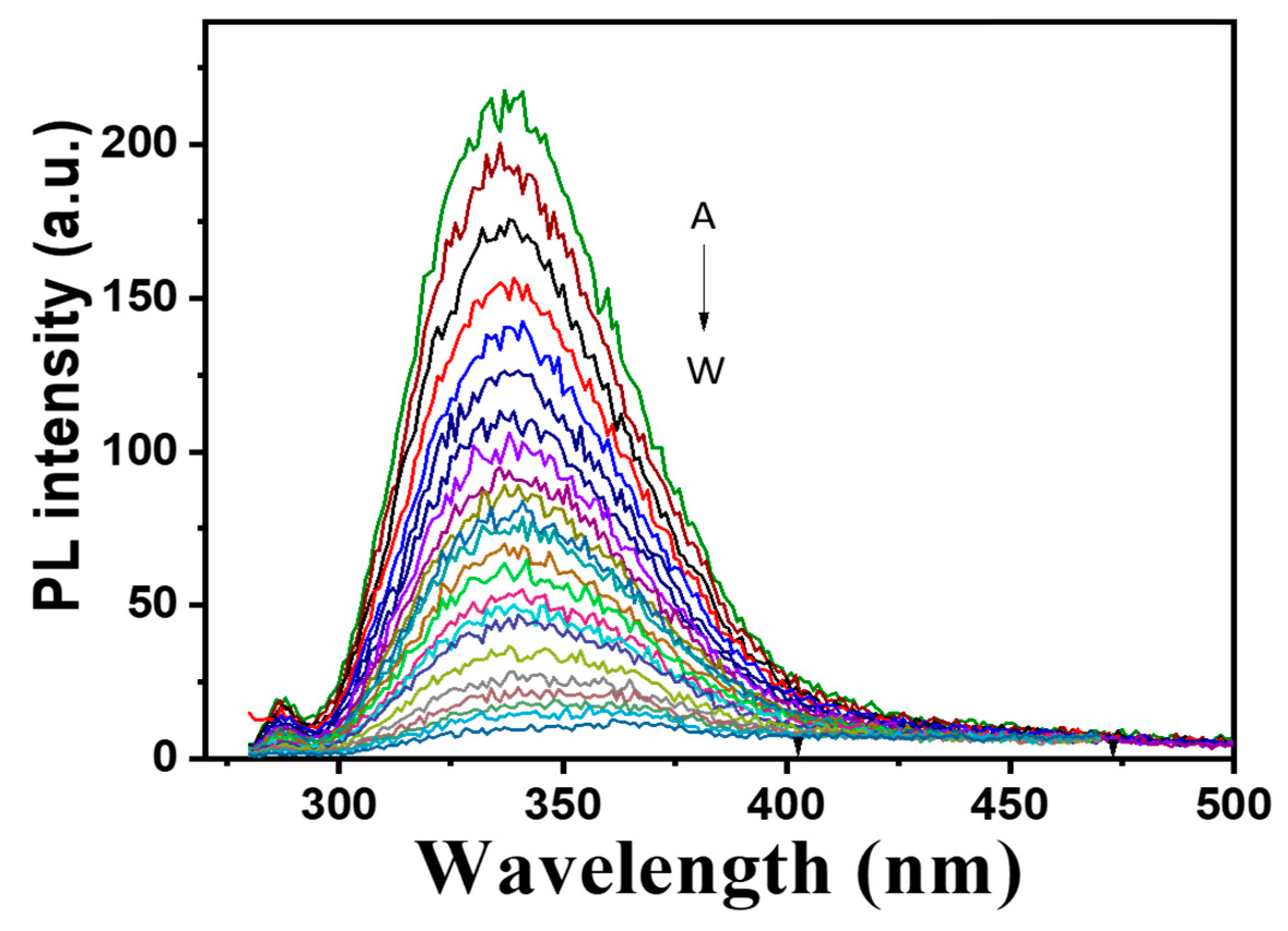
2.5. Spectral Properties, Metal Content and Electron Paramagnetic Resonance (EPR) Analysis of Purified Melac13220
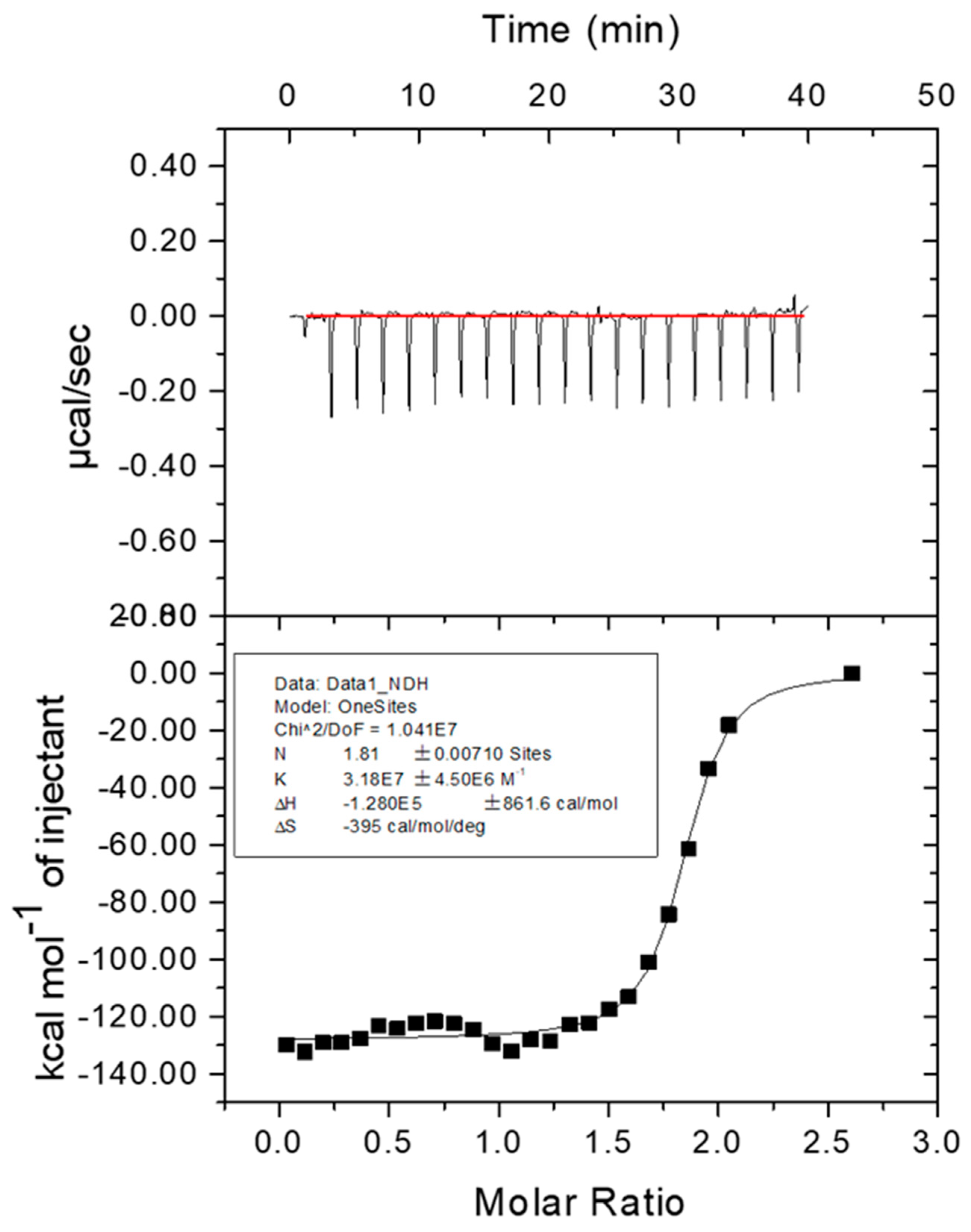
2.6. Affinity of Fe2+ to Melac13220
2.7. Decolorization of Dyes
3. Discussion
3.1. Enzymatic Characteristics of Melac13220
3.2. Effect of Metal Ions (Fe2+) and Organic Solvent on Enzyme Activity
3.3. Substrate and Kinetic Analysis
3.4. Spectral Properties of Purifies Melac13220
3.5. Decolorization of Synthetic Dyes
4. Materials and Methods
4.1. Materials
4.2. Cloning, Expression, and Purification of Melac13220
4.3. Enzyme Assay
4.4. Effects of Temperature and pH on Laccase Activity and Stability
4.5. Effects of Metal Ions and Organic Solvent on Melac13220
4.6. Enzyme Kinetic Studies
4.7. Spectral Characteristics, Metal Content and Electron Paramagnetic Resonance (EPR) Analysis of Melac13220
4.8. Affinity Analysis of Fe2+ to Melac13220
4.9. Dye Decolorization by Melac13220
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bello-Gil, D.; Roig-Molina, E.; Fonseca, J.; Sarmiento-Ferrández, M.D.; Ferrándiz, M.; Franco, E.; Mira, E.; Maestro, B.; Sanz, J.M. An enzymatic system for decolorization of wastewater dyes using immobilized CueO laccase-like multicopper oxidase on poly-3-hydroxybutyrate. Microb. Biotechnol. 2018, 11, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Kudanga, T.; Nemadziva, B.; Le Roes-Hill, M. Laccase catalysis for the synthesis of bioactive compounds. Appl. Microbiol. Biotechnol. 2017, 101, 13–33. [Google Scholar] [CrossRef] [PubMed]
- Osma, J.F.; Toca-Herrera, J.L.; Rodríguez-Couto, S. Uses of Laccases in the Food Industry. Enzym. Res. 2010, 2010, 918769. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, L.N.; Campana, A.L.; Cruz, J.C.; Soto, N.O.; Osma, J.F. Enhanced Catalytic Dye Decolorization by Microencapsulation of Laccase from P. Sanguineus CS43 in Natural and Synthetic Polymers. Polymers 2020, 12, 1353. [Google Scholar] [CrossRef]
- Homaei, A.; Qeshmi, F.I. Purification and characterization of a robust thermostable protease isolated from Bacillus subtilis strain HR02 as an extremozyme. J. Appl. Microbiol. 2022, 1–11. [Google Scholar] [CrossRef]
- Qeshmi, F.I.; Homaei, A.; Khajeh, K.; Kamrani, E.; Fernandes, P. Production of a Novel Marine Pseudomonas aeruginosa Recombicc. Mar. Biotechnol. 2022, 24, 599–613. [Google Scholar] [CrossRef]
- Li, J.; Xie, Y.; Wang, R.; Fang, Z.; Fang, W.; Zhang, X.; Xiao, Y. Mechanism of salt-induced activity enhancement of a marine-derived laccase, Lac15. Eur. Biophys. J. 2018, 47, 225–236. [Google Scholar] [CrossRef]
- Assadi, G.; Vesterlund, L.; Bonfiglio, F.; Mazzurana, L.; Cordeddu, L.; Schepis, D.; Mjösberg, J.; Ruhrmann, S.; Fabbri, A.; Vukojevic, V.; et al. Functional Analyses of the Crohn’s Disease Risk Gene LACC1. PLoS ONE 2016, 11, e0168276. [Google Scholar] [CrossRef]
- Zhang, H.B.; Zhang, Y.L.; Huang, F.; Gao, P.J.; Chen, J.C. Purification and characterization of a thermostable laccase with unique oxidative characteristics from Trametes hirsute. Biotechnol. Lett. 2009, 31, 837–843. [Google Scholar] [CrossRef]
- Ikeda, R.; Uyama, H.; Kobayashi, S. Novel Synthetic Pathway to a Poly(phenylene oxide). Laccase-Catalyzed Oxidative Polymerization of Syringic Acid. Macromolecules 1996, 29, 3053–3054. [Google Scholar] [CrossRef]
- Scheibel, D.M.; Guo, D.; Luo, J.; Gitsov, I. A Single Enzyme Mediates the “Quasi-Living” Formation of Multiblock Copolymers with a Broad Biomedical Potential. Biomacromolecules 2020, 21, 2132–2146. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.M.; Solomon, E.I. Electron transfer and reaction mechanism of laccases. Cell. Mol. Life Sci. 2015, 72, 869–883. [Google Scholar] [CrossRef] [PubMed]
- Viswanath, B.; Rajesh, B.; Janardhan, A.; Kumar, A.P.; Narasimha, G. Fungal Laccases and Their Applications in Bioremediation. Enzyme Res. 2014, 2014, 163242–163263. [Google Scholar] [CrossRef] [PubMed]
- Iark, D.; dos Reis Buzzo, A.J.; Garcia, J.A.; Côrrea, V.G.; Helm, C.V.; Corrêa, R.C.; Peralta, R.A.; Moreira, R.D.; Bracht, A.; Peralta, R.M. Enzymatic degradation and detoxification of azo dye Congo red by a new laccase from Oudemansiella canarii. Bioresour. Technol. 2019, 289, 121655. [Google Scholar] [CrossRef]
- Witayakran, S.; Ragauskas, A.J. Synthetic Applications of Laccase in Green Chemistry. Adv. Synth. Catal. 2009, 351, 1187–1209. [Google Scholar] [CrossRef]
- Patel, S.K.S.; Gupta, R.K.; Kim, S.-Y.; Kim, I.-W.; Kalia, V.C.; Lee, J.-K. Rhus vernicifera Laccase Immobilization on Magnetic Nanoparticles to Improve Stability and Its Potential Application in Bisphenol A Degradation. Indian J. Microbiol. 2020, 61, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, L.; Guo, W.; Jia, L.; Fu, Y.; Gui, S.; Lu, F. Cloning, expression, and characterization of a thermostable and pH-stable laccase from Klebsiella pneumoniae and its application to dye decolorization. Process Biochem. 2017, 53, 125–134. [Google Scholar] [CrossRef]
- Mehandia, S.; Sharma, S.C.; Arya, S.K. Isolation and characterization of an alkali and thermostable laccase from a novel Alcaligenes faecalis and its application in decolorization of synthetic dyes. Biotechnol. Rep. 2020, 25, e00413–e00421. [Google Scholar] [CrossRef]
- Gupta, M.N.; Bisaria, V.S. Stable cellulolytic enzymes and their application in hydrolysis of lignocellulosic biomass. Biotechnol. J. 2018, 13, 1700633–1700642. [Google Scholar] [CrossRef]
- Hossain, L.; Sarker, S.K.; Khan, M.S. Evaluation of present and future wastewater impacts of textile dyeing industries in Bangladesh. Environ. Dev. 2018, 26, 23–33. [Google Scholar] [CrossRef]
- Kumar, M.; Mishra, A.; Singh, S.S.; Srivastava, S.; Thakur, I.S. Expression and characterization of novel laccase gene from Pandoraea sp. ISTKB and its application. Int. J. Biol. Macromol. 2018, 115, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Ning, Y.J.; Wang, S.N.; Zhang, J.; Zhang, G.Q.; Chen, Q.J. Purification, characterization, and cloning of an extracellular laccase with potent dye decolorizing ability from white rot fungus Cerrena unicolor GSM-01. Int. J. Biol. Macromol. 2017, 95, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-N.; Chen, Q.-J.; Zhu, M.-J.; Xue, F.-Y.; Li, W.-C.; Zhao, T.-J.; Li, G.-D.; Zhang, G.-Q. An extracellular yellow laccase from white rot fungus Trametes sp. F1635 and its mediator systems for dye decolorization. Biochimie 2018, 148, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Huang, L.; Li, Y.; Ma, J.; Wang, S.; Zhang, Y.; Ge, X.; Wang, N.; Lu, F.; Liu, Y. Characterization and application of a novel laccase derived from Bacillus amyloliquefaciens. Int. J. Biol. Macromol. 2020, 150, 982–990. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Wang, J.; Yin, Q.; Fang, W.; Xiao, Y.; Fang, Z. Expression of a thermo- and alkali-philic fungal laccase in Pichia pastoris and its application. Protein Expr. Purif. 2019, 154, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; An, Q.; Meng, G.; Wu, X.J.; Dai, Y.C.; Si, J.; Cui, B.K. A novel laccase from white rot fungus Trametes orientalis: Purification, characterization, and application. Int. J. Biol. Macromol. 2017, 102, 758–770. [Google Scholar] [CrossRef]
- Brissos, V.; Goncalves, N.; Melo, E.P.; Martins, L.O. Improving Kinetic or Thermodynamic Stability of an Azoreductase by Directed Evolution. PLoS ONE 2014, 9, e87209. [Google Scholar] [CrossRef]
- Murugesan, K.; Kim, Y.-M.; Jeon, J.-R.; Chang, Y.-S. Effect of metal ions on reactive dye decolorization by laccase from Ganoderma lucidum. J. Hazard. Mater. 2009, 168, 523–529. [Google Scholar] [CrossRef]
- Shafiei, M.; Afzali, F.; Karkhane, A.A.; Ebrahimi, S.M.; Haghbeen, K.; Aminzadeh, S. Cohnella sp. A01 laccase: Thermostable, detergent resistant, anti-environmental and industrial pollutants enzyme. Heliyon 2019, 5, e02543. [Google Scholar] [CrossRef]
- Yang, J.; Lin, Q.; Ng, T.B.; Ye, X.; Lin, J. Purification and Characterization of a Novel Laccase from Cerrena sp. HYB07 with Dye Decolorizing Ability. PLoS ONE 2014, 9, e110834. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.J.; Kim, H.W.; Jeon, S.J. Biochemical characterization of a thermostable cobalt- or copper-dependent polyphenol oxidase with dye decolorizing ability from Geobacillus sp. JS12. Enzyme Microb. Technol. 2018, 118, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P.; Gabriel, J. Copper and cadmium increase laccase activity in Pleurotus ostreatus. FEMS Microbiol. Lett. 2002, 206, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Meng, X.; Zhou, H.; Liu, Y.; Secundo, F.; Liu, Y. Enzyme Stability and Activity in Non-Aqueous Reaction Systems: A Mini Review. Catalysts 2016, 6, 32. [Google Scholar] [CrossRef]
- Palmieri, G.; Giardina, P.; Bianco, C.; Scaloni, A.; Capasso, A.; Sannia, G. A Novel White Laccase from Pleurotus ostreatus. J. Biol. Chem. 1997, 272, 31301–31307. [Google Scholar] [CrossRef]
- Min, K.-L.; Kim, Y.-H.; Kim, Y.W.; Jung, H.S.; Hah, Y.C. Characterization of a Novel Laccase Produced by the Wood-Rotting Fungus Phellinus ribis. Arch. Biochem. Biophys. 2001, 392, 279–286. [Google Scholar] [CrossRef]
- Sonica, S.; Randhir, K.; Jitendra, M. Purification and characterization of a novel white highly thermo stable laccase from a novel Bacillus sp. MSK-01 having potential to be used as anticancer agent. Int. J. Biol. Macromol. 2021, 170, 232–238. [Google Scholar] [CrossRef]
- Zhao, D.; Zhang, X.; Cui, D.; Zhao, M. Characterisation of a Novel White Laccase from the Deuteromycete Fungus Myrothecium verrucaria NF-05 and Its Decolourisation of Dyes. PLoS ONE 2016, 7, e38817. [Google Scholar] [CrossRef]
- Diamantidis, G.; Effosse, A.; Potier, P.; Bally, R. Purification and characterization of the first bacterial laccase in the rhizospheric bacterium Azospirillum lipoferum. Soil Biol. Biochem. 2000, 32, 919–927. [Google Scholar] [CrossRef]
- Rai, R.; Bibra, M.; Chadha, B.S.; Sani, R.K. Enhanced hydrolysis of lignocellulosic biomass with doping of a highly thermostable recombinant laccase. Int. J. Biol. Macromol. 2019, 137, 232–237. [Google Scholar] [CrossRef]
- Margot, J.; Bennati-Granier, C.; Maillard, J.; Blánquez, P.; Barry, D.A.; Holliger, C. Bacterial versus fungal laccase: Potential for micropollutant degradation. AMB Express 2013, 3, 63. [Google Scholar] [CrossRef] [Green Version]
- Sadhasivam, S.; Savitha, S.; Swaminathan, K.; Lin, F.-H. Production, purification and characterization of mid-redox potential laccase from a newly isolated Trichoderma harzianum WL1. Process Biochem. 2008, 43, 736–742. [Google Scholar] [CrossRef]
- Lu, C.; Cao, L.; Liu, R.; Lei, Y.; Ding, G.J. Effect of common metal ions on the rate of degradation of 4-nitrophenol by a laccase-Cu2+ synergistic system. J. Environ. Manag. 2012, 113, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Lu, Y.; Zhang, R.; Wang, H.; Liu, Q. Characterization of a novel laccase purified from the fungus Hohenbuehelia serotina and its decolourisation of dyes. Acta Biochim. Pol. 2016, 63, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Shleev, S.; Jarosz-Wilkolazka, A.; Khalunina, A.; Morozova, O.; Yaropolov, A.; Ruzgas, T.; Gorton, L. Direct electron transfer reactions of laccases from different origins on carbon electrodes. Bioelectrochemistry 2005, 67, 115–124. [Google Scholar] [CrossRef]
- Ghatge, S.; Yang, Y.; Song, W.-Y.; Kim, T.-Y.; Hur, H.-G. A novel laccase from thermoalkaliphilic bacterium Caldalkalibacillus thermarum strain TA2.A1 able to catalyze dimerization of a lignin model compound. Appl. Microbiol. Biotechnol. 2018, 102, 4075–4086. [Google Scholar] [CrossRef]
- DeRose, V.J.; Liu, K.E.; Lippard, S.J.; Hoffman, B.M. Investigation of the Dinuclear Fe Center of Methane Monooxygenase by Advanced Paramagnetic Resonance Techniques: On the Geometry of DMSO Binding. J. Am. Chem. Soc. 1996, 118, 121–134. [Google Scholar] [CrossRef]
- Secundo, F.; Carrea, G. Optimization of Hydrolase Efficiency in Organic Solvents. Chem. Eur. J. 2003, 9, 3194–3199. [Google Scholar] [CrossRef]
- Bourbonnais, R.; Paice, M.G. Oxidation of non-phenolic substrates. An expanded role for laccase in lignin biodegradation. FEBS Lett. 1990, 267, 99–102. [Google Scholar] [CrossRef]
- Guan, Z.B.; Luo, Q.; Wang, H.R.; Chen, Y.; Liao, X.R. Bacterial laccases: Promising biological green tools for industrial applications. Cell. Mol. Life Sci. 2018, 75, 3569–3592. [Google Scholar] [CrossRef]
- Mohammadian, M.; Fathi-Roudsari, M.; Mollania, N.; Badoei-Dalfard, A.; Khajeh, K. Enhanced expression of a recombinant bacterial laccase at low temperature and microaerobic conditions: Purification and biochemical characterization. J. Ind. Microbiol. Biotechnol. 2010, 37, 863–869. [Google Scholar] [CrossRef]
- Komal, A.; Nisha, B.; Bikash, K.; Venkatesh, C.; Pradeep, V. Process optimization, purification and characterization of alkaline stable white laccase from Myrothecium verrucaria ITCC-8447 and its application in delignification of agroresidues. Int. J. Biol. Macromol. 2019, 125, 1042–1055. [Google Scholar] [CrossRef]
- Zhao, Y.; Abdurahman, A.; Zhang, Y.; Zheng, P.; Zhang, M.; Li, F. Highly Efficient Multifunctional Luminescent Radicals. CCS Chem. 2022, 4, 722–731. [Google Scholar] [CrossRef]
- Mtibaà, R.; Barriuso, J.; de Eugenio, L.; Aranda, E.; Belbahri, L.; Nasri, M.; Martínez, M.J.; Mechichi, T. Purification and characterization of a fungal laccase from the ascomycete Thielavia sp. and its role in the decolorization of a recalcitrant dye. Int. J. Biol. Macromol. 2018, 120, 1744–1751. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Km (mM) | kcat (s−1) | kcat/Km (mM−1 s−1) |
|---|---|---|---|
| ABTS ABTS (+Fe2+) | 7.65 × 10−2 9.76 × 10−2 | 14.6 74.5 | 190.85 763.32 |
| 2,6-DMP 2,6-DMP (+Fe2+) | 9.18 × 10−2 1.59 × 10−1 | 12.1 33.8 | 130.81 214.06 |
| Microorganism | Molecular Weight (kDa) | Optimum Temperature (°C) | Optimum pH | Km (mM) | kcat (s−1) | kcat/Km (mM−1 s−1) | References |
|---|---|---|---|---|---|---|---|
| Methylobacterium extorquens | 50 | 65 | 1.5 | 7.65 × 10−2 | 14.6 | 190.85 | This study |
| Pleurotus ostreatus | 61 | 60 | 7.0 | 9.0 × 10−2 | 5833 | 64811.1 | [34] |
| Phellinus ribis | 76 | 65 | 5.0 | 0.207 | 1333 | 6439.6 | [35] |
| Bacillus sp. MSK-01 | 32 | 75 | 4.5 | 1.624 | 177 | 109 | [36] |
| Myrothecium verrucaria NF-05 | 66 | 30 | 4.0 | 8.59 × 10−2 | 267.1 | 3109.4 | [37] |
| Trametes hirsuta | 90 | 85 | 2.4 | 7 × 10−2 | 197 | 2800 | [9] |
| Dyes | λmax (nm) | Decolorization (%) | |
|---|---|---|---|
| 10 h | 24 h | ||
| Remazol Brilliant Blue R | 595 | 13.9 ± 0.7 | 29.1 ± 1.3 |
| Crystal Violet | 590 | 10.7 ± 1.1 | 30.5 ± 1.5 |
| Indigo Carmine | 610 | 37 ± 0.6 | 54 ± 0.9 |
| Congo Red | 488 | 93 ± 1.2 | 99 ± 0.7 |
| Origin | Decolorization (%) | Reference | |
|---|---|---|---|
| 10 h | 24 h | ||
| Methylobacterium extorquens | 93% | 99% | This study |
| Pandoraea sp. ISTKB | 20% | 10% | [21] |
| Myrothecium verrucaria NF-05 | / | 58% | [37] |
| Trametes orientalis | / | 60% | [26] |
| Cerrena sp. HYB07 | / | 57% | [30] |
| Alcaligenes faecalis XF1 | / | 31% | [18] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ainiwaer, A.; Liang, Y.; Ye, X.; Gao, R. Characterization of a Novel Fe2+ Activated Non-Blue Laccase from Methylobacterium extorquens. Int. J. Mol. Sci. 2022, 23, 9804. https://doi.org/10.3390/ijms23179804
Ainiwaer A, Liang Y, Ye X, Gao R. Characterization of a Novel Fe2+ Activated Non-Blue Laccase from Methylobacterium extorquens. International Journal of Molecular Sciences. 2022; 23(17):9804. https://doi.org/10.3390/ijms23179804
Chicago/Turabian StyleAiniwaer, Abidan, Yue Liang, Xiao Ye, and Renjun Gao. 2022. "Characterization of a Novel Fe2+ Activated Non-Blue Laccase from Methylobacterium extorquens" International Journal of Molecular Sciences 23, no. 17: 9804. https://doi.org/10.3390/ijms23179804
APA StyleAiniwaer, A., Liang, Y., Ye, X., & Gao, R. (2022). Characterization of a Novel Fe2+ Activated Non-Blue Laccase from Methylobacterium extorquens. International Journal of Molecular Sciences, 23(17), 9804. https://doi.org/10.3390/ijms23179804