

Comprehensive Transcriptome Analysis of Hair Follicle Morphogenesis Reveals That lncRNA-H19 Promotes Dermal Papilla Cell Proliferation through the Chi-miR-214-3p/β-Catenin Axis in Cashmere Goats


Abstract
:1. Introduction
2. Results
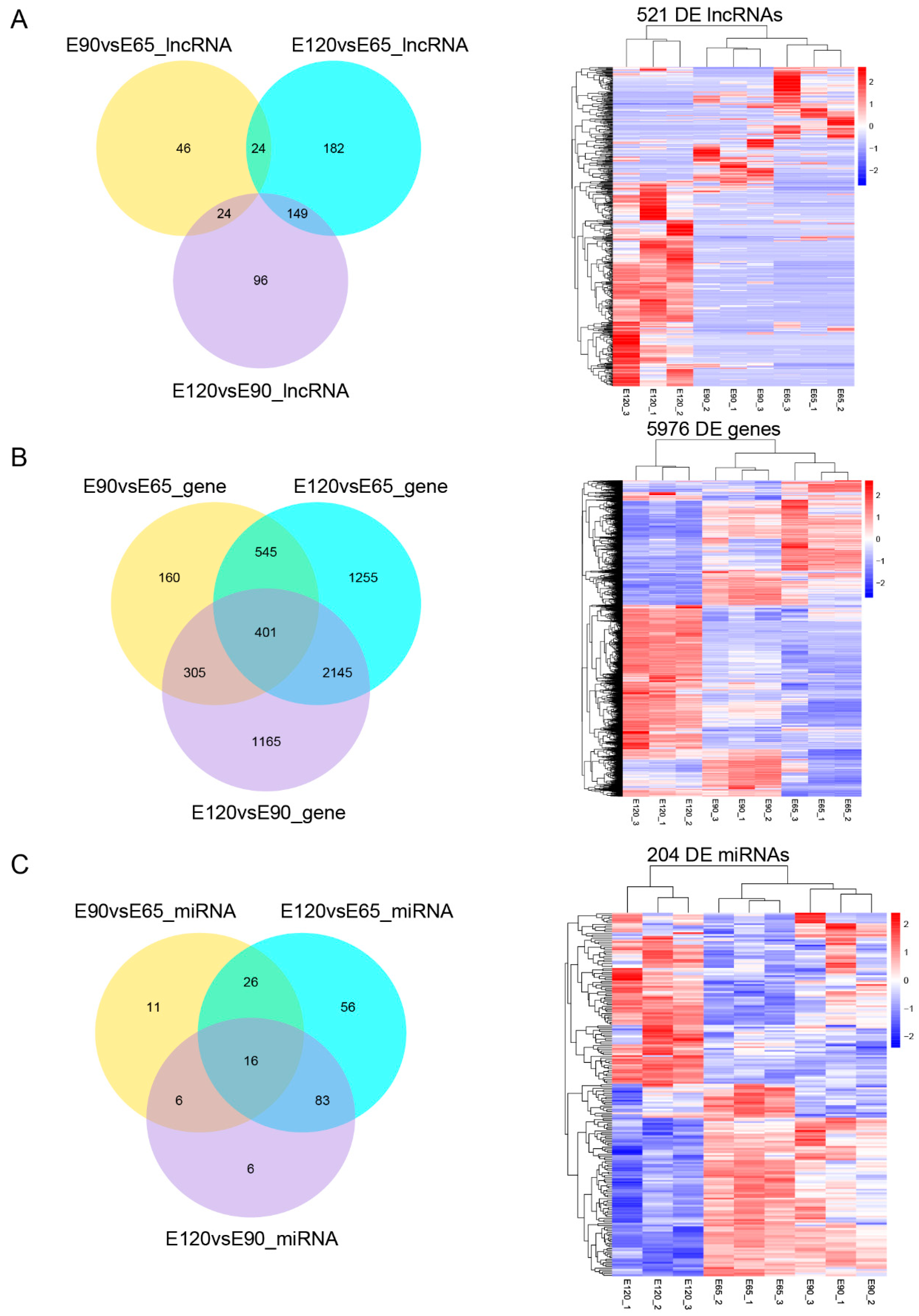
2.1. Identification of lncRNAs and miRNAs in Goat Skin
2.2. Differentially Expressed Analysis and lncRNA Target Prediction
2.3. KEGG Pathway Analysis of DE Genes
2.4. lncRNA–miRNA–mRNA Networks
2.5. LncRNA H19 Promotes the Proliferation of DP Cells
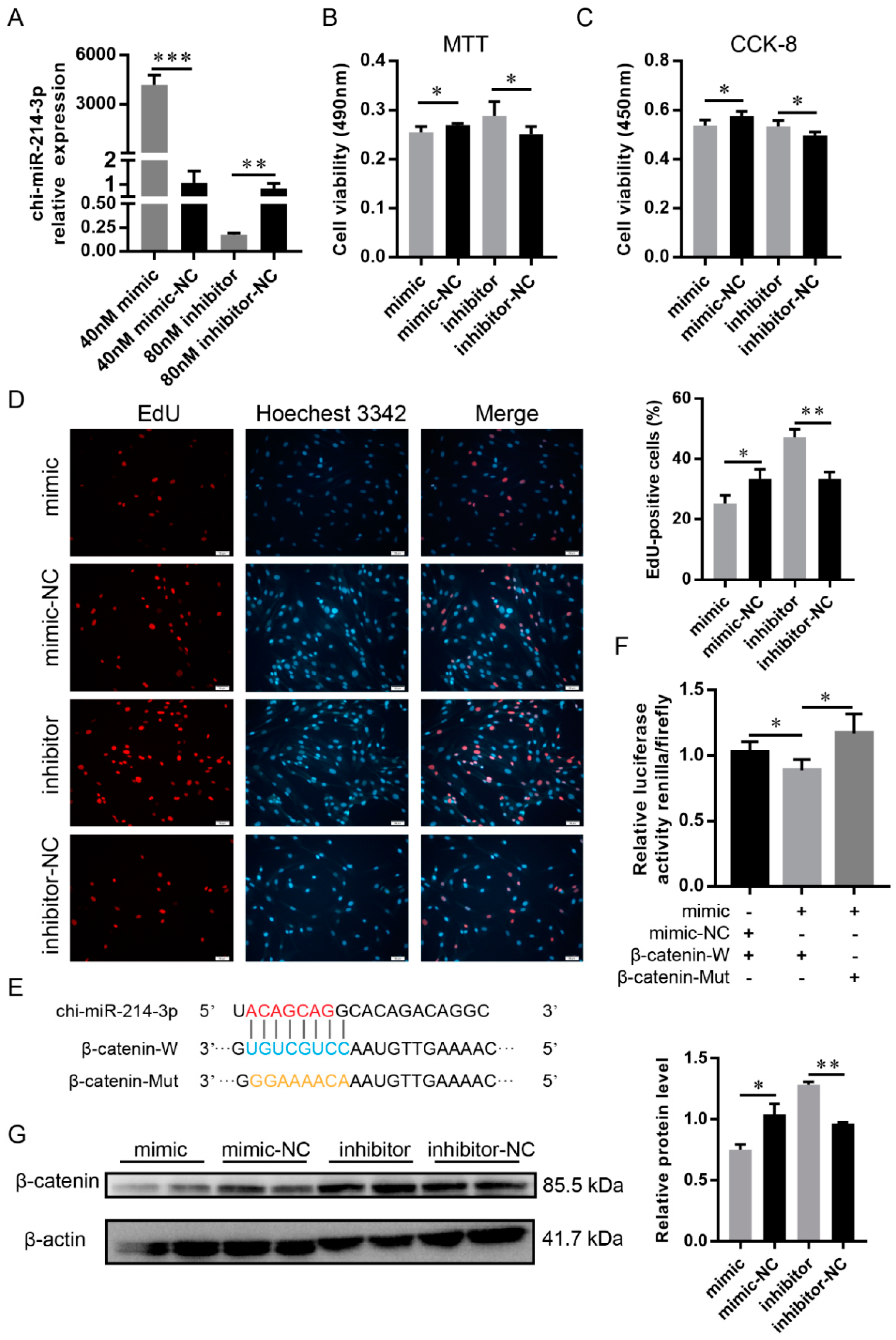
2.6. LncRNA H19 Functioned as a ceRNA for Chi-miR-214-3p
2.7. Chi-miR-214-3p Suppressed the Proliferation of DP Cells by Targeting β-Catenin
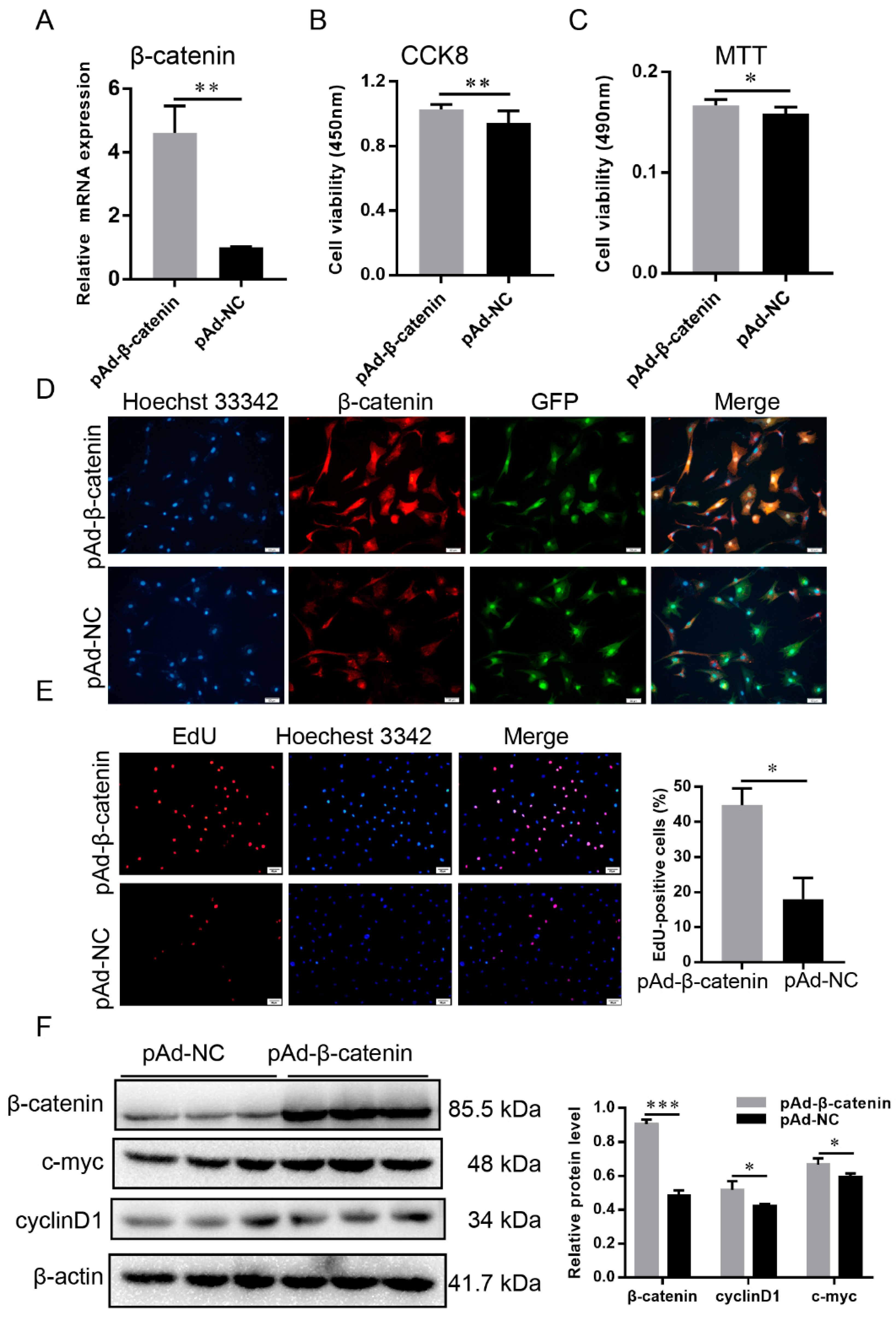
2.8. β-Catenin Promoted the Proliferation of DP Cells
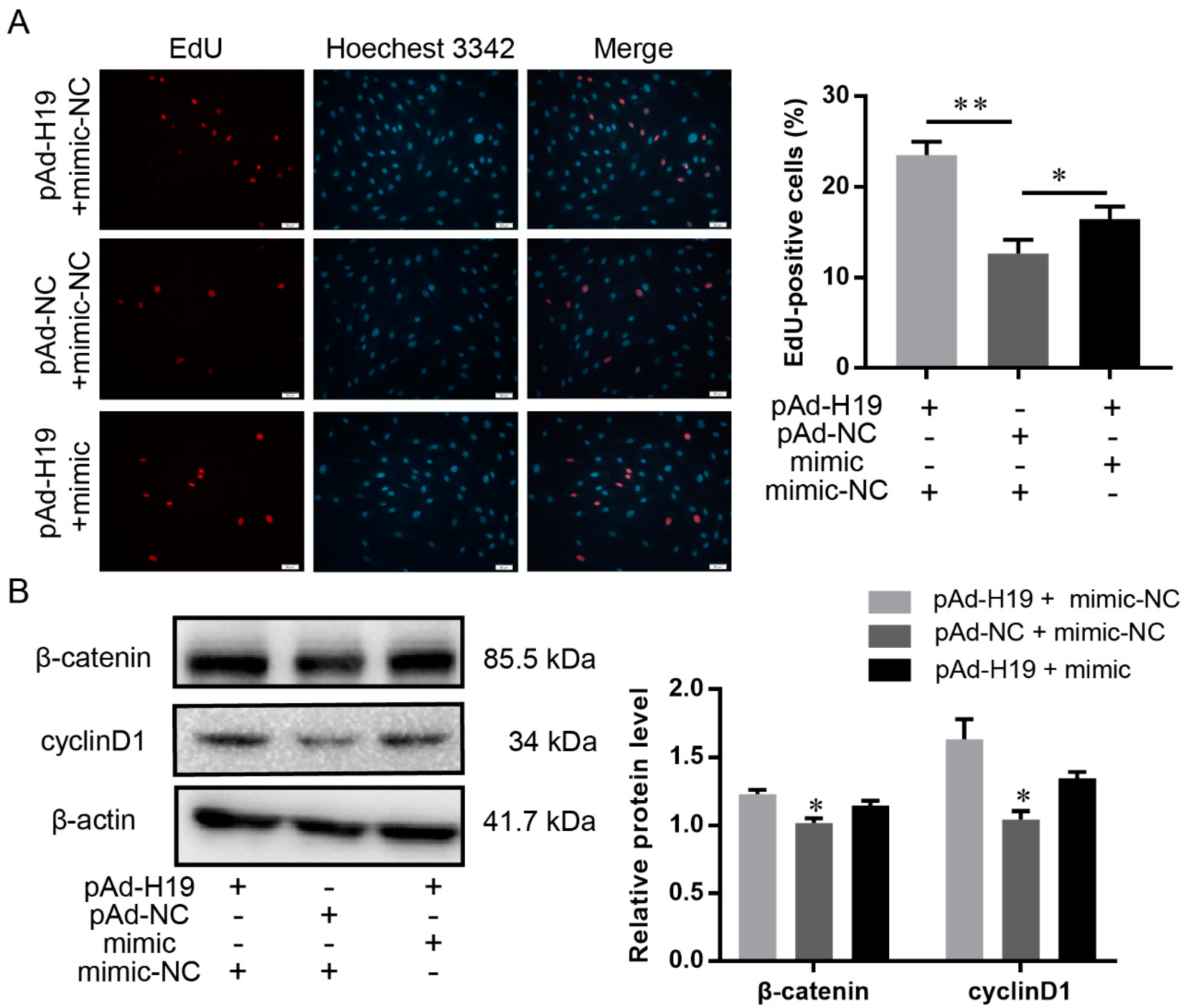
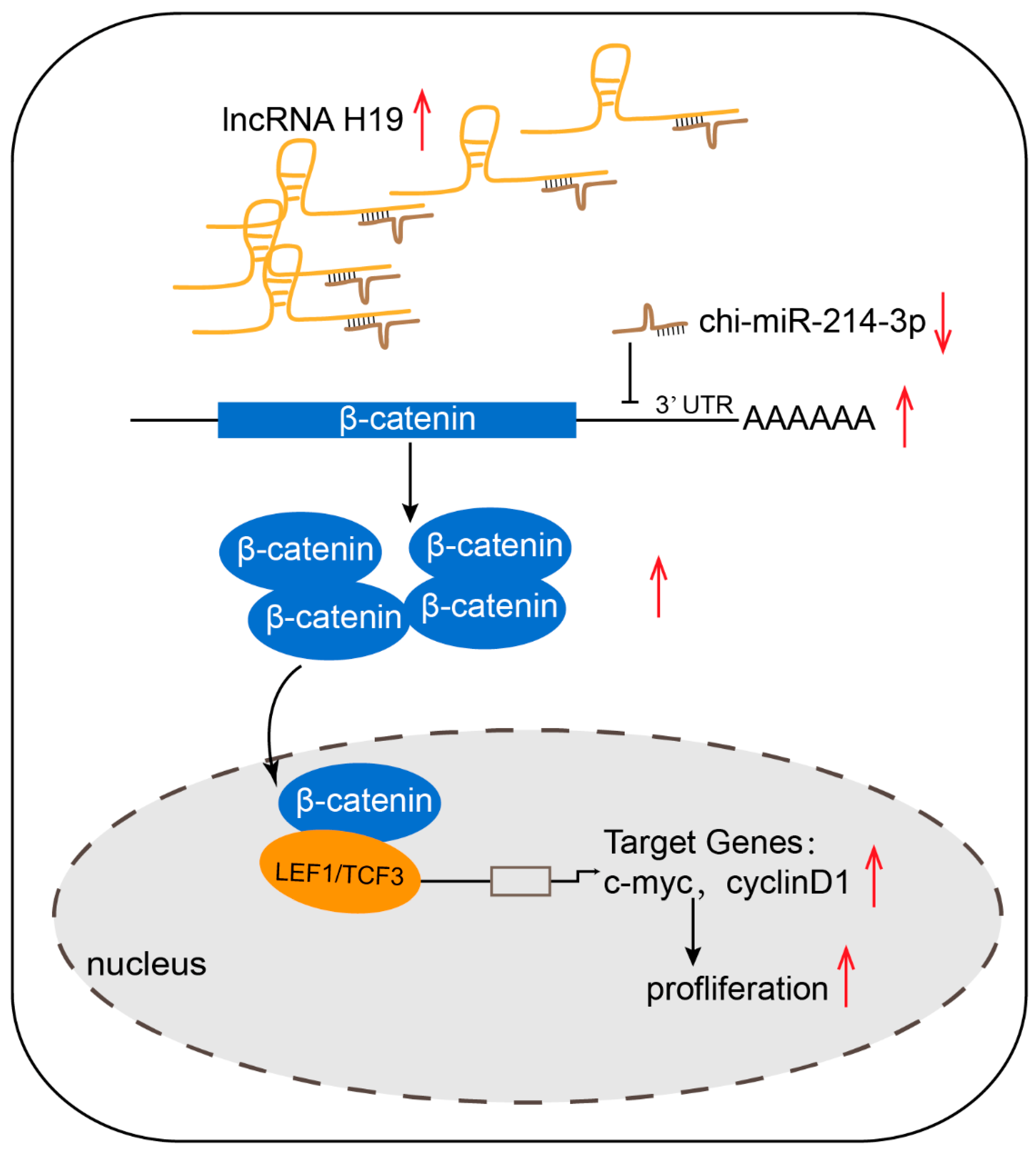
2.9. LncRNA H19 Promoted Dermal Papilla Cell Proliferation through the Chi-miR-214-3p/β-Catenin Axis
3. Discussion
3.1. Regulation of HF Development
3.2. LncRNAs and miRNAs Play an Important Role in HF Development
3.3. LncRNA Could Function as a ceRNA during HF Development
4. Materials and Methods
4.1. Animals and Samples
4.2. Transcriptome Sequencing and Bioinformatics Analysis
4.3. Validation of Gene Expression by Real-Time Quantitative PCR (RT-qPCR) Analysis
4.4. KEGG Pathway and lncRNA Target Analysis
4.5. Competing Endogenous RNA (CeRNA) Network Analysis
4.6. Vector Construction
4.7. MTT Assay
4.8. CCK-8 Assay
4.9. EdU Assay
4.10. Dual-Luciferase Assay
4.11. Western Blot
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berger, J.; Buuveibaatar, B.; Mishra, C. Globalization of the cashmere market and the decline of large mammals in central Asia. Conserv. Biol. 2013, 27, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E. Skin stem cells: Rising to the surface. J. Cell Biol. 2008, 180, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, X.; Yan, H.; Zeng, J.; Ma, S.; Niu, Y.; Zhou, G.; Jiang, Y.; Chen, Y. Comparative Transcriptome Analysis of Fetal Skin Reveals Key Genes Related to Hair Follicle Morphogenesis in Cashmere Goats. PLoS ONE 2016, 11, e0151118. [Google Scholar] [CrossRef]
- Wang, S.; Li, F.; Liu, J.; Zhang, Y.; Zheng, Y.; Ge, W.; Qu, L.; Wang, X. Integrative Analysis of Methylome and Transcriptome Reveals the Regulatory Mechanisms of Hair Follicle Morphogenesis in Cashmere Goat. Cells 2020, 9, 969. [Google Scholar] [CrossRef]
- Ge, W.; Zhang, W.; Zhang, Y.; Zheng, Y.; Li, F.; Wang, S.; Liu, J.; Tan, S.; Yan, Z.; Wang, L.; et al. A Single-cell Transcriptome Atlas of Cashmere Goat Hair Follicle Morphogenesis. Genom. Proteom. Bioinform. 2021, 19, 437–451. [Google Scholar] [CrossRef]
- Gupta, K.; Levinsohn, J.; Linderman, G.; Chen, D.; Sun, T.Y.; Dong, D.; Taketo, M.M.; Bosenberg, M.; Kluger, Y.; Choate, K.; et al. Single-Cell Analysis Reveals a Hair Follicle Dermal Niche Molecular Differentiation Trajectory that Begins Prior to Morphogenesis. Dev. Cell 2019, 48, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Duverger, O.; Morasso, M.I. Epidermal patterning and induction of different hair types during mouse embryonic development. Birth Defects Res. C Embryo Today 2009, 87, 263–272. [Google Scholar] [CrossRef]
- Chen, D.; Jarrell, A.; Guo, C.; Lang, R.; Atit, R. Dermal beta-catenin activity in response to epidermal Wnt ligands is required for fibroblast proliferation and hair follicle initiation. Development 2012, 139, 1522–1533. [Google Scholar] [CrossRef]
- Botchkarev, V.A.; Botchkareva, N.V.; Sharov, A.A.; Funa, K.; Huber, O.; Gilchrest, B.A. Modulation of BMP signaling by noggin is required for induction of the secondary (nontylotrich) hair follicles. J. Investig. Dermatol. 2002, 118, 3–10. [Google Scholar] [CrossRef]
- Huh, S.H.; Närhi, K.; Lindfors, P.H.; Häärä, O.; Yang, L.; Ornitz, D.M.; Mikkola, M.L. Fgf20 governs formation of primary and secondary dermal condensations in developing hair follicles. Genes Dev. 2013, 27, 450–458. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.Y.; Kunisada, M.; Childress, V.; Michel, M.; Schlessinger, D. Shh is required for Tabby hair follicle development. Cell Cycle 2011, 10, 3379–3386. [Google Scholar] [CrossRef] [PubMed]
- Kobielak, K.; Pasolli, H.A.; Alonso, L.; Polak, L.; Fuchs, E. Defining BMP functions in the hair follicle by conditional ablation of BMP receptor IA. J. Cell Biol. 2003, 163, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Teta, M.; Choi, Y.S.; Okegbe, T.; Wong, G.; Tam, O.H.; Chong, M.M.; Seykora, J.T.; Nagy, A.; Littman, D.R.; Andl, T.; et al. Inducible deletion of epidermal Dicer and Drosha reveals multiple functions for miRNAs in postnatal skin. Development 2012, 139, 1405–1416. [Google Scholar] [CrossRef] [PubMed]
- Andl, T.; Murchison, E.P.; Liu, F.; Zhang, Y.; Yunta-Gonzalez, M.; Tobias, J.W.; Andl, C.D.; Seykora, J.T.; Hannon, G.J.; Millar, S.E. The miRNA-processing enzyme dicer is essential for the morphogenesis and maintenance of hair follicles. Curr. Biol. 2006, 16, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.I.; Alam, M.; Emelianov, V.U.; Poterlowicz, K.; Patel, A.; Sharov, A.A.; Mardaryev, A.N.; Botchkareva, N.V. MicroRNA-214 controls skin and hair follicle development by modulating the activity of the Wnt pathway. J. Cell Biol. 2014, 207, 549–567. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Li, F.; Meng, Q.; Zhao, Y.; Chen, L.; Zhang, H.; Xue, L.; Zhang, X.; Lengner, C.; Yu, Z. Post-transcriptional Regulation of Keratinocyte Progenitor Cell Expansion, Differentiation and Hair Follicle Regression by miR-22. PLoS Genet. 2015, 11, e1005253. [Google Scholar] [CrossRef]
- Akhade, V.S.; Pal, D.; Kanduri, C. Long Noncoding RNA: Genome Organization and Mechanism of Action. Adv. Exp. Med. Biol. 2017, 1008, 47–74. [Google Scholar] [CrossRef]
- Lin, C.M.; Liu, Y.; Huang, K.; Chen, X.C.; Cai, B.Z.; Li, H.H.; Yuan, Y.P.; Zhang, H.; Li, Y. Long noncoding RNA expression in dermal papilla cells contributes to hairy gene regulation. Biochem. Biophys. Res. Commun. 2014, 453, 508–514. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, Z.; Zhu, Y.; Wang, W.; Bai, M.; Jiao, Q.; Wang, Y.; Zhao, S.; Yin, X.; Guo, D.; et al. LncRNA-000133 from secondary hair follicle of Cashmere goat: Identification, regulatory network and its effects on inductive property of dermal papilla cells. Anim. Biotechnol. 2020, 31, 122–134. [Google Scholar] [CrossRef]
- Wang, S.; Ge, W.; Luo, Z.; Guo, Y.; Jiao, B.; Qu, L.; Zhang, Z.; Wang, X. Integrated analysis of coding genes and non-coding RNAs during hair follicle cycle of cashmere goat (Capra hircus). BMC Genom. 2017, 18, 767. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Kang, D.; Ma, S.; Wang, X.; Gao, Y.; Yang, Y.; Wang, X.; Chen, Y. Integrative analysis reveals ncRNA-mediated molecular regulatory network driving secondary hair follicle regression in cashmere goats. BMC Genom. 2018, 19, 222. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Gao, Y.; Ding, Q.; Liu, J.; Li, Y.; Jin, M.; Xu, H.; Ma, S.; Wang, X.; Zeng, W.; et al. Exosomal Micro RNAs Derived from Dermal Papilla Cells Mediate Hair Follicle Stem Cell Proliferation and Differentiation. Int. J. Biol. Sci. 2019, 15, 1368–1382. [Google Scholar] [CrossRef] [PubMed]
- Hoefert, J.E.; Bjerke, G.A.; Wang, D.; Yi, R. The microRNA-200 family coordinately regulates cell adhesion and proliferation in hair morphogenesis. J. Cell Biol. 2018, 217, 2185–2204. [Google Scholar] [CrossRef]
- Wong, R.; Geyer, S.; Weninger, W.; Guimberteau, J.C.; Wong, J.K. The dynamic anatomy and patterning of skin. Exp. Dermatol. 2016, 25, 92–98. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, X.; Geng, R.; He, X.; Qu, L.; Chen, Y. Discovery of cashmere goat (Capra hircus) microRNAs in skin and hair follicles by Solexa sequencing. BMC Genom. 2013, 14, 511. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yang, F.; Zhao, M.; Ma, L.; Li, H.; Xie, Y.; Nai, R.; Che, T.; Su, R.; Zhang, Y.; et al. The intragenic mRNA-microRNA regulatory network during telogen-anagen hair follicle transition in the cashmere goat. Sci. Rep. 2018, 8, 14227. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Li, S.; Zheng, X.; Chen, W.; Li, X.; Liu, Z.; Hu, Y.; Qiao, H.; Qi, Q.; Pei, Q.; et al. Transcriptome Reveals Long Non-coding RNAs and mRNAs Involved in Primary Wool Follicle Induction in Carpet Sheep Fetal Skin. Front. Physiol. 2018, 9, 446. [Google Scholar] [CrossRef]
- Sulayman, A.; Tian, K.; Huang, X.; Tian, Y.; Xu, X.; Fu, X.; Zhao, B.; Wu, W.; Wang, D.; Yasin, A.; et al. Genome-wide identification and characterization of long non-coding RNAs expressed during sheep fetal and postnatal hair follicle development. Sci. Rep. 2019, 9, 8501. [Google Scholar] [CrossRef]
- Schneider, M.R.; Schmidt-Ullrich, R.; Paus, R. The hair follicle as a dynamic miniorgan. Curr. Biol. 2009, 19, R132–R142. [Google Scholar] [CrossRef]
- Reddy, S.; Andl, T.; Bagasra, A.; Lu, M.M.; Epstein, D.J.; Morrisey, E.E.; Millar, S.E. Characterization of Wnt gene expression in developing and postnatal hair follicles and identification of Wnt5a as a target of Sonic hedgehog in hair follicle morphogenesis. Mech. Dev. 2001, 107, 69–82. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Pispa, J.; Hartung, A.J.; Du, Y.; Ezer, S.; Jenks, T.; Shimada, T.; Pekkanen, M.; Mikkola, M.L.; Ko, M.S.; et al. The Tabby phenotype is caused by mutation in a mouse homologue of the EDA gene that reveals novel mouse and human exons and encodes a protein (ectodysplasin-A) with collagenous domains. Proc. Natl. Acad. Sci. USA 1997, 94, 13069–13074. [Google Scholar] [CrossRef]
- Lin, M.H.; Leimeister, C.; Gessler, M.; Kopan, R. Activation of the Notch pathway in the hair cortex leads to aberrant differentiation of the adjacent hair-shaft layers. Development 2000, 127, 2421–2432. [Google Scholar] [CrossRef]
- Sellheyer, K.; Krahl, D. Expression pattern of GATA-3 in embryonic and fetal human skin suggests a role in epidermal and follicular morphogenesis. J. Cutan. Pathol. 2010, 37, 357–361. [Google Scholar] [CrossRef]
- Kaufman, C.K.; Zhou, P.; Pasolli, H.A.; Rendl, M.; Bolotin, D.; Lim, K.C.; Dai, X.; Alegre, M.L.; Fuchs, E. GATA-3: An unexpected regulator of cell lineage determination in skin. Genes Dev. 2003, 17, 2108–2122. [Google Scholar] [CrossRef]
- Potter, C.S.; Peterson, R.L.; Barth, J.L.; Pruett, N.D.; Jacobs, D.F.; Kern, M.J.; Argraves, W.S.; Sundberg, J.P.; Awgulewitsch, A. Evidence that the satin hair mutant gene Foxq1 is among multiple and functionally diverse regulatory targets for Hoxc13 during hair follicle differentiation. J. Biol. Chem. 2006, 281, 29245–29255. [Google Scholar] [CrossRef]
- Hong, H.K.; Noveroske, J.K.; Headon, D.J.; Liu, T.; Sy, M.S.; Justice, M.J.; Chakravarti, A. The winged helix/forkhead transcription factor Foxq1 regulates differentiation of hair in satin mice. Genesis 2001, 29, 163–171. [Google Scholar] [CrossRef]
- Tkatchenko, A.V.; Visconti, R.P.; Shang, L.; Papenbrock, T.; Pruett, N.D.; Ito, T.; Ogawa, M.; Awgulewitsch, A. Overexpression of Hoxc13 in differentiating keratinocytes results in downregulation of a novel hair keratin gene cluster and alopecia. Development 2001, 128, 1547–1558. [Google Scholar] [CrossRef]
- Cai, J.; Lee, J.; Kopan, R.; Ma, L. Genetic interplays between Msx2 and Foxn1 are required for Notch1 expression and hair shaft differentiation. Dev. Biol. 2009, 326, 420–430. [Google Scholar] [CrossRef]
- Wang, C.; Wang, L.; Ding, Y.; Lu, X.; Zhang, G.; Yang, J.; Zheng, H.; Wang, H.; Jiang, Y.; Xu, L. LncRNA Structural Characteristics in Epigenetic Regulation. Int. J. Mol. Sci. 2017, 18, 2659. [Google Scholar] [CrossRef]
- Zhu, Y.B.; Wang, Z.Y.; Yin, R.H.; Jiao, Q.; Zhao, S.J.; Cong, Y.Y.; Xue, H.L.; Guo, D.; Wang, S.Q.; Zhu, Y.X.; et al. A lncRNA-H19 transcript from secondary hair follicle of Liaoning cashmere goat: Identification, regulatory network and expression regulated potentially by its promoter methylation. Gene 2018, 641, 78–85. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef]
- Zhang, Y.; Xia, S.; Wang, T.; Wang, S.; Yuan, D.; Li, F.; Wang, X. Chi-miR-30b-5p inhibits dermal papilla cells proliferation by targeting CaMKIIδ gene in cashmere goat. BMC Genom. 2020, 21, 430. [Google Scholar] [CrossRef]
- Wang, S.; Luo, Z.; Zhang, Y.; Yuan, D.; Ge, W.; Wang, X. The inconsistent regulation of HOXC13 on different keratins and the regulation mechanism on HOXC13 in cashmere goat (Capra hircus). BMC Genom. 2018, 19, 630. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Known | Novel | Known Different | Novel Different | |
|---|---|---|---|---|
| mRNAs | 40,364 | 275 | 5940 | 36 |
| lncRNAs | 2227 | 8258 | 91 | 430 |
| miRNAs | 422 | 183 | 180 | 24 |
| Transcript_id | E65_FPKM/TPM | E90_FPKM/TPM | E120_FPKM/TPM |
|---|---|---|---|
| lncRNAs | |||
| H19 | 843.0740 | 941.2135 | 114.6905 |
| TCONS_00573233 | 508.9460 | 37.8758 | 380.9373 |
| XR_001296756.2 | 0.4040 | 4.7004 | 133.6244 |
| TCONS_00790678 | 23.2770 | 0.0000 | 99.5389 |
| TCONS_00796832 | 1.0006 | 101.5070 | 1.8325 |
| TCONS_00794094 | 12.0705 | 0.0000 | 76.5603 |
| TCONS_00267940 | 0.2418 | 82.8900 | 0.9865 |
| TCONS_00799045 | 5.7793 | 0.0000 | 37.3490 |
| TCONS_00021925 | 0.0205 | 0.0074 | 30.0650 |
| TCONS_00267941 | 0.0122 | 27.4923 | 0.0000 |
| TCONS_00462852 | 0.2009 | 0.6194 | 24.8990 |
| XR_001297629.2 | 0.0161 | 0.0000 | 23.0225 |
| TCONS_00579021 | 15.4707 | 0.1352 | 4.9993 |
| XR_001917911.1 | 0.0000 | 0.6697 | 16.3470 |
| TCONS_00794114 | 0.0000 | 15.9658 | 0.0000 |
| miRNAs | |||
| chi-miR-143-3p | 67,656.2395 | 53,405.3408 | 143,004.1167 |
| chi-miR-26a-5p | 56,081.5294 | 69,262.7051 | 71,726.3097 |
| chi-miR-27b-3p | 18,121.1064 | 31,704.0191 | 48,005.4398 |
| novel_1 | 9700.2197 | 21,113.9625 | 51,116.4677 |
| chi-let-7f-5p | 18,722.4699 | 24,936.8240 | 29,656.2387 |
| chi-miR-100-5p | 34,055.8992 | 17,383.0249 | 13,167.1150 |
| chi-miR-411a-5p | 27,666.1047 | 25,927.8426 | 9216.2797 |
| chi-let-7g-5p | 15,255.1331 | 17,077.0222 | 21,381.1897 |
| chi-let-7a-5p | 9843.2445 | 13,049.8691 | 22,008.5678 |
| chi-miR-126-3p | 13,413.4404 | 10,380.2313 | 19,570.4819 |
| chi-miR-379-5p | 16,120.5727 | 13,927.9794 | 6478.6473 |
| chi-miR-24-3p | 6192.5776 | 8893.4012 | 13,070.6241 |
| chi-miR-378-3p | 14,459.7160 | 7918.5068 | 4337.4918 |
| chi-let-7b-5p | 3520.0832 | 6329.7777 | 11,805.9702 |
| chi-miR-200b | 4858.8871 | 5182.6072 | 11,212.8627 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Li, F.; Shi, Y.; Zhang, T.; Wang, X. Comprehensive Transcriptome Analysis of Hair Follicle Morphogenesis Reveals That lncRNA-H19 Promotes Dermal Papilla Cell Proliferation through the Chi-miR-214-3p/β-Catenin Axis in Cashmere Goats. Int. J. Mol. Sci. 2022, 23, 10006. https://doi.org/10.3390/ijms231710006
Zhang Y, Li F, Shi Y, Zhang T, Wang X. Comprehensive Transcriptome Analysis of Hair Follicle Morphogenesis Reveals That lncRNA-H19 Promotes Dermal Papilla Cell Proliferation through the Chi-miR-214-3p/β-Catenin Axis in Cashmere Goats. International Journal of Molecular Sciences. 2022; 23(17):10006. https://doi.org/10.3390/ijms231710006
Chicago/Turabian StyleZhang, Yuelang, Fang Li, Yujie Shi, Tongtong Zhang, and Xin Wang. 2022. "Comprehensive Transcriptome Analysis of Hair Follicle Morphogenesis Reveals That lncRNA-H19 Promotes Dermal Papilla Cell Proliferation through the Chi-miR-214-3p/β-Catenin Axis in Cashmere Goats" International Journal of Molecular Sciences 23, no. 17: 10006. https://doi.org/10.3390/ijms231710006
APA StyleZhang, Y., Li, F., Shi, Y., Zhang, T., & Wang, X. (2022). Comprehensive Transcriptome Analysis of Hair Follicle Morphogenesis Reveals That lncRNA-H19 Promotes Dermal Papilla Cell Proliferation through the Chi-miR-214-3p/β-Catenin Axis in Cashmere Goats. International Journal of Molecular Sciences, 23(17), 10006. https://doi.org/10.3390/ijms231710006