
The VEGF Inhibitor Soluble Fms-like Tyrosine Kinase 1 Does Not Promote AKI-to-CKD Transition

 , , and
, , and
Abstract
:1. Introduction
2. Results
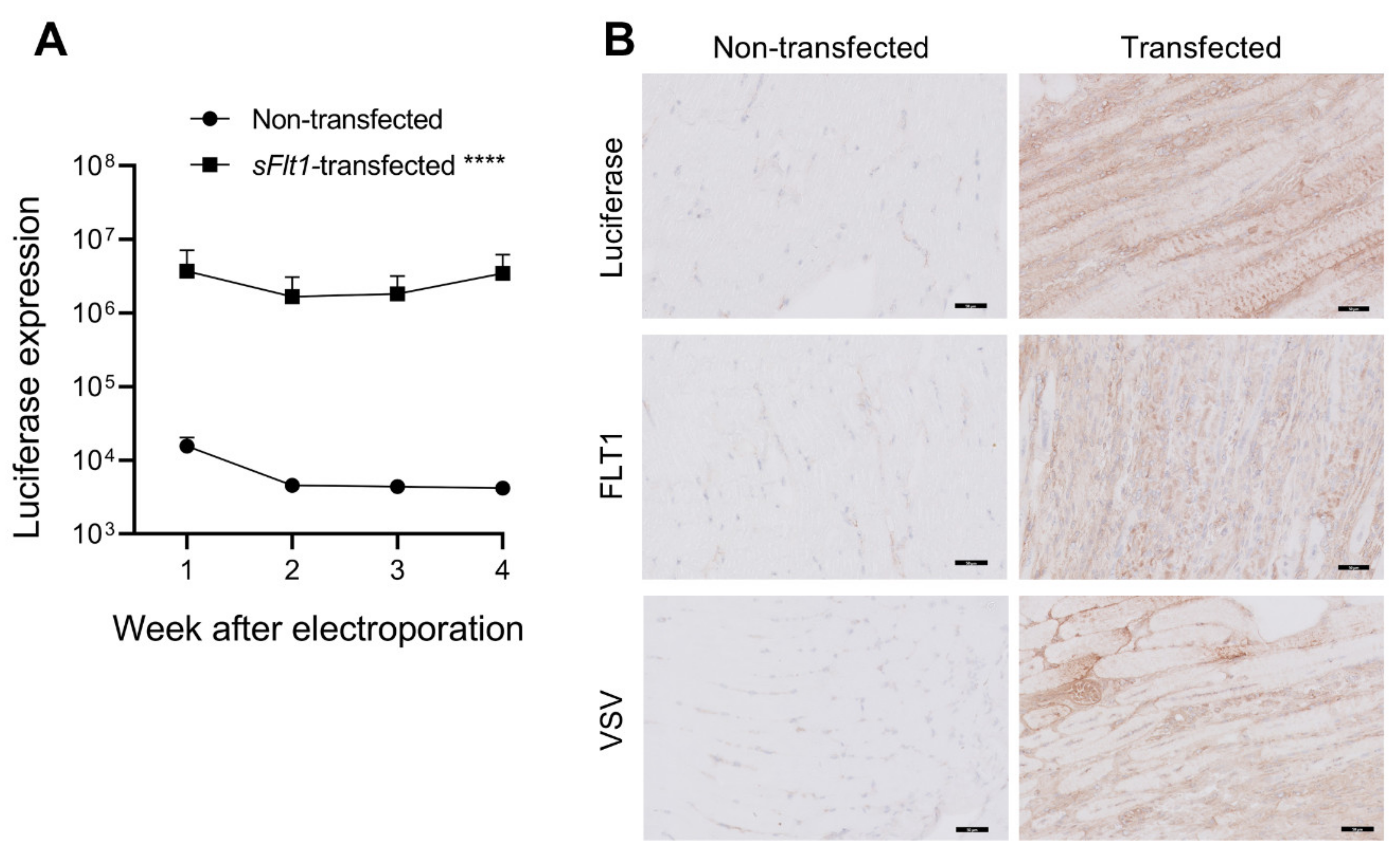
2.1. Gene Delivery of sFlt1-VSV and Luciferase
2.2. sFLT1 Does Not Aggravate Acute and Chronic Kidney Damage following IRI
2.3. sFLT1 Has No Effect on Peritubular Capillary Loss after IRI
2.4. sFLT1 Treatment Does Not Alter Macrophage Infiltration after IRI
2.5. Transfection Efficiency Correlates with Reduced Kidney Damage and Inflammation
2.6. Renal sFLT1 mRNA Expression Is Not Associated with Human CKD
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. sFlt1-VSV Transfection
4.3. Unilateral IRI Model
4.4. ELISA of Serum Kidney Injury Molecule-1
4.5. Histology and Immunohistochemistry
4.6. Digital Image Analysis
4.7. Quantitative PCR
4.8. Human CKD Biopsy Cohort
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cudmore, M.J.; Hewett, P.W.; Ahmad, S.; Wang, K.-Q.; Cai, M.; Al-Ani, B.; Fujisawa, T.; Ma, B.; Sissaoui, S.; Ramma, W.; et al. The role of heterodimerization between VEGFR-1 and VEGFR-2 in the regulation of endothelial cell homeostasis. Nat. Commun. 2012, 3, 972. [Google Scholar] [CrossRef] [PubMed]
- Ambati, B.K.; Nozaki, M.; Singh, N.; Takeda, A.; Jani, P.D.; Suthar, T.; Albuquerque, R.J.; Richter, E.; Sakurai, E.; Newcomb, M.T.; et al. Corneal avascularity is due to soluble VEGF receptor-1. Nature 2006, 443, 993–997. [Google Scholar] [CrossRef] [PubMed]
- Chappell, J.C.; Taylor, S.M.; Ferrara, N.; Bautch, V.L. Local Guidance of Emerging Vessel Sprouts Requires Soluble Flt-1. Dev. Cell 2009, 17, 377–386. [Google Scholar] [CrossRef]
- Jin, J.; Sison, K.; Li, C.; Tian, R.; Wnuk, M.; Sung, H.-K.; Jeansson, M.; Zhang, C.; Tucholska, M.; Jones, N.; et al. Soluble FLT1 Binds Lipid Microdomains in Podocytes to Control Cell Morphology and Glomerular Barrier Function. Cell 2012, 151, 384–399. [Google Scholar] [CrossRef]
- Seno, A.; Takeda, Y.; Matsui, M.; Okuda, A.; Nakano, T.; Nakada, Y.; Kumazawa, T.; Nakagawa, H.; Nishida, T.; Onoue, K.; et al. Suppressed Production of Soluble Fms-Like Tyrosine Kinase-1 Contributes to Myocardial Remodeling and Heart Failure. Hypertension 2016, 68, 678–687. [Google Scholar] [CrossRef]
- Bus, P.; Scharpfenecker, M.; Van Der Wilk, P.; Wolterbeek, R.; Bruijn, J.A.; Baelde, H.J. The VEGF-A inhibitor sFLT-1 improves renal function by reducing endothelial activation and inflammation in a mouse model of type 1 diabetes. Diabetologia 2017, 60, 1813–1821. [Google Scholar] [CrossRef]
- Onoue, K.; Uemura, S.; Takeda, Y.; Somekawa, S.; Iwama, H.; Imagawa, K.; Nishida, T.; Morikawa, Y.; Takemoto, Y.; Asai, O.; et al. Reduction of Circulating Soluble Fms-Like Tyrosine Kinase-1 Plays a Significant Role in Renal Dysfunction—Associated Aggravation of Atherosclerosis. Circulation 2009, 120, 2470–2477. [Google Scholar] [CrossRef]
- Tsao, P.N.; Chan, F.T.; Wei, S.C.; Hsieh, W.S.; Chou, H.C.; Su, Y.N.; Chen, C.Y.; Hsu, W.M.; Hsieh, F.J.; Hsu, S.M. Soluble vascular endothelial growth factor receptor-1 protects mice in sepsis. Crit. Care Med. 2007, 35, 1955–1960. [Google Scholar] [CrossRef] [PubMed]
- Biscetti, F.; Flex, A.; Pecorini, G.; Angelini, F.; Arena, V.; Stigliano, E.; Gremese, E.; Tolusso, B.; Ferraccioli, G. The role of high-mobility group box protein 1 in collagen antibody-induced arthritis is dependent on vascular endothelial growth factor. Clin. Exp. Immunol. 2016, 184, 62–72. [Google Scholar] [CrossRef]
- van Aanhold, C.; Bus, P.; Zandbergen, M.; Bos, M.; Berbée, J.; Quint, K.; Bruijn, J.; Baelde, H. The VEGF Inhibitor Soluble FLT-1 Ameliorates Atopic Dermatitis in APOC1 Transgenic Mice. J. Investig. Dermatol. 2019, 140, 491–494.e4. [Google Scholar] [CrossRef]
- Levine, R.J.; Maynard, S.E.; Qian, C.; Lim, K.-H.; England, L.J.; Yu, K.F.; Schisterman, E.F.; Thadhani, R.; Sachs, B.P.; Epstein, F.H.; et al. Circulating Angiogenic Factors and the Risk of Preeclampsia. N. Engl. J. Med. 2004, 350, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Shye, M.; Hanna, R.M.; Patel, S.S.; Tram-Tran, N.; Hou, J.; Mccannel, C.; Khalid, M.; Hanna, M.; Abdelnour, L.; Kurtz, I. Worsening proteinuria and renal function after intravitreal vascular endothelial growth factor blockade for diabetic proliferative retinopathy. Clin. Kidney J. 2020, 13, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Eremina, V.; Jefferson, J.A.; Kowalewska, J.; Hochster, H.; Haas, M.; Weisstuch, J.; Richardson, C.; Kopp, J.B.; Kabir, M.G.; Backx, P.H.; et al. VEGF inhibition and renal thrombotic microangiopathy. N. Engl. J. Med. 2008, 358, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Wewers, T.M.; Schulz, A.; Nolte, I.; Pavenstädt, H.; Brand, M.; Di Marco, G.S. Circulating Soluble Fms-like Tyrosine Kinase in Renal Diseases Other than Preeclampsia. J. Am. Soc. Nephrol. 2021, 32, 1853–1863. [Google Scholar] [CrossRef]
- Chapal, M.; Néel, M.; Le Borgne, F.; Meffray, E.; Carceles, O.; Hourmant, M.; Giral, M.; Foucher, Y.; Moreau, A.; Fakhouri, F. Increased Soluble Flt-1 Correlates With Delayed Graft Function and Early Loss of Peritubular Capillaries in the Kidney Graft. Transplantation 2013, 96, 739–744. [Google Scholar] [CrossRef]
- Wewers, T.M.; Mayer, A.B.; Pfleiderer, A.; Beul, K.; Schmidt, R.; Heitplatz, B.; Van Marck, V.; Nolte, I.; Pavenstädt, H.; Reuter, S.; et al. Increased soluble fms-like tyrosine kinase 1 after ischemia reperfusion contributes to adverse clinical outcomes following kidney transplantation. Kidney Int. 2019, 95, 1091–1102. [Google Scholar] [CrossRef]
- Di Marco, G.S.; Reuter, S.; Hillebrand, U.; Amler, S.; König, M.; Larger, E.; Oberleithner, H.; Brand, E.; Pavenstädt, H.; Brand, M. The soluble VEGF receptor sFlt1 contributes to endothelial dysfunction in CKD. J. Am. Soc. Nephrol. 2009, 20, 2235–2245. [Google Scholar] [CrossRef]
- Baelde, H.J.; Eikmans, M.; Lappin, D.W.P.; Doran, P.P.; Hohenadel, D.; Brinkkoetter, P.T.; van der Woude, F.J.; Waldherr, R.; Rabelink, T.J.; de Heer, E.; et al. Reduction of VEGF-A and CTGF expression in diabetic nephropathy is associated with podocyte loss. Kidney Int. 2007, 71, 637–645. [Google Scholar] [CrossRef]
- Yang, S.-M.; Ka, S.-M.; Wu, H.-L.; Yeh, Y.-C.; Kuo, C.-H.; Hua, K.-F.; Shi, G.-Y.; Hung, Y.-J.; Hsiao, F.-C.; Yang, S.-S.; et al. Thrombomodulin domain 1 ameliorates diabetic nephropathy in mice via anti-NF-κB/NLRP3 inflammasome-mediated inflammation, enhancement of NRF2 antioxidant activity and inhibition of apoptosis. Diabetologia 2014, 57, 424–434. [Google Scholar] [CrossRef]
- Masola, V.; Gambaro, G.; Tibaldi, E.; Brunati, A.M.; Gastaldello, A.; D’Angelo, A.; Onisto, M.; Lupo, A. Heparanase and syndecan-1 interplay orchestrates fibroblast growth factor-2-induced epithelial-mesenchymal transition in renal tubular cells. J. Biol. Chem. 2012, 287, 1478–1488. [Google Scholar] [CrossRef] [Green Version]
- Bus, P.; Pierneef, L.; Bor, R.; Wolterbeek, R.; van Es, L.A.; Rensen, P.C.; de Heer, E.; Havekes, L.M.; Bruijn, J.A.; Berbée, J.F.; et al. Apolipoprotein C-I plays a role in the pathogenesis of glomerulosclerosis. J. Pathol. 2017, 241, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, H.; Hamano, Y.; Charytan, D.; Cosgrove, D.; Kieran, M.; Sudhakar, A.; Kalluri, R. Neutralization of Circulating Vascular Endothelial Growth Factor (VEGF) by Anti-VEGF Antibodies and Soluble VEGF Receptor 1 (sFlt-1) Induces Proteinuria. J. Biol. Chem. 2003, 278, 12605–12608. [Google Scholar] [CrossRef] [PubMed]
- Holash, J.; Davis, S.; Papadopoulos, N.; Croll, S.D.; Ho, L.; Russell, M.; Boland, P.; Leidich, R.; Hylton, D.; Burova, E.; et al. VEGF-Trap: A VEGF blocker with potent antitumor effects. Proc. Natl. Acad. Sci. USA 2002, 99, 11393–11398. [Google Scholar] [CrossRef]
- Markovic-Mueller, S.; Stuttfeld, E.; Asthana, M.; Weinert, T.; Bliven, S.; Goldie, K.N.; Kisko, K.; Capitani, G.; Ballmer-Hofer, K. Structure of the Full-length VEGFR-1 Extracellular Domain in Complex with VEGF-A. Structure 2017, 25, 341–352. [Google Scholar] [CrossRef]
- Lin, S.-L.; Chang, F.-C.; Schrimpf, C.; Chen, Y.-T.; Wu, C.-F.; Wu, V.-C.; Chiang, W.-C.; Kuhnert, F.; Kuo, C.J.; Chen, Y.-M.; et al. Targeting endothelium-pericyte cross talk by inhibiting VEGF receptor signaling attenuates kidney microvascular rarefaction and fibrosis. Am. J. Pathol. 2011, 178, 911–923. [Google Scholar] [CrossRef]
- Matsui, M.; Takeda, Y.; Uemura, S.; Matsumoto, T.; Seno, A.; Onoue, K.; Tsushima, H.; Morimoto, K.; Soeda, T.; Okayama, S.; et al. Suppressed soluble Fms-like tyrosine kinase-1 production aggravates atherosclerosis in chronic kidney disease. Kidney Int. 2014, 85, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, O.; Afsar, B.; Ortiz, A.; Kanbay, M. The role of endothelial glycocalyx in health and disease. Clin. Kidney J. 2019, 12, 611–619. [Google Scholar] [CrossRef]
- Wei, Q.; Dong, Z. Mouse model of ischemic acute kidney injury: Technical notes and tricks. Am. J. Physiol. Ren. Physiol. 2012, 303, F1487–F1494. [Google Scholar] [CrossRef]
- Eefting, D.; Grimbergen, J.M.; de Vries, M.R.; van Weel, V.; Kaijzel, E.L.; Que, I.; Moon, R.T.; Löwik, C.W.; van Bockel, J.H.; Quax, P.H.A. Prolonged In Vivo Gene Silencing by Electroporation-Mediated Plasmid Delivery of Small Interfering RNA. Hum. Gene Ther. 2007, 18, 861–869. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bivariate Correlation with Luciferase Expression | ||
|---|---|---|
| Pearson’s r | p-Value | |
| Serum KIM-1 day 2 | 0.626 | 0.071 |
| Kidney size (mm) | 0.394 | 0.295 |
| Outer medulla Sirius red | −0.514 | 0.157 |
| Outer medulla endomucin | 0.053 | 0.893 |
| Outer medulla F4/80 | −0.091 | 0.817 |
| Col1a1 | −0.751 | 0.020 |
| Col4a2 | −0.756 | 0.018 |
| Fn1 | −0.759 | 0.018 |
| Pgf | −0.889 | 0.001 |
| Adgre1 | −0.863 | 0.003 |
| Mrc1 | −0.884 | 0.002 |
| Il1b | 0.074 | 0.850 |
| Vegfa | −0.667 | 0.050 |
| Vcam1 | −0.839 | 0.005 |
| Target | Forward | Reverse |
|---|---|---|
| Hprt | AGATGGTCAAGGTCGCAAGC | TCAAGGGCATATCCTACAACAAAC |
| Col1a1 | TGACTGGAAGAGCGGAGAGT | AGACGGCTGAGTAGGGAACA |
| Fn1 | GGCAGGCTCAGCAAATCG | CATAGCAGGTACAAACCAGGG |
| Col4a1 | CTGGGATCATGGACCGAGTG | CCTTTCTCCGGGTAGCACTG |
| Vegfa | CTGGACCCTGGCTTTACTGC’ | GCTTCGCTGGTAGACATCCA |
| Pgf | TTCTGGAGACGACAAAGGCA | GCTGGTTACCTCCGGGAAAT |
| Adgre1 | GGCAGGGATCTTGGTTATGCT | GCTGCACTCTGTAAGGACACT |
| Mrc1 | GCTGGCGAGCATCAAGAGTA | CATCACTCCAGGTGAACCCC |
| Vcam1 | GAAATGCCACCCTCACCTTA | ACGTCAGAACAACCGAATCC |
| GAPDH | CGACCACTTTGTCAAGCTCA | AGGGGTCTACATGGCAACTG |
| sFLT1-i13 | ACAATCAGAGGTGAGCACTGCAA | TCCGAGCCTGAAAGTTAGCAA |
| sFLT1-e15a | CGAGCCTCAGATCACTTGGT | CGATGACGATGGTGACGTT |
| THBD | ACATCCTGGACGACGGTTTC | CGCAGATGCACTCGAAGGTA |
| SDC1 | TGCCGCAAATTGTGGCTACTAAT | GAGCTGCGTGTCCTTCCAAG |
| Clinical Characteristics | Controls (n = 10) | CKD (n = 30) | p Value |
|---|---|---|---|
| Age (years) | 58.3 ± 14.2 | 59.9 ± 9.8 | 0.698 |
| Female, n (%) | 3 (30) | 15 (50) | 0.271 |
| Hypertension, n (%) | 3/7 (43) | 14/20 (70) | 0.201 |
| Systolic blood pressure, mmHg | 135 ± 31 | 137 ± 17 | 0.884 |
| Diastolic blood pressure, mmHg | 74 ± 16 | 80 ± 10 | 0.329 |
| Serum creatinine, mmol/L | 81 ± 48 | 202 ± 138 | 0.015 |
| Proteinuria, g/L | 0.03 ± 0.05 | 3.7 ± 1.9 | <0.001 |
| Histopathological characteristics | Controls (n = 10) | CKD (n = 28) | p value |
| Glomerulosclerosis, n (%) | |||
| Absent | 10 (100) | 0 | |
| Diffuse | 11 (39) | ||
| Diffuse and nodular | 2 (7) | ||
| Nodular | 15 (54) | ||
| FSGS | 1 (4) | ||
| Tubular atrophy | 13 (46) | ||
| Interstitial fibrosis | 12 (43) | ||
| Arteriosclerosis | 14 (50) | ||
| Arteriolar hyalinosis | 2 (7) | ||
| IgA GN | 2 (7) | ||
| Membranous GN | 1 (4) | ||
| Interstitial Sirius red (%) | 19.5 ± 7.8 | 33.3 ± 9.8 | <0.001 |
| Interstitial CD31 (%) | 9.6 ± 2.2 | 5.5 ± 3.4 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Aanhold, C.C.L.; Koudijs, A.; Dijkstra, K.L.; Wolterbeek, R.; Bruijn, J.A.; van Kooten, C.; Baelde, H.J. The VEGF Inhibitor Soluble Fms-like Tyrosine Kinase 1 Does Not Promote AKI-to-CKD Transition. Int. J. Mol. Sci. 2022, 23, 9660. https://doi.org/10.3390/ijms23179660
van Aanhold CCL, Koudijs A, Dijkstra KL, Wolterbeek R, Bruijn JA, van Kooten C, Baelde HJ. The VEGF Inhibitor Soluble Fms-like Tyrosine Kinase 1 Does Not Promote AKI-to-CKD Transition. International Journal of Molecular Sciences. 2022; 23(17):9660. https://doi.org/10.3390/ijms23179660
Chicago/Turabian Stylevan Aanhold, Cleo C. L., Angela Koudijs, Kyra L. Dijkstra, Ron Wolterbeek, Jan A. Bruijn, Cees van Kooten, and Hans J. Baelde. 2022. "The VEGF Inhibitor Soluble Fms-like Tyrosine Kinase 1 Does Not Promote AKI-to-CKD Transition" International Journal of Molecular Sciences 23, no. 17: 9660. https://doi.org/10.3390/ijms23179660
APA Stylevan Aanhold, C. C. L., Koudijs, A., Dijkstra, K. L., Wolterbeek, R., Bruijn, J. A., van Kooten, C., & Baelde, H. J. (2022). The VEGF Inhibitor Soluble Fms-like Tyrosine Kinase 1 Does Not Promote AKI-to-CKD Transition. International Journal of Molecular Sciences, 23(17), 9660. https://doi.org/10.3390/ijms23179660