
Nutritional Component Analyses in Different Varieties of Actinidia eriantha Kiwifruit by Transcriptomic and Metabolomic Approaches

 and
and
Abstract
:1. Introduction
2. Results
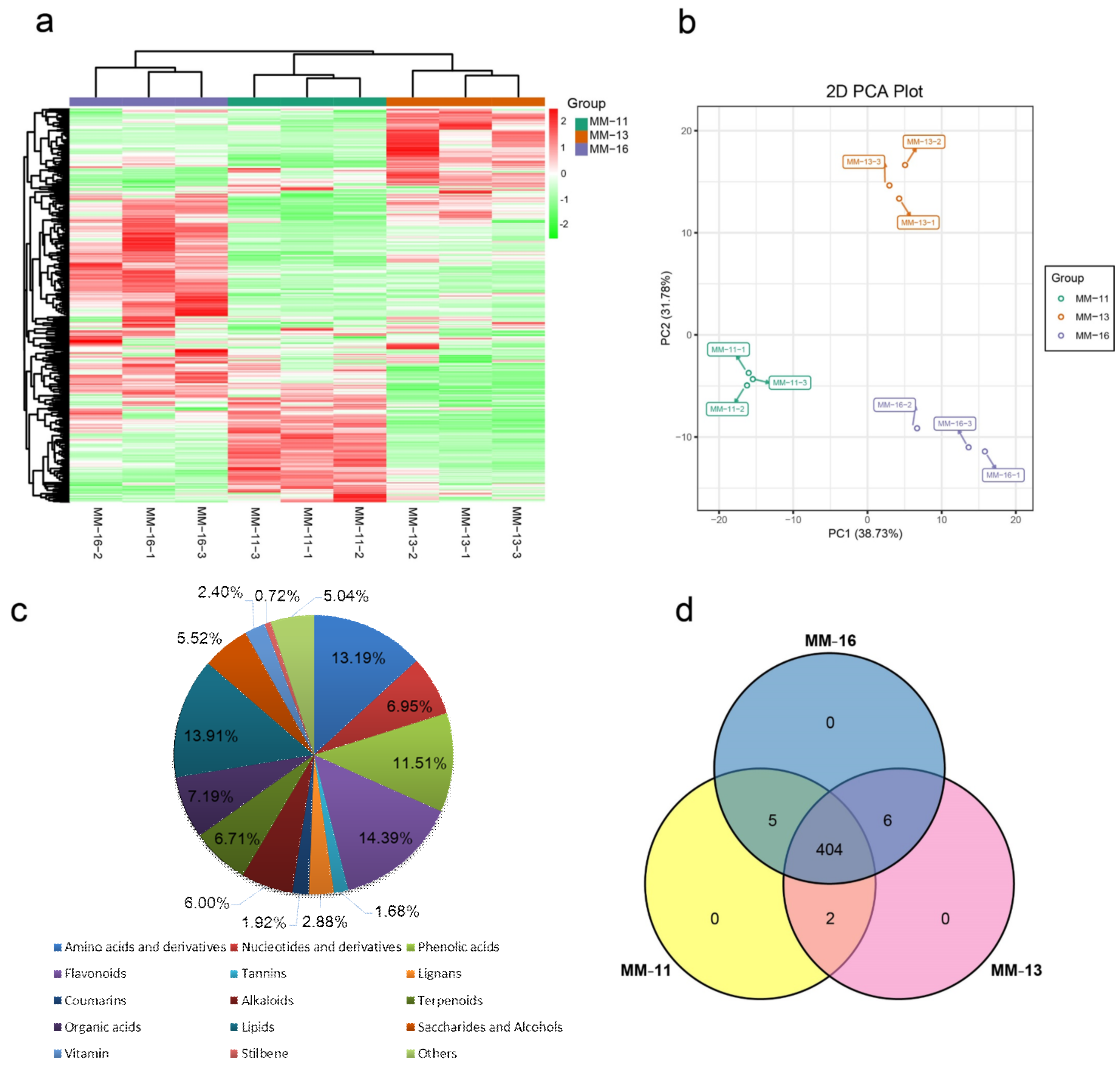
2.1. The Main Nutritional Components of A. eriantha Fruit Detected by Metabolomic Analyses
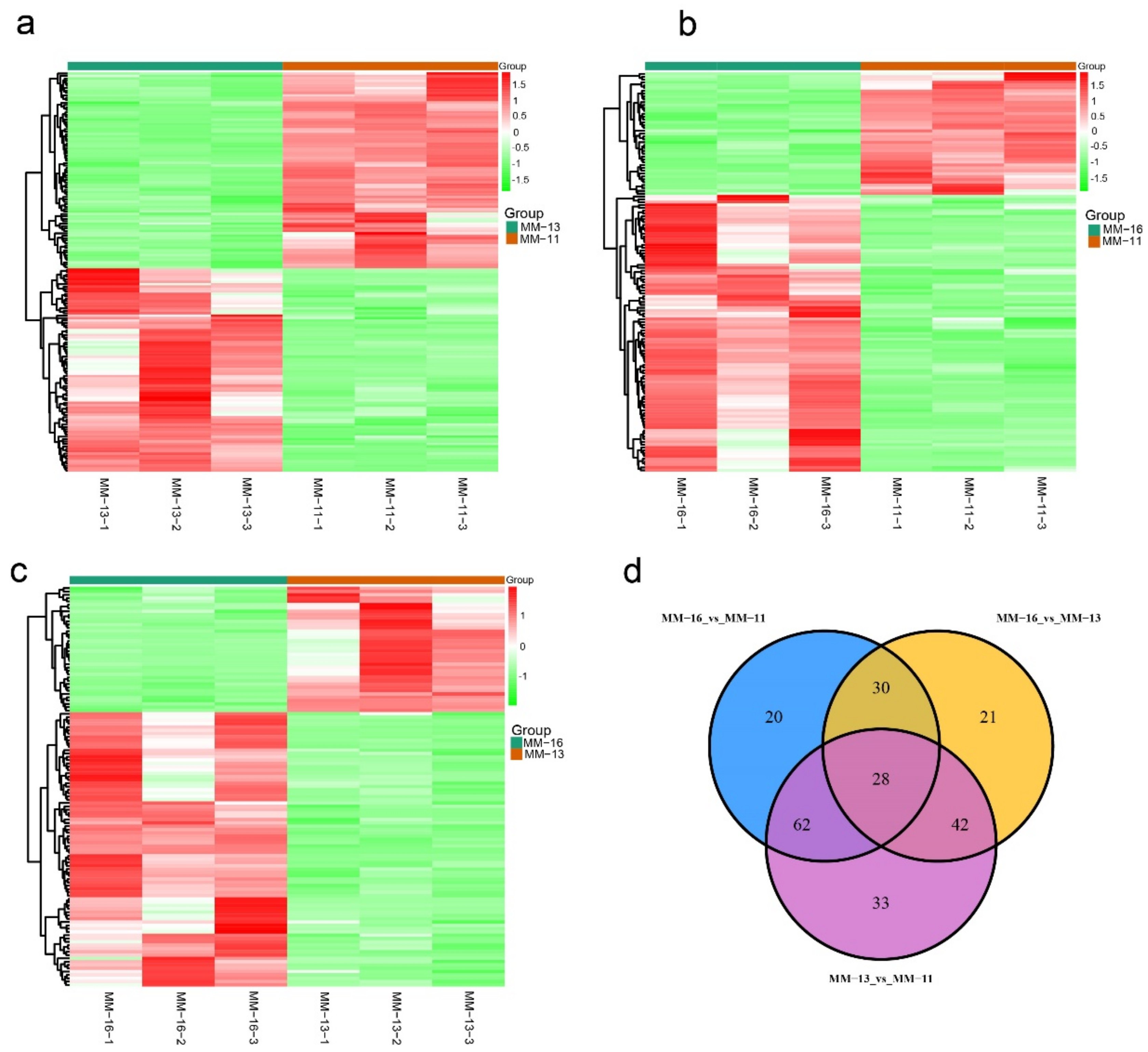
2.2. Dynamic Metabolic Changes of Different Varieties
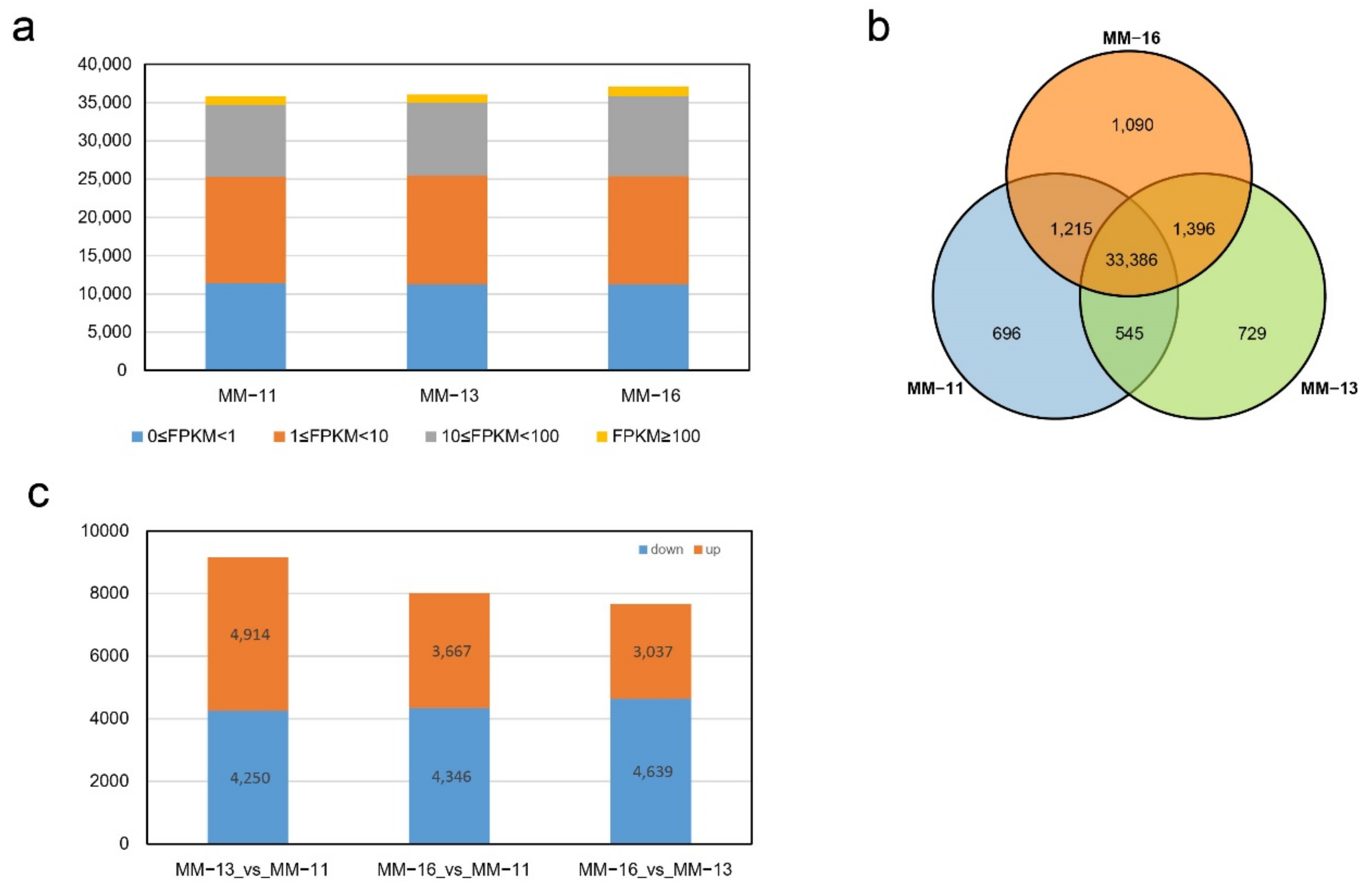
2.3. Transcriptome Analysis
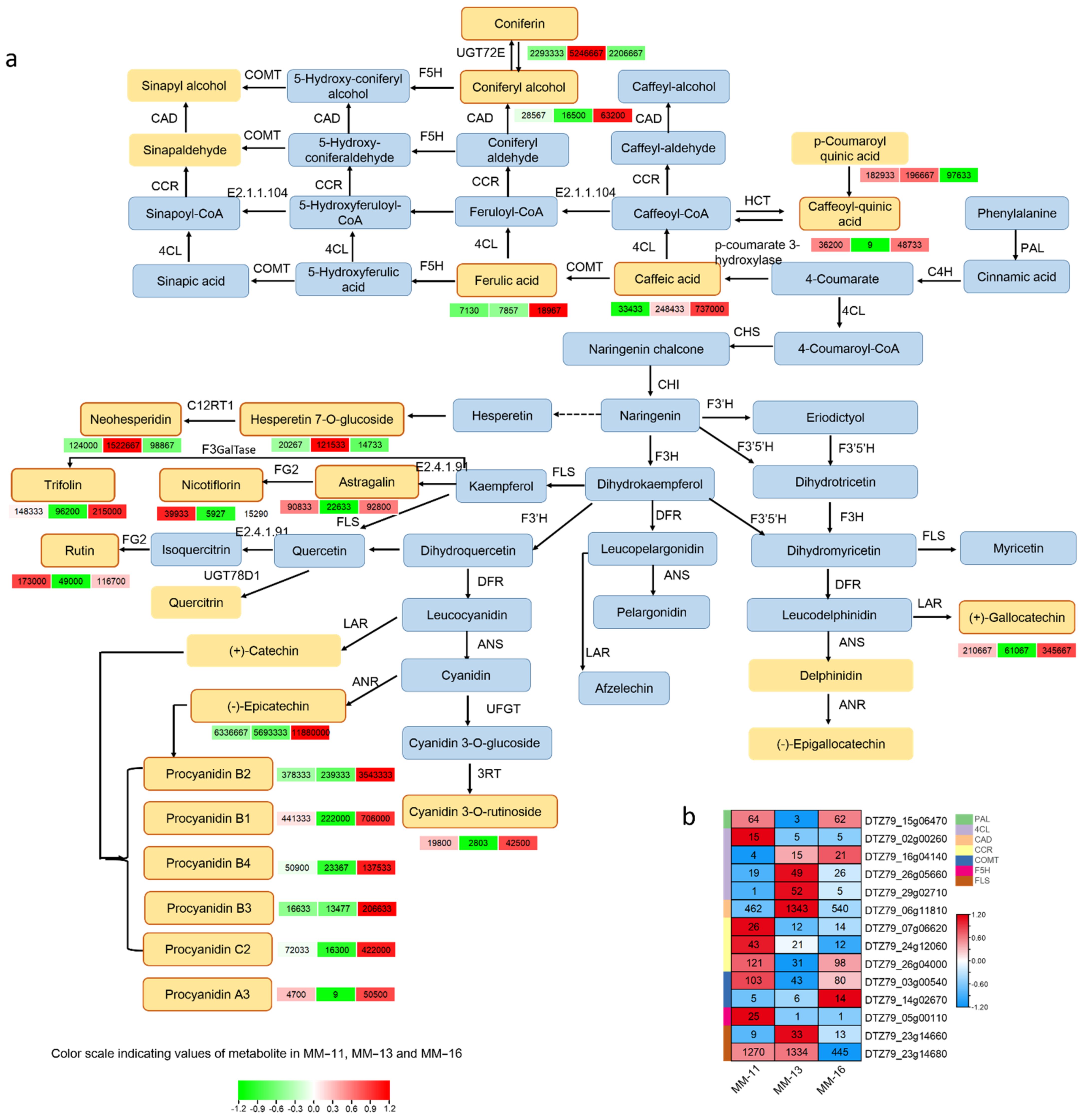
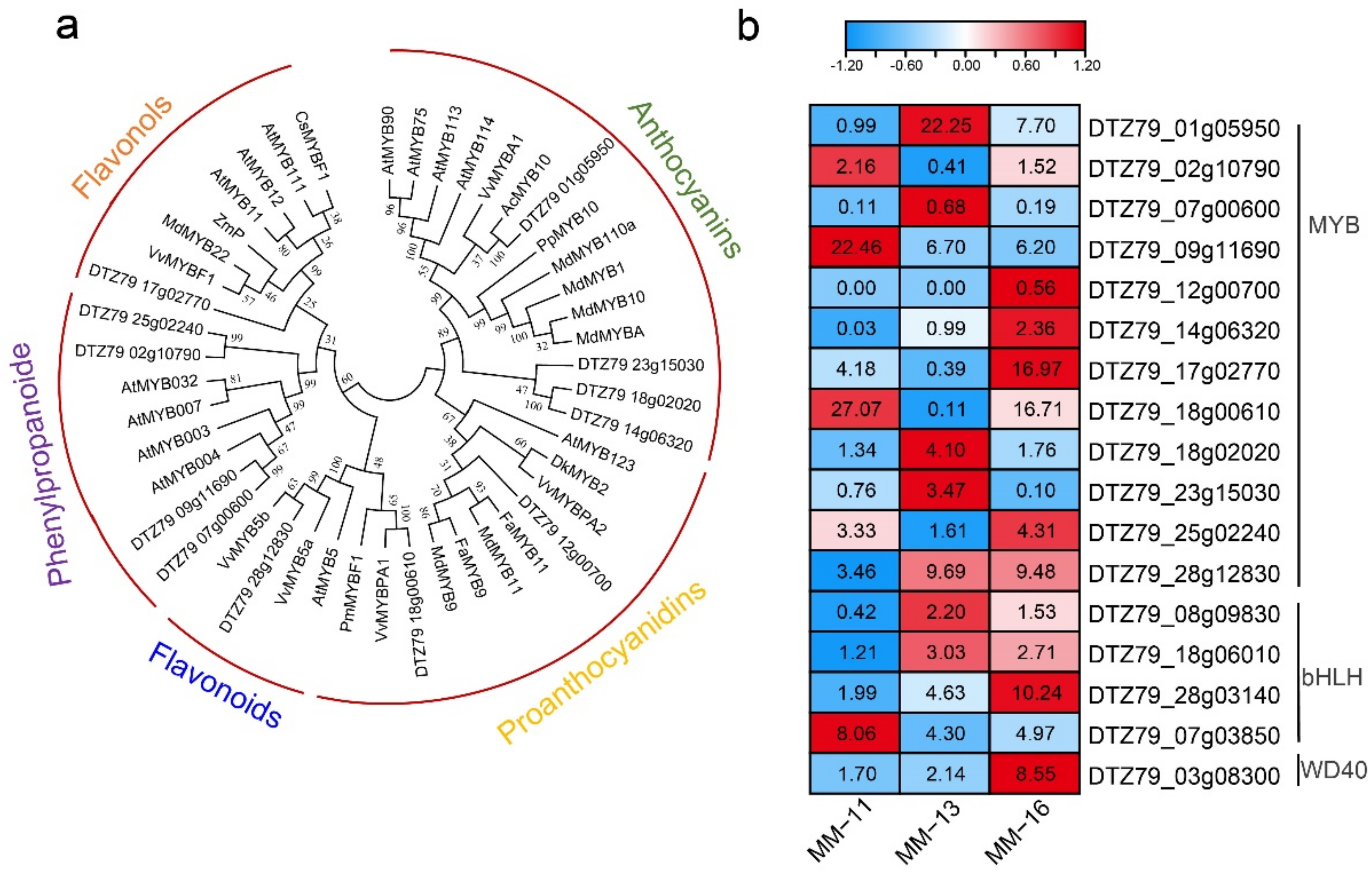
2.4. Identified Metabolites Involved in Phenolic Acid, Flavonoid and Anthocyanin Biosynthesis
2.5. Expression of Phenolic Acid, Flavonoid and Aanthocyanidin Biosynthesis Genes in Three A. eriantha Varieties
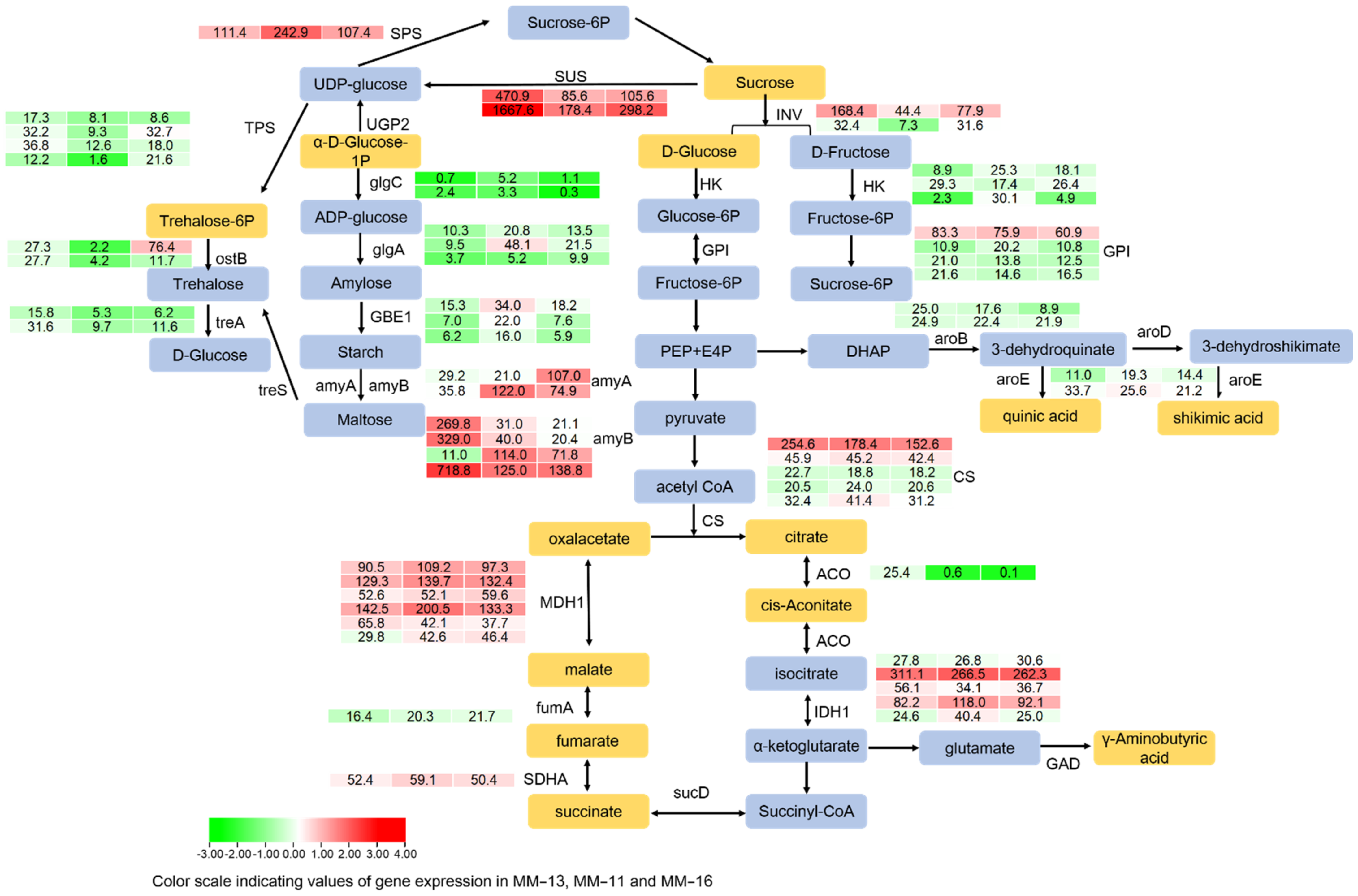
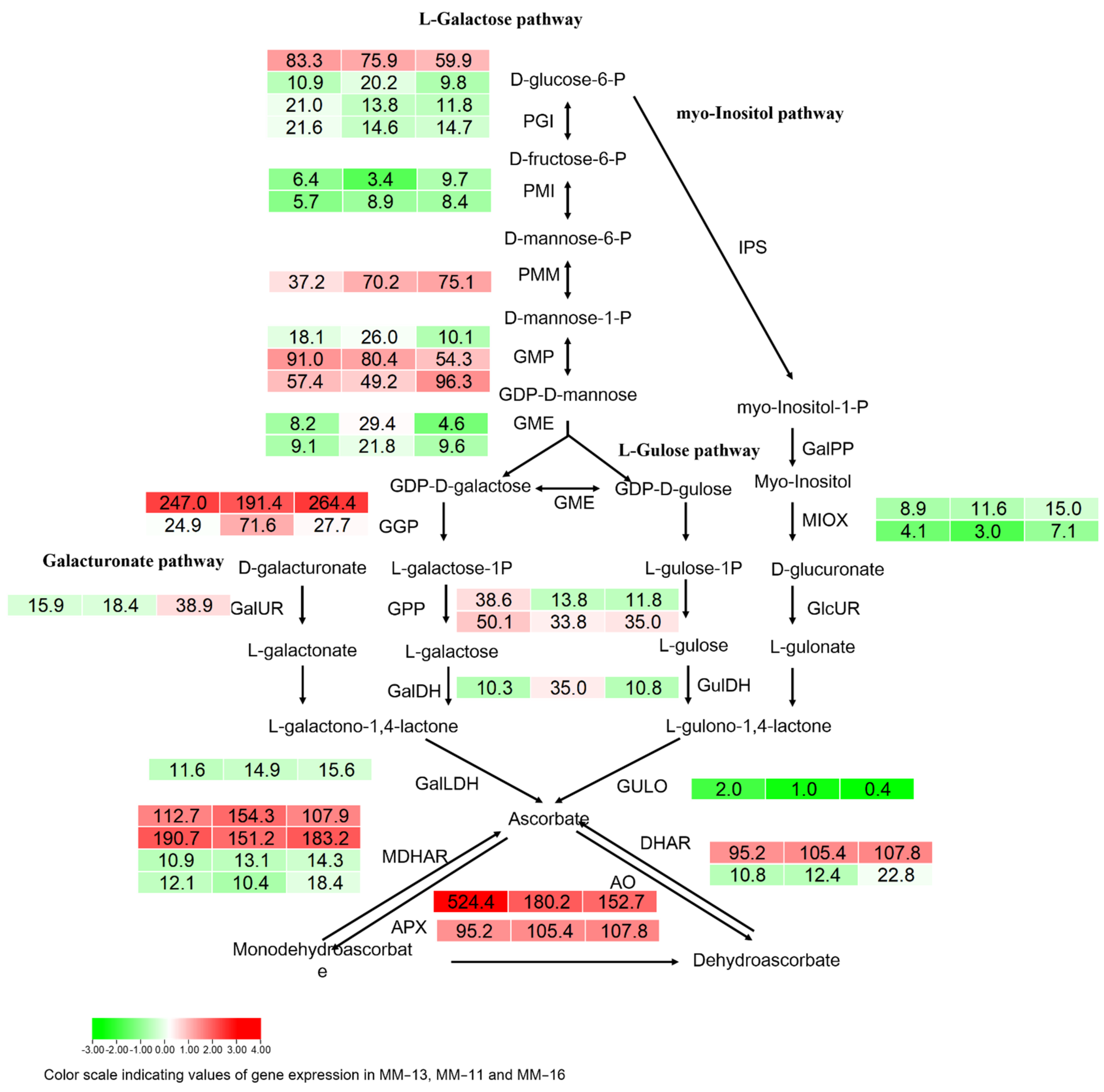
2.6. Soluble Sugar, Organic Acid and AsA in Three A. eriantha Varieties
2.7. Validation of Gene Expression Level in Three Varieties
3. Discussion
4. Materials and Methods
4.1. Fruit Materials
4.2. Metabolome Data Analysis
4.3. Transcriptome Sequencing and Analysis
4.4. Quantitative Real-Time PCR (qRT-PCR) Validation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Binbin, L.; Bo, Z.; Min, X.; Wenbiao, Z.; Seyrek, U.A.; Chunhui, H.; Xiaobiao, X. Variation and probability grading of the main quantitative characteristics of wild Actinidia eriantha germplasm resources. Int. J. Fruit Sci. 2016, 33, 8–15. [Google Scholar] [CrossRef]
- Tang, J.; Wu, H.; Lang, B.; Qu, X.; Huang, C.; Xu, X. Association analysis on leaf and fruit AsA content and SSR markers of wild Actinidia eriantha. Acta Hortic. Sin. 2014, 41, 833–840. [Google Scholar] [CrossRef]
- Zhong, C.; Zhang, P.; Jiang, Z.; Wang, S.; Han, F.; Xu, L.; Huang, H. Dynamic Changes of Carbohydrate and Vitamin C in Fruits of Actinidia chinensis and A. eriantha during Growing Season. Plant Sci. J. 2011, 29, 370–376. [Google Scholar]
- Huang, C.-H.; Tao, J.-J.; Liao, G.-L.; Xie, M.; Qu, X.-Y.; Chen, L.; Xu, X.-B. Dynamic changes of phenol and antioxidant capacity during fruit development of three Actinidia species (kiwifruit). Sci. Hortic. 2020, 273, 109571. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, J.; Chen, F.; Chen, X.; Zhou, Z.; Wang, H. Activation of RAW264.7 macrophages by the polysaccharide from the roots of Actinidia eriantha and its molecular mechanisms. Carbohydr. Polym. 2015, 121, 388–402. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yao, L.; Sun, H.; Wu, Y. Chemical composition and antitumor activity of different polysaccharides from the roots of Actinidia eriantha. Carbohydr. Polym. 2009, 78, 316–322. [Google Scholar] [CrossRef]
- Liao, G.; Li, Z.; Huang, C.; Zhong, M.; Tao, J.; Qu, X.; Chen, L.; Xu, X. Genetic diversity of inner quality and SSR association analysis of wild kiwifruit (Actinidia eriantha). Sci. Hortic. 2019, 248, 241–247. [Google Scholar] [CrossRef]
- Tang, J.; Huang, C.; Wu, H.; Lang, B.; Qu, X.; Xu, X. Genetic Diversity of Wild Actinidia eriantha Germplasm Based on Fruit Traits and SSR Markers. Acta Hortic. Sin. 2014, 41, 1198–1206. [Google Scholar] [CrossRef]
- Mason, A.S. Interploid and Interspecific Hybridization for Kiwifruit Improvement; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Liao, G.; Xu, X.; Huang, C.; Zhong, M.; Jia, D. Resource evaluation and novel germplasm mining of Actinidia eriantha. Sci. Hortic. 2021, 282, 110037. [Google Scholar] [CrossRef]
- Zhong, C.; Wang, S.; Jiang, Z.; Huang, H. ‘Jinyan’, an Interspecific Hybrid Kiwifruit with Brilliant Yellow Flesh and Good Storage Quality. HortSci. Horts 2012, 47, 1187–1190. [Google Scholar] [CrossRef]
- Xu, X.; Liao, G.; Huang, C.; Zhong, M.; Jia, D.; Qu, X.; Liu, Q.; He, Y.; Li, Y. Differences of sucrose accumulation concentration and related genes expression between two sucrose accumulation types of Actinidia eriantha. Sci. Rep. 2020, 10, 20474. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Yan, P.; Du, K.; Li, M.; Xie, Y.; Gao, P. Nutritional component analyses of kiwifruit in different development stages by metabolomic and transcriptomic approaches. J. Sci. Food Agric. 2020, 100, 2399–2409. [Google Scholar] [CrossRef] [PubMed]
- Li, A.N.; Li, S.; Zhang, Y.J.; Xu, X.R.; Chen, Y.M.; Li, H.B. Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef] [PubMed]
- Hano, C.; Tungmunnithum, D. Plant Polyphenols, More Than Just Simple Natural Antioxidants: Oxidative Stress, Aging and Age-Related Diseases. Medicines 2020, 7, 26. [Google Scholar] [CrossRef]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Tapiero, H.; Tew, K.D.; Ba, G.N.; Mathé, G. Polyphenols: Do they play a role in the prevention of human pathologies? Biomed. Pharmacother. Biomed. Pharmacother. 2002, 56, 200–207. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef]
- Shin, H.S.; Satsu, H.; Bae, M.-J.; Zhao, Z.; Ogiwara, H.; Totsuka, M.; Shimizu, M. Anti-inflammatory effect of chlorogenic acid on the IL-8 production in Caco-2 cells and the dextran sulphate sodium-induced colitis symptoms in C57BL/6 mice. Food Chem. 2015, 168, 167–175. [Google Scholar] [CrossRef]
- Grosso, G.; Micek, A.; Godos, J.; Pajak, A.; Sciacca, S.; Galvano, F.; Giovannucci, E.L. Dietary Flavonoid and Lignan Intake and Mortality in Prospective Cohort Studies: Systematic Review and Dose-Response Meta-Analysis. Am. J. Epidemiol. 2017, 185, 1304–1316. [Google Scholar] [CrossRef]
- Bondonno, N.P.; Dalgaard, F.; Kyrø, C.; Murray, K.; Bondonno, C.P.; Lewis, J.R.; Croft, K.D.; Gislason, G.; Scalbert, A.; Cassidy, A.; et al. Flavonoid intake is associated with lower mortality in the Danish Diet Cancer and Health Cohort. Nat. Commun. 2019, 10, 3651. [Google Scholar] [CrossRef]
- Liu, Y.; Qi, Y.; Chen, X.; He, H.; Liu, Z.; Zhang, Z.; Ren, Y.; Ren, X. Phenolic compounds and antioxidant activity in red- and in green-fleshed kiwifruits. Food Res. Int. 2019, 116, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.-R.; Li, H.-Y.; Wei, S.-Y.; Han, Q.-H.; Zhao, L.; Zhang, Q.; Li, S.-Q.; Qin, W.; Wu, D.-T. Changes of phenolic compounds, antioxidant capacities, and inhibitory effects on digestive enzymes of kiwifruits (Actinidia chinensis) during maturation. J. Food Meas. Charact. 2020, 14, 1765–1774. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, R.; Zhang, Y.; Yang, Y.; Sun, X.; Zhang, Q.; Yang, N. Biotransformation of phenolics and metabolites and the change in antioxidant activity in kiwifruit induced by Lactobacillus plantarum fermentation. J. Sci. Food Agric. 2020, 100, 3283–3290. [Google Scholar] [CrossRef] [PubMed]
- Wegrzyn, T.; MacRae, E. Alpha-amylase and Starch Degradation in Kiwifruit. J. Plant Physiol. 1995, 147, 19–28. [Google Scholar] [CrossRef]
- Hu, X.; Kuang, S.; Zhang, A.-D.; Zhang, W.-S.; Chen, M.-J.; Yin, X.-R.; Chen, K.-S. Characterization of Starch Degradation Related Genes in Postharvest Kiwifruit. Int. J. Mol. Sci. 2016, 17, 2112. [Google Scholar] [CrossRef]
- Nielsen, T.H.; Deiting, U.; Stitt, M. A [beta]-Amylase in Potato Tubers Is Induced by Storage at Low Temperature. Plant Physiol. 1997, 113, 503–510. [Google Scholar] [CrossRef]
- Lorberth, R.; Ritte, G.; Willmitzer, L.; Kossmann, J. Inhibition of a starch-granule-bound protein leads to modified starch and repression of cold sweetening. Nat. Biotechnol. 1998, 16, 473–477. [Google Scholar] [CrossRef]
- Chia, T.; Thorneycroft, D.; Chapple, A.; Messerli, G.; Chen, J.; Zeeman, S.C.; Smith, S.M.; Smith, A.M. A cytosolic glucosyltransferase is required for conversion of starch to sucrose in Arabidopsis leaves at night. Plant J. Cell Mol. Biol. 2004, 37, 853–863. [Google Scholar] [CrossRef]
- Yu, T.S.; Kofler, H.; Häusler, R.E.; Hille, D.; Flügge, U.I.; Zeeman, S.C.; Smith, A.M.; Kossmann, J.; Lloyd, J.; Ritte, G.; et al. The Arabidopsis sex1 mutant is defective in the R1 protein, a general regulator of starch degradation in plants, and not in the chloroplast hexose transporter. Plant Cell 2001, 13, 1907–1918. [Google Scholar] [CrossRef]
- Zhang, A.D.; Wang, W.Q.; Tong, Y.; Li, M.J.; Grierson, D.; Ferguson, I.; Chen, K.S.; Yin, X.R. Transcriptome Analysis Identifies a Zinc Finger Protein Regulating Starch Degradation in Kiwifruit. Plant Physiol. 2018, 178, 850–863. [Google Scholar] [CrossRef]
- Nishiyama, I.; Fukuda, T.; Shimohashi, A.; Oota, T. Sugar and Organic Acid Composition in the Fruit Juice of Different Actinidia Varieties. Food Sci. Technol. Res. 2008, 14, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Liu, Y.; Zeng, S.; Xiao, G.; Wang, G.; Wang, Y.; Peng, M.; Huang, H. Gene Expression Profiling of Development and Anthocyanin Accumulation in Kiwifruit (Actinidia chinensis) Based on Transcriptome Sequencing. PLoS ONE 2015, 10, e0136439. [Google Scholar] [CrossRef]
- Jiang, Z.Y.; Zhong, Y.; Zheng, J.; Ali, M.; Liu, G.D.; Zheng, X.L. L-ascorbic acid metabolism in an ascorbate-rich kiwifruit (Actinidia. eriantha Benth.) cv. ‘White’ during postharvest. Plant Physiol. Biochem. PPB 2018, 124, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fang, J.; Qi, X.; Lin, M.; Zhong, Y.; Sun, L.; Cui, W. Combined Analysis of the Fruit Metabolome and Transcriptome Reveals Candidate Genes Involved in Flavonoid Biosynthesis in Actinidia arguta. Int. J. Mol. Sci. 2018, 19, 1471. [Google Scholar] [CrossRef]
- Payyavula, R.S.; Singh, R.K.; Navarre, D.A. Transcription factors, sucrose, and sucrose metabolic genes interact to regulate potato phenylpropanoid metabolism. J. Exp. Bot. 2013, 64, 5115–5131. [Google Scholar] [CrossRef] [PubMed]
- Kulshrestha, S.; Tyagi, P.; Sindhi, V.; Yadavilli, K.S. Invertase and its applications—A brief review. J. Pharm. Res. 2013, 7, 792–797. [Google Scholar] [CrossRef]
- Huang, S.; Ding, J.; Deng, D.; Tang, W.; Sun, H.; Liu, D.; Zhang, L.; Niu, X.; Zhang, X.; Meng, M.; et al. Draft genome of the kiwifruit Actinidia chinensis. Nat. Commun. 2013, 4, 2640. [Google Scholar] [CrossRef]
- Wang, R.; Shu, P.; Zhang, C.; Zhang, J.; Chen, Y.; Zhang, Y.; Du, K.; Xie, Y.; Li, M.; Ma, T.; et al. Integrative analyses of metabolome and genome-wide transcriptome reveal the regulatory network governing flavor formation in kiwifruit (Actinidia chinensis). New Phytol. 2022, 233, 373–389. [Google Scholar] [CrossRef]
- Liao, G.; Liu, Q.; Xu, X.; He, Y.; Li, Y.; Wang, H.; Ye, B.; Huang, C.; Zhong, M.; Jia, D. Metabolome and Transcriptome Reveal Novel Formation Mechanism of Early Mature Trait in Kiwifruit (Actinidia eriantha). Front. Plant Sci. 2021, 12, 760496. [Google Scholar] [CrossRef]
- Yu, M.; Man, Y.; Lei, R.; Lu, X.; Wang, Y. Metabolomics Study of Flavonoids and Anthocyanin-Related Gene Analysis in Kiwifruit (Actinidia chinensis) and Kiwiberry (Actinidia arguta). Plant Mol. Biol. Report 2020, 38, 353–369. [Google Scholar] [CrossRef]
- Gómez-Míguez, M.; González-Manzano, S.; Escribano-Bailón, M.T.; Heredia, F.J.; Santos-Buelga, C. Influence of Different Phenolic Copigments on the Color of Malvidin 3-Glucoside. J. Agric. Food Chem. 2006, 54, 5422–5429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Wang, L.; Liu, Z.; Zhao, Z.; Zhao, J.; Wang, Z.; Zhou, G.; Liu, P.; Liu, M. Transcriptome and metabolome profiling unveil the mechanisms of Ziziphus jujuba Mill. peel coloration. Food Chem. 2020, 312, 125903. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Reichelt, M.; Yoshida, K.; Gershenzon, J.; Constabel, C.P. Two R2R3-MYB proteins are broad repressors of flavonoid and phenylpropanoid metabolism in poplar. Plant J. Cell Mol. Biol. 2018, 96, 949–965. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef]
- Kayesh, E.; Shangguan, L.; Korir, N.K.; Sun, X.; Bilkish, N.; Zhang, Y.; Han, J.; Song, C.; Cheng, Z.-M.; Fang, J. Fruit skin color and the role of anthocyanin. Acta Physiol. Plant 2013, 35, 2879–2890. [Google Scholar] [CrossRef]
- Schijlen, E.G.; Ric de Vos, C.H.; van Tunen, A.J.; Bovy, A.G. Modification of flavonoid biosynthesis in crop plants. Phytochemistry 2004, 65, 2631–2648. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.; Ming, M.; Allan, A.C.; Gu, C.; Li, L.; Wu, X.; Wang, R.; Chang, Y.; Qi, K.; Zhang, S.; et al. Map-based cloning of the pear gene MYB114 identifies an interaction with other transcription factors to coordinately regulate fruit anthocyanin biosynthesis. Plant J. 2017, 92, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Brendolise, C.; Espley, R.V.; Lin-Wang, K.; Laing, W.; Peng, Y.; McGhie, T.; Dejnoprat, S.; Tomes, S.; Hellens, R.P.; Allan, A.C. Multiple Copies of a Simple MYB-Binding Site Confers Trans-regulation by Specific Flavonoid-Related R2R3 MYBs in Diverse Species. Front. Plant Sci. 2017, 8, 1864. [Google Scholar] [CrossRef] [PubMed]
- Montefiori, M.; Brendolise, C.; Dare, A.P.; Lin-Wang, K.; Davies, K.M.; Hellens, R.P.; Allan, A.C. In the Solanaceae, a hierarchy of bHLHs confer distinct target specificity to the anthocyanin regulatory complex. J. Exp. Bot. 2015, 66, 1427–1436. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, K.; Qi, Y.; Lv, G.; Ren, X.; Liu, Z.; Ma, F. Transcriptional Regulation of Anthocyanin Synthesis by MYB-bHLH-WDR Complexes in Kiwifruit (Actinidia chinensis). J. Agric. Food Chem. 2021, 69, 3677–3691. [Google Scholar] [CrossRef]
- Wei, X.; Mao, L.; Wei, X.; Guan, W.; Chen, R.; Luo, Z. AchMYC2 promotes JA-mediated suberin polyphenolic accumulation via the activation of phenylpropanoid metabolism-related genes in the wound healing of kiwifruit (Actinidia chinensis). Postharvest Biol. Technol. 2022, 188, 111896. [Google Scholar] [CrossRef]
- Passeri, V.; Martens, S.; Carvalho, E.; Bianchet, C.; Damiani, F.; Paolocci, F. The R2R3MYB VvMYBPA1 from grape reprograms the phenylpropanoid pathway in tobacco flowers. Planta 2017, 246, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Bogs, J.; Jaffé, F.W.; Takos, A.M.; Walker, A.R.; Robinson, S.P. The grapevine transcription factor VvMYBPA1 regulates proanthocyanidin synthesis during fruit development. Plant Physiol. 2007, 143, 1347–1361. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.C.; Marsh, K.B.; Boldingh, H.L.; Pickering, A.H.; Bulley, S.M.; Frearson, N.J.; Ferguson, A.R.; Thornber, S.E.; Bolitho, K.M.; Macrae, E.A. High growing temperatures reduce fruit carbohydrate and vitamin C in kiwifruit. Plant Cell Environ. 2004, 27, 423–435. [Google Scholar] [CrossRef]
- Lintas, C.; Adorisio, S.; Cappelloni, M.; Monastra, E. Composition and nutritional evaluation of kiwifruit grown in Italy. N. Z. J. Crop Hortic. Sci. 1991, 19, 341–344. [Google Scholar] [CrossRef]
- He, Y.; Li, Z.; Liao, G.; Chen, L.; Zhong, M.; Huang, C.; Jia, D.; Qu, X.; Xu, X. Variation in fruit quality within wild Actinidia eriantha germplasm. N. Z. J. Crop Hortic. Sci. 2020, 48, 153–163. [Google Scholar] [CrossRef]
- Bulley, S.M.; Rassam, M.; Hoser, D.; Otto, W.; Schünemann, N.; Wright, M.; MacRae, E.; Gleave, A.; Laing, W. Gene expression studies in kiwifruit and gene over-expression in Arabidopsis indicates that GDP-L-galactose guanyltransferase is a major control point of vitamin C biosynthesis. J. Exp. Bot. 2009, 60, 765–778. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | MM-13 vs.MM-11) | MM-16 vs. MM-11) | MM-16 vs. MM-13) | |||
|---|---|---|---|---|---|---|
| Up | Down | Up | Down | Up | Down | |
| Flavonoids | 20 | 16 | 4 | 10 | 14 | 19 |
| Lipids | 12 | 7 | 7 | 2 | 6 | 1 |
| Phenolic acids | 9 | 13 | 9 | 18 | 5 | 14 |
| Amino acids and derivatives | 6 | 17 | 7 | 15 | 6 | 5 |
| Terpenoids | 6 | 11 | 5 | 11 | 1 | 6 |
| Nucleotides and derivatives | 6 | 2 | 2 | 6 | 0 | 7 |
| Organic acids | 6 | 3 | 2 | 7 | 1 | 7 |
| Others | 4 | 4 | 1 | 4 | 1 | 3 |
| Tannins | 3 | 0 | 0 | 4 | 0 | 6 |
| Alkaloids | 2 | 6 | 1 | 7 | 0 | 5 |
| Coumarins | 2 | 1 | 0 | 4 | 1 | 4 |
| Saccharides and Alcohols | 2 | 1 | 2 | 1 | 1 | 0 |
| Vitamin | 2 | 2 | 1 | 1 | 0 | 1 |
| Lignans | 1 | 1 | 2 | 7 | 1 | 5 |
| Stilbene | 0 | 0 | 0 | 0 | 1 | 0 |
| Total | 81 | 84 | 43 | 97 | 38 | 83 |
| Compounds | Class | Content | Type | ||
|---|---|---|---|---|---|
| MM-13 | MM-11 | MM-16 | |||
| Coniferin | Phenolic acids | 5,246,667 | 2,293,333 | 2,206,667 | DAM |
| Caffeic acid | Phenolic acids | 248,433 | 33,433 | 737,000 | DAM |
| 3-O-p-Coumaroyl quinic acid | Phenolic acids | 196,667 | 182,933 | 97,633 | DAM |
| Sinapyl alcohol | Phenolic acids | 212,333 | 388,000 | 281,000 | |
| Coniferyl alcohol | Phenolic acids | 16,500 | 28,567 | 63,200 | DAM |
| Sinapinaldehyde | Phenolic acids | 12,120 | 20,200 | 18,267 | |
| Ferulic acid | Phenolic acids | 7857 | 7130 | 18,967 | DAM |
| 4-Caffeoylquinic acid * | Phenolic acids | 9 | 36,200 | 48,733 | DAM |
| Hesperetin 7-O-neohesperidoside (Neohesperidin) * | Dihydroflavone | 1,522,667 | 124,000 | 98,867 | DAM |
| Delphinidin chloride | Anthocyanins | 14,567 | 17,067 | 15,933 | |
| Cyanidin-3-O-rutinoside (Keracyanin) | Anthocyanins | 2803 | 19,800 | 42,500 | DAM |
| Hesperetin-7-O-glucoside | Flavonoid | 121,533 | 20,267 | 14,733 | DAM |
| Kaempferol-3-O-galactoside (Trifolin) | Flavonols | 96,200 | 148,333 | 215,000 | DAM |
| Quercetin-3-O-rutinoside (Rutin) | Flavonols | 49,000 | 173,000 | 116,700 | DAM |
| Quercetin-3-O-α-L-rhamnoside (Quercitrin) | Flavonols | 29,267 | 15,243 | 25,867 | |
| Kaempferol-3-O-glucoside (Astragalin) | Flavonols | 22,633 | 90,833 | 92,800 | DAM |
| Kaempferol-3-O-rutinoside (Nicotiflorin) | Flavonols | 5927 | 39,933 | 15,290 | DAM |
| Epicatechin * | Flavanols | 5,693,333 | 6,336,667 | 11,880,000 | DAM |
| Catechin * | Flavanols | 4,053,333 | 3,630,000 | 6,743,333 | |
| Gallocatechin * | Flavanols | 61,067 | 210,667 | 345,667 | DAM |
| Epigallocatechin * | Flavanols | 42,400 | 45,200 | 51,633 | |
| Procyanidin B1 | Proanthocyanidins | 222,000 | 441,333 | 706,000 | DAM |
| Procyanidin A3 | Proanthocyanidins | 9 | 4700 | 50,500 | DAM |
| Procyanidin B2 | Proanthocyanidins | 239,333 | 378,333 | 3,543,333 | DAM |
| Procyanidin B4 | Proanthocyanidins | 23,367 | 50,900 | 137,533 | DAM |
| Procyanidin B3 | Proanthocyanidins | 13,477 | 16,633 | 206,633 | DAM |
| Procyanidin C2 | Proanthocyanidins | 16,300 | 72,033 | 422,000 | DAM |
| Gene Name | Gene ID | Matoblics | Correlation | p-Value |
|---|---|---|---|---|
| 4CL | DTZ79_29g02710 | Delphinidin chloride | −0.99905815 | 0.027632 |
| 4CL | DTZ79_26g05660 | Quercetin-3-O-rutinoside (Rutin) | −0.99996421 | 0.005386 |
| CAD | DTZ79_06g11810 | Sinapinaldehyde | −0.99866962 | 0.032842 |
| COMT | DTZ79_14g02670 | Ferulic acid | 0.999602413 | 0.017953 |
| FLS | DTZ79_23g14680 | Procyanidin B3 | −0.99957299 | 0.018605 |
| LAR | DTZ79_13g00660 | Epigallocatechin | −0.99999998 | 0.000135 |
| PAL | DTZ79_15g06470 | 4-Caffeoylquinic acid | 0.99921024 | 0.025303 |
| PAL | DTZ79_15g06470 | Kaempferol-3-O-glucoside (Astragalin) | 0.999746251 | 0.014342 |
| ACO | DTZ79_17g07470 | Citric Acid | −0.99901742 | 0.028224 |
| ACO | DTZ79_17g07470 | L-(-)-Malic acid | 0.997522695 | 0.04482 |
| CS | DTZ79_12g12350 | L-(-)-Malic acid | 0.998746142 | 0.031883 |
| CS | DTZ79_17g00890 | Oxaloacetic acid | −0.99920132 | 0.025446 |
| fumA | DTZ79_17g10730 | L-(-)-Malic acid | −0.99975265 | 0.01416 |
| MDH1 | DTZ79_23g14440 | L-(-)-Malic acid | 0.997955145 | 0.040719 |
| GPI | DTZ79_04g02580 | Quinic Acid | −0.99814646 | 0.038767 |
| SUS | DTZ79_23g00730 | Trehalose 6-phosphate | 0.999359474 | 0.022787 |
| TPS | DTZ79_18g05470 | Trehalose 6-phosphate | 0.997125182 | 0.048284 |
| GMP | DTZ79_24g08440 | L-Ascorbic acid | 0.999898684 | 0.009062 |
| MDHAR | DTZ79_27g01630 | L-Ascorbic acid | 0.999853612 | 0.010893 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, H.; Tao, J.; Zhong, W.; Jiao, X.; Chen, S.; Wu, M.; Gao, Z.; Huang, C. Nutritional Component Analyses in Different Varieties of Actinidia eriantha Kiwifruit by Transcriptomic and Metabolomic Approaches. Int. J. Mol. Sci. 2022, 23, 10217. https://doi.org/10.3390/ijms231810217
Jia H, Tao J, Zhong W, Jiao X, Chen S, Wu M, Gao Z, Huang C. Nutritional Component Analyses in Different Varieties of Actinidia eriantha Kiwifruit by Transcriptomic and Metabolomic Approaches. International Journal of Molecular Sciences. 2022; 23(18):10217. https://doi.org/10.3390/ijms231810217
Chicago/Turabian StyleJia, Huimin, Junjie Tao, Wenqi Zhong, Xudong Jiao, Shuangshuang Chen, Mengting Wu, Zhongshan Gao, and Chunhui Huang. 2022. "Nutritional Component Analyses in Different Varieties of Actinidia eriantha Kiwifruit by Transcriptomic and Metabolomic Approaches" International Journal of Molecular Sciences 23, no. 18: 10217. https://doi.org/10.3390/ijms231810217
APA StyleJia, H., Tao, J., Zhong, W., Jiao, X., Chen, S., Wu, M., Gao, Z., & Huang, C. (2022). Nutritional Component Analyses in Different Varieties of Actinidia eriantha Kiwifruit by Transcriptomic and Metabolomic Approaches. International Journal of Molecular Sciences, 23(18), 10217. https://doi.org/10.3390/ijms231810217