
Sugarcane Rhizosphere Bacteria Community Migration Correlates with Growth Stages and Soil Nutrient

 , , and
, , and
Abstract
:1. Introduction
2. Results
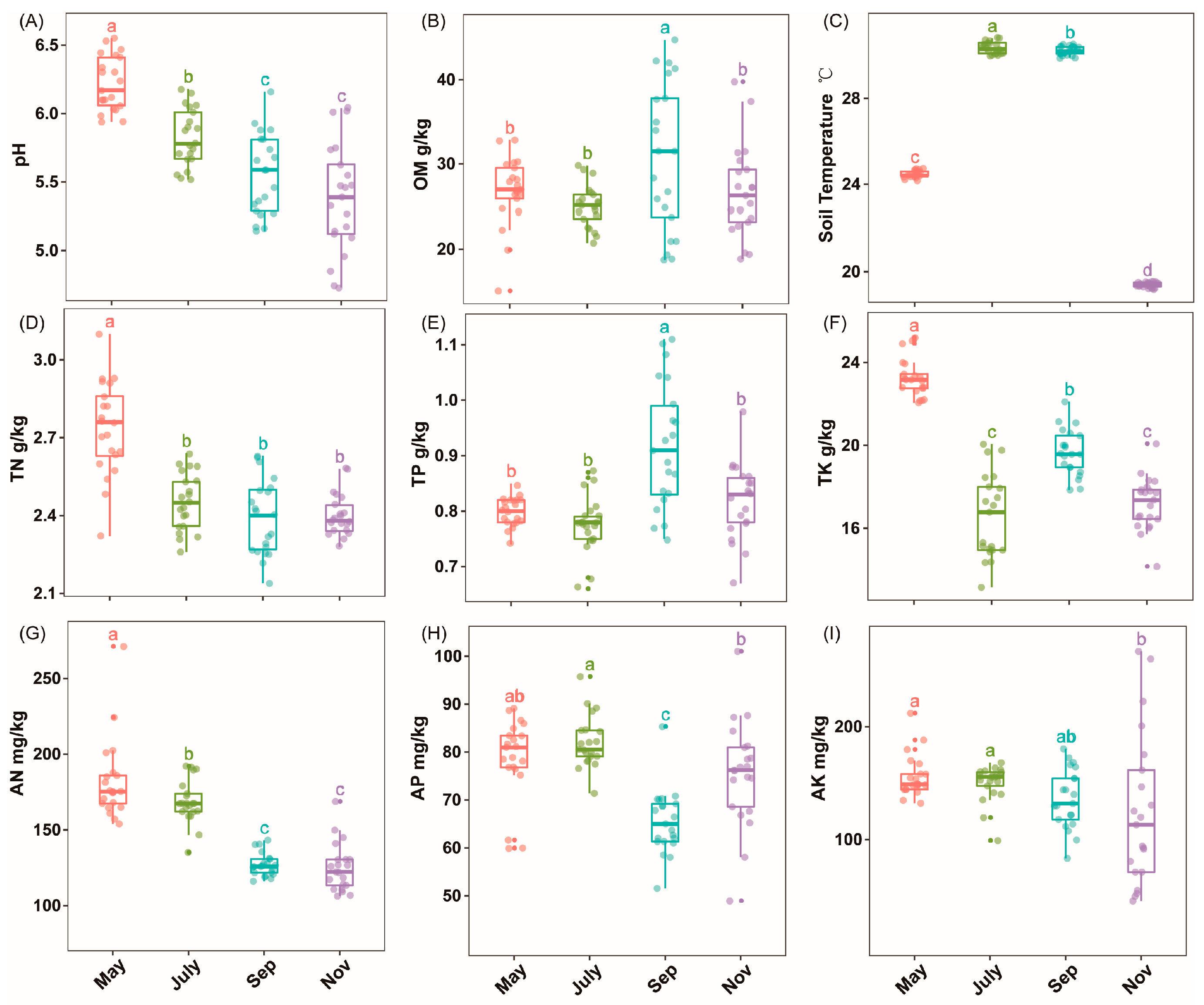
2.1. Soil Physiochemical Properties
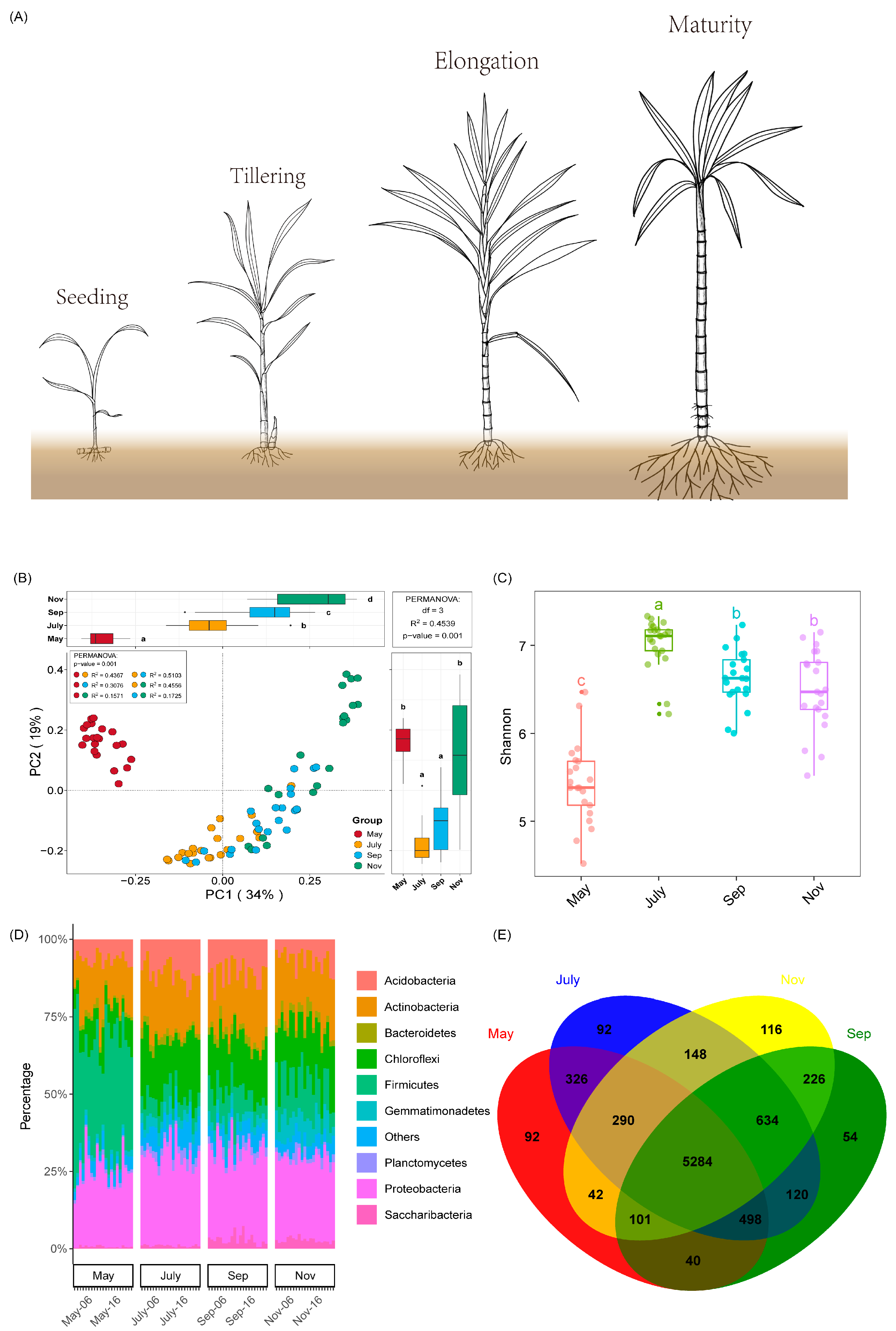
2.2. Rhizosphere Bacteria Community Structure, Diversity and Richness
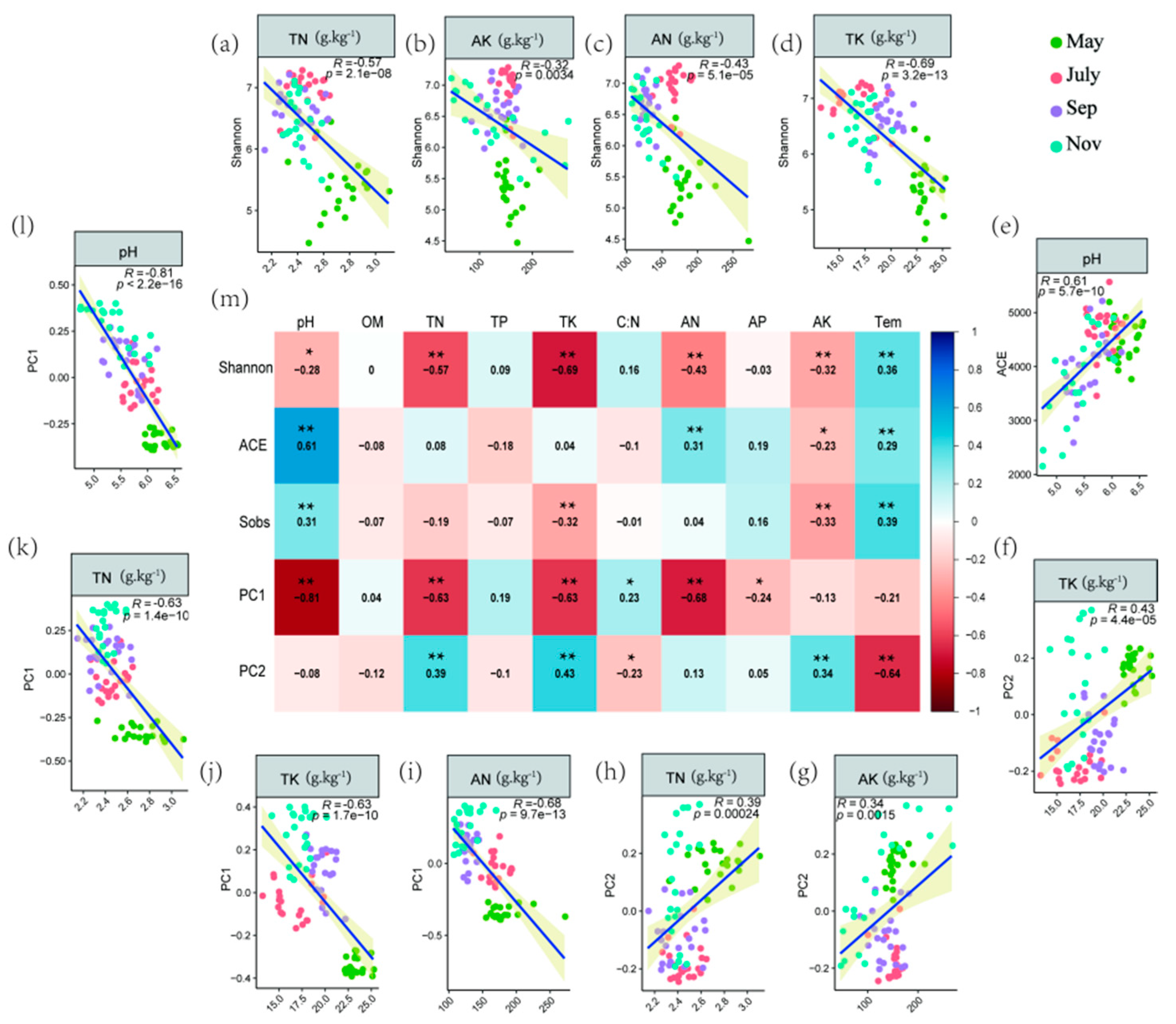
2.3. Correlations of Soil Properties with Bacterial Communities
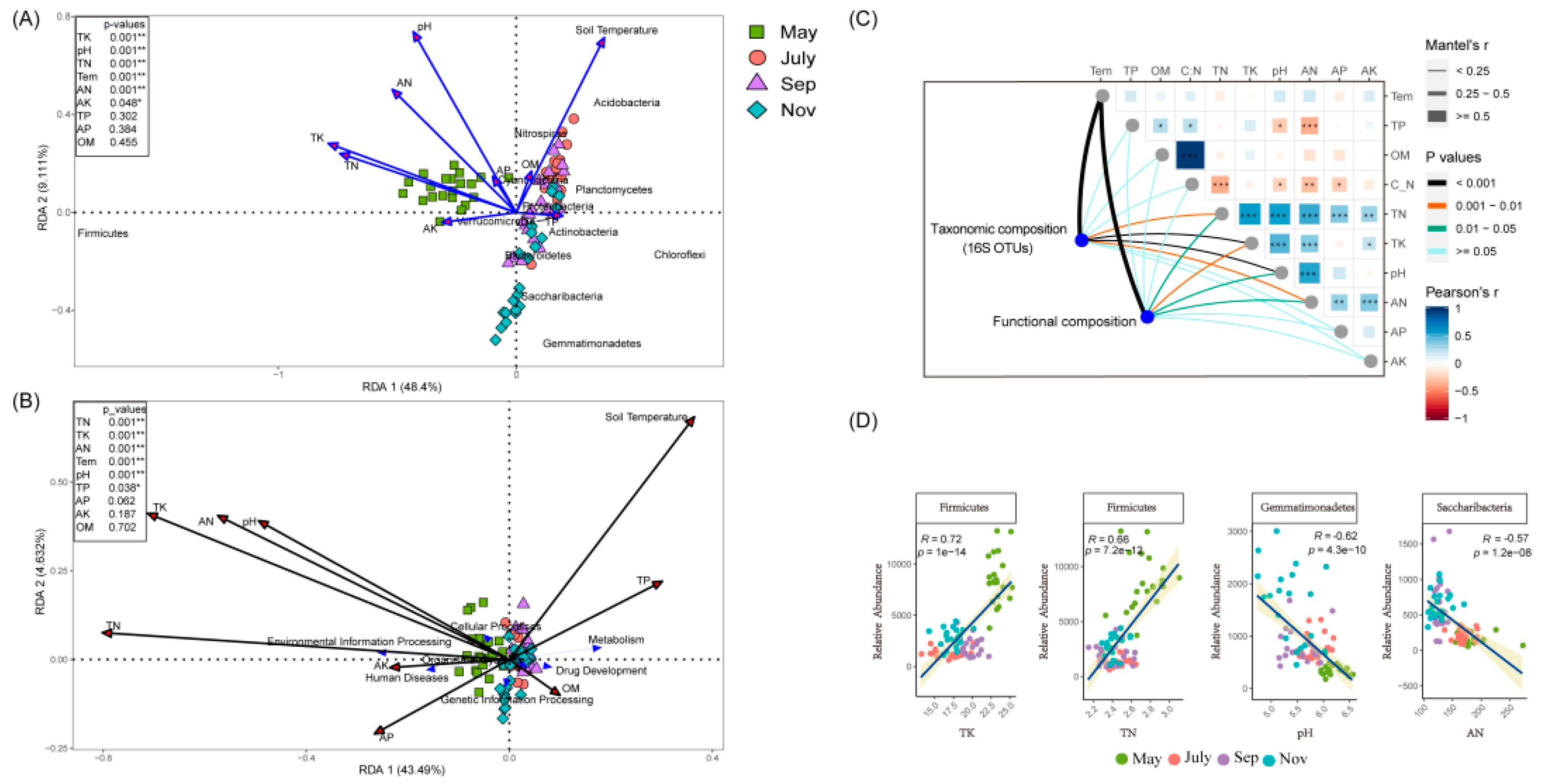
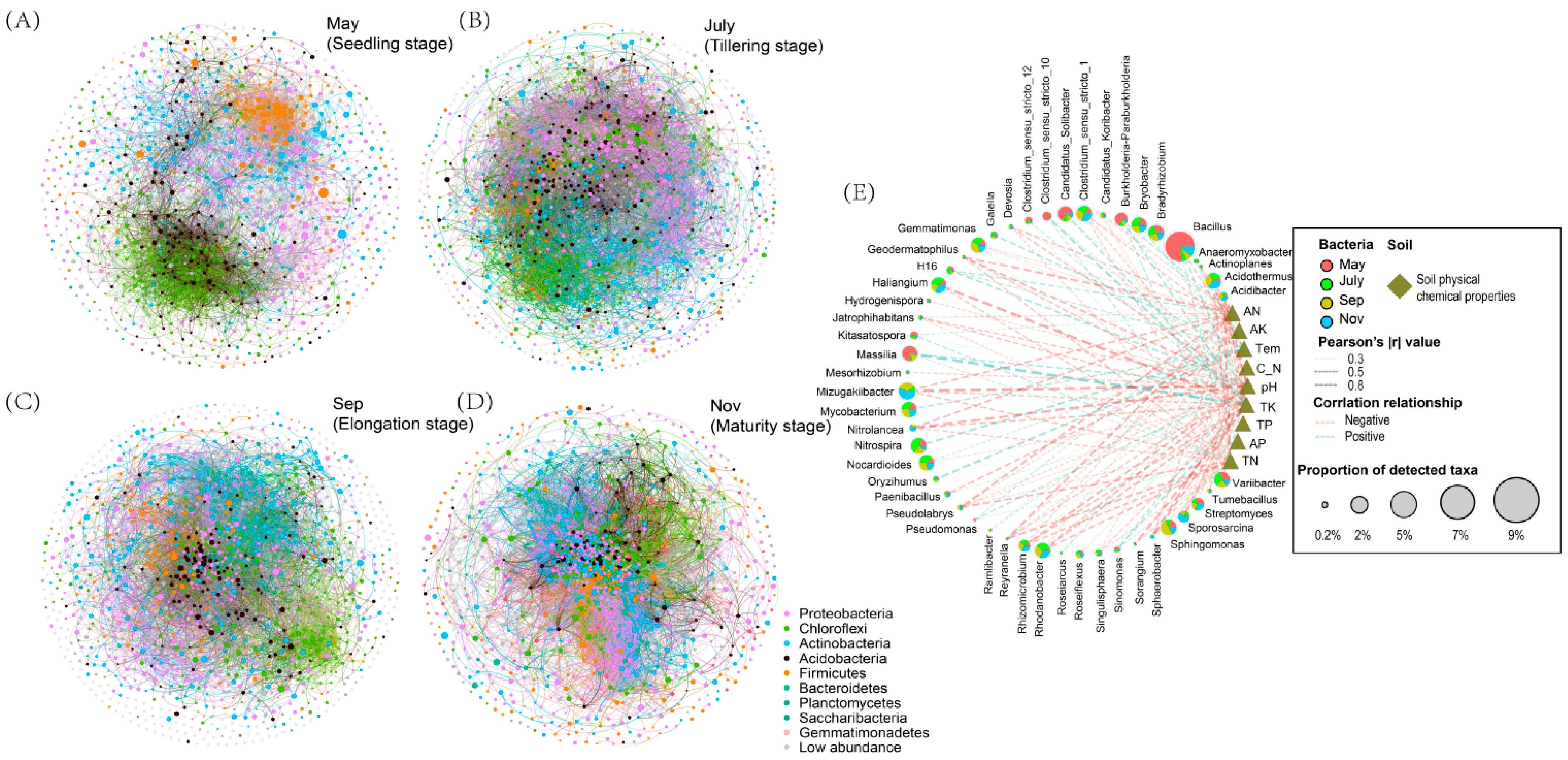
2.4. Soil Environmental Variables and Rhizosphere Bacteria Co-Occurrence Network
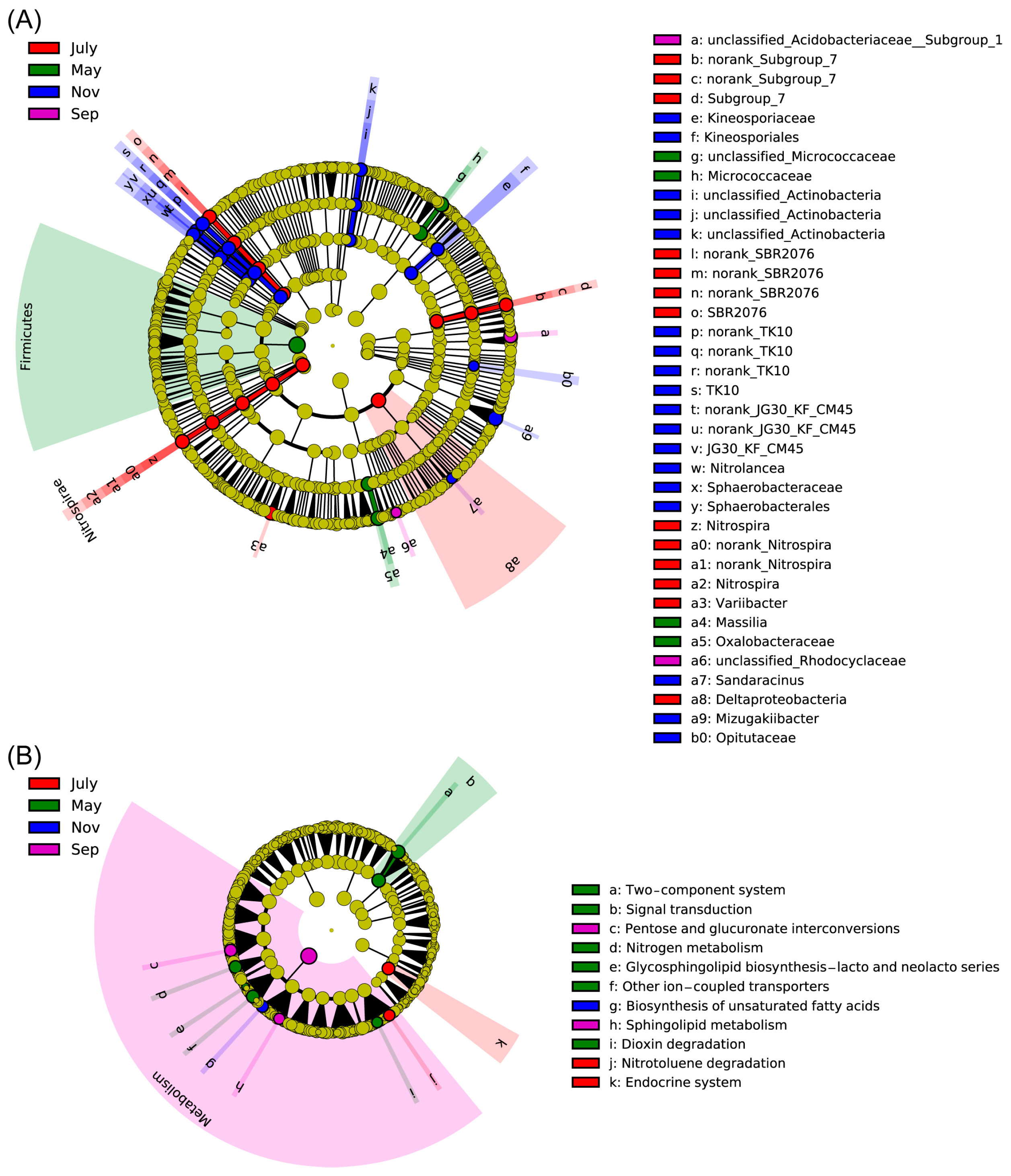
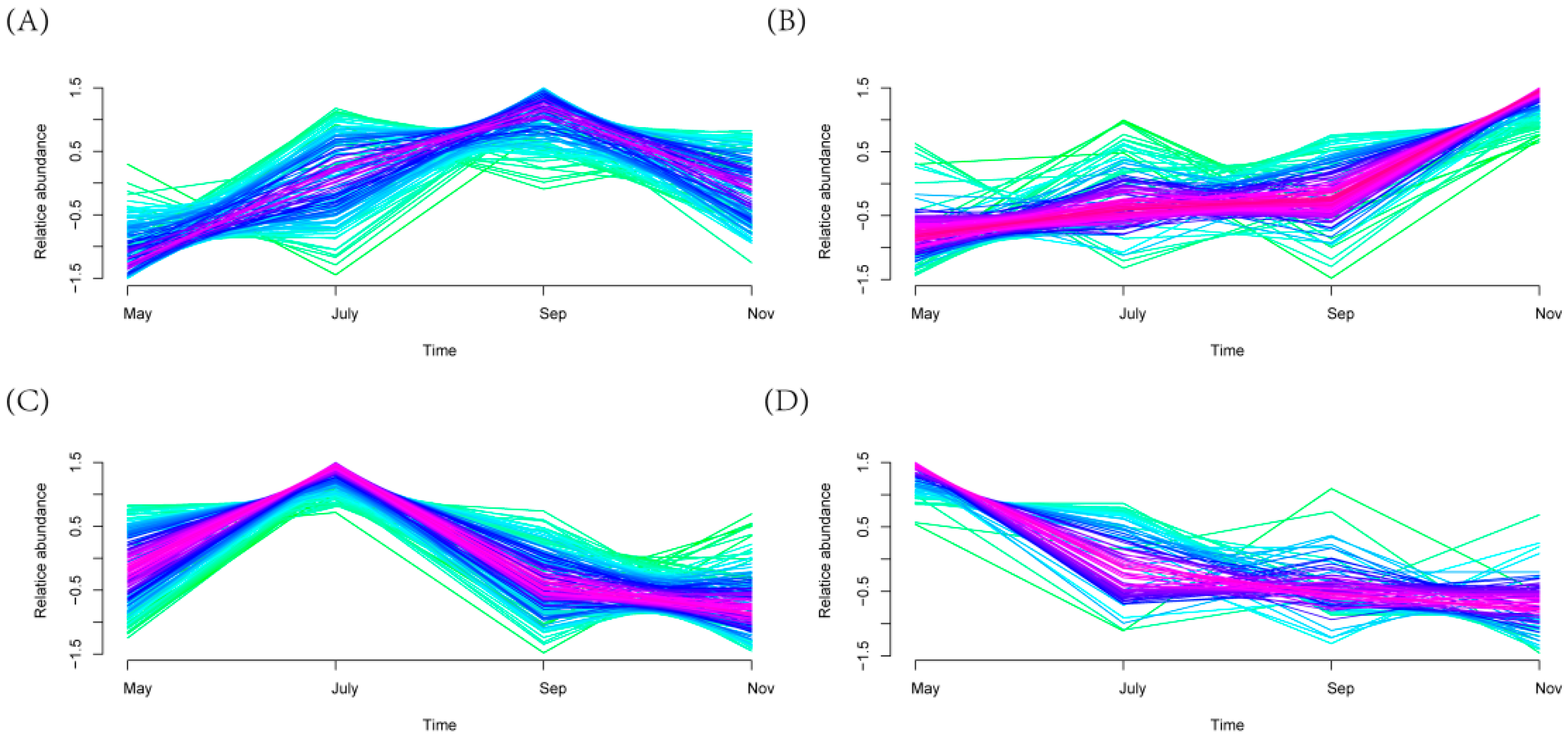
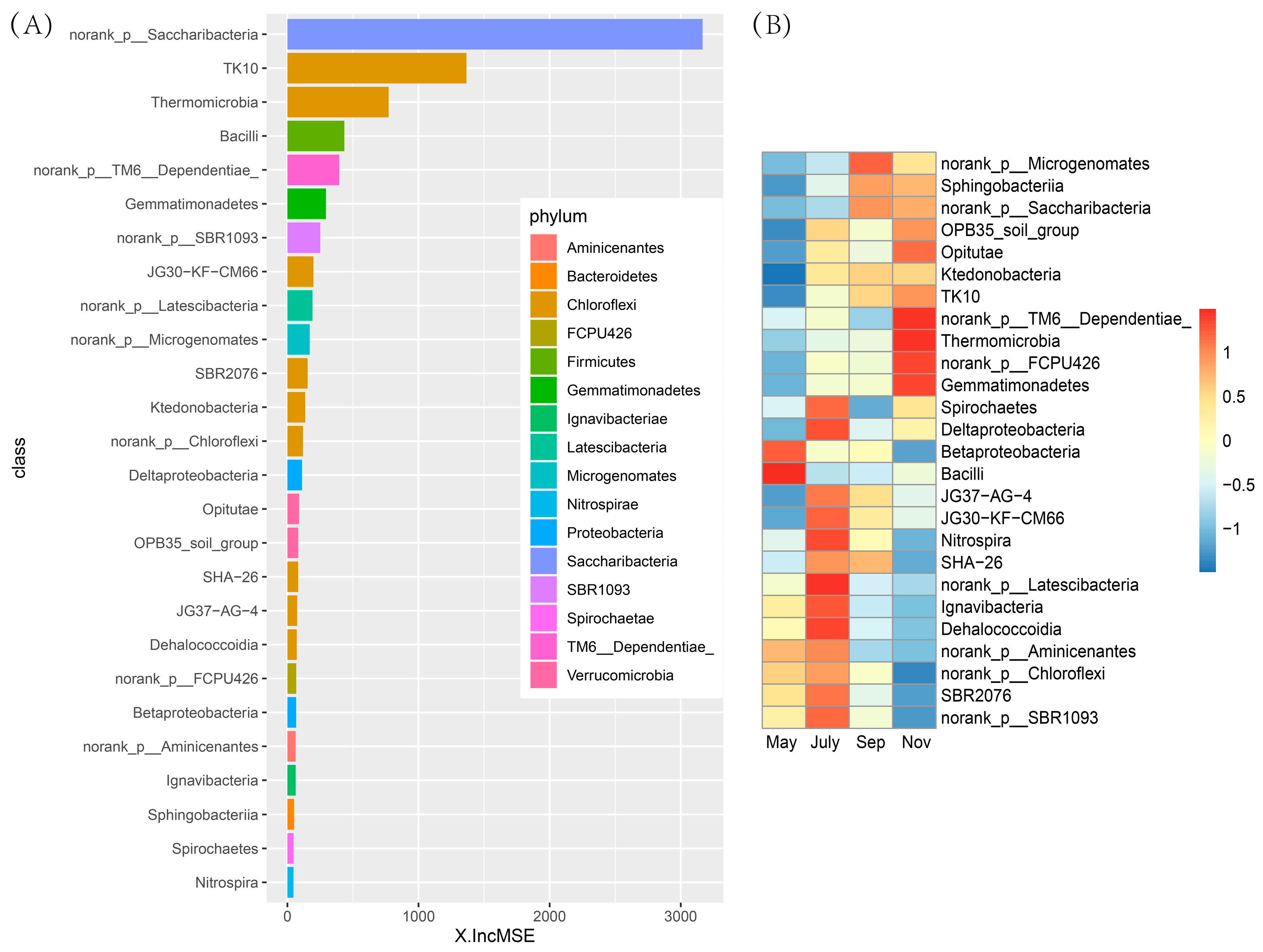
2.5. Differential Rhizosphere Bacteria Abundance Detected during Various Seasons
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Soil Physiochemical Analysis
4.2. Extraction and Electrophoretic Detection of Soil DNA
4.3. 16S rRNA PCR Amplification and Sequencing
4.4. Data Quality Control and Filtering
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manners, J.M.; Casu, R.E. Transcriptome Analysis and Functional Genomics of Sugarcane. Trop. Plant Biol. 2011, 4, 9–21. [Google Scholar] [CrossRef]
- Linnenluecke, M.K.; Zhou, C.; Smith, T.; Thompson, N.; Nucifora, N. The impact of climate change on the Australian sugarcane industry. J. Clean. Prod. 2020, 246, 118974. [Google Scholar] [CrossRef]
- Lopes, C.M.; Silva, A.M.M.; Estrada-Bonilla, G.A.; Ferraz-Almeida, R.; Vieira, J.L.V.; Otto, R.; Vitti, G.C.; Cardoso, E.J.B.N. Improving the fertilizer value of sugarcane wastes through phosphate rock amendment and phosphate-solubilizing bacteria inoculation. J. Clean. Prod. 2021, 298, 126821. [Google Scholar] [CrossRef]
- Damaj, M.B.; Jifon, J.L.; Woodard, S.L.; Vargas-Bautista, C.; Barros, G.O.F.; Molina, J.; White, S.G.; Damaj, B.B.; Nikolov, Z.L.; Mandadi, K.K. Unprecedented enhancement of recombinant protein production in sugarcane culms using a combinatorial promoter stacking system. Sci. Rep. UK 2020, 10, 13713. [Google Scholar] [CrossRef] [PubMed]
- Naseri, H.; Parashkoohi, M.G.; Ranjbar, I.; Zamani, D.M. Sustainability of quantitative and qualitative indicators of sugarcane production under different tillage systems (case study: Khuzestan province of Iran). Environ. Sustain. Indic. 2020, 8, 100046. [Google Scholar] [CrossRef]
- Shukla, S.K.; Sharma, L.; Jaiswal, V.P.; Pathak, A.D.; Tiwari, R.; Awasthi, S.K.; Gaur, A. Soil quality parameters vis-a-vis growth and yield attributes of sugarcane as influenced by integration of microbial consortium with NPK fertilizers. Sci. Rep. UK 2020, 10, 19180. [Google Scholar] [CrossRef] [PubMed]
- Daniel, D.R.; Neung-Hwan, O.; Ryan, F.; Jason, J. The Rhizosphere and Soil Formation. In The Rhizosphere; Academic Press: Cambridge, MA, USA, 2007; pp. 179–182. [Google Scholar]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and heritability of the maize rhizosphere microbiome under field conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef]
- Yang, C.; Crowley, D.E. Rhizosphere Microbial Community Structure in Relation to Root Location and Plant Iron Nutritional Status. Appl. Environ. Microb. 2000, 66, 345–351. [Google Scholar] [CrossRef]
- Singh, B.K.; Millard, P.; Whiteley, A.S.; Murrell, J.C. Unravelling rhizosphere–microbial interactions: Opportunities and limitations. Trends Microbiol. 2004, 12, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Westover, K.M.; Kennedy, A.C.; Kelley, S.E. Patterns of Rhizosphere Microbial Community Structure Associated with Co-Occurring Plant Species. J. Ecol. 1997, 85, 863–873. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Q.; Wang, Z.; Zheng, H.; Chen, Y.; Chen, X.; Wang, L.; Li, H.; Zhang, J. Nitrogen Addition Alleviates Microbial Nitrogen Limitations and Promotes Soil Respiration in a Subalpine Coniferous Forest. Forests 2019, 10, 1038. [Google Scholar] [CrossRef]
- Veen, J.A.V.I.; Kuikman, P.J. Soil structural aspects of decomposition of organic matter by micro-organisms. Biogeochemistry 1990, 11, 213–233. [Google Scholar] [CrossRef]
- Kuske, C.R.; Ticknor, L.O.; Miller, M.E.; Dunbar, J.M.; Davis, J.A.; Barns, S.M.; Belnap, J. Comparison of Soil Bacterial Communities in Rhizospheres of Three Plant Species and the Interspaces in an Arid Grassland. Appl. Environ. Microb. 2002, 68, 1854–1863. [Google Scholar] [CrossRef] [PubMed]
- Aira, M.; Gómez-Brandón, M.; Lazcano, C.; Bååth, E.; Domínguez, J. Plant genotype strongly modifies the structure and growth of maize rhizosphere microbial communities. Soil Biol. Biochem. 2010, 42, 2276–2281. [Google Scholar] [CrossRef]
- Gqozo, M.P.; Bill, M.; Siyoum, N.; Labuschagne, N.; Korsten, L. Fungal diversity and community composition of wheat rhizosphere and non-rhizosphere soils from three different agricultural production regions of South Africa. Appl. Soil Ecol. 2020, 151, 103543. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, N.; Liu, Y.; Zhang, X.; Hu, B.; Qin, Y.; Xu, H.; Wang, H.; Guo, X.; Qian, J.; et al. Root microbiota shift in rice correlates with resident time in the field and developmental stage. Sci. China Life Sci. 2018, 61, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Dombrowski, N.; Schlaeppi, K.; Agler, M.T.; Hacquard, S.; Kemen, E.; Garrido-Oter, R.; Wunder, J.; Coupland, G.; Schulze-Lefert, P. Root microbiota dynamics of perennial Arabis alpina are dependent on soil residence time but independent of flowering time. ISME J. 2017, 11, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Kraffczyk, I.; Trolldenier, G.; Beringer, H. Soluble root exudates of maize: Influence of potassium supply and rhizosphere microorganisms. Soil Biol. Biochem. 1984, 16, 315–322. [Google Scholar] [CrossRef]
- Holden, P.A.U.O.; Firestone, M.K. Soil microorganisms in soil cleanup: How can we improve our understanding. J. Environ. Qual. 1997, 26, 32–40. [Google Scholar] [CrossRef]
- Yu, Y.L.; Chen, Y.X.; Luo, Y.M.; Pan, X.D.; He, Y.F.; Wong, M.H. Rapid degradation of butachlor in wheat rhizosphere soil. Chemosphere 2003, 50, 771–774. [Google Scholar] [CrossRef]
- Kertesz, M.A. The role of soil microbes in plant sulphur nutrition. J. Exp. Bot. 2004, 55, 1939–1945. [Google Scholar] [CrossRef]
- Smith, M.E.; Facelli, J.M.; Cavagnaro, T.R. Interactions between soil properties, soil microbes and plants in remnant-grassland and old-field areas: A reciprocal transplant approach. Plant Soil 2018, 433, 127–145. [Google Scholar] [CrossRef]
- Hill, P.W.; Marsden, K.A.; Jones, D.L. How significant to plant N nutrition is the direct consumption of soil microbes by roots? New Phytol. 2013, 199, 948–955. [Google Scholar] [CrossRef]
- Khan, A.G. Role of soil microbes in the rhizospheres of plants growing on trace metal contaminated soils in phytoremediation. J. Trace Elem. Med. Biol. 2005, 18, 355–364. [Google Scholar] [CrossRef]
- Perry, G.; Perry, D. Ethylene induced nitrile and VOC synthesis by soil microbes; Improved root elongation & reduced risk of fungal infection in plants. PeerJ. Prepr. 2019, 7, e543v2. [Google Scholar] [CrossRef]
- Bitas, V.; Kim, H.; Bennett, J.W.; Kang, S. Sniffing on Microbes: Diverse Roles of Microbial Volatile Organic Compounds in Plant Health. Mol. Plant Microbe Interact. 2013, 26, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Ronald, P.C.; Shirasu, K. Front-runners in plant–microbe interactions. Plant Biol. 2012, 15, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Jones, D.L.; Nguyen, C.; Finlay, R.D. Carbon flow in the rhizosphere: Carbon trading at the soil–root interface. Plant Soil 2009, 321, 5–33. [Google Scholar] [CrossRef]
- Marasco, R.; Rolli, E.; Ettoumi, B.; Vigani, G.; Mapelli, F.; Borin, S.; Abou-Hadid, A.F.; El-Behairy, U.A.; Sorlini, C.; Cherif, A.; et al. A drought resistance-promoting microbiome is selected by root system under desert farming. PLoS ONE 2012, 7, e48479. [Google Scholar] [CrossRef] [PubMed]
- Xiao, E.; Ning, Z.; Xiao, T.; Sun, W.; Qiu, Y. Variation in rhizosphere microbiota correlates with edaphic factor in an abandoned antimony tailing dump. Environ. Pollut. 2019, 253, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Lazzari, A.; Prigione, V.; Voyron, S.; Spadaro, D.; Varese, G.C. Influence of plant genotype on the cultivable fungi associated to tomato rhizosphere and roots in different soils. Fungal. Biol. UK 2016, 120, 862–872. [Google Scholar] [CrossRef]
- Yuan, J.; Wen, T.; Zhang, H.; Zhao, M.; Penton, C.R.; Thomashow, L.S.; Shen, Q. Predicting disease occurrence with high accuracy based on soil macroecological patterns of Fusarium wilt. ISME J. 2020, 14, 2936–2950. [Google Scholar] [CrossRef] [PubMed]
- Ziqin, P.; Jinwen, H.; Nyumah, F.; Wenxiong, L.; Zhaonian, Y.; Chaohua, H. Combining N fertilization with biochar affects root-shoot growth, rhizosphere soil properties and bacterial communities under sugarcane monocropping. Ind. Crops Prod. 2022, 182, 114899. [Google Scholar] [CrossRef]
- Dakora, F.D.; Phillips, D.A. Root exudates as mediators of mineral acquisition in low-nutrient environments. Plant Soil 2002, 245, 201–213. [Google Scholar] [CrossRef]
- Lorenz, K.; And, K.; Kandeler, E. The response of soil microbial biomass and activity of a Norway spruce forest to liming and drought. J. Plant Nutr. Soil Sci. 2001, 164, 9–19. [Google Scholar] [CrossRef]
- Liu, X.; Li, Z.Y.; Liu, R.R.; Li, L.L.; Wang, W.W. Changes of bacterial flora structure in rhizosphere soil of soybean at different growth stages. Guangxi Plant 2018, 38, 107–114. [Google Scholar]
- Lin, M.; Xiong, H.; Xiang, X.; Zhou, Z.; Liang, L.; Mei, Z. The Effect of Plant Geographical Location and Developmental Stage on Root-Associated Microbiomes of Gymnadenia conopsea. Front. Microbiol. 2020, 11, 1257. [Google Scholar] [CrossRef] [PubMed]
- Chaparro, J.M.; Badri, D.V.; Bakker, M.G.; Sugiyama, A.; Manter, D.K.; Vivanco, J.M. Root Exudation of Phytochemicals in Arabidopsis Follows Specific Patterns That Are Developmentally Programmed and Correlate with Soil Microbial Functions. PLoS ONE 2013, 8, e55731. [Google Scholar] [CrossRef]
- Hamman, S.T.; Burke, I.C.; Stromberger, M.E. Relationships between microbial community structure and soil environmental conditions in a recently burned system. Soil Biol. Biochem. 2007, 39, 1703–1711. [Google Scholar] [CrossRef]
- Tayyab, M.; Yang, Z.; Zhang, C.; Islam, W.; Zhang, H. Sugarcane monoculture drives microbial community composition, activity and abundance of agricultural-related microorganisms. Environ. Sci. Pollut. Res. 2021, 28, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, N.; Liu, H.; Zhang, Y.; Yu, L. Soil pH is a Key Determinant of Soil Fungal Community Composition in the Ny-Ålesund Region, Svalbard (High Arctic). Front. Microbiol. 2016, 7, 227. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [PubMed]
- Chaparro, J.; Badri, D.; Vivanco, J. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef]
- Shi, S.; Nuccio, E.; Herman, D.J.; Rijkers, R.; Firestone, M. Successional Trajectories of Rhizosphere Bacterial Communities over Consecutive Seasons. mBio 2015, 6, 500–503. [Google Scholar] [CrossRef]
- Faust, K.; Raes, J. Microbial interactions: From networks to models. Nat. Rev. Microbiol. 2012, 10, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Ofek, M.; Hadar, Y.; Minz, D. Ecology of Root Colonizing Massilia (Oxalobacteraceae). PLoS ONE 2012, 7, e40117. [Google Scholar] [CrossRef]
- Hejdová, B. Ekologie Aktinobakteriálních čeledí Micrococcaceae, Streptomycetaceae a Nocardiaceae v terestrických ekosystémech. České Budějovice, Bachelor’s Thesis. University of South Bohemia in České Budějovice, Faculty of Science. Thesis Supervisor Ing. Jiří Bárta, Ph.D. 2014. Available online: https://theses.cz/id/rxhrtv (accessed on 4 February 2022).
- Hu, J.; Zhao, Y.; Yao, X.; Wang, J.; Hu, B. Dominance of Comammox Nitrospira in Soil Nitrification. Sci. Total Environ. 2021, 780, 146558. [Google Scholar] [CrossRef]
- Lueders, T.; Pommerenke, B.; Friedrich, M.W. Stable-Isotope Probing of Microorganisms Thriving at Thermodynamic Limits: Syntrophic Propionate Oxidation in Flooded Soil. Appl. Environ. Microb. 2004, 70, 5778–5786. [Google Scholar] [CrossRef]
- Yousuf, B.; Kumar, R.; Mishra, A.; Jha, B. Differential distribution and abundance of diazotrophic bacterial communities across different soil niches using a gene-targeted clone library approach. FEMS Microbiol. Lett. 2014, 360, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Jayamani, I.; Cupples, A.M. Stable Isotope Probing and High-Throughput Sequencing ImplicateXanthomonadaceae and Rhodocyclaceae in Ethylbenzene Degradation. Environ. Eng. Sci. 2015, 32, 240–249. [Google Scholar] [CrossRef]
- Liu, F.; Conrad, R. Chemolithotrophic acetogenic H2/CO2 utilization in Italian rice field soil. ISME J. 2011, 5, 1526–1539. [Google Scholar] [CrossRef]
- Kumar, L.; Futschik, M. Mfuzz: A software package for soft clustering of microarray data. Bioinformation 2007, 2, 5–7. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Fujisawa, S.; Nakai, S.; Hosomi, M. Biodegradation of natural and synthetic estrogens by nitrifying activated sludge and ammonia-oxidizing bacterium Nitrosomonas europaea. Water Res. 2004, 38, 2322–2329. [Google Scholar] [CrossRef] [PubMed]
- Mayilraj, S.; Brandt, E.S.E. The Family Paenibacillaceae; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Goldfarb, K.C.; Karaoz, U.; Hanson, C.A.; Santee, C.A.; Bradford, M.A.; Treseder, K.K.; Wallenstein, M.D.; Brodie, E.L. Differential Growth Responses of Soil Bacterial Taxa to Carbon Substrates of Varying Chemical Recalcitrance. Front. Microbiol. 2011, 2, 94. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Chen, S.; Liu, X.; Wang, K.; Yang, Y. Effect of soil sampling designs on soil microbial alpha diversity. Microbiol. China 2020, 47, 1699–1708. [Google Scholar] [CrossRef]
- Riley, D.; Barber, S.A. Bicarbonate Accumulation and pH Changes at the Soybean (Glycine max (L.) Merr.) Root-Soil Interface1. Soil Sci. Soc. Am. J. 1969, 33, 905–908. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000; pp. 265–267. [Google Scholar]
- Qin, L.; Huang, S.Q.; Zhong, L.L.; Zhou, H.; Zhao, S.; Xiang, B.; Lei, S.R. Comparison of Dumas combustion and Kjeldahl methods for determining total nitrogen content in soil. Soil Fertil. Sci. China 2020, 4, 258–265. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Du, Q.W.; Liu, Y.; Han, G.; Wang, W. Comparison of Three Commonly Used Methods for Measuring Soil Organic Matter. Heilongjiang Agric. Sci. 2014, 46, 163. [Google Scholar]
- Ye, X.S.; Wu, X.R.; Zhao, Z.Q. Comparison on the Determination of Soil Total Phosphorus Concentration by Continuous Flow Injection Analyzer and Traditional Analysis Methods. Lab. Res. Explor. 2010, 29, 50–53. [Google Scholar]
- Wang, F. Study on the Potassium Determination in Soil by Automatic Digestion Equipment-Flame Photometry Method. Ningxia Agric. For. Sci. Technol. 2017, 58, 38–39. [Google Scholar]
- Guo, Y.F. Discussion on the Determination of Quick-acting Potassium in Soil by Using Ammonium Acetate Flame Photometry and M3 Method and the Correlation Study. Shanxi Sci. Technol. 2014, 29, 54–56. [Google Scholar]
- Bu, Y.S.; Magdoff, F.R. A comparison of ten methods for determination of available phosphorus in soils. Acta Pedol. Sin. 2003, 40, 140–146. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, J.; Lu, M.; Qin, C.; Chen, Y.; Yang, L.; Huang, Q.; Wang, J.; Shen, Z.; Shen, Q. Microbial communities of an arable soil treated for 8 years with organic and inorganic fertilizers. Biol. Fert. Soils 2016, 52, 455–467. [Google Scholar] [CrossRef]
- Wang, Y.; Li, T.; Li, C.; Song, F. Differences in Microbial Community and Metabolites in Litter Layer of Plantation and Original Korean Pine Forests in North Temperate Zone. Microorganisms 2020, 8, 2023. [Google Scholar] [CrossRef]
- Knights, D.; Walters, W.A.; Peña, A.G.; Pirrung, M.; Stombaugh, J.; Caporaso, J.G.; Goodrich, J.K.; Bittinger, K.; Lozupone, C.A.; Costello, E.K.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Yang, S.; Liebner, S.; Alawi, M.; Ebenh, H.O.; Wagner, D. Taxonomic database and cut-off value for processing mcrA gene 454 pyrosequencing data by MOTHUR. J. Microbiol. Meth. 2014, 103, 3–5. [Google Scholar] [CrossRef]
- Zhu, H.T.; Chen, L.; Tu, Z.L.; Deng, X.H. Application of DPS Data Processing System in Data Processing of Pharmaceutical Research. Her. Med. 2006, 363–365. [Google Scholar]
- Jia, G.H.; Wang, Y.R.; Han, G.D. Application comparison of Excel_2003 and SPSS_13.0 software in t test. Chin. Med. J. Metall. Indus. 2014, 31, 113–114. [Google Scholar]
- Press, P. MyGraphicsLab Illustrator Course with Adobe Illustrator CS6 Classroom in a Book (Student Edition). Pearson Schweiz Ag, 10 March 2014. [Google Scholar]
- Chen, H.; Boutros, P.C. VennDiagram: A package for the generation of highly-customizable Venn and Euler diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef]
- Pang, Z.; Tayyab, M.; Kong, C.; Hu, C.; Zhu, Z.; Wei, X.; Yuan, Z. Liming Positively Modulates Microbial Community Composition and Function of Sugarcane Fields. Agronomy 2019, 9, 808. [Google Scholar] [CrossRef]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed]
- Caumo, M.; Freitas, E.M.D.; Silva, V.L.D.; Toldi, M.; Alves, L.D.S.; Orlandi, C.R.; Fior, C.S. Grassland community structure in Permanent Preservation Areas associated with forestry and livestock in the Pampa biome, Southern Brazil. S. Afr. J. Bot. 2021, 139, 442–448. [Google Scholar] [CrossRef]
- Wang, Y.; Ouyang, Z.; Zheng, H.; Wang, X.; Chen, F.; Zeng, J. Relationships between carbon source utilization of soil microbial communities and environmental factors in natural secondary forest in subtropical area, China. Acta Ecol. Sin. 2012, 32, 1839–1845. [Google Scholar] [CrossRef]
- Sunagawa, S.; Coelho, L.P.; Chaffron, S.; Kultima, J.R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D.R.; Alberti, A.; et al. Ocean plankton. Structure and function of the global ocean microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xiang, C.J.; Zhang, C.; Xu, B.Q.; Wu, J.; Wang, R.P.; Yang, Y.P.; Shi, L.Y.; Zhang, J.F.; Zhan, Z. Microbial biomarkers of common tongue coatings in patients with gastric cancer. Microb. Pathog. 2018, 127, 97–105. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| May | July | Sep | ||||
|---|---|---|---|---|---|---|
| R2 | p | R2 | p | R2 | p | |
| July | 0.967 | 0.001 ** | ||||
| Sep | 0.972 | 0.001 ** | 0.382 | 0.001 ** | ||
| Nov | 0.952 | 0.001 ** | 0.618 | 0.001 ** | 0.373 | 0.001 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Z.; Liu, Q.; Pang, Z.; Fallah, N.; Liu, Y.; Hu, C.; Lin, W. Sugarcane Rhizosphere Bacteria Community Migration Correlates with Growth Stages and Soil Nutrient. Int. J. Mol. Sci. 2022, 23, 10303. https://doi.org/10.3390/ijms231810303
Yuan Z, Liu Q, Pang Z, Fallah N, Liu Y, Hu C, Lin W. Sugarcane Rhizosphere Bacteria Community Migration Correlates with Growth Stages and Soil Nutrient. International Journal of Molecular Sciences. 2022; 23(18):10303. https://doi.org/10.3390/ijms231810303
Chicago/Turabian StyleYuan, Zhaonian, Qiang Liu, Ziqin Pang, Nyumah Fallah, Yueming Liu, Chaohua Hu, and Wenxiong Lin. 2022. "Sugarcane Rhizosphere Bacteria Community Migration Correlates with Growth Stages and Soil Nutrient" International Journal of Molecular Sciences 23, no. 18: 10303. https://doi.org/10.3390/ijms231810303
APA StyleYuan, Z., Liu, Q., Pang, Z., Fallah, N., Liu, Y., Hu, C., & Lin, W. (2022). Sugarcane Rhizosphere Bacteria Community Migration Correlates with Growth Stages and Soil Nutrient. International Journal of Molecular Sciences, 23(18), 10303. https://doi.org/10.3390/ijms231810303