
MC180295 Inhibited Epstein–Barr Virus-Associated Gastric Carcinoma Cell Growth by Suppressing DNA Repair and the Cell Cycle
 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Growth Inhibition by MC180295 in EBV-Associated Gastric Cancer Cells
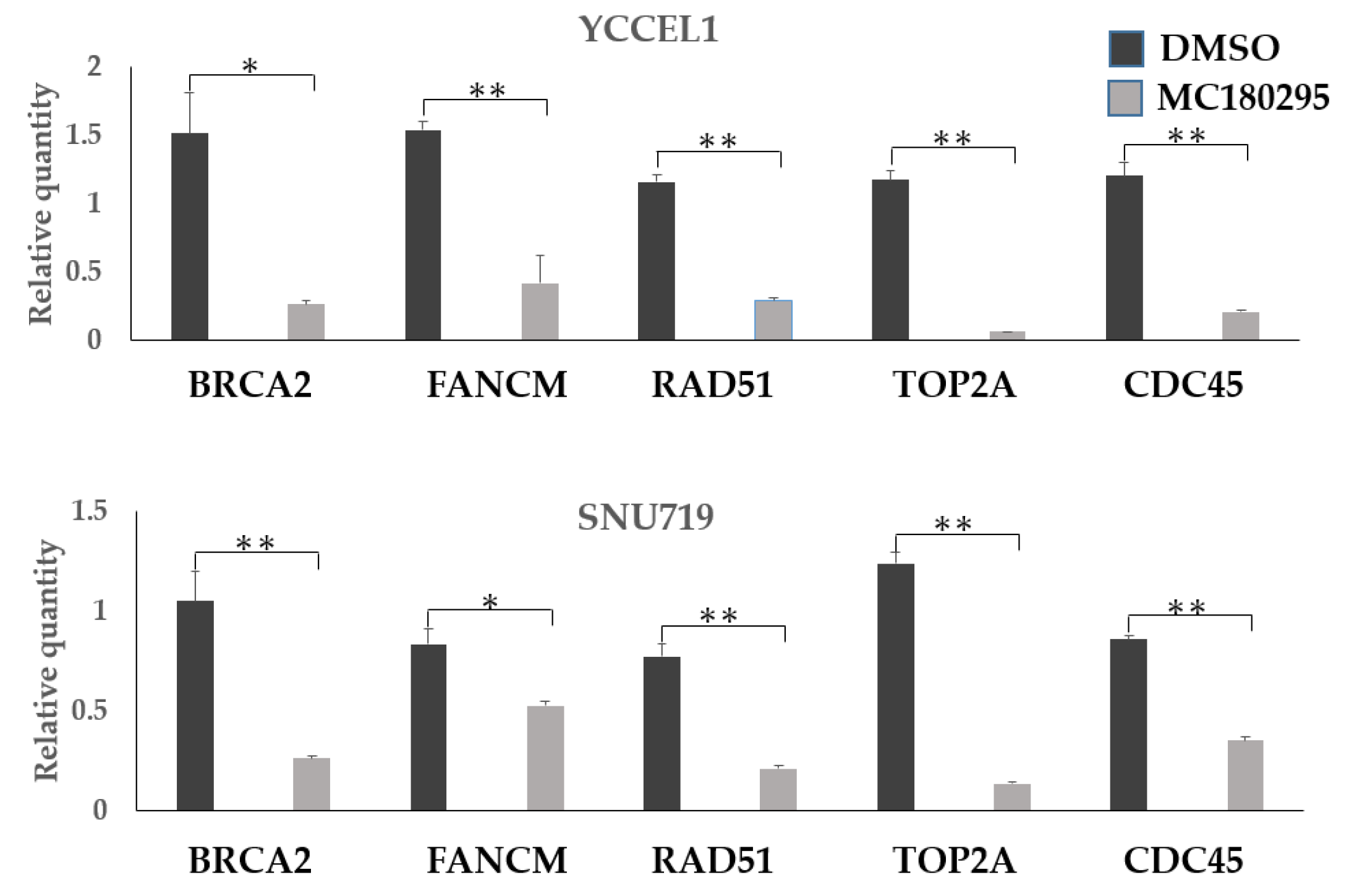
2.2. Patterns of Gene Expression after MC180295 Treatment
2.3. Protein Expression of γH2A Histone Family Member X(γH2AX) and Caspase-3 after MC180295 Treatment
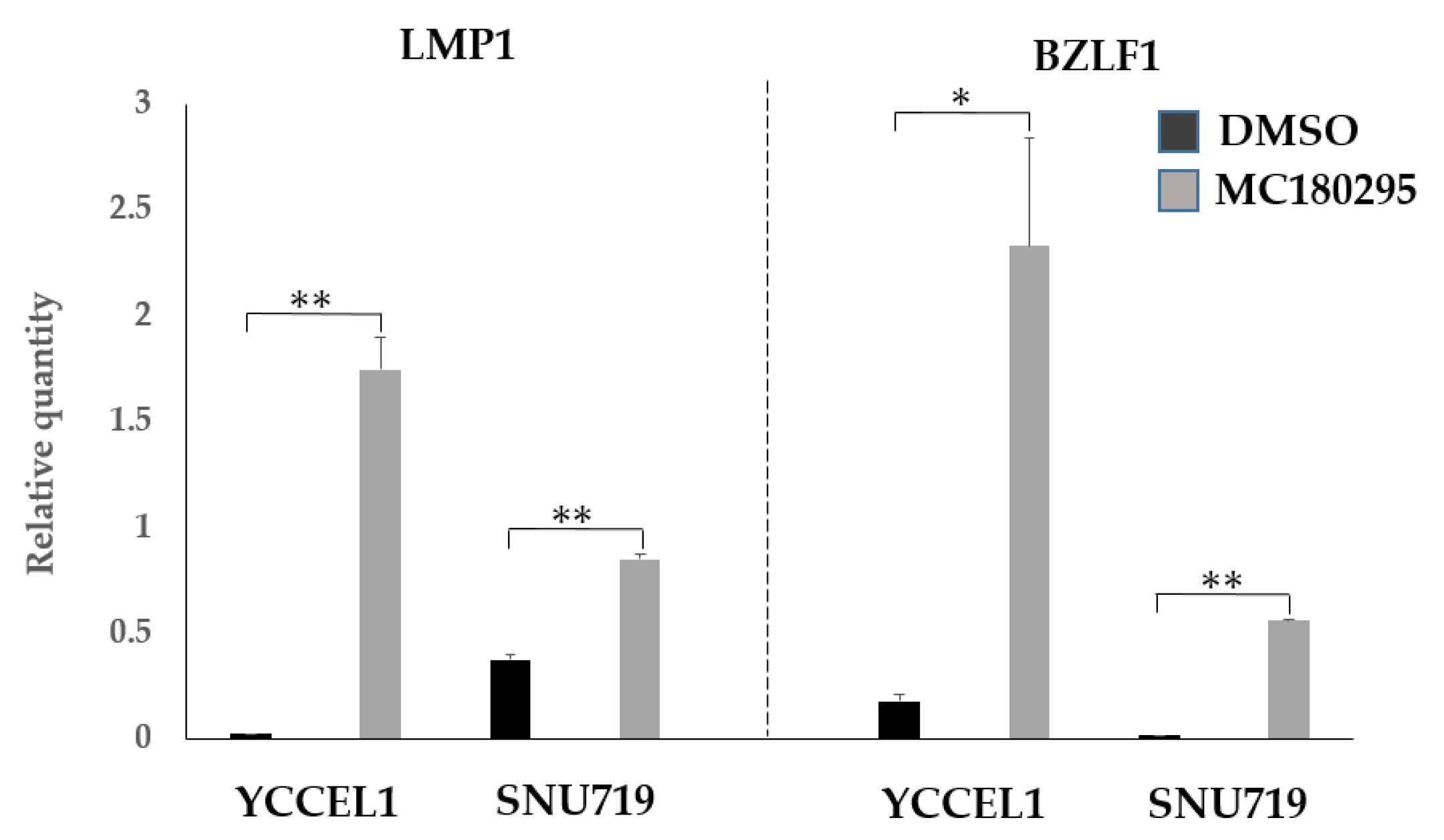
2.4. Expression of EBV Genes LMP1 and BZLF1 after MC180295 Treatment
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Cycle Analysis
4.3. RNA Extraction and SAGE Analysis
4.4. Real-Time PCR
4.5. Western Blot for γH2AX and Caspase-3
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zur Hausen, H.; Schulte-Holthausen, H.; Klein, G.; Henle, W.; Henle, G.; Clifford, P.; Santesson, L. EBV DNA in biopsies of Burkitt tumours and anaplastic carcinomas of the nasopharynx. Nature 1970, 228, 1056–1058. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Murphy, G.; Pfeiffer, R.; Camargo, M.C.; Rabkin, C.S. Meta-analysis shows that prevalence of Epstein-Barr virus-positive gastric cancer differs based on sex and anatomic location. Gastroenterology 2009, 137, 824–833. [Google Scholar] [CrossRef]
- Ricci, A.D.; Rizzo, A.; Rojas Llimpe, F.L.; Di Fabio, F.; De Biase, D.; Rihawi, K. Novel HER2-directed treatments in advanced gastric carcinoma: AnotHER paradigm shift? Cancers 2021, 13, 1664. [Google Scholar] [CrossRef]
- Rihawi, K.; Ricci, A.D.; Rizzo, A.; Brocchi, S.; Marasco, G.; Pastore, L.V.; Llimpe, F.L.R.; Golfieri, R.; Renzulli, M. Tumor-associated macrophages and inflammatory microenvironment in gastric cancer: Novel translational implications. Int. J. Mol. Sci. 2021, 22, 3805. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network. Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014, 513, 202–209. [Google Scholar] [CrossRef]
- Masucci, M.G.; Contreras-Salazar, B.; Ragnar, E.; Falk, K.; Minarovits, J.; Ernberg, I.; Klein, G. 5-Azacytidine up regulates the expression of Epstein-Barr virus nuclear antigen 2 (EBNA-2) through EBNA-6 and latent membrane protein in the Burkitt’s lymphoma line rael. J. Virol. 1989, 63, 3135–3141. [Google Scholar] [CrossRef]
- Imai, S.; Koizumi, S.; Sugiura, M.; Tokunaga, M.; Uemura, Y.; Yamamoto, N.; Tanaka, S.; Sato, E.; Osato, T. Gastric carcinoma: Monoclonal epithelial malignant cells expressing Epstein-Barr virus latent infection protein. Proc. Natl. Acad. Sci. USA 1994, 91, 9131–9135. [Google Scholar] [CrossRef]
- Kaneda, A.; Matsusaka, K.; Aburatani, H.; Fukayama, M. Epstein-Barr virus infection as an epigenetic driver of tumorigenesis. Cancer Res. 2012, 72, 3445–3450. [Google Scholar] [CrossRef]
- Nishikawa, J.; Iizasa, H.; Yoshiyama, H.; Nakamura, M.; Saito, M.; Sasaki, S.; Shimokuri, K.; Yanagihara, M.; Sakai, K.; Suehiro, Y.; et al. The role of epigenetic regulation in Epstein-Barr virus-associated gastric cancer. Int. J. Mol. Sci. 2017, 18, 1606. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Pandey, S.; Travers, M.; Sun, H.; Morton, G.; Madzo, J.; Chung, W.; Khowsathit, J.; Perez-Leal, O.; Barrero, C.A.; et al. Targeting CDK9 reactivates epigenetically silenced genes in cancer. Cell 2018, 175, 1244–1258. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, N.; Ushijima, T.; Tan, P. How to stomach an epigenetic insult: The gastric cancer epigenome. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Schneider, B.J.; Shah, M.A.; Klute, K.; Ocean, A.; Popa, E.; Altorki, N.; Lieberman, M.; Schreiner, A.; Yantiss, R.; Christos, P.J.; et al. Phase I study of epigenetic priming with azacitidine prior to standard neoadjuvant chemotherapy for patients with resectable gastric and esophageal adenocarcinoma: Evidence of tumor hypomethylation as an indicator of major histopathologic response. Clin. Cancer Res. 2017, 23, 2673–2680. [Google Scholar] [CrossRef] [PubMed]
- Yoo, C.; Ryu, M.H.; Na, Y.S.; Ryoo, B.Y.; Lee, C.W.; Kang, Y.K. Vorinostat in combination with capecitabine plus cisplatin as a first-line chemotherapy for patients with metastatic or unresectable gastric cancer: Phase II study and biomarker analysis. Br. J. Cancer 2016, 114, 1185–1190. [Google Scholar] [CrossRef]
- Zhao, W.; Steinfeld, J.B.; Liang, F.; Chen, X.; Maranon, D.G.; Jian Ma, C.; Kwon, Y.; Rao, T.; Wang, W.; Sheng, C.; et al. BRCA1-BARD1 promotes RAD51-mediated homologous DNA pairing. Nature 2017, 550, 360–365. [Google Scholar] [CrossRef]
- Nepomuceno, T.C.; Fernandes, V.C.; Gomes, T.T.; Carvalho, R.S.; Suarez-Kurtz, G.; Monteiro, A.N.; Carvalho, M.A. BRCA1 recruitment to damaged DNA sites is dependent on CDK9. Cell Cycle 2017, 16, 665–672. [Google Scholar] [CrossRef]
- Lord, C.J.; Ashworth, A. PARP inhibitors: Synthetic lethality in the clinic. Science 2017, 355, 1152–1158. [Google Scholar] [CrossRef]
- Godon, C.; Cordelières, F.P.; Biard, D.; Giocanti, N.; Mégnin-Chanet, F.; Hall, J.; Favaudon, V. PARP inhibition versus PARP-1 silencing: Different outcomes in terms of single-strand break repair and radiation susceptibility. Nucleic Acids Res. 2008, 36, 4454–4464. [Google Scholar] [CrossRef]
- Schultz, N.; Lopez, E.; Saleh-Gohari, N.; Helleday, T. Poly(ADP-ribose) polymerase (PARP-1) has a controlling role in homologous recombination. Nucleic Acids Res. 2003, 31, 4959–4964. [Google Scholar] [CrossRef]
- Keszthelyi, A.; Minchell, N.E.; Baxter, J. The causes and consequences of topological stress during DNA replication. Genes 2016, 7, 134. [Google Scholar] [CrossRef] [Green Version]
- Broderick, R.; Nasheuer, H.P. Regulation of Cdc45 in the cell cycle and after DNA damage. Biochem. Soc. Trans. 2009, 37, 926–930. [Google Scholar] [CrossRef]
- Zhang, R.; Xu, J.; Zhao, J.; Bai, J.H. Proliferation and invasion of colon cancer cells are suppressed by knockdown of TOP2A. J. Cell. Biochem. 2018, 119, 7256–7263. [Google Scholar] [CrossRef] [PubMed]
- Hao, T.; Yang, Y.; He, J.; Bai, J.; Zheng, Y.; Luo, Z. Knockdown of circular RNA hsa_circ_0062270 suppresses the progression of melanoma via downregulation of CDC45. Histol. Histopathol. 2022, 37, 373–383. [Google Scholar] [PubMed]
- Mah, L.J.; El-Osta, A.; Karagiannis, T.C. γH2AX: A sensitive molecular marker of DNA damage and repair. Leukemia 2010, 24, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Plesca, D.; Mazumder, S.; Almasan, A. DNA damage response and apoptosis. Methods Enzymol. 2008, 446, 107–122. [Google Scholar]
- Kalla, M.; Schmeinck, A.; Bergbauer, M.; Pich, D.; Hammerschmidt, W. AP-1 homolog BZLF1 of Epstein-Barr virus has two essential functions dependent on the epigenetic state of the viral genome. Proc. Natl. Acad. Sci. USA 2010, 107, 850–855. [Google Scholar] [CrossRef]
- Hagemeier, S.R.; Barlow, E.A.; Meng, Q.; Kenney, S.C. The cellular ataxia telangiectasia-mutated kinase promotes Epstein-Barr virus lytic reactivation in response to multiple different types of lytic reactivation-inducing stimuli. J. Virol. 2012, 86, 13360–13370. [Google Scholar] [CrossRef]
- Hu, L.F.; Minarovits, J.; Cao, S.L.; Contreras-Salazar, B.; Rymo, L.; Falk, K.; Klein, G.; Ernberg, I. Variable expression of latent membrane protein in nasopharyngeal carcinoma can be related to methylation status of the Epstein-Barr virus BNLF-1 5′-flanking region. J. Virol. 1991, 65, 1558–1567. [Google Scholar] [CrossRef]
- Bailey, S.G.; Verrall, E.; Schelcher, C.; Rhie, A.; Doherty, A.J.; Sinclair, A.J. Functional interaction between Epstein-Barr virus replication protein Zta and host DNA damage response protein 53BP1. J. Virol. 2009, 83, 11116–11122. [Google Scholar] [CrossRef]
- Chen, C.C.; Yang, Y.C.; Wang, W.H.; Chen, C.S.; Chang, L.K. Enhancement of Zta-activated lytic transcription of Epstein-Barr virus by Ku80. J. Gen. Virol. 2011, 92, 661–668. [Google Scholar] [CrossRef]
- Dreyfus, D.H.; Liu, Y.; Ghoda, L.Y.; Chang, J.T. Analysis of an ankyrin-like region in Epstein Barr Virus encoded (EBV) BZLF-1 (ZEBRA) protein: Implications for interactions with NF-κB and p53. Virol. J. 2011, 8, 422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Shirata, N.; Kudoh, A.; Iwahori, S.; Nakayama, S.; Murata, T.; Isomura, H.; Nishiyama, Y.; Tsurumi, T. Expression of Epstein-Barr virus BZLF1 immediate-early protein induces p53 degradation independent of MDM2, leading to repression of p53-mediated transcription. Virology 2009, 388, 204–211. [Google Scholar] [CrossRef]
- Yang, J.; Deng, W.; Hau, P.M.; Liu, J.; Lau, V.M.; Cheung, A.L.; Huen, M.S.; Tsao, S.W. Epstein-Barr virus BZLF1 protein impairs accumulation of host DNA damage proteins at damage sites in response to DNA damage. Lab. Investig. 2015, 95, 937–950. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.H.; Sitz, J.; Shen, Q.; Leblanc-Lacroix, A.; Campos, E.I.; Borozan, I.; Marcon, E.; Greenblatt, J.; Fradet-Turcotte, A.; Jin, D.Y.; et al. A Screen for Epstein-Barr Virus Proteins That Inhibit the DNA Damage Response Reveals a Novel Histone Binding Protein. J. Virol. 2018, 92, e00262-18. [Google Scholar] [CrossRef]
- Yoshiyama, H.; Imai, S.; Shimizu, N.; Takada, K. Epstein-Barr virus infection of human gastric carcinoma cells: Implication of the existence of a new virus receptor different from CD21. J. Virol. 1997, 71, 5688–5691. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Expression Fold Change | |||
|---|---|---|---|
| Symbol | Entrez Gene Name | YCCEL1 | SNU719 |
| E2F8 | E2F transcription factor 8 | −17.625 | −59.15 |
| FANCM | FA complementation group M | −13.337 | −12.558 |
| BRCA2 | BRCA2 DNA repair associated | −10.78 | −24.684 |
| BLM | BLM RecQ-like helicase | −6.618 | −10.5 |
| RAD51 | RAD51 recombinase | −6.016 | −9.869 |
| PLK1 | Polo-like kinase 1 | −5.603 | −4.065 |
| FANCA | FA complementation group A | −5.155 | −5.353 |
| E2F1 | E2F transcription factor 1 | −5.017 | −6.087 |
| MSH6 | mutS homolog 6 | −4.992 | −3.919 |
| BRIP1 | BRCA1-interacting protein C-terminal helicase 1 | −4.92 | −5.305 |
| Expression Fold Change | |||
|---|---|---|---|
| Symbol | Entrez Gene Name | YCCEL1 | SNU719 |
| TOP2A | DNA topoisomerase II alpha | −23.748 | −72.009 |
| CDC45 | Cell division cycle 45 | −20.479 | −16.356 |
| CDC6 | Cell division cycle 6 | −6.911 | −3.132 |
| MCM6 | Minichromosome maintenance complex component 6 | −6.511 | −5.254 |
| CDC7 | Cell division cycle 7 | −6.142 | −2.967 |
| PRIM1 | DNA primase subunit 1 | −5.285 | −3.952 |
| MCM4 | Minichromosome maintenance complex component 4 | −4.906 | −6.035 |
| POLA2 | DNA polymerase alpha 2, accessory subunit | −4.752 | −6.194 |
| POLE | DNA polymerase epsilon, catalytic subunit | −4.632 | −3.361 |
| MCM9 | Minichromosome maintenance 9 homologous recombination repair factor | −4.153 | −3.712 |
| Genes | Primer Sequence (5′–3′) | Annealing Temperature, °C | |
|---|---|---|---|
| BRCA2 | Forward | GGCTTCAAAAAGCACTCCAGATG | 60 |
| Reverse | GGATTCTGTATCTCTTGACGTTCC | ||
| FANCM | Forward | GCCGTAAACGTCAAGGCAGGAT | 60 |
| Reverse | CCATCAGGAACCATTCGTGGAC | ||
| RAD51 | Forward | TCTCTGGCAGTGATGTCCTGGA | 60 |
| Reverse | TAAAGGGCGGTGGCACTGTCTA | ||
| TOP2A | Forward | GTGGCAAGGATTCTGCTAGTCC | 60 |
| Reverse | ACCATTCAGGCTCAACACGCTG | ||
| CDC45 | Forward | TGGATGCTGTCCAAGGACCTGA | 60 |
| Reverse | CAGGACACCAACATCAGTCACG | ||
| LMP1 | Forward | AATCTGGATGTATTACCATGGACAAC | 60 |
| Reverse | GCGGGAGGGAGTCATCGT | ||
| BZLF1 | Forward | CTGCGCCTCCTGTTGAAG | 60 |
| Reverse | TTAAGAGATCCTCGTGTAAAACATCT | ||
| β-actin | Forward | GCTCCTCCTGAGCGCAAG | 60 |
| Reverse | CATCTGCTGGAAGGTGGACA | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujii, T.; Nishikawa, J.; Fukuda, S.; Kubota, N.; Nojima, J.; Fujisawa, K.; Ogawa, R.; Goto, A.; Hamabe, K.; Hashimoto, S.; et al. MC180295 Inhibited Epstein–Barr Virus-Associated Gastric Carcinoma Cell Growth by Suppressing DNA Repair and the Cell Cycle. Int. J. Mol. Sci. 2022, 23, 10597. https://doi.org/10.3390/ijms231810597
Fujii T, Nishikawa J, Fukuda S, Kubota N, Nojima J, Fujisawa K, Ogawa R, Goto A, Hamabe K, Hashimoto S, et al. MC180295 Inhibited Epstein–Barr Virus-Associated Gastric Carcinoma Cell Growth by Suppressing DNA Repair and the Cell Cycle. International Journal of Molecular Sciences. 2022; 23(18):10597. https://doi.org/10.3390/ijms231810597
Chicago/Turabian StyleFujii, Tomohiro, Jun Nishikawa, Soichiro Fukuda, Naoto Kubota, Junzo Nojima, Koichi Fujisawa, Ryo Ogawa, Atsushi Goto, Koichi Hamabe, Shinichi Hashimoto, and et al. 2022. "MC180295 Inhibited Epstein–Barr Virus-Associated Gastric Carcinoma Cell Growth by Suppressing DNA Repair and the Cell Cycle" International Journal of Molecular Sciences 23, no. 18: 10597. https://doi.org/10.3390/ijms231810597
APA StyleFujii, T., Nishikawa, J., Fukuda, S., Kubota, N., Nojima, J., Fujisawa, K., Ogawa, R., Goto, A., Hamabe, K., Hashimoto, S., Wai, A. P., Iizasa, H., Yoshiyama, H., Sakai, K., Suehiro, Y., Yamasaki, T., & Takami, T. (2022). MC180295 Inhibited Epstein–Barr Virus-Associated Gastric Carcinoma Cell Growth by Suppressing DNA Repair and the Cell Cycle. International Journal of Molecular Sciences, 23(18), 10597. https://doi.org/10.3390/ijms231810597