
Gene Expression and Chondrogenic Potential of Cartilage Cells: Osteoarthritis Grade Differences



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
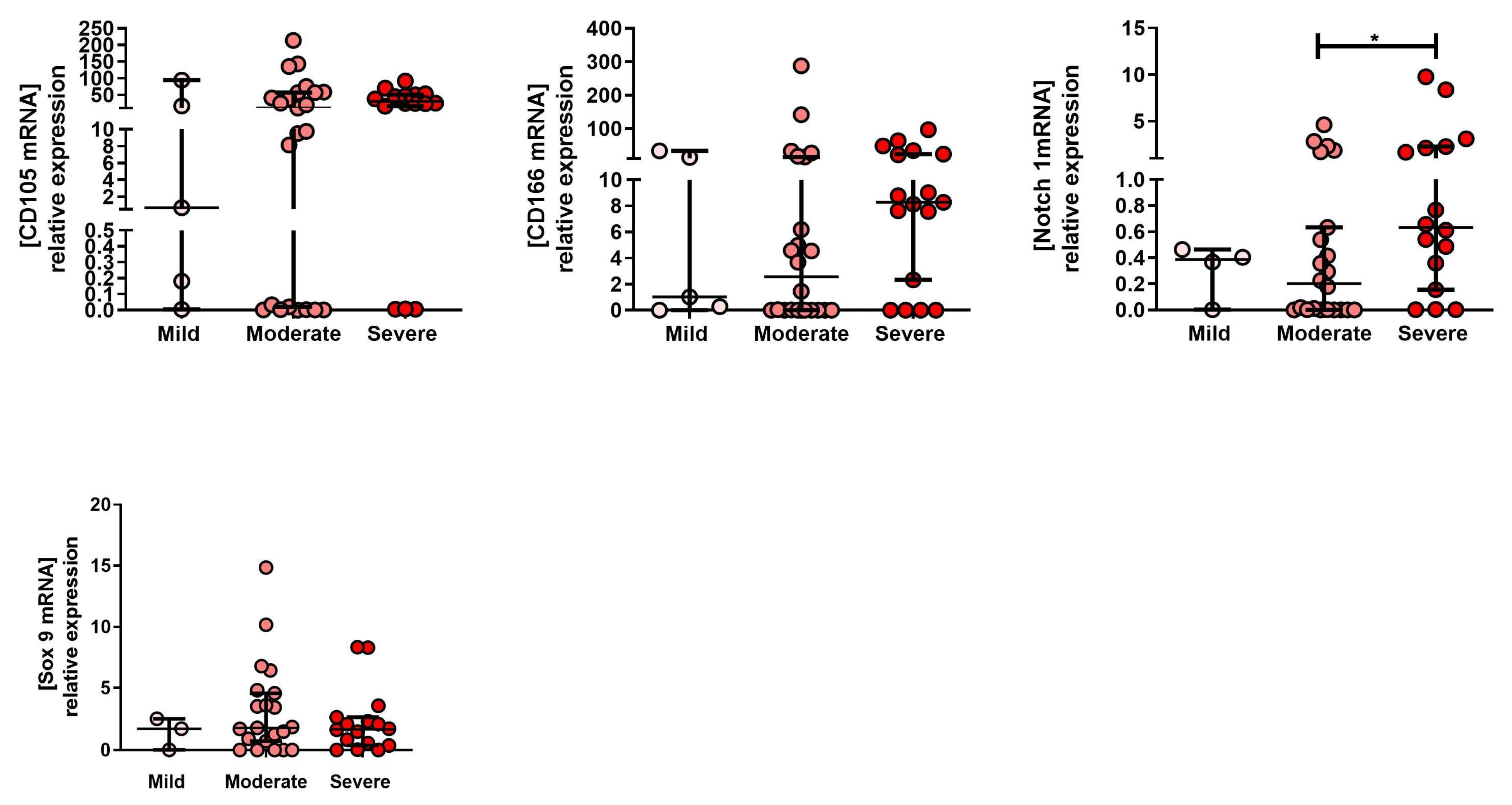
2.1. mRNA Expression Profile of MPC Markers in Tissue Isolated from Mild, Moderate and severe OA Grade
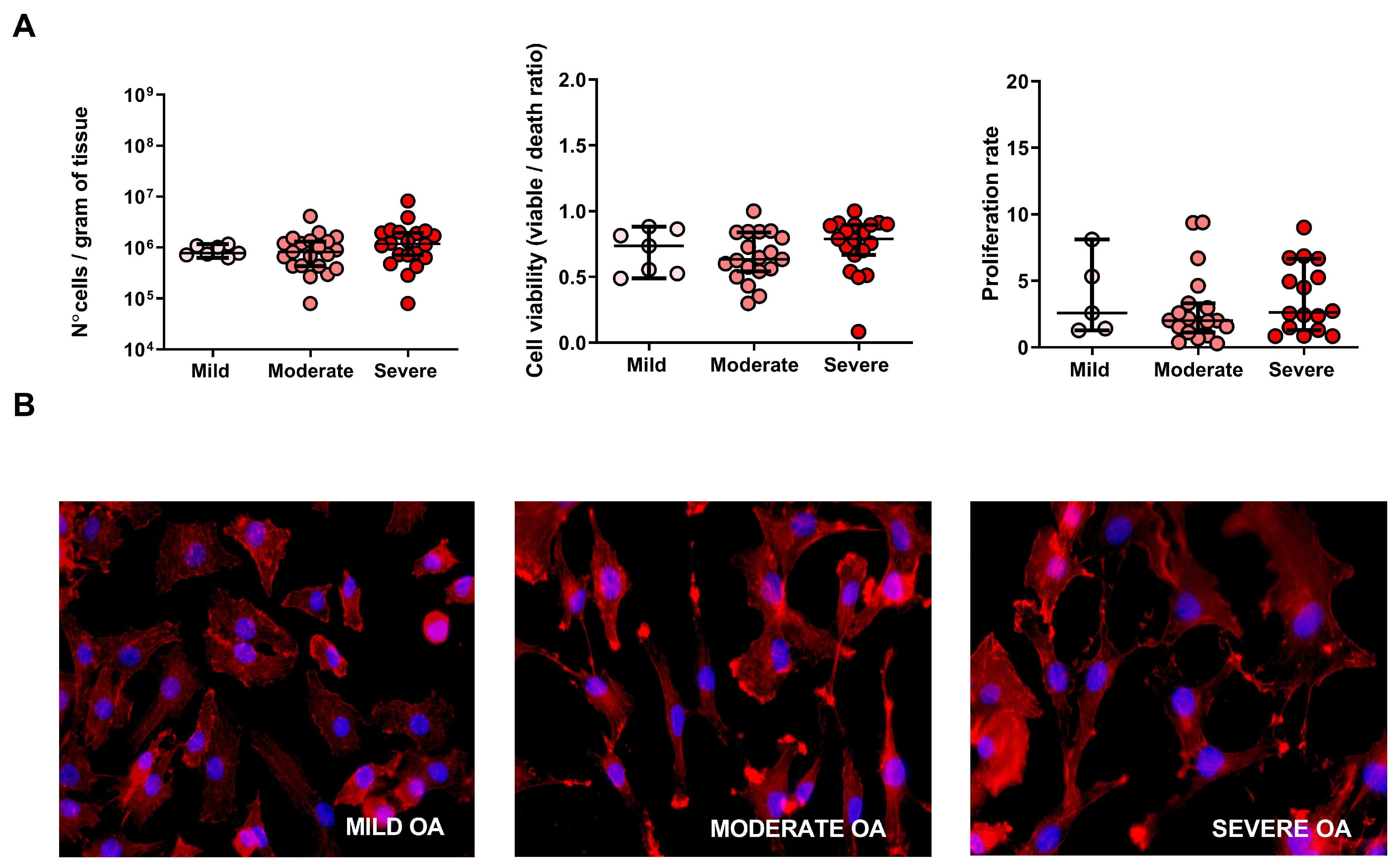
2.2. Cell Number, Viability, Morphology and Proliferation Rate of Cells Derived from Mild, Moderate and Severe OA Grade
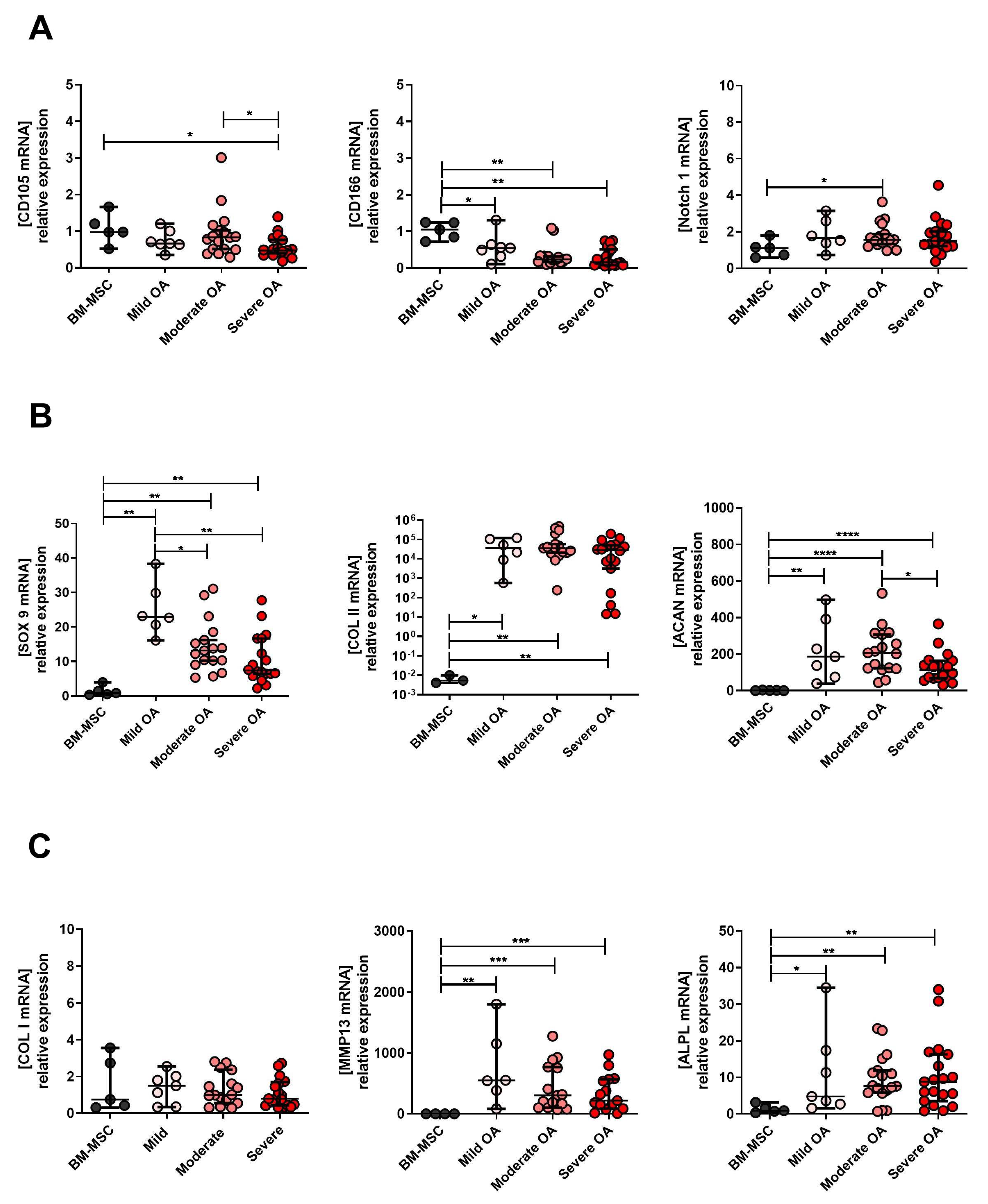
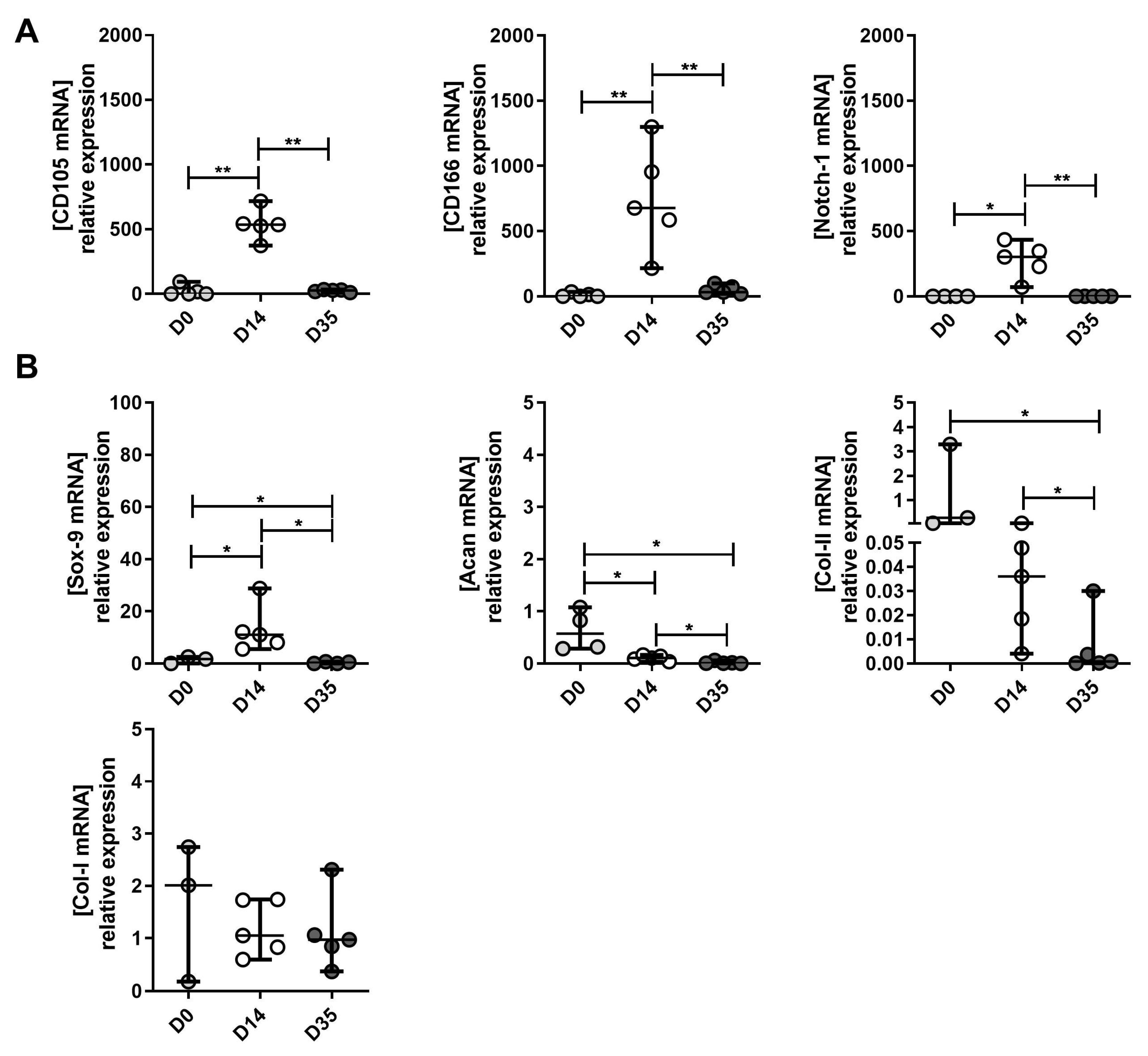
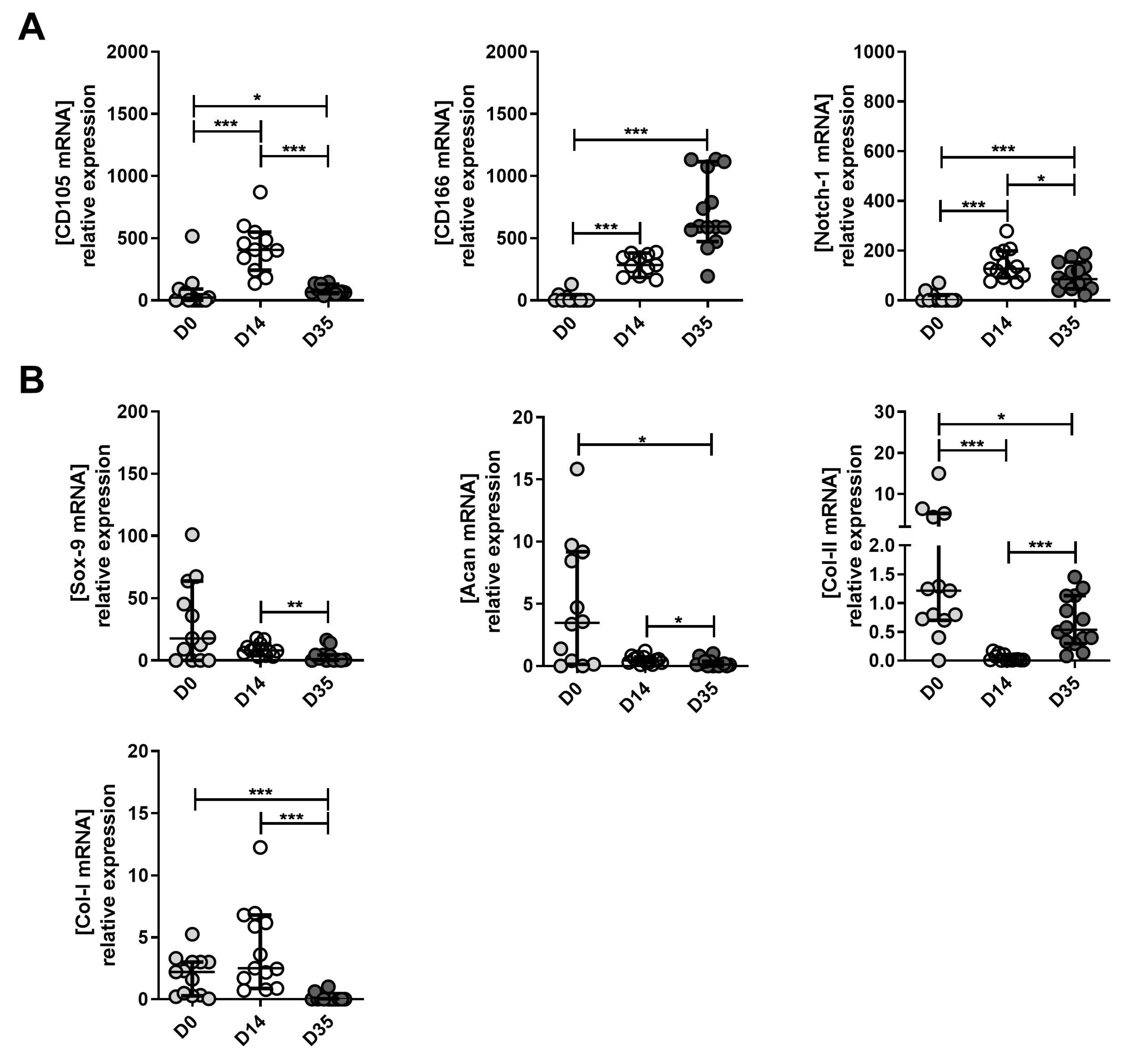
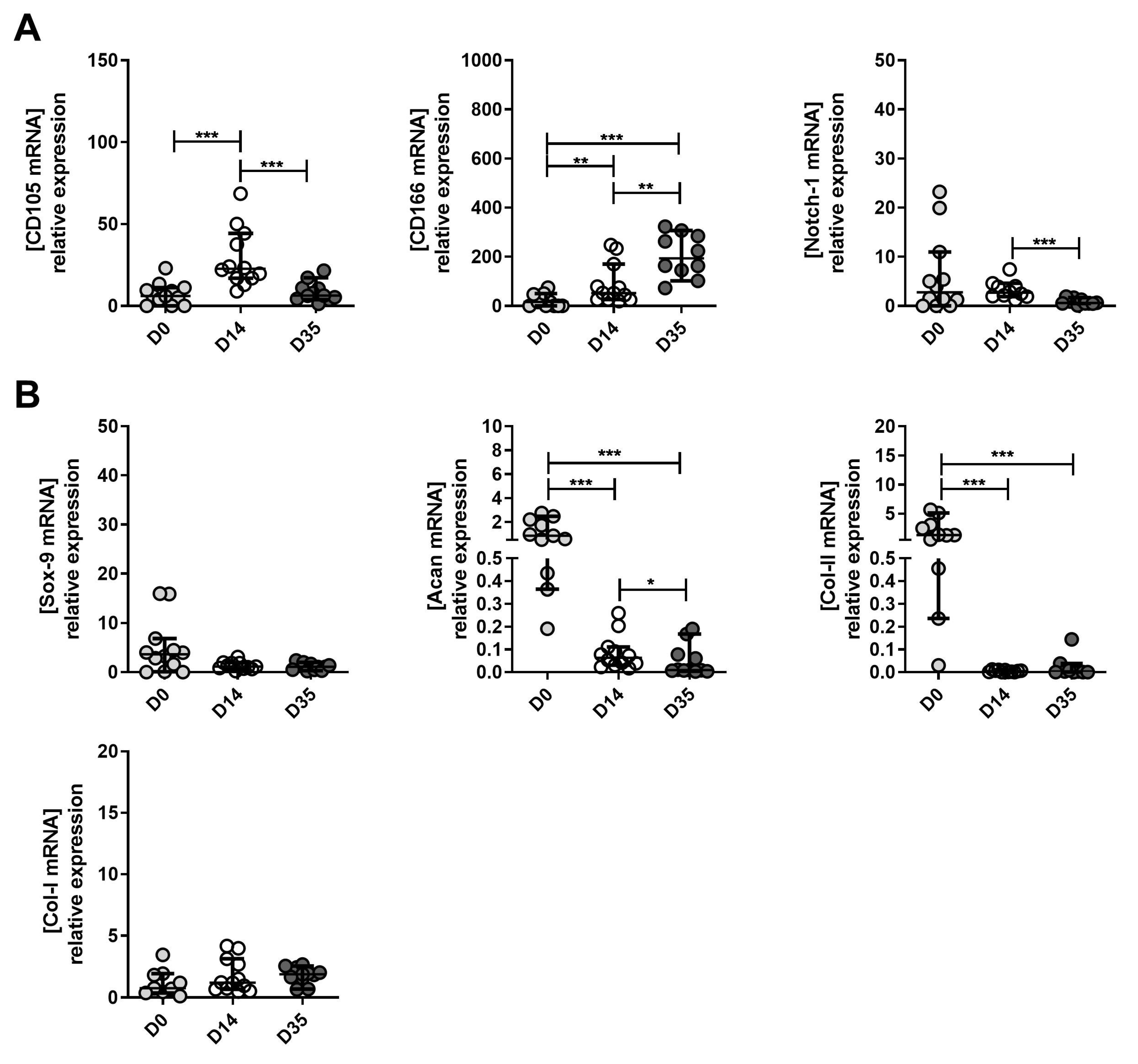
2.3. Expression Levels of MPC, Chondrocyte and Hypertrophic Chondroyte Markers after In Vitro Expansion of Cells Derived from Mild, Moderate and Severe OA Grades
2.4. The Changes in MPC, Chondrocyte and Hypertrophic Chondroyte Marker Expression between Tissue (D0), Expanded (D14) and Re-Differentiated (D35) Cells Derived from Mild, Moderate and Severe OA
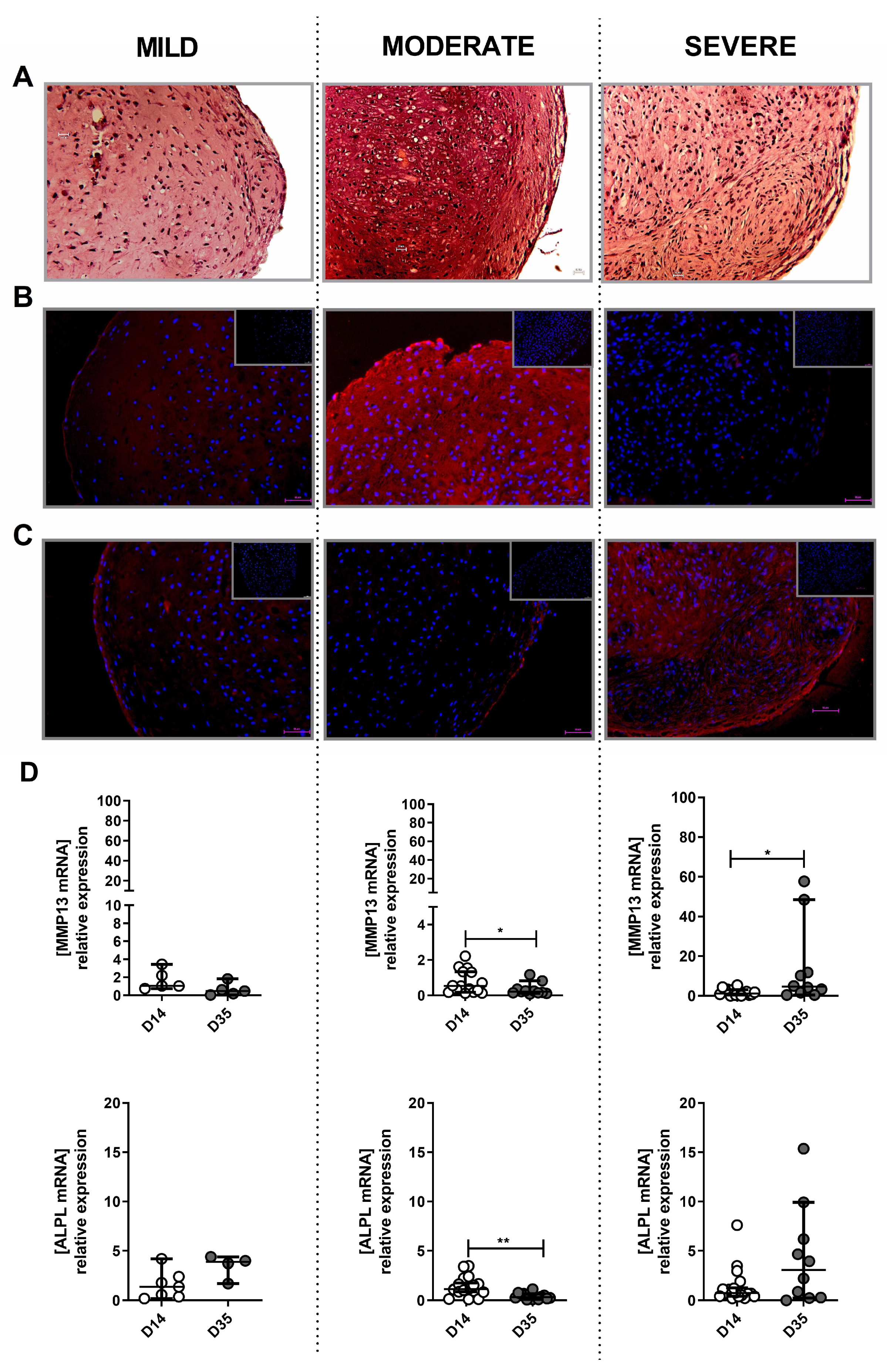
2.5. The Potential of Mild, Moderate and Severe OA-Derived Cells to Form Hyaline Cartilage-Like Tissue In Vitro
3. Discussion
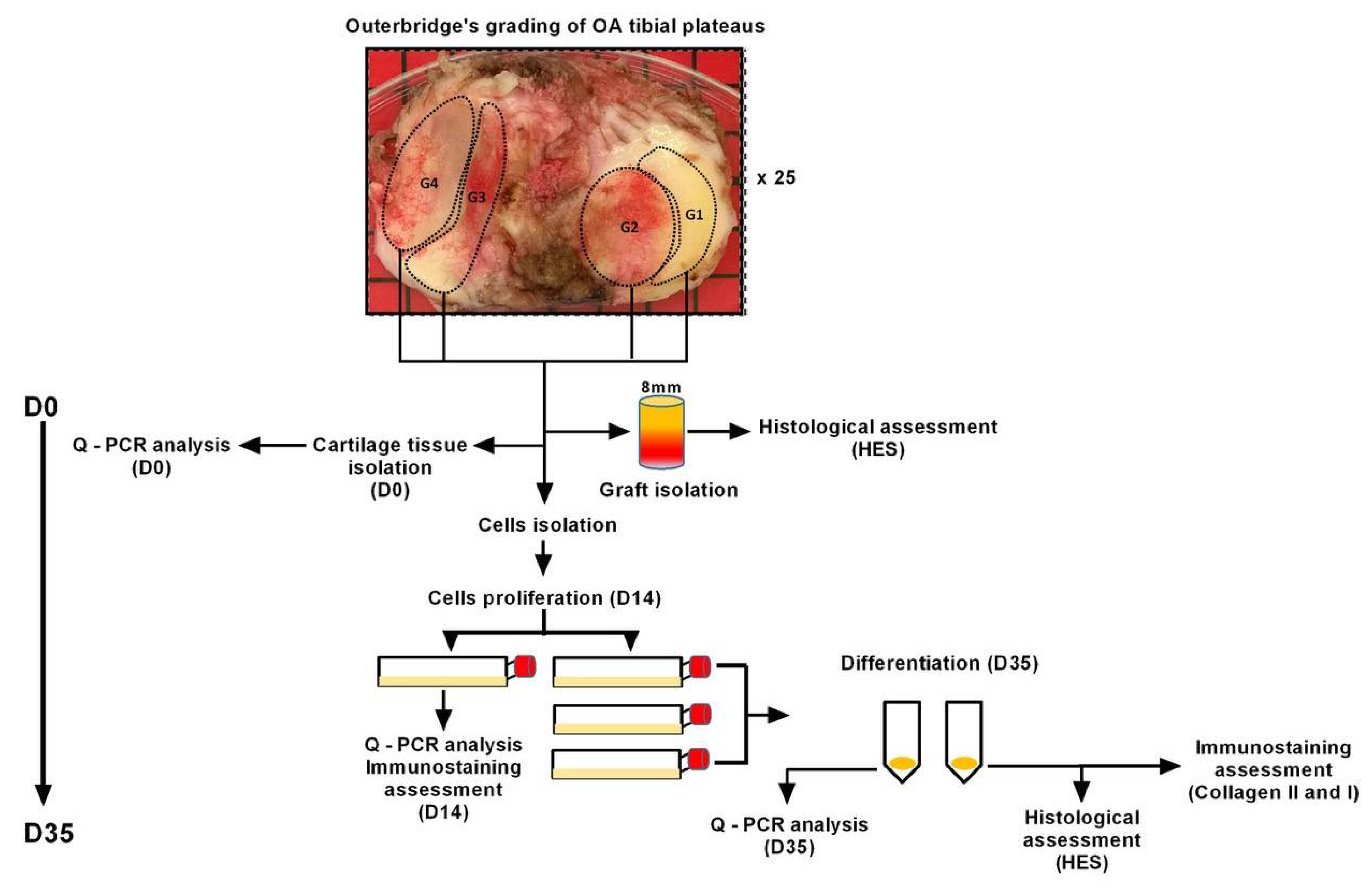
4. Materials and Methods
4.1. Cartilage Preparation
4.2. Histological Assessment of OA Grafts
4.3. Isolation and Culture of Cells from Cartilage Samples
4.4. Cell Differentiation Assay
4.5. mRNA Expression/Quantitative Real-Time PCR (qPCR)
4.6. Histology and Immunostaining
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jiang, Y.; Cai, Y.; Zhang, W.; Yin, Z.; Hu, C.; Tong, T.; Lu, P.; Zhang, S.; Neculai, D.; Tuan, R.S.; et al. Human Cartilage-Derived Progenitor Cells From Committed Chondrocytes for Efficient Cartilage Repair and Regeneration. Stem Cells Transl. Med. 2016, 5, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Goldring, M.B. Chondrogenesis, chondrocyte differentiation, and articular cartilage metabolism in health and osteoarthritis. Ther. Adv. Musculoskelet Dis. 2012, 4, 269–285. [Google Scholar] [CrossRef] [PubMed]
- Phull, A.R.; Eo, S.H.; Abbas, Q.; Ahmed, M.; Kim, S.J. Applications of Chondrocyte-Based Cartilage Engineering: An Overview. Biomed. Res. Int. 2016, 2016, 1879837. [Google Scholar] [CrossRef] [PubMed]
- Dewan, A.K.; Gibson, M.A.; Elisseeff, J.H.; Trice, M.E. Evolution of autologous chondrocyte repair and comparison to other cartilage repair techniques. Biomed. Res. Int. 2014, 2014, 272481. [Google Scholar] [CrossRef] [PubMed]
- Pretzel, D.; Linss, S.; Rochler, S.; Endres, M.; Kaps, C.; Alsalameh, S.; Kinne, R.W. Relative percentage and zonal distribution of mesenchymal progenitor cells in human osteoarthritic and normal cartilage. Arthritis Res. Ther. 2011, 13, R64. [Google Scholar] [CrossRef]
- Alsalameh, S.; Amin, R.; Gemba, T.; Lotz, M. Identification of mesenchymal progenitor cells in normal and osteoarthritic human articular cartilage. Arthritis Rheum. 2004, 50, 1522–1532. [Google Scholar] [CrossRef]
- Bernstein, P.; Sperling, I.; Corbeil, D.; Hempel, U.; Fickert, S. Progenitor cells from cartilage-no osteoarthritis-grade-specific differences in stem cell marker expression. Biotechnol. Prog. 2013, 29, 206–212. [Google Scholar] [CrossRef]
- Otsuki, S.; Grogan, S.P.; Miyaki, S.; Kinoshita, M.; Asahara, H.; Lotz, M.K. Tissue neogenesis and STRO-1 expression in immature and mature articular cartilage. J. Orthop. Res. 2010, 28, 96–102. [Google Scholar] [CrossRef]
- Li, S.; Sengers, B.G.; Oreffo, R.O.; Tare, R.S. Chondrogenic potential of human articular chondrocytes and skeletal stem cells: A comparative study. J. Biomater. Appl. 2015, 29, 824–836. [Google Scholar] [CrossRef]
- Fickert, S.; Fiedler, J.; Brenner, R.E. Identification of subpopulations with characteristics of mesenchymal progenitor cells from human osteoarthritic cartilage using triple staining for cell surface markers. Arthritis Res. Ther. 2004, 6, R422–R432. [Google Scholar] [CrossRef] [Green Version]
- Stoop, R.; Albrecht, D.; Gaissmaier, C.; Fritz, J.; Felka, T.; Rudert, M.; Aicher, W.K. Comparison of marker gene expression in chondrocytes from patients receiving autologous chondrocyte transplantation versus osteoarthritis patients. Arthritis Res. Ther. 2007, 9, R60. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.M.; Herlofsen, S.R.; Karlsen, T.A.; Küchler, A.M.; Fløisand, Y.; Brinchmann, J.E. Similar properties of chondrocytes from osteoarthritis joints and mesenchymal stem cells from healthy donors for tissue engineering of articular cartilage. PLoS ONE 2013, 8, e62994. [Google Scholar] [CrossRef] [PubMed]
- Hu, N.; Gao, Y.; Jayasuriya, C.T.; Liu, W.; Du, H.; Ding, J.; Feng, M.; Chen, Q. Chondrogenic induction of human osteoarthritic cartilage-derived mesenchymal stem cells activates mineralization and hypertrophic and osteogenic gene expression through a mechanomiR. Arthritis Res. Ther. 2019, 21, 167. [Google Scholar] [CrossRef]
- Schminke, B.; Miosge, N. Cartilage Repair In Vivo: The Role of Migratory Progenitor Cells. Curr. Rheumatol. Rep. 2014, 16. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, K.; Grogan, S.; Olee, T.; Lotz, M. Mesenchymal progenitor cells in adult human articular cartilage. Biorheology 2006, 43, 447–454. [Google Scholar] [PubMed]
- Mantripragada, V.P.; Bova, W.A.; Boehm, C.; Piuzzi, N.S.; Obuchowski, N.A.; Midura, R.J.; Muschler, G.F. Primary Cells Isolated from Human Knee Cartilage Reveal Decreased Prevalence of Progenitor Cells but Comparable Biological Potential During Osteoarthritic Disease Progression. J. Bone Joint Surg. Am. 2018, 100, 1771–1780. [Google Scholar] [CrossRef]
- Mantripragada, V.P.; Bova, W.A.; Boehm, C.; Piuzzi, N.S.; Obuchowski, N.A.; Midura, R.J.; Muschler, G.F. Progenitor cells from different zones of human cartilage and their correlation with histopathological osteoarthritis progression. J. Orthop. Res. 2018, 36, 1728–1738. [Google Scholar] [CrossRef]
- Mantripragada, V.P.; Bova, W.A.; Piuzzi, N.S.; Boehm, C.; Obuchowski, N.A.; Midura, R.J.; Muschler, G.F. Native-Osteoarthritic Joint Resident Stem and Progenitor Cells for Cartilage Cell-Based Therapies: A Quantitative Comparison With Respect to Concentration and Biological Performance. Am. J. Sports Med. 2019, 47, 3521–3530. [Google Scholar] [CrossRef]
- Grogan, S.P.; Miyaki, S.; Asahara, H.; D’Lima, D.D.; Lotz, M.K. Mesenchymal progenitor cell markers in human articular cartilage: Normal distribution and changes in osteoarthritis. Arthritis Res. Ther. 2009, 11, R85. [Google Scholar] [CrossRef]
- Agar, G.; Blumenstein, S.; Bar-Ziv, Y.; Kardosh, R.; Schrift-Tzadok, M.; Gal-Levy, R.; Fischler, T.; Goldschmid, R.; Yayon, A. The Chondrogenic Potential of Mesenchymal Cells and Chondrocytes from Osteoarthritic Subjects: A Comparative Analysis. Cartilage 2011, 2, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Vinod, E.; Amirtham, S.M.; Kachroo, U. An assessment of bone marrow mesenchymal stem cell and human articular cartilage derived chondroprogenitor cocultures vs. monocultures. Knee 2021, 29, 418–425. [Google Scholar] [CrossRef]
- Sassi, N.; Laadhar, L.; Driss, M.; Kallel-Sellami, M.; Sellami, S.; Makni, S. The role of the Notch pathway in healthy and osteoarthritic articular cartilage: From experimental models to ex vivo studies. Arthritis Res. Ther. 2011, 13, 208. [Google Scholar] [CrossRef]
- Ustunel, I.; Ozenci, A.M.; Sahin, Z.; Ozbey, O.; Acar, N.; Tanriover, G.; Celik-Ozenci, C.; Demir, R. The immunohistochemical localization of notch receptors and ligands in human articular cartilage, chondroprogenitor culture and ultrastructural characteristics of these progenitor cells. Acta Histochem. 2008, 110, 397–407. [Google Scholar] [CrossRef]
- Dehne, T.; Karlsson, C.; Ringe, J.; Sittinger, M.; Lindahl, A. Chondrogenic differentiation potential of osteoarthritic chondrocytes and their possible use in matrix-associated autologous chondrocyte transplantation. Arthritis Res. Ther. 2009, 11, R133. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, J.; Mirando, A.J.; Wang, C.; Zuscik, M.J.; O’Keefe, R.J.; Hilton, M.J. A dual role for NOTCH signaling in joint cartilage maintenance and osteoarthritis. Sci. Signal. 2015, 8, ra71. [Google Scholar] [CrossRef]
- Liu, C.-F.; Samsa, W.E.; Zhou, G.; Lefebvre, V. Transcriptional control of chondrocyte specification and differentiation. Semin. Cell Dev. Biol. 2017, 62, 34–49. [Google Scholar] [CrossRef]
- Akiyama, H.; Chaboissier, M.C.; Martin, J.F.; Schedl, A.; de Crombrugghe, B. The transcription factor Sox9 has essential roles in successive steps of the chondrocyte differentiation pathway and is required for expression of Sox5 and Sox6. Genes Dev. 2002, 16, 2813–2828. [Google Scholar] [CrossRef]
- Vinod, E.; Padmaja, K.; Livingston, A.; James, J.V.; Amirtham, S.M.; Sathishkumar, S.; Kachroo, U. Prospective Isolation and Characterization of Chondroprogenitors from Human Chondrocytes Based on CD166/CD34/CD146 Surface Markers. Cartilage 2021, 13 (Suppl. S2), 808S–817S. [Google Scholar] [CrossRef]
- Xu, J.; Wang, W.; Ludeman, M.; Cheng, K.; Hayami, T.; Lotz, J.C.; Kapila, S. Chondrogenic differentiation of human mesenchymal stem cells in three-dimensional alginate gels. Tissue Eng. Part A 2008, 14, 667–680. [Google Scholar] [CrossRef]
- Mwale, F.; Stachura, D.; Roughley, P.; Antoniou, J. Limitations of using aggrecan and type X collagen as markers of chondrogenesis in mesenchymal stem cell differentiation. J. Orthop. Res. 2006, 24, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- Brew, C.J.; Clegg, P.D.; Boot-Handford, R.P.; Andrew, J.G.; Hardingham, T. Gene expression in human chondrocytes in late osteoarthritis is changed in both fibrillated and intact cartilage without evidence of generalised chondrocyte hypertrophy. Ann. Rheum. Dis. 2010, 69, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Pritzker, K.P.H.; Gay, S.; Jimenez, S.A.; Ostergaard, K.; Pelletier, J.-P.; Revell, P.A.; Salter, D.; Van Den Berg, W.B. Osteoarthritis cartilage histopathology: Grading and staging. Osteoarthr. Cartil. 2006, 14, 13–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazor, M.; Lespessailles, E.; Best, T.M.; Ali, M.; Toumi, H. Gene Expression and Chondrogenic Potential of Cartilage Cells: Osteoarthritis Grade Differences. Int. J. Mol. Sci. 2022, 23, 10610. https://doi.org/10.3390/ijms231810610
Mazor M, Lespessailles E, Best TM, Ali M, Toumi H. Gene Expression and Chondrogenic Potential of Cartilage Cells: Osteoarthritis Grade Differences. International Journal of Molecular Sciences. 2022; 23(18):10610. https://doi.org/10.3390/ijms231810610
Chicago/Turabian StyleMazor, Marija, Eric Lespessailles, Thomas M. Best, Mazen Ali, and Hechmi Toumi. 2022. "Gene Expression and Chondrogenic Potential of Cartilage Cells: Osteoarthritis Grade Differences" International Journal of Molecular Sciences 23, no. 18: 10610. https://doi.org/10.3390/ijms231810610
APA StyleMazor, M., Lespessailles, E., Best, T. M., Ali, M., & Toumi, H. (2022). Gene Expression and Chondrogenic Potential of Cartilage Cells: Osteoarthritis Grade Differences. International Journal of Molecular Sciences, 23(18), 10610. https://doi.org/10.3390/ijms231810610