
Restoration of Cathepsin D Level via L-Serine Attenuates PPA-Induced Lysosomal Dysfunction in Neuronal Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
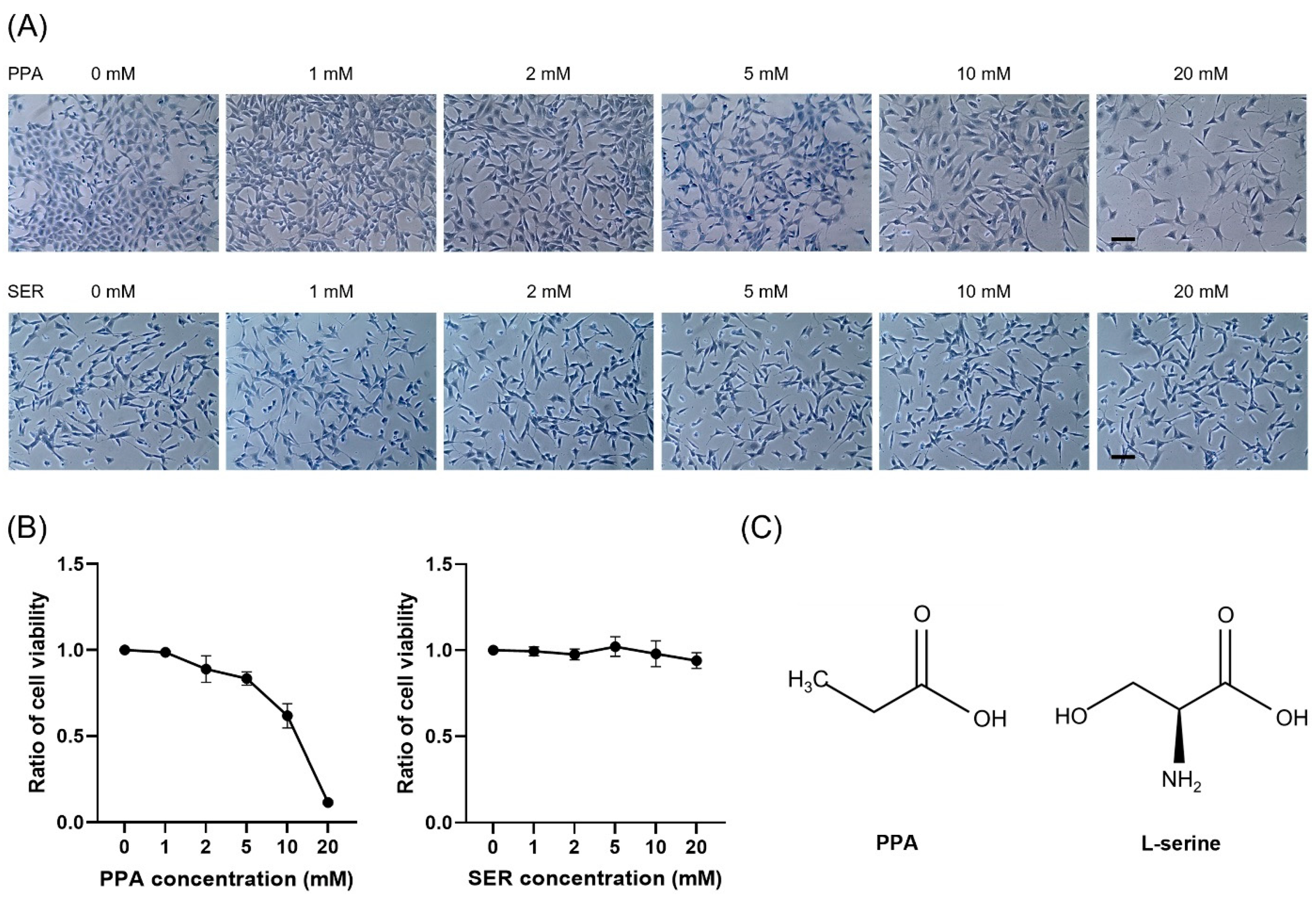
2.1. Toxicological Effects of Propionic Acid and L-Serine on HT22 Cells
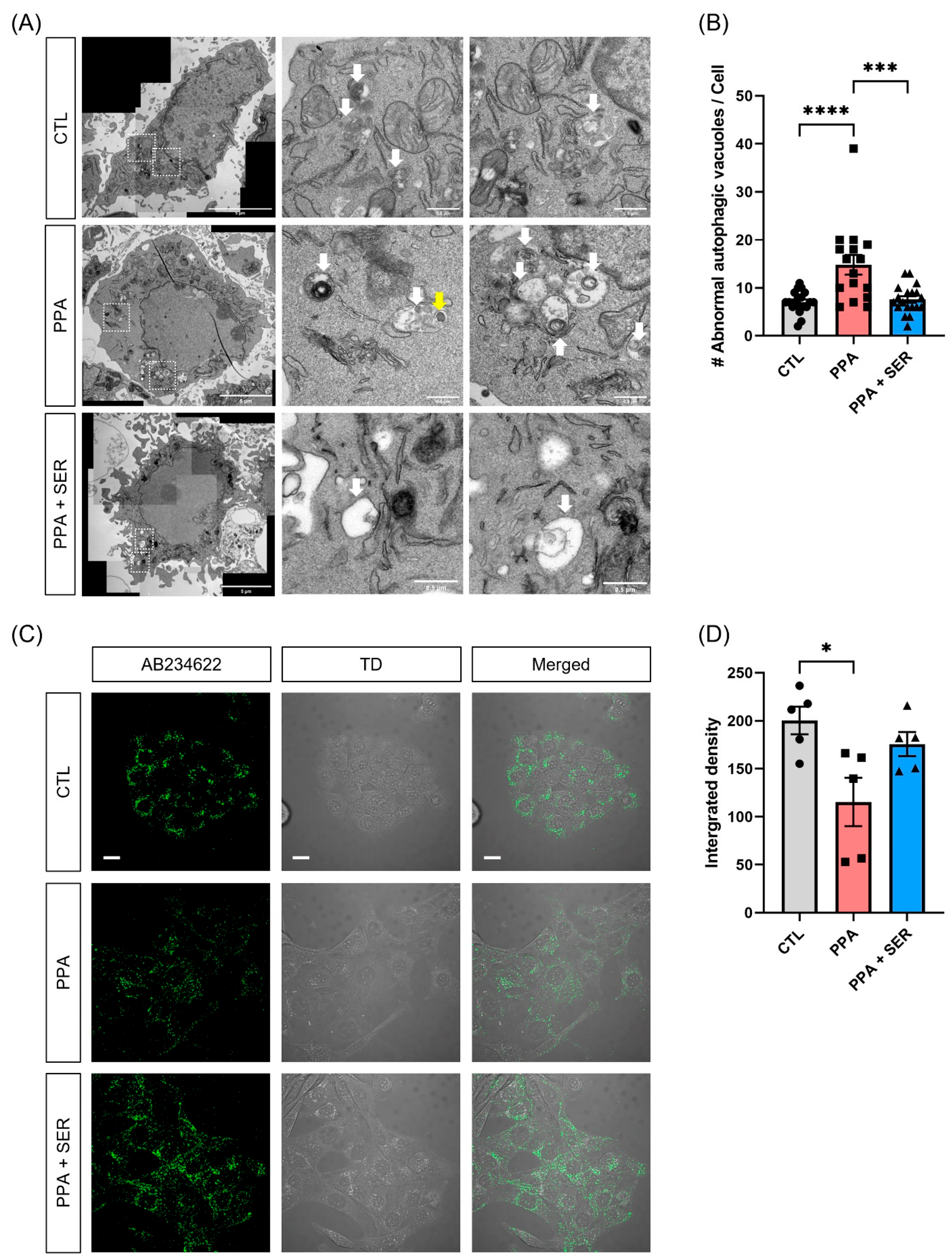
2.2. PPA Decreased Lysosomal Activity in PPA-Treated HT22 Cells, and L-Serine Attenuated the Dysfunction
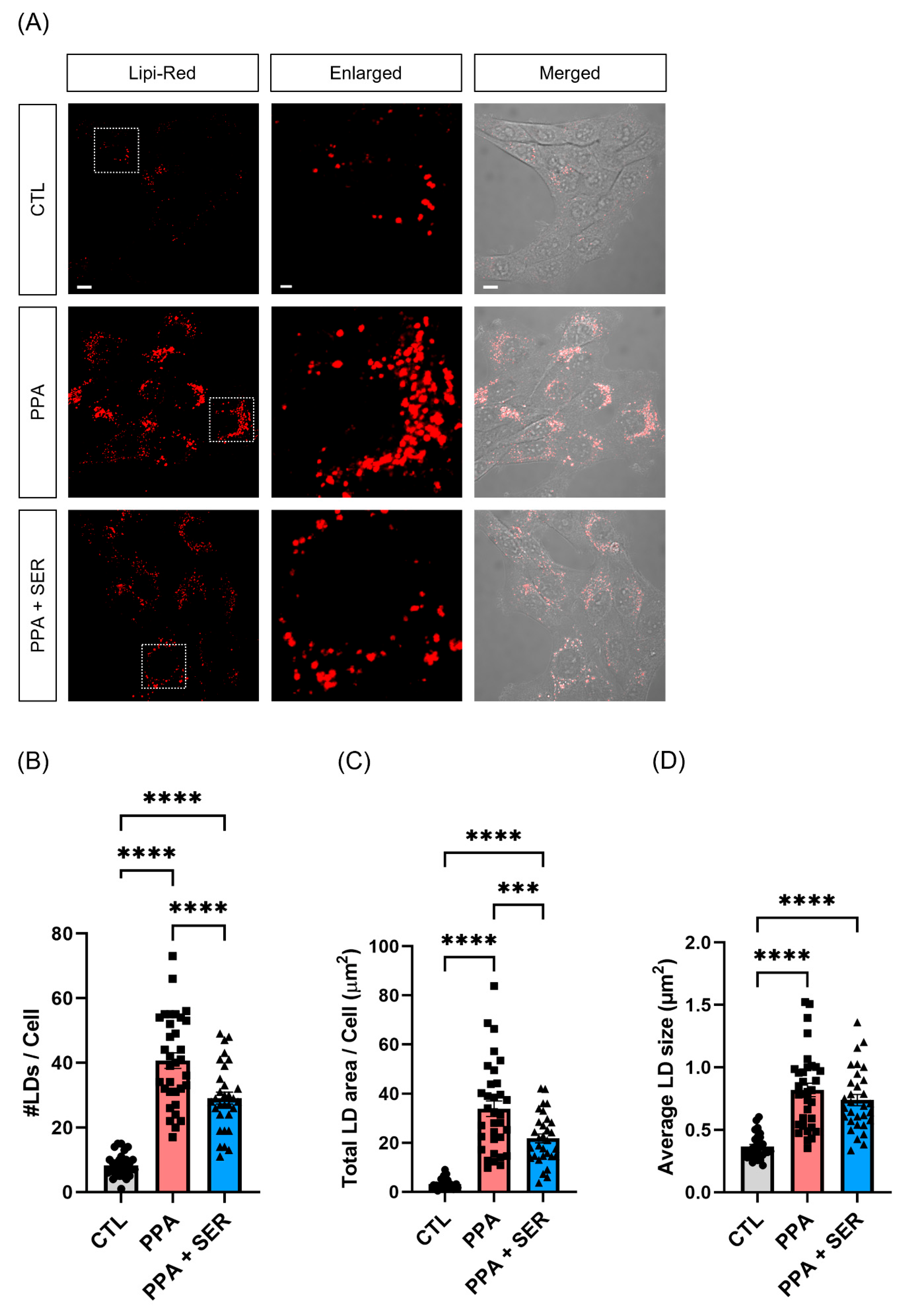
2.3. PPA Increased Lipid Droplets in HT22 Cells, and L-Serine Attenuated the Lipid Accumulation
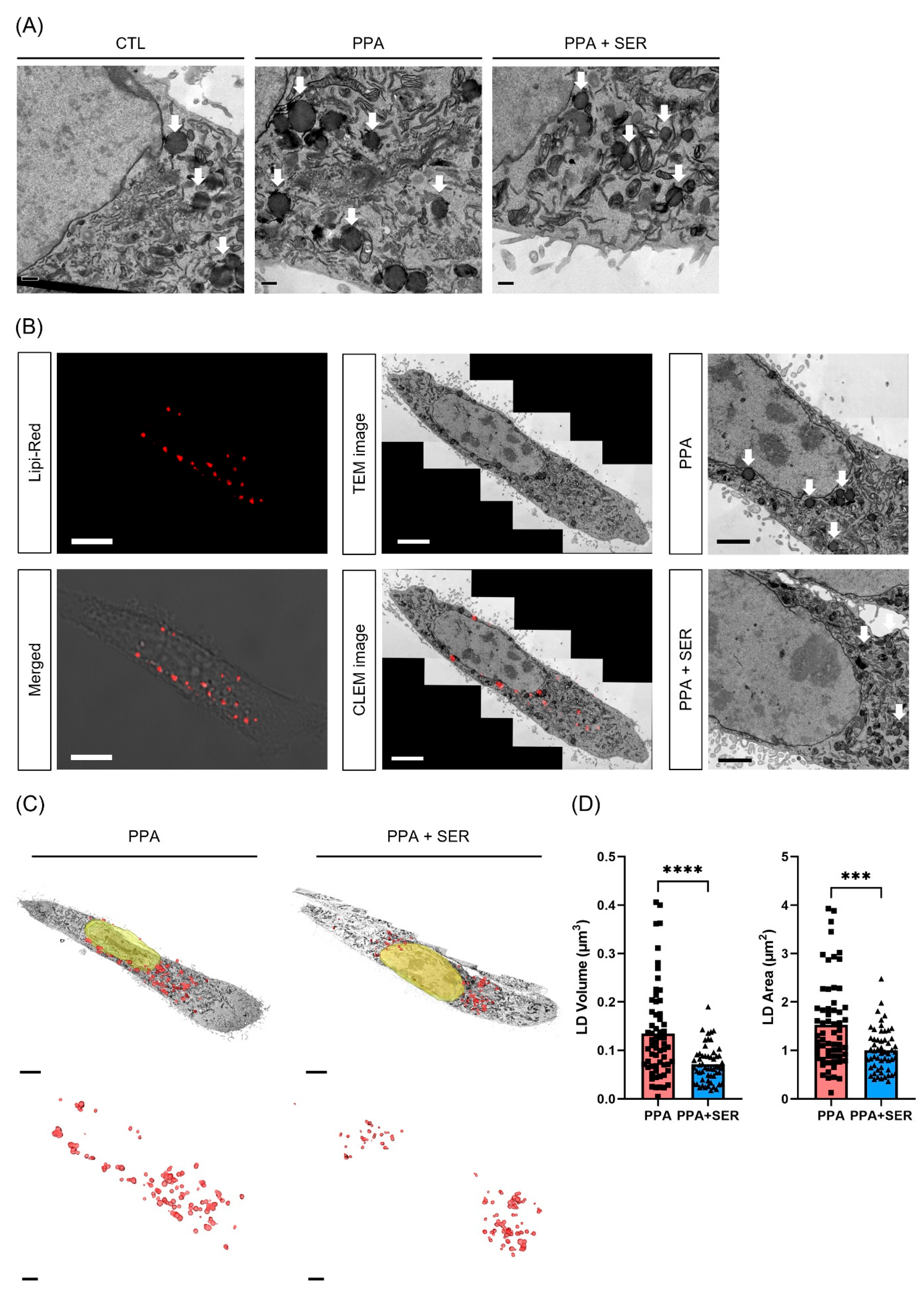
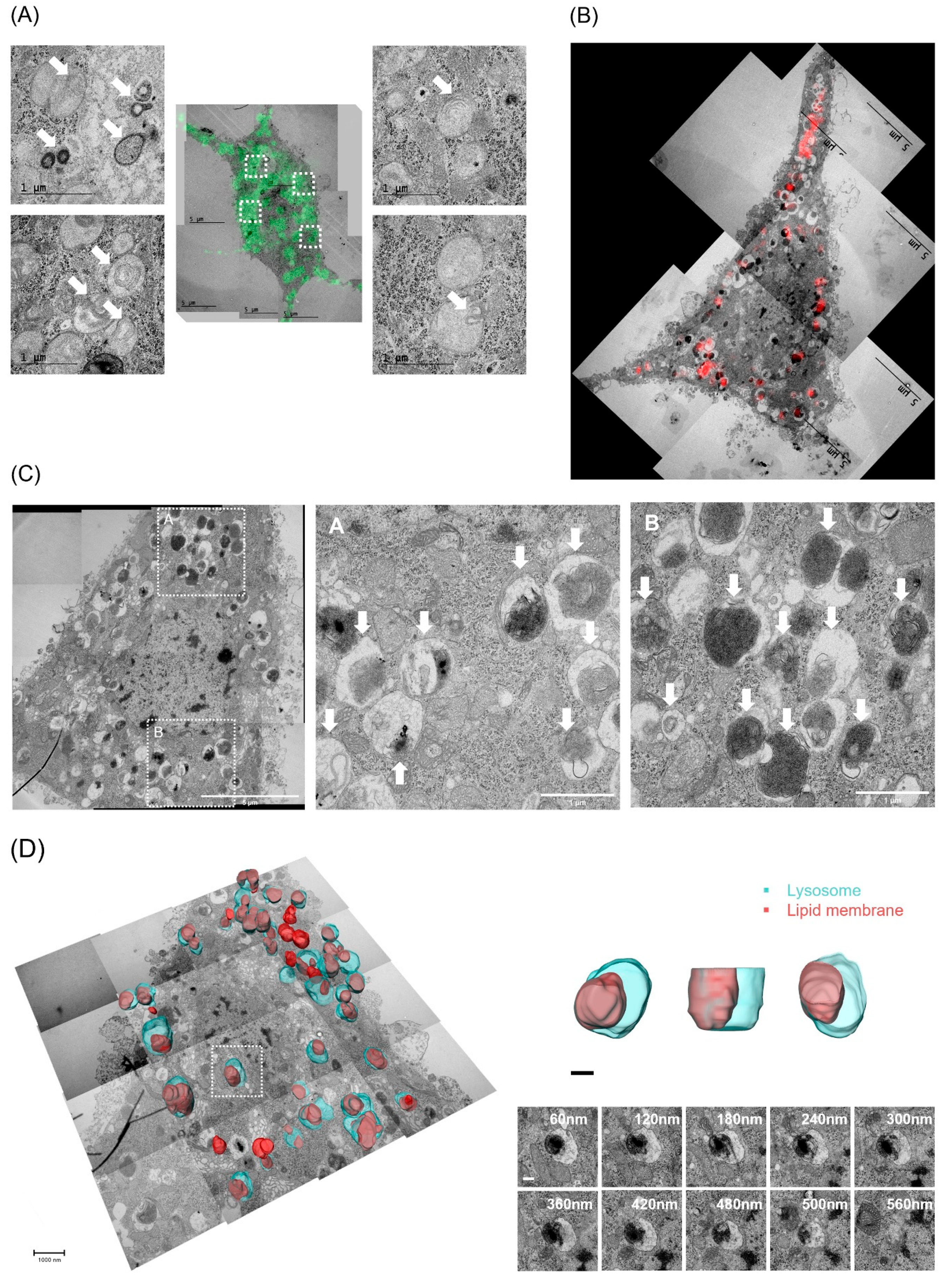
2.4. Three-Dimensional Volume Reconstruction in Serial TEM Showed Effects of L-Serine in Decreasing Lipid Droplets in PPA-Treated HT22 Cells
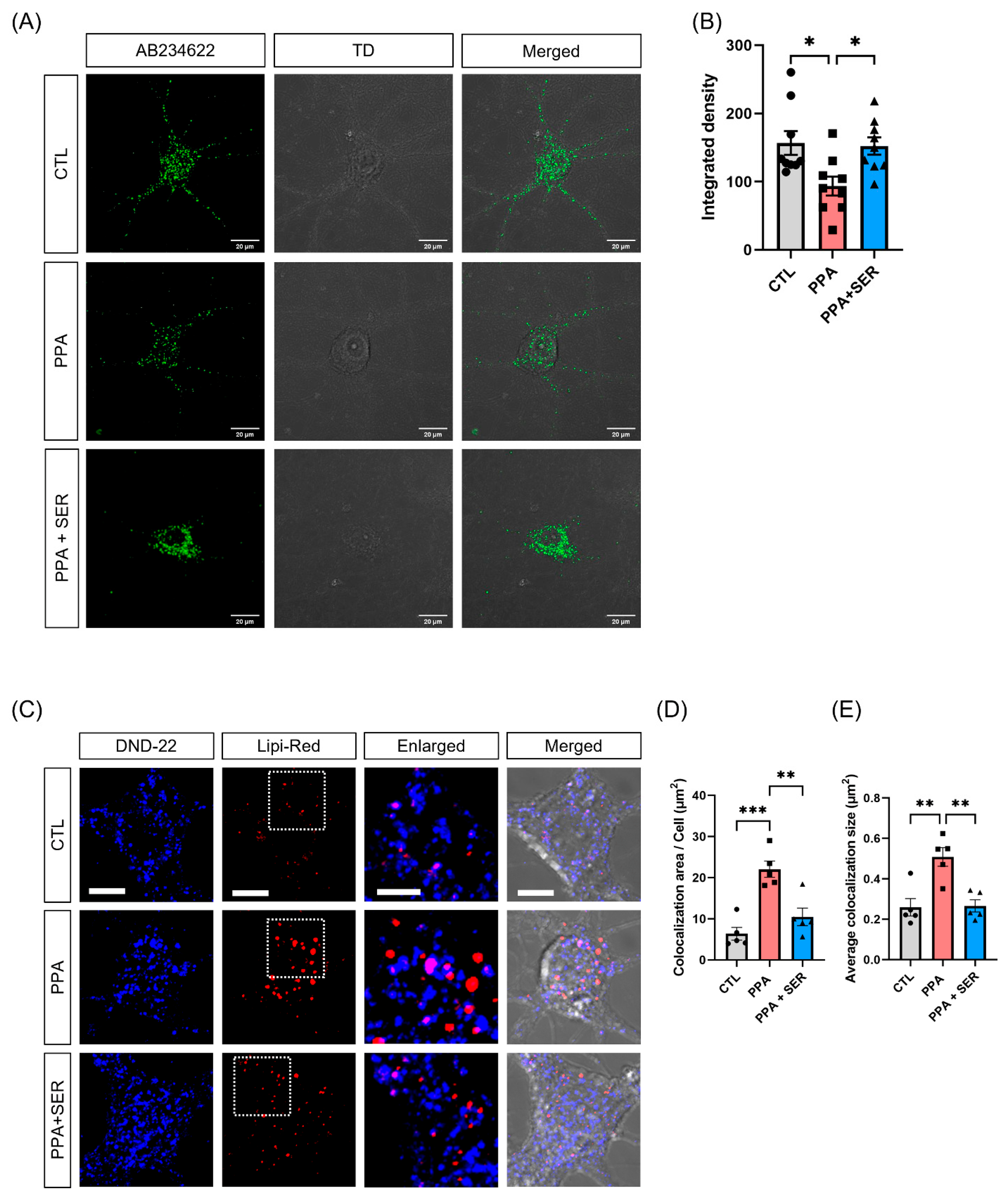
2.5. L-Serine Decreased the Colocalization of Lipi-Red and Lysotracker in PPA-Treated Hippocampal Neurons
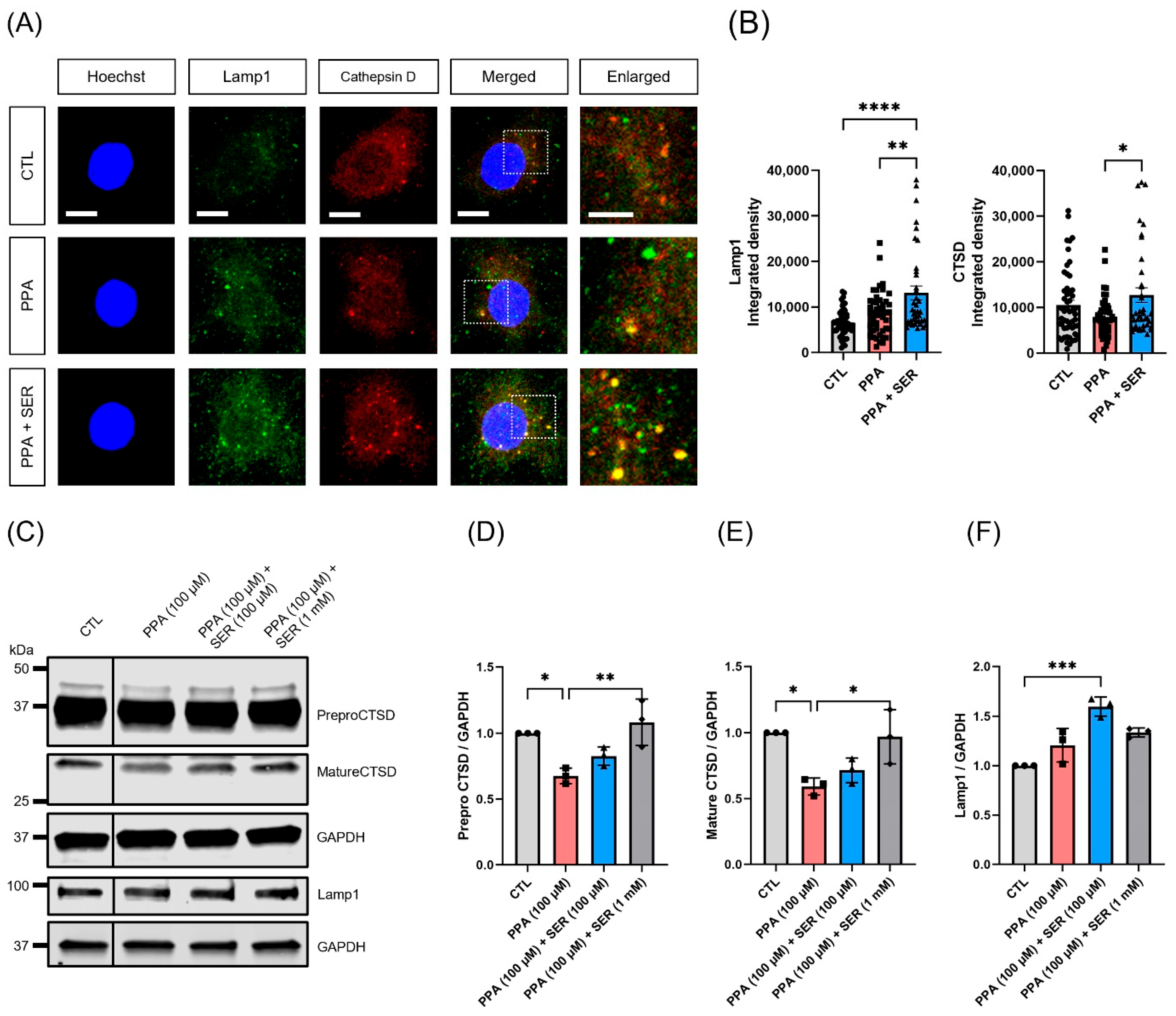
2.6. Attenuation of Lysosomal Dysfunction by L-Serine in PPA-Treated Hippocampal Neurons
2.7. CLEM and Serial TEM Showed Increase in Lysosomal Lipid Membrane Accumulation by PPA in Hippocampal Neurons
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Pharmacological Treatment
4.3. Cell Viability Assays
4.4. Trypan Blue Staining
4.5. Lysosomal Activity Assays
4.6. Immunocytochemistry
4.7. Confocal Microscopy Analysis
4.8. Western Blotting Analysis
4.9. Transmission Electron Microscopy (TEM)
4.10. Three-Dimensional Reconstruction of Serial TEM Images
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| PPA | propionic acid |
| SER | L-serine |
| CTSD | cathepsin D |
| LD | lipid droplet |
| SCFAs | short-chain fatty acids |
| TEM | transmission electron microscopy |
| CLEM | correlative light and electron microscopy |
| V-ATPase | vacuolar-type ATPase |
| AB234622 | lysosomal intracellular activity assay kit |
References
- Tabatabaie, L.; Klomp, L.W.; Berger, R.; de Koning, T.J. L-serine synthesis in the central nervous system: A review on serine deficiency disorders. Mol. Genet. Metab. 2010, 99, 256–262. [Google Scholar] [CrossRef]
- Metcalf, S.J.; Dunlop, R.A.; Powell, J.T.; Banack, S.A.; Cox, P.A. L-serine: A naturally-occurring amino acid with therapeutic potential. Neurotox. Res. 2018, 33, 213–221. [Google Scholar] [CrossRef]
- Meftah, A.; Hasegawa, H.; Kantrowitz, J.T. D-serine: A cross species review of safety. Front. Psychiatry 2021, 12, 726365. [Google Scholar] [CrossRef]
- Pernot, P.; Maucler, C.; Tholance, Y.; Vasylieva, N.; Debilly, G.; Pollegioni, L.; Cespuglio, R.; Marinesco, S. D-serine diffusion through the blood-brain barrier: Effect on d-serine compartmentalization and storage. Neurochem. Int. 2012, 60, 837–845. [Google Scholar] [CrossRef]
- Furuya, S.; Tabata, T.; Mitoma, J.; Yamada, K.; Yamasaki, M.; Makino, A.; Yamamoto, T.; Watanabe, M.; Kano, M.; Hirabayashi, Y. L-serine and glycine serve as major astroglia-derived trophic factors for cerebellar purkinje neurons. Proc. Natl. Acad. Sci. USA 2000, 97, 11528–11533. [Google Scholar] [CrossRef]
- De Koning, T.J.; Klomp, L.W. Serine-deficiency syndromes. Curr. Opin. Neurol. 2004, 17, 197–204. [Google Scholar] [CrossRef]
- Maugard, M.; Vigneron, P.-A.; Bolaños, J.P.; Bonvento, G. L-serine links metabolism with neurotransmission. Prog. Neurobiol. 2021, 197, 101896. [Google Scholar] [CrossRef]
- Ye, L.; Sun, Y.; Jiang, Z.; Wang, G. L-serine, an endogenous amino acid, is a potential neuroprotective agent for neurological disease and injury. Front. Mol. Neurosci. 2021, 14. [Google Scholar] [CrossRef]
- Sun, L.; Qiang, R.; Yang, Y.; Jiang, Z.-L.; Wang, G.-H.; Zhao, G.-W.; Ren, T.-J.; Jiang, R.; Xu, L.-H. L-serine treatment may improve neurorestoration of rats after permanent focal cerebral ischemia potentially through improvement of neurorepair. PLoS ONE 2014, 9, e93405. [Google Scholar] [CrossRef]
- Levine, D.T.; Miller, R.G.; Bradley, W.G.; Moore, D.H.; Saperstein, D.S.; Flynn, L.E.; Katz, J.S.; Forshew, D.A.; Metcalf, J.S.; Banack, S.A. Phase i clinical trial of safety of l-serine for als patients. Amyotroph. Lateral Scler. Front. Degener. 2017, 18, 107–111. [Google Scholar] [CrossRef]
- Douce, L.J.; Maugard, M.; Veran, J.; Matos, M.; Jégo, P.; Vigneron, P.A.; Faivre, E.; Toussay, X.; Vandenberghe, M.; Balbastre, Y. Impairment of glycolysis-derived l-serine production in astrocytes contributes to cognitive deficits in alzheimer’s disease. Cell Metab. 2020, 31, 503–517.e8. [Google Scholar] [CrossRef]
- Mitsui, R.; Ono, S.; Karaki, S.-I.; Kuwahara, A. Propionate modulates spontaneous contractions via enteric nerves and prostaglandin release in the rat distal colon. Jpn. J. Physiol. 2005, 55, 331–338. [Google Scholar] [CrossRef]
- Vadder, D.F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef]
- Sunkara, T.L.; Jiang, W.; Zhang, G. Modulation of antimicrobial host defense peptide gene expression by free fatty acids. PLoS ONE 2012, 7, e49558. [Google Scholar] [CrossRef]
- Grünert, C.S.; Müllerleile, S.; de Silva, L.; Barth, M.; Walter, M.; Walter, K.; Meissner, T.; Lindner, M.; Ensenauer, R.; Santer, R. Propionic acidemia: Clinical course and outcome in 55 pediatric and adolescent patients. Orphanet J. Rare Dis. 2013, 8, 6. [Google Scholar] [CrossRef]
- Morland, C.; Frøland, A.-S.; Pettersen, M.N.; Storm-Mathisen, J.; Gundersen, V.; Rise, F.; Hassel, B. Propionate enters gabaergic neurons, inhibits gaba transaminase, causes gaba accumulation and lethargy in a model of propionic acidemia. Biochem. J. 2018, 475, 749–758. [Google Scholar] [CrossRef]
- MacFabe, D.F. Short-chain fatty acid fermentation products of the gut microbiome: Implications in autism spectrum disorders. Microb. Ecol. Health Dis. 2012, 23, 19260. [Google Scholar] [CrossRef]
- Kim, A.S.; Jang, E.H.; Mun, J.Y.; Choi, H. Propionic acid induces mitochondrial dysfunction and affects gene expression for mitochondria biogenesis and neuronal differentiation in sh-sy5y cell line. Neurotoxicology 2019, 75, 116–122. [Google Scholar] [CrossRef]
- Zhou, H.; Yu, B.; Sun, J.; Liu, Z.; Chen, H.; Ge, L.; Chen, D. Short-chain fatty acids can improve lipid and glucose metabolism independently of the pig gut microbiota. J. Anim. Sci. Biotechnol. 2021, 12, 61. [Google Scholar] [CrossRef]
- Choi, H.; Kim, I.S.; Mun, J.Y. Propionic acid induces dendritic spine loss by mapk/erk signaling and dysregulation of autophagic flux. Mol. Brain 2020, 13, 86. [Google Scholar] [CrossRef]
- Thelen, M.A.; Zoncu, R. Emerging roles for the lysosome in lipid metabolism. Trends Cell Biol. 2017, 27, 833–850. [Google Scholar] [CrossRef]
- Yan, Y.; Jiang, K.; Liu, P.; Zhang, X.; Dong, X.; Gao, J.; Liu, Q.; Barr, M.P.; Zhang, Q.; Hou, X. Bafilomycin a1 induces caspase-independent cell death in hepatocellular carcinoma cells via targeting of autophagy and mapk pathways. Sci. Rep. 2016, 6, 37052. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Luo, J.; Zhu, J.; Shi, H.; Li, J.; Qiu, S.; Wang, P.; Loor, J.J. Effect of short-chain fatty acids on triacylglycerol accumulation, lipid droplet formation and lipogenic gene expression in goat mammary epithelial cells. Anim. Sci. J. 2016, 87, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Boer, d.P.; Hoogenboom, J.P.; Giepmans, B.N.G. Correlated light and electron microscopy: Ultrastructure lights up! Nat. Methods 2015, 12, 503–513. [Google Scholar] [CrossRef]
- Kitazawa, S.; Nishizawa, S.; Nakagawa, H.; Funata, M.; Nishimura, K.; Soga, T.; Hara, T. Cancer with low cathepsin d levels is susceptible to vacuolar (h(+) )-atpase inhibition. Cancer Sci. 2017, 108, 1185–1193. [Google Scholar] [CrossRef]
- Guerreiro, F.J.; Mira, N.P.; Santos, A.X.S.; Riezman, H.; Sá-Correia, I. Membrane phosphoproteomics of yeast early response to acetic acid: Role of hrk1 kinase and lipid biosynthetic pathways, in particular sphingolipids. Front. Microbiol. 2017, 8, 1302. [Google Scholar] [CrossRef]
- Godinho, P.C.; Prata, C.S.; Pinto, S.N.; Cardoso, C.; Bandarra, N.M.; Fernandes, F.; Sá-Correia, I. Pdr18 is involved in yeast response to acetic acid stress counteracting the decrease of plasma membrane ergosterol content and order. Sci. Rep. 2018, 8, 7860. [Google Scholar] [CrossRef]
- Guo, P.Z.; Khoomrung, S.; Nielsen, J.; Olsson, L. Changes in lipid metabolism convey acid tolerance in saccharomyces cerevisiae. Biotechnol. Biofuels 2018, 11, 297. [Google Scholar] [CrossRef]
- Lindberg, L.; Santos, A.X.S.; Riezman, H.; Olsson, L.; Bettiga, M. Lipidomic profiling of saccharomyces cerevisiae and zygosaccharomyces bailii reveals critical changes in lipid composition in response to acetic acid stress. PLoS ONE 2013, 8, e73936. [Google Scholar] [CrossRef]
- Hong, H.Y.; Nishimura, Y.; Hishikawa, D.; Tsuzuki, H.; Miyahara, H.; Gotoh, C.; Choi, K.C.; Feng, D.D.; Chen, C.; Lee, H.G.; et al. Acetate and propionate short chain fatty acids stimulate adipogenesis via gpcr43. Endocrinology 2005, 146, 5092–5099. [Google Scholar] [CrossRef] [Green Version]
- Yonezawa, T.; Yonekura, S.; Sanosaka, M.; Hagino, A.; Katoh, K.; Obara, Y. Octanoate stimulates cytosolic triacylglycerol accumulation and cd36 mrna expression but inhibits acetyl coenzyme a carboxylase activity in primary cultured bovine mammary epithelial cells. J Dairy Res 2004, 71, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Araujo, G.M.E.; Liebscher, G.; Hess, M.W.; Huber, L.A. Lysosomal size matters. Traffic 2020, 21, 60–75. [Google Scholar] [CrossRef] [PubMed]
- Heuser, J. Changes in lysosome shape and distribution correlated with changes in cytoplasmic ph. J. Cell Biol. 1989, 108, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Gottschling, E.D.; Nyström, T. The upsides and downsides of organelle interconnectivity. Cell 2017, 169, 24–34. [Google Scholar] [CrossRef]
- Halcrow, P.; Khan, N.; Datta, G.; Ohm, J.E.; Chen, X.; Geiger, J.D. Importance of measuring endolysosome, cytosolic, and extracellular ph in understanding the pathogenesis of and possible treatments for glioblastoma multiforme. Cancer Rep. 2019, 2, e1193. [Google Scholar] [CrossRef]
- Lakpa, L.K.; Khan, N.; Afghah, Z.; Chen, X.; Geiger, J.D. Lysosomal stress response (lsr): Physiological importance and pathological relevance. J. Neuroimmune Pharmacol. 2021, 16, 219–237. [Google Scholar] [CrossRef]
- Song, Q.; Meng, B.; Xu, H.; Mao, Z. The emerging roles of vacuolar-type atpase-dependent lysosomal acidification in neurodegenerative diseases. Transl. Neurodegener. 2020, 9, 17. [Google Scholar] [CrossRef]
- Zoncu, R.; Bar-Peled, L.; Efeyan, A.; Wang, S.; Sancak, Y.; Sabatini, D.M. Mtorc1 senses lysosomal amino acids through an inside-out mechanism that requires the vacuolar h(+)-atpase. Science 2011, 334, 678–683. [Google Scholar] [CrossRef]
- Stransky, A.L.; Forgac, M. Amino acid availability modulates vacuolar h+-atpase assembly. J. Biol. Chem. 2015, 290, 27360–27369. [Google Scholar] [CrossRef]
- Muench, P.S.; Rawson, S.; Eyraud, V.; Delmas, A.F.; da Silva, P.; Phillips, C.; Trinick, J.; Harrison, M.A.; Gressent, F.; Huss, M. Pa1b inhibitor binding to subunits c and e of the vacuolar atpase reveals its insecticidal mechanism. J. Biol. Chem. 2014, 289, 16399–16408. [Google Scholar] [CrossRef] [Green Version]
- Hanessian, S.; Ma, J.; Wang, W.; Gai, Y. Total synthesis of bafilomycin a(1) relying on iterative 1,2-induction in acyclic precursors. J. Am. Chem. Soc. 2001, 123, 10200–10206. [Google Scholar] [CrossRef] [PubMed]
- Huss, M.; Wieczorek, H. Inhibitors of v-atpases: Old and new players. J. Exp. Biol. 2009, 212, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Cullen, V.; Lindfors, M.; Ng, J.; Paetau, A.; Swinton, E.; Kolodziej, P.; Boston, H.; Saftig, P.; Woulfe, J.; Feany, M.B.; et al. Cathepsin d expression level affects alpha-synuclein processing, aggregation, and toxicity in vivo. Mol. Brain 2009, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Hamamichi, S.; Caldwell, K.A.; Caldwell, G.A.; Yacoubian, T.A.; Wilson, S.; Xie, Z.-L.; Speake, L.D.; Parks, R.; Crabtree, D. Lysosomal enzyme cathepsin d protects against alpha-synuclein aggregation and toxicity. Mol. Brain 2008, 1, 17. [Google Scholar] [CrossRef] [PubMed]
- Sevlever, D.; Jiang, P.; Yen, S.H. Cathepsin d is the main lysosomal enzyme involved in the degradation of alpha-synuclein and generation of its carboxy-terminally truncated species. Biochemistry 2008, 47, 9678–9687. [Google Scholar] [CrossRef]
- Koike, M.; Nakanishi, H.; Saftig, P.; Ezaki, J.; Isahara, K.; Ohsawa, Y.; Schulz-Schaeffer, W.; Watanabe, T.; Waguri, S.; Kametaka, S. Cathepsin d deficiency induces lysosomal storage with ceroid lipofuscin in mouse cns neurons. J. Neurosci. 2000, 20, 6898–6906. [Google Scholar] [CrossRef]
- Dunlop, A.R.; Carney, J.M. Mechanisms of l-serine-mediated neuroprotection include selective activation of lysosomal cathepsins b and l. Neurotox. Res. 2021, 39, 17–26. [Google Scholar] [CrossRef]
- Stoka, V.; Turk, V.; Turk, B. Lysosomal cathepsins and their regulation in aging and neurodegeneration. Ageing Res. Rev. 2016, 32, 22–37. [Google Scholar] [CrossRef]
- Houben, T.; Oligschlaeger, Y.; Hendrikx, T.; Bitorina, A.V.; Walenbergh, S.M.A.; van Gorp, P.J.; Gijbels, M.J.J.; Friedrichs, S.; Plat, J.; Schaap, F.G. Cathepsin d regulates lipid metabolism in murine steatohepatitis. Sci. Rep. 2017, 7, 3494. [Google Scholar] [CrossRef]
- Haidar, B.; Kiss, R.S.; Sarov-Blat, L.; Brunet, R.; Harder, C.; McPherson, R.; Marcel, Y.L. Cathepsin d, a lysosomal protease, regulates abca1-mediated lipid efflux*. J. Biol. Chem. 2006, 281, 39971–39981. [Google Scholar] [CrossRef] [Green Version]
- Lorkowski, W.S.; Brubaker, G.; Gulshan, K.; Smith, J.D. V-atpase (vacuolar atpase) activity required for abca1 (atp-binding cassette protein a1)-mediated cholesterol efflux. Arterioscler Thromb. Vasc. Biol. 2018, 38, 2615–2625. [Google Scholar] [CrossRef] [PubMed]
- Jang, N.Y.; Jang, H.; Kim, G.H.; Noh, J.E.; Chang, K.A.; Lee, K.J. Rapgef2 mediates oligomeric aβ-induced synaptic loss and cognitive dysfunction in the 3xtg-ad mouse model of alzheimer’s disease. Neuropathol. Appl. Neurobiol. 2021, 47, 625–639. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeon, H.; Kim, Y.J.; Hwang, S.-K.; Seo, J.; Mun, J.Y. Restoration of Cathepsin D Level via L-Serine Attenuates PPA-Induced Lysosomal Dysfunction in Neuronal Cells. Int. J. Mol. Sci. 2022, 23, 10613. https://doi.org/10.3390/ijms231810613
Jeon H, Kim YJ, Hwang S-K, Seo J, Mun JY. Restoration of Cathepsin D Level via L-Serine Attenuates PPA-Induced Lysosomal Dysfunction in Neuronal Cells. International Journal of Molecular Sciences. 2022; 23(18):10613. https://doi.org/10.3390/ijms231810613
Chicago/Turabian StyleJeon, Hyunbum, Yeo Jin Kim, Su-Kyeong Hwang, Jinsoo Seo, and Ji Young Mun. 2022. "Restoration of Cathepsin D Level via L-Serine Attenuates PPA-Induced Lysosomal Dysfunction in Neuronal Cells" International Journal of Molecular Sciences 23, no. 18: 10613. https://doi.org/10.3390/ijms231810613
APA StyleJeon, H., Kim, Y. J., Hwang, S. -K., Seo, J., & Mun, J. Y. (2022). Restoration of Cathepsin D Level via L-Serine Attenuates PPA-Induced Lysosomal Dysfunction in Neuronal Cells. International Journal of Molecular Sciences, 23(18), 10613. https://doi.org/10.3390/ijms231810613