
An Axis between the Long Non-Coding RNA HOXA11-AS and NQOs Enhances Metastatic Ability in Oral Squamous Cell Carcinoma

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Expression of HOXA11-AS and NQOs in Human OSCCs
2.2. Role of HOXA11-AS in NQO1 Activation in Human OSCC Cell Lines
2.3. Role of HOXA11-AS in NQO2 Repression in Human OSCC Cell Line
2.4. Effect of HOXA11-AS on Malignant Phenotypes of OSCC Cells
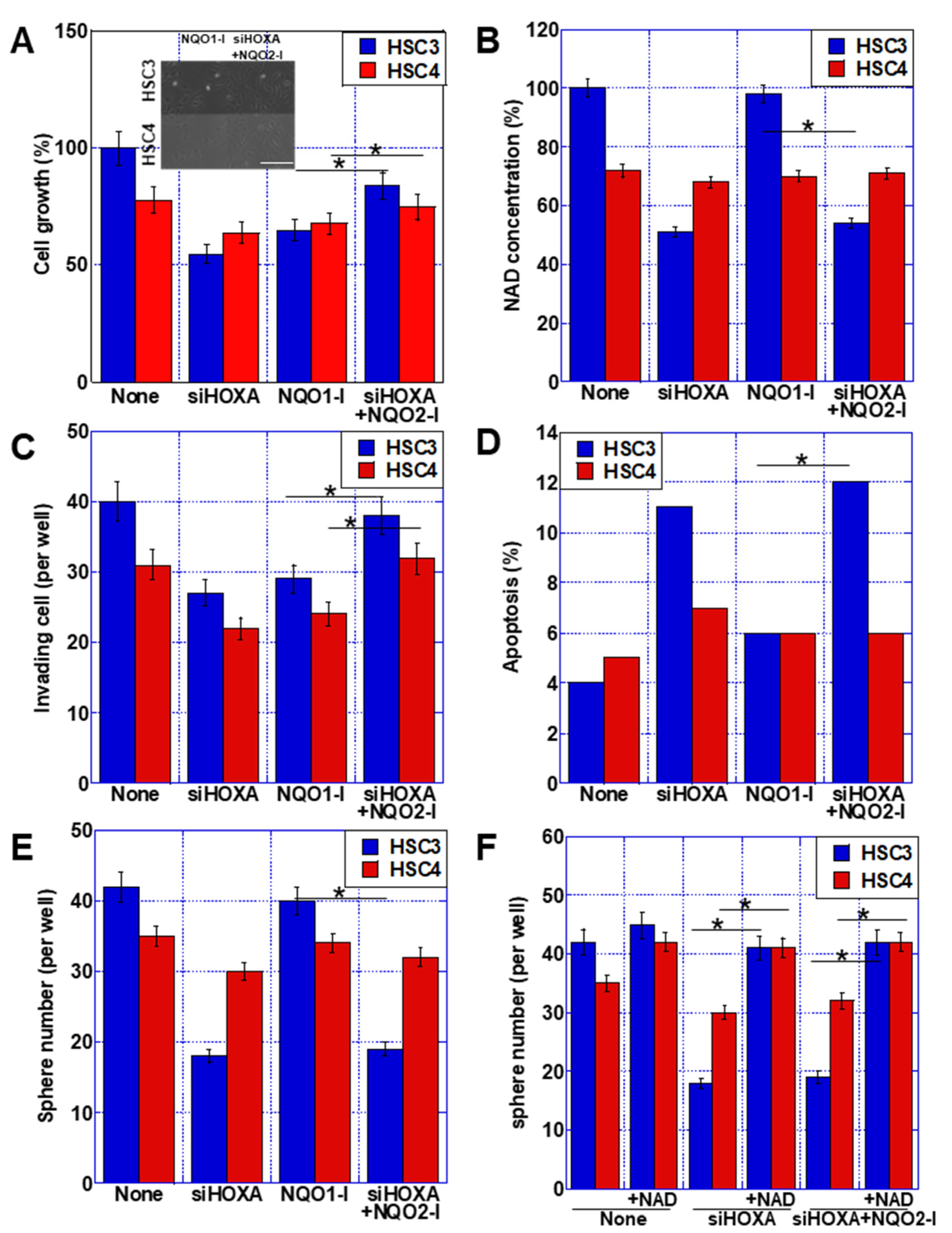
2.5. Role of NQO1 in OSCC Cell Lines
2.6. Role of NQO2 in OSCC Cell Lines
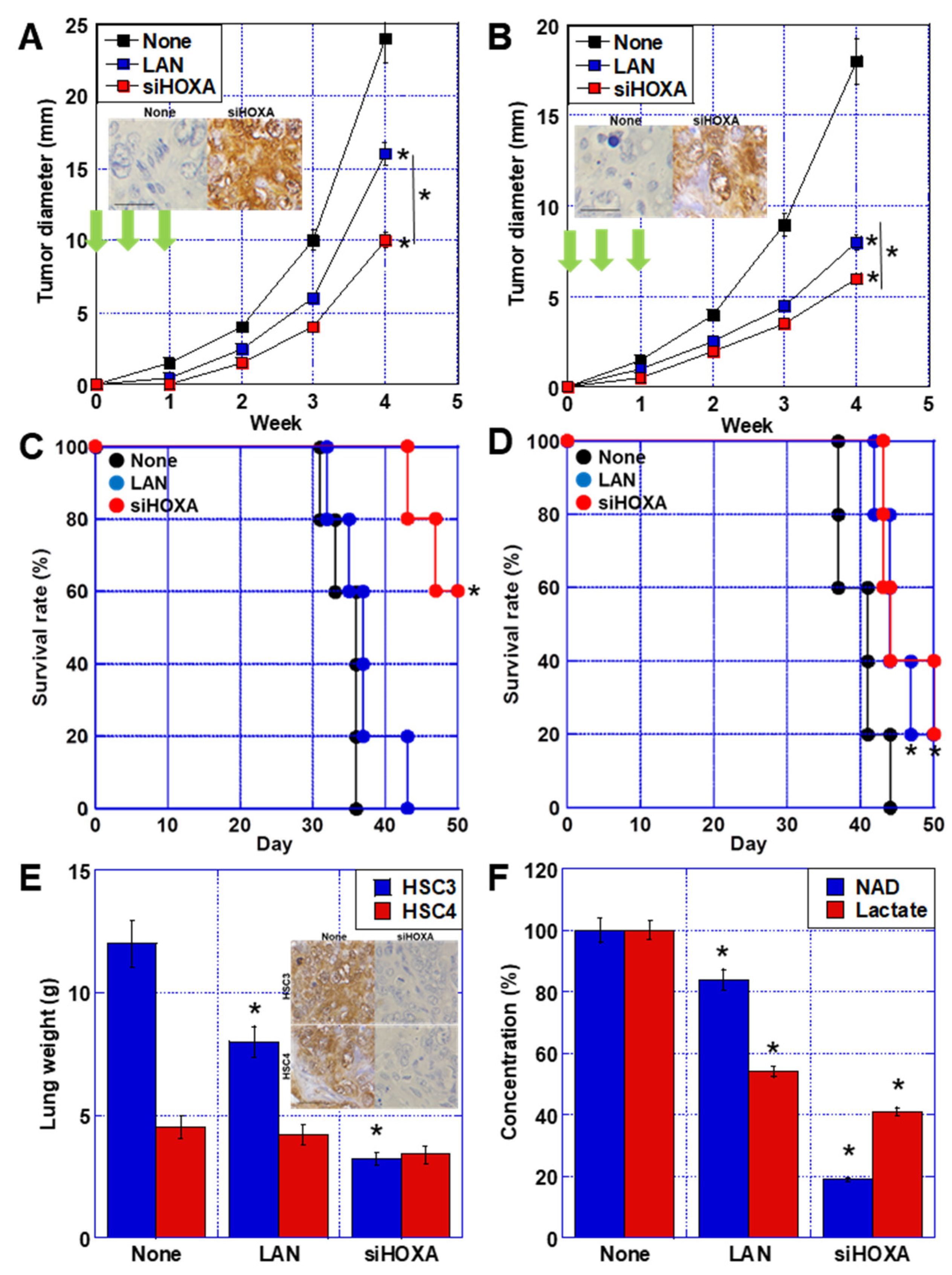
2.7. Effect of HOXA11-AS Treatment of OSCC
3. Discussion
4. Materials and Methods
4.1. Surgical Specimens
4.2. Cell Culture
4.3. Reagents
4.4. Cell Proliferation, Cell Infiltration, and Apoptosis
4.5. Sphere Formation Assay
4.6. Small Interfering RNA
4.7. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.8. qRT-PCR for lncRNA
4.9. Detection of miRNA
4.10. Protein Extraction
4.11. Determination of Lactate, NAD, and FAD Concentrations, and Activity of NQO1 and GAPDH
4.12. Chromatin Immunoprecipitation Assay
4.13. Animal Models
4.14. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviation
| lncRNA | Long non-coding RNA |
| HOX | homeobox |
| HOXA11-AS | HOX-A11 antisense |
| OSCC | oral squamous cell carcinoma |
| NQO1 | NAD(P)H: quinone oxidoreductase 1 |
| NQO2 | NRH: quinone oxidoreductase 2 |
| EZH2 | enhancer of zeste homolog 2 |
| H3K27me3 | trimethylation of histone H3 at lysine 4 |
| CDDP | cisplatin |
| LAN | lanthanide |
| ME | malic enzyme |
| FAD | flavin adenine dinucleotide |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| NAD | nicotinamide adenine dinucleotide |
References
- Kocarnik, J.M.; Compton, K.; Dean, F.E.; Fu, W.; Gaw, B.L.; Harvey, J.D.; Henrikson, H.J.; Lu, D.; Pennini, A.; Xu, R.; et al. Cancer Incidence, Mortality, Years of Life Lost, Years Lived with Disability, and Disability-Adjusted Life Years for 29 Cancer Groups From 2010 to 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. JAMA Oncol. 2022, 8, 420–444. [Google Scholar] [PubMed]
- Stokes, W.A.; Fuller, C.; Day, T.A.; Gillespie, M.B. Perioperative survival of elderly head and neck squamous cell carcinoma patients. Laryngoscope 2014, 124, 2281–2286. [Google Scholar] [CrossRef] [PubMed]
- Prince, A.; Aguirre-Ghizo, J.; Genden, E.; Posner, M.; Sikora, A. Head and neck squamous cell carcinoma: New translational therapies. Mt. Sinai J. Med. 2010, 77, 684–699. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, N.; Sento, S.; Yoshizawa, Y.; Sasabe, E.; Kudo, Y.; Yamamoto, T. Current Trends and Future Prospects of Molecular Targeted Therapy in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2020, 22, 240. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, Y.; Song, G.; Zhang, X.; Gao, S.; Liu, H. HOX cluster-embedded antisense long non-coding RNAs in lung cancer. Cancer Lett. 2019, 450, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.Y.; Huang, C.; Wang, W.; Li, H.B.; Sun, M.; Xie, M. HOXA11-AS: A novel regulator in human cancer proliferation and metastasis. OncoTargets Ther. 2018, 11, 4387–4393. [Google Scholar] [CrossRef]
- Chen, D.; Sun, Q.; Cheng, X.; Zhang, L.; Song, W.; Zhou, D.; Lin, J.; Wang, W. Genome-wide analysis of long noncoding RNA (lncRNA) expression in colorectal cancer tissues from patients with liver metastasis. Cancer Med. 2016, 5, 1629–1639. [Google Scholar] [CrossRef]
- Wei, C.; Zhao, L.; Liang, H.; Zhen, Y.; Han, L. Recent advances in unraveling the molecular mechanisms and functions of HOXA11-AS in human cancers and other diseases (Review). Oncol. Rep. 2020, 43, 1737–1754. [Google Scholar] [CrossRef]
- Lu, S.; Jiang, X.; Su, Z.; Cui, Z.; Fu, W.; Tai, S. The role of the long non-coding RNA HOXA11-AS in promoting proliferation and metastasis of malignant tumors. Cell Biol. Int. 2018, 42, 1596–1601. [Google Scholar] [CrossRef]
- Long, D.J., 2nd; Jaiswal, A.K. NRH:quinone oxidoreductase2 (NQO2). Chem.-Biol. Interact. 2000, 129, 99–112. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, D.; Ma, K.; Wu, X.; Hao, H.; Jiang, S. NAD(P)H:Quinone Oxidoreductase 1 (NQO1) as a Therapeutic and Diagnostic Target in Cancer. J. Med. Chem. 2018, 61, 6983–7003. [Google Scholar] [CrossRef] [PubMed]
- Hirose, Y.; Sakata, J.; Kobayashi, T.; Miura, K.; Yuza, K.; Nakano, M.; Ichikawa, H.; Nagahashi, M.; Shimada, Y.; Kameyama, H.; et al. NQO1 as a Marker of Chemosensitivity and Prognosis for Colorectal Liver Metastasis. Anticancer Res. 2021, 41, 1563–1570. [Google Scholar] [CrossRef]
- Sobin, L.H.; Gospodarowicz, M.; Wittekind, C. UICC TNM Classification of Malignant Tumours, 7th ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2009. [Google Scholar]
- Zhao, X.; Jin, Y.; Li, L.; Xu, L.; Tang, Z.; Qi, Y.; Yin, L.; Peng, J. MicroRNA-128-3p aggravates doxorubicin-induced liver injury by promoting oxidative stress via targeting Sirtuin-1. Pharmacol. Res. 2019, 146, 104276. [Google Scholar] [CrossRef] [PubMed]
- Jadeja, R.N.; Jones, M.A.; Abdelrahman, A.A.; Powell, F.L.; Thounaojam, M.C.; Gutsaeva, D.; Bartoli, M.; Martin, P.M. Inhibiting microRNA-144 potentiates Nrf2-dependent antioxidant signaling in RPE and protects against oxidative stress-induced outer retinal degeneration. Redox Biol. 2020, 28, 101336. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Tao, X.; Qi, Y.; Xu, L.; Yin, L.; Peng, J. Protective effect of dioscin against doxorubicin-induced cardiotoxicity via adjusting microRNA-140-5p-mediated myocardial oxidative stress. Redox Biol. 2018, 16, 189–198. [Google Scholar] [CrossRef]
- Li, Y.; Gao, M.; Yin, L.H.; Xu, L.N.; Qi, Y.; Sun, P.; Peng, J.Y. Dioscin ameliorates methotrexate-induced liver and kidney damages via adjusting miRNA-145-5p-mediated oxidative stress. Free Radic. Biol. Med. 2021, 169, 99–109. [Google Scholar] [CrossRef]
- Yang, J.J.; Tao, H.; Hu, W.; Liu, L.P.; Shi, K.H.; Deng, Z.Y.; Li, J. MicroRNA-200a controls Nrf2 activation by target Keap1 in hepatic stellate cell proliferation and fibrosis. Cell. Signal. 2014, 26, 2381–2389. [Google Scholar] [CrossRef]
- Huang, W.; Shi, G.; Yong, Z.; Li, J.; Qiu, J.; Cao, Y.; Zhao, Y.; Yuan, L. Downregulation of RKIP promotes radioresistance of nasopharyngeal carcinoma by activating NRF2/NQO1 axis via downregulating miR-450b-5p. Cell Death Dis. 2020, 11, 504. [Google Scholar] [CrossRef]
- Ling, Y.; Li, Z.Z.; Zhang, J.F.; Zheng, X.W.; Lei, Z.Q.; Chen, R.Y.; Feng, J.H. MicroRNA-494 inhibition alleviates acute lung injury through Nrf2 signaling pathway via NQO1 in sepsis-associated acute respiratory distress syndrome. Life Sci. 2018, 210, 1–8. [Google Scholar] [CrossRef]
- Wu, G.D.; Li, Z.H.; Li, X.; Zheng, T.; Zhang, D.K. microRNA-592 blockade inhibits oxidative stress injury in Alzheimer’s disease astrocytes via the KIAA0319-mediated Keap1/Nrf2/ARE signaling pathway. Exp. Neurol. 2020, 324, 113128. [Google Scholar] [CrossRef]
- Liang, J.Q.; Zhou, Z.T.; Bo, L.; Tan, H.N.; Hu, J.H.; Tan, M.S. Phosphoglycerate kinase 1 silencing by a novel microRNA microRNA-4523 protects human osteoblasts from dexamethasone through activation of Nrf2 signaling cascade. Cell Death Dis. 2021, 12, 964. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, C.; Yamamoto, K.; Fujiwara-Tani, R.; Luo, Y.; Matsushima, S.; Fujii, K.; Ohmori, H.; Sasahira, T.; Sasaki, T.; Kitadai, Y.; et al. Expression of cytosolic malic enzyme (ME1) is associated with disease progression in human oral squamous cell carcinoma. Cancer Sci. 2018, 109, 2036–2045. [Google Scholar] [CrossRef] [PubMed]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.J.; Lin, X.D.; Xu, L.Y.; Zhu, S.Q. Long Noncoding RNA HOXA11-AS Modulates the Resistance of Nasopharyngeal Carcinoma Cells to Cisplatin via miR-454-3p/c-Met. Mol. Cells 2020, 43, 856–869. [Google Scholar] [PubMed]
- Wang, J.; Shen, J. LncRNA HOXA11-AS aggravates the keloid formation by targeting miR-148b-3p/IGFBP5 axis. Biochem. Biophys. Res. Commun. 2021, 581, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Lv, H.; Yang, Z.; Shu, J.; Zhang, H. LncRNA PCAT29 Up-Regulates the Expression of PTEN by Down-Regulating miR-494 in Non-Small-Cell Lung Cancer to Suppress Tumor Progression. Crit. Rev. Eukaryot. Gene Expr. 2021, 31, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.W.; Liu, A.C. LncRNA SBF2-AS1 Promotes Diffuse Large B-Cell Lymphoma Growth by Regulating FGFR2 via Sponging miR-494-3p. Cancer Manag. Res. 2021, 13, 571–578. [Google Scholar] [CrossRef]
- Ogata, R.; Mori, S.; Kishi, S.; Sasaki, R.; Iwata, N.; Ohmori, H.; Sasaki, T.; Nishiguchi, Y.; Nakashima, C.; Goto, K.; et al. Linoleic Acid Upregulates Microrna-494 to Induce Quiescence in Colorectal Cancer. Int. J. Mol. Sci. 2021, 23, 225. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Q.; Li, L. Functional and therapeutic significance of EZH2 in urological cancers. Oncotarget 2017, 8, 38044–38055. [Google Scholar] [CrossRef]
- Jenuwein, T.; Allis, C.D. Translating the histone code. Science 2001, 293, 1074–1080. [Google Scholar] [CrossRef] [Green Version]
- Bainomugisa, C.K.; Sutherland, H.G.; Parker, R.; McRae, A.F.; Haupt, L.M.; Griffiths, L.R.; Heath, A.; Nelson, E.C.; Wright, M.J.; Hickie, I.B.; et al. Using Monozygotic Twins to Dissect Common Genes in Posttraumatic Stress Disorder and Migraine. Front. Neurosci. 2021, 15, 678350. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Venneti, S.; Nagrath, D. Glutaminolysis: A Hallmark of Cancer Metabolism. Ann. Rev. Biomed. Eng. 2017, 19, 163–194. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, C.; Kirita, T.; Yamamoto, K.; Mori, S.; Luo, Y.; Sasaki, T.; Fujii, K.; Ohmori, H.; Kawahara, I.; Mori, T.; et al. Malic Enzyme 1 Is Associated with Tumor Budding in Oral Squamous Cell Carcinomas. Int. J. Mol. Sci. 2020, 21, 7149. [Google Scholar] [CrossRef] [PubMed]
- Provenzani, A.; Fronza, R.; Loreni, F.; Pascale, A.; Amadio, M.; Quattrone, A. Global alterations in mRNA polysomal recruitment in a cell model of colorectal cancer progression to metastasis. Carcinogenesis 2006, 27, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Long, D.J., 2nd; Iskander, K.; Gaikwad, A.; Arin, M.; Roop, D.R.; Knox, R.; Barrios, R.; Jaiswal, A.K. Disruption of dihydronicotinamide riboside:quinone oxidoreductase 2 (NQO2) leads to myeloid hyperplasia of bone marrow and decreased sensitivity to menadione toxicity. J. Biol. Chem. 2002, 277, 46131–46139. [Google Scholar] [CrossRef]
- Chini, C.C.S.; Peclat, T.R.; Gomez, L.S.; Zeidler, J.D.; Warner, G.M.; Kashyap, S.; Mazdeh, D.Z.; Hayat, F.; Migaud, M.E.; Paulus, A.; et al. Dihydronicotinamide Riboside Is a Potent NAD(+) Precursor Promoting a Pro-Inflammatory Phenotype in Macrophages. Front. Immunol. 2022, 13, 840246. [Google Scholar] [CrossRef] [PubMed]
- Ciarlo, E.; Joffraud, M.; Hayat, F.; Giner, M.P.; Giroud-Gerbetant, J.; Sanchez-Garcia, J.L.; Rumpler, M.; Moco, S.; Migaud, M.E.; Cantó, C. Nicotinamide Riboside and Dihydronicotinic Acid Riboside Synergistically Increase Intracellular NAD(+) by Generating Dihydronicotinamide Riboside. Nutrients 2022, 14, 2752. [Google Scholar] [CrossRef]
- Yang, Y.; Mohammed, F.S.; Zhang, N.; Sauve, A.A. Dihydronicotinamide riboside is a potent NAD(+) concentration enhancer in vitro and in vivo. J. Biol. Chem. 2019, 294, 9295–9307. [Google Scholar] [CrossRef]
- Navas, L.E.; Carnero, A. NAD(+) metabolism, stemness, the immune response, and cancer. Signal Transduct. Target. Ther. 2021, 6, 2. [Google Scholar] [CrossRef]
- Lucena-Cacace, A.; Otero-Albiol, D.; Jiménez-García, M.P.; Peinado-Serrano, J.; Carnero, A. NAMPT overexpression induces cancer stemness and defines a novel tumor signature for glioma prognosis. Oncotarget 2017, 8, 99514–99530. [Google Scholar] [CrossRef] [Green Version]
- Heske, C.M. Beyond Energy Metabolism: Exploiting the Additional Roles of NAMPT for Cancer Therapy. Front. Oncol. 2019, 9, 1514. [Google Scholar] [CrossRef] [PubMed]
- Momtazmanesh, S.; Rezaei, N. Long Non-Coding RNAs in Diagnosis, Treatment, Prognosis, and Progression of Glioma: A State-of-the-Art Review. Front. Oncol. 2021, 11, 712786. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.J.; You, G.R.; Chang, J.T.; Cheng, A.J. Systematic Analysis and Identification of Dysregulated Panel lncRNAs Contributing to Poor Prognosis in Head-Neck Cancer. Front. Oncol. 2021, 11, 731752. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Mori, S.; Kishi, S.; Honoki, K.; Fujiwara-Tani, R.; Moriguchi, T.; Sasaki, T.; Fujii, K.; Tsukamoto, S.; Fujii, H.; Kido, A.; et al. Anti-Stem Cell Property of Pterostilbene in Gastrointestinal Cancer Cells. Int. J. Mol. Sci. 2020, 21, 9347. [Google Scholar] [CrossRef] [PubMed]
- Kuniyasu, H.; Oue, N.; Wakikawa, A.; Shigeishi, H.; Matsutani, N.; Kuraoka, K.; Ito, R.; Yokozaki, H.; Yasui, W. Expression of receptors for advanced glycation end-products (RAGE) is closely associated with the invasive and metastatic activity of gastric cancer. J. Pathol. 2002, 196, 163–170. [Google Scholar] [CrossRef]
- Kuniyasu, H.; Oue, N.; Sasahira, T.; Luo, Y.; Moriwaka, Y.; Shimomoto, T.; Fujii, K.; Ohmori, H.; Yasui, W. Reg IV enhances peritoneal metastasis in gastric carcinomas. Cell Prolif. 2009, 42, 110–121. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Forward (5′–3′) | Reference | |
|---|---|---|---|---|
| Promoter DNA | ||||
| NQO2 promoter | AY334547.1 | Upper | TGGCATCTCACAAAGGACAG | |
| Lower | GCCGCTGGTGTACTGGTATT | |||
| RNA | ||||
| nucleostemin (NS) | BC001024.2 | Upper | ATTGCCAACAGTGGTGTTCA | |
| Lower | AATGGCTTTGCTGCAAGTTT | |||
| CD44 | FJ216964.1 | Upper | CATTCAAATCCGGAAGTGCT | |
| Lower | GTTGCCAAACCACTGTTCCT | |||
| NQO1 | BC007659.2 | Upper | AAAGGACCCTTCCGGAGTAA | |
| Lower | CCATCCTTCCAGGATTTGAA | |||
| NQO2 | BC006096.2 | Upper | CACACCAGGAACCCAAGTCT | |
| Lower | TTGTAGGCTTCGTGGGTTTC | |||
| ACTB | NM_001101.3 | Upper | GGACTTCGAGCAAGAGATGG | |
| Lower | AGCACTGTGTTGGCGTACAG | |||
| lncRNA | ||||
| HOXA11AS | NR_002795.2 | Upper | TCTCCTGGAGTCTCGCATTT | |
| Lower | TCGGAAGTGACCATGAATGA | |||
| miRNA | ||||
| mir-128 | NR_029672.1 | Upper | GCCGTAGCACTGTCTGAGAG | [14] |
| Lower | GCAGCTGAAAAAGAGACCGG | |||
| mir-140 | NR_029681.1 | Upper | TGTGTCCTGCCAGTGGTTTT | [15] |
| Lower | GTCCGTGGTTCTACCCTGTG | |||
| mir-144 | NR_029685.1 | Upper | AGTTTGCGATGAGACACTACAGT | [16] |
| Lower | GGTGCCCGGACTAGTACATC | |||
| mir-145 | NR_029686.1 | Upper | CTTGTCCTCACGGTCCAGTT | [17] |
| Lower | TTCCTGGGAAAACTGGACCG | |||
| mir-200a | NR_029834.1 | Upper | AGCATCTTACCGGACAGTGC | [18] |
| Lower | TGGGAAATCCAGCACTGTCC | |||
| mir-450b | LM609945.1 | Upper | AAGTGTATTGGGATCATTTTGCA | [19] |
| Lower | ACTATGGATGCAAAATGATCCCA | |||
| mir-494 | NR_030174.1 | Upper | CTCGAAGGAGAGGTTGTCCG | [20] |
| Lower | AGAAGACAACACGGACAACCT | |||
| mir-592 | NR_030323.1 | Upper | GCGATGATGTGTTGTGATGGC | [21] |
| Lower | CGTCATGATGTTGCGTCACC | |||
| mir-4523 | NR_039749.1 | Upper | TCGGCTGTGTGAGGACTAGA | [22] |
| Lower | CTCGGCCGCCTCTAGTCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakashima, C.; Fujiwara-Tani, R.; Mori, S.; Kishi, S.; Ohmori, H.; Fujii, K.; Mori, T.; Miyagawa, Y.; Yamamoto, K.; Kirita, T.; et al. An Axis between the Long Non-Coding RNA HOXA11-AS and NQOs Enhances Metastatic Ability in Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2022, 23, 10704. https://doi.org/10.3390/ijms231810704
Nakashima C, Fujiwara-Tani R, Mori S, Kishi S, Ohmori H, Fujii K, Mori T, Miyagawa Y, Yamamoto K, Kirita T, et al. An Axis between the Long Non-Coding RNA HOXA11-AS and NQOs Enhances Metastatic Ability in Oral Squamous Cell Carcinoma. International Journal of Molecular Sciences. 2022; 23(18):10704. https://doi.org/10.3390/ijms231810704
Chicago/Turabian StyleNakashima, Chie, Rina Fujiwara-Tani, Shiori Mori, Shingo Kishi, Hitoshi Ohmori, Kiyomu Fujii, Takuya Mori, Yoshihiro Miyagawa, Kazuhiko Yamamoto, Tadaaki Kirita, and et al. 2022. "An Axis between the Long Non-Coding RNA HOXA11-AS and NQOs Enhances Metastatic Ability in Oral Squamous Cell Carcinoma" International Journal of Molecular Sciences 23, no. 18: 10704. https://doi.org/10.3390/ijms231810704
APA StyleNakashima, C., Fujiwara-Tani, R., Mori, S., Kishi, S., Ohmori, H., Fujii, K., Mori, T., Miyagawa, Y., Yamamoto, K., Kirita, T., Luo, Y., & Kuniyasu, H. (2022). An Axis between the Long Non-Coding RNA HOXA11-AS and NQOs Enhances Metastatic Ability in Oral Squamous Cell Carcinoma. International Journal of Molecular Sciences, 23(18), 10704. https://doi.org/10.3390/ijms231810704