
Emerging Roles of Ceramides in Breast Cancer Biology and Therapy


Abstract
:
1. Introduction
2. Ceramides: Structure and Production
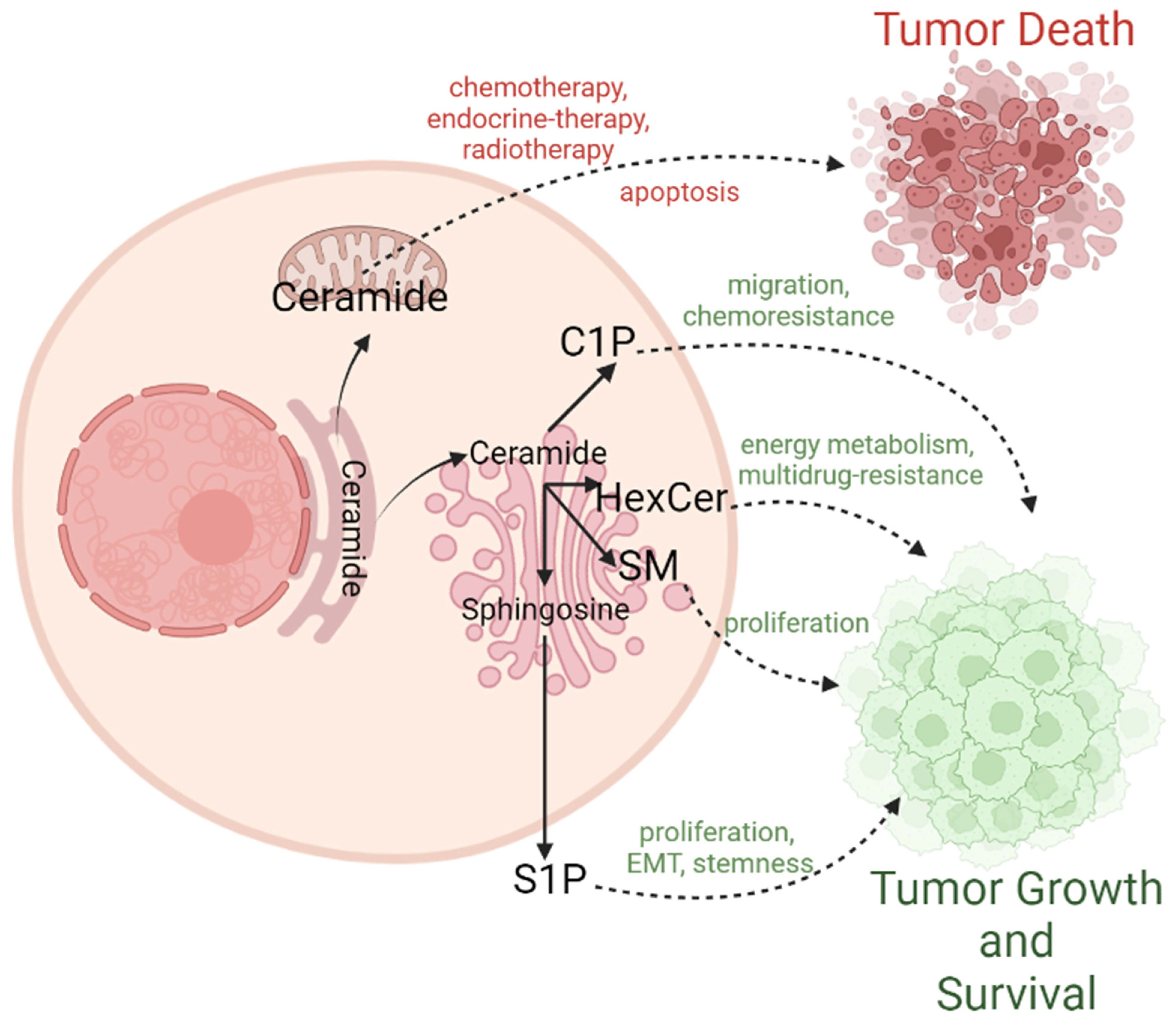
3. Canonical Role of Ceramides in BC: A Bona Fide Inducer of Cell Death
4. The Other Role of Ceramides: Conversion to Pro-Survival Sphingolipids
4.1. Sphingosine-1-Phosphate (S1P)
4.2. Ceramide-1-Phosphate (C1P)
4.3. Sphingomyelins (SM)
4.4. Hexosylceramides (HexCer)
5. Therapeutic Implications of Ceramides in Breast Cancer
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- National Cancer Institute (NCI) Surveillance; End Results Program. Cancer Stat Facts: Female Breast Cancer; National Cancer Institute (NCI): Bethesda, MD, USA, 2021.
- Espaillat, M.P.; Shamseddine, A.A.; Adada, M.M.; Hannun, Y.A.; Obeid, L.M. Ceramide and sphingosine-1-phosphate in cancer, two faces of the sphinx. Transl. Cancer Res. 2015, 4, 484–499. [Google Scholar]
- Sheridan, M.; Ogretmen, B. The Role of Ceramide Metabolism and Signaling in the Regulation of Mitophagy and Cancer Therapy. Cancers 2021, 13, 2475. [Google Scholar] [CrossRef] [PubMed]
- Moro, K.; Nagahashi, M.; Gabriel, E.; Takabe, K.; Wakai, T. Clinical application of ceramide in cancer treatment. Breast Cancer 2019, 26, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Mashhadi Akbar Boojar, M.; Mashhadi Akbar Boojar, M.; Golmohammad, S. Ceramide pathway: A novel approach to cancer chemotherapy. Egypt. J. Basic Appl. Sci. 2018, 5, 237–244. [Google Scholar] [CrossRef]
- Fahy, E.; Subramaniam, S.; Brown, H.A.; Glass, C.K.; Merrill, A.H., Jr.; Murphy, R.C.; Raetz, C.R.; Russell, D.W.; Seyama, Y.; Shaw, W.; et al. A comprehensive classification system for lipids. J. Lipid. Res. 2005, 46, 839–861. [Google Scholar] [CrossRef]
- Goni, F.M.; Contreras, F.X.; Montes, L.R.; Sot, J.; Alonso, A. Biophysics (and sociology) of ceramides. Biochem. Soc. Symp. 2005, 72, 177–188. [Google Scholar] [CrossRef]
- Vielhaber, G.; Pfeiffer, S.; Brade, L.; Lindner, B.; Goldmann, T.; Vollmer, E.; Hintze, U.; Wittern, K.P.; Wepf, R. Localization of ceramide and glucosylceramide in human epidermis by immunogold electron microscopy. J. Investig. Derm. 2001, 117, 1126–1136. [Google Scholar] [CrossRef]
- Obeid, L.M.; Linardic, C.M.; Karolak, L.A.; Hannun, Y.A. Programmed cell death induced by ceramide. Science 1993, 259, 1769–1771. [Google Scholar] [CrossRef]
- Modur, V.; Zimmerman, G.A.; Prescott, S.M.; McIntyre, T.M. Endothelial cell inflammatory responses to tumor necrosis factor alpha. Ceramide-dependent and -independent mitogen-activated protein kinase cascades. J. Biol. Chem. 1996, 271, 13094–13102. [Google Scholar] [CrossRef]
- Garcia-Ruiz, C.; Colell, A.; Mari, M.; Morales, A.; Fernandez-Checa, J.C. Direct effect of ceramide on the mitochondrial electron transport chain leads to generation of reactive oxygen species. Role of mitochondrial glutathione. J. Biol. Chem. 1997, 272, 11369–11377. [Google Scholar] [CrossRef]
- Geilen, C.C.; Bektas, M.; Wieder, T.; Kodelja, V.; Goerdt, S.; Orfanos, C.E. 1alpha,25-dihydroxyvitamin D3 induces sphingomyelin hydrolysis in HaCaT cells via tumor necrosis factor alpha. J. Biol. Chem. 1997, 272, 8997–9001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, M.; Nakashima, S.; Banno, Y.; Yamakawa, H.; Hayashi, K.; Takenaka, K.; Nishimura, Y.; Sakai, N.; Nozawa, Y. Ordering of ceramide formation, caspase activation, and Bax/Bcl-2 expression during etoposide-induced apoptosis in C6 glioma cells. Cell Death Differ. 2000, 7, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Wiesner, D.A.; Dawson, G. Staurosporine induces programmed cell death in embryonic neurons and activation of the ceramide pathway. J. Neurochem. 1996, 66, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- Come, M.G.; Bettaieb, A.; Skladanowski, A.; Larsen, A.K.; Laurent, G. Alteration of the daunorubicin-triggered sphingomyelin-ceramide pathway and apoptosis in MDR cells: Influence of drug transport abnormalities. Int. J. Cancer 1999, 81, 580–587. [Google Scholar] [CrossRef]
- Cifone, M.G.; Migliorati, G.; Parroni, R.; Marchetti, C.; Millimaggi, D.; Santoni, A.; Riccardi, C. Dexamethasone-induced thymocyte apoptosis: Apoptotic signal involves the sequential activation of phosphoinositide-specific phospholipase C, acidic sphingomyelinase, and caspases. Blood 1999, 93, 2282–2296. [Google Scholar] [CrossRef]
- Alphonse, G.; Aloy, M.T.; Broquet, P.; Gerard, J.P.; Louisot, P.; Rousson, R.; Rodriguez-Lafrasse, C. Ceramide induces activation of the mitochondrial/caspases pathway in Jurkat and SCC61 cells sensitive to gamma-radiation but activation of this sequence is defective in radioresistant SQ20B cells. Int. J. Radiat. Biol. 2002, 78, 821–835. [Google Scholar] [CrossRef]
- Vit, J.P.; Rosselli, F. Role of the ceramide-signaling pathways in ionizing radiation-induced apoptosis. Oncogene 2003, 22, 8645–8652. [Google Scholar] [CrossRef]
- Takeda, Y.; Tashima, M.; Takahashi, A.; Uchiyama, T.; Okazaki, T. Ceramide generation in nitric oxide-induced apoptosis. Activation of magnesium-dependent neutral sphingomyelinase via caspase-3. J. Biol. Chem. 1999, 274, 10654–10660. [Google Scholar] [CrossRef]
- Masamune, A.; Igarashi, Y.; Hakomori, S. Regulatory role of ceramide in interleukin (IL)-1 beta-induced E-selectin expression in human umbilical vein endothelial cells. Ceramide enhances IL-1 beta action, but is not sufficient for E-selectin expression. J. Biol. Chem. 1996, 271, 9368–9375. [Google Scholar] [CrossRef]
- Birbes, H.; El Bawab, S.; Obeid, L.M.; Hannun, Y.A. Mitochondria and ceramide: Intertwined roles in regulation of apoptosis. Adv. Enzym. Regul. 2002, 42, 113–129. [Google Scholar] [CrossRef]
- Sautin, Y.; Takamura, N.; Shklyaev, S.; Nagayama, Y.; Ohtsuru, A.; Namba, H.; Yamashita, S. Ceramide-induced apoptosis of human thyroid cancer cells resistant to apoptosis by irradiation. Thyroid 2000, 10, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Lozano, J.; Menendez, S.; Morales, A.; Ehleiter, D.; Liao, W.C.; Wagman, R.; Haimovitz-Friedman, A.; Fuks, Z.; Kolesnick, R. Cell autonomous apoptosis defects in acid sphingomyelinase knockout fibroblasts. J. Biol. Chem. 2001, 276, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Selzner, M.; Bielawska, A.; Morse, M.A.; Rudiger, H.A.; Sindram, D.; Hannun, Y.A.; Clavien, P.A. Induction of apoptotic cell death and prevention of tumor growth by ceramide analogues in metastatic human colon cancer. Cancer Res. 2001, 61, 1233–1240. [Google Scholar] [PubMed]
- Kimura, K.; Markowski, M.; Edsall, L.C.; Spiegel, S.; Gelmann, E.P. Role of ceramide in mediating apoptosis of irradiated LNCaP prostate cancer cells. Cell Death Differ. 2003, 10, 240–248. [Google Scholar] [CrossRef]
- Morstein, J.; Kol, M.; Novak, A.J.E.; Feng, S.; Khayyo, S.; Hinnah, K.; Li-Purcell, N.; Pan, G.; Williams, B.M.; Riezman, H.; et al. Short Photoswitchable Ceramides Enable Optical Control of Apoptosis. ACS Chem. Biol. 2021, 16, 452–456. [Google Scholar] [CrossRef]
- Chmura, S.J.; Mauceri, H.J.; Advani, S.; Heimann, R.; Beckett, M.A.; Nodzenski, E.; Quintans, J.; Kufe, D.W.; Weichselbaum, R.R. Decreasing the apoptotic threshold of tumor cells through protein kinase C inhibition and sphingomyelinase activation increases tumor killing by ionizing radiation. Cancer Res. 1997, 57, 4340–4347. [Google Scholar]
- Alphonse, G.; Bionda, C.; Aloy, M.T.; Ardail, D.; Rousson, R.; Rodriguez-Lafrasse, C. Overcoming resistance to gamma-rays in squamous carcinoma cells by poly-drug elevation of ceramide levels. Oncogene 2004, 23, 2703–2715. [Google Scholar] [CrossRef]
- Rodriguez-Lafrasse, C.; Alphonse, G.; Aloy, M.T.; Ardail, D.; Gerard, J.P.; Louisot, P.; Rousson, R. Increasing endogenous ceramide using inhibitors of sphingolipid metabolism maximizes ionizing radiation-induced mitochondrial injury and apoptotic cell killing. Int. J. Cancer 2002, 101, 589–598. [Google Scholar] [CrossRef]
- Raisova, M.; Goltz, G.; Bektas, M.; Bielawska, A.; Riebeling, C.; Hossini, A.M.; Eberle, J.; Hannun, Y.A.; Orfanos, C.E.; Geilen, C.C. Bcl-2 overexpression prevents apoptosis induced by ceramidase inhibitors in malignant melanoma and HaCaT keratinocytes. FEBS Lett. 2002, 516, 47–52. [Google Scholar] [CrossRef]
- Wang, S.; Su, X.; Xu, M.; Xiao, X.; Li, X.; Li, H.; Keating, A.; Zhao, R.C. Exosomes secreted by mesenchymal stromal/stem cell-derived adipocytes promote breast cancer cell growth via activation of Hippo signaling pathway. Stem. Cell Res. 2019, 10, 117. [Google Scholar] [CrossRef]
- Cheng, Q.; Li, X.; Wang, Y.; Dong, M.; Zhan, F.H.; Liu, J. The ceramide pathway is involved in the survival, apoptosis and exosome functions of human multiple myeloma cells in vitro. Acta Pharm. Sin. 2018, 39, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Bruno, A.P.; Laurent, G.; Averbeck, D.; Demur, C.; Bonnet, J.; Bettaieb, A.; Levade, T.; Jaffrezou, J.P. Lack of ceramide generation in TF-1 human myeloid leukemic cells resistant to ionizing radiation. Cell Death Differ. 1998, 5, 172–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettus, B.J.; Chalfant, C.E.; Hannun, Y.A. Ceramide in apoptosis: An overview and current perspectives. Biochim. Biophys. Acta 2002, 1585, 114–125. [Google Scholar] [CrossRef]
- Siskind, L.J.; Kolesnick, R.N.; Colombini, M. Ceramide channels increase the permeability of the mitochondrial outer membrane to small proteins. J. Biol. Chem. 2002, 277, 26796–26803. [Google Scholar] [CrossRef]
- Di Paola, M.; Zaccagnino, P.; Montedoro, G.; Cocco, T.; Lorusso, M. Ceramide induces release of pro-apoptotic proteins from mitochondria by either a Ca2+-dependent or a Ca2+-independent mechanism. J. Bioenergy Biomembr. 2004, 36, 165–170. [Google Scholar] [CrossRef]
- Siskind, L.J.; Colombini, M. The lipids C2- and C16-ceramide form large stable channels. Implications for apoptosis. J. Biol. Chem. 2000, 275, 38640–38644. [Google Scholar] [CrossRef]
- Contreras, F.X.; Basanez, G.; Alonso, A.; Herrmann, A.; Goni, F.M. Asymmetric addition of ceramides but not dihydroceramides promotes transbilayer (flip-flop) lipid motion in membranes. Biophys. J. 2005, 88, 348–359. [Google Scholar] [CrossRef]
- De Mario, A.; Tosatto, A.; Hill, J.M.; Kriston-Vizi, J.; Ketteler, R.; Vecellio Reane, D.; Cortopassi, G.; Szabadkai, G.; Rizzuto, R.; Mammucari, C. Identification and functional validation of FDA-approved positive and negative modulators of the mitochondrial calcium uniporter. Cell Rep. 2021, 35, 109275. [Google Scholar] [CrossRef]
- Sot, J.; Aranda, F.J.; Collado, M.I.; Goni, F.M.; Alonso, A. Different effects of long- and short-chain ceramides on the gel-fluid and lamellar-hexagonal transitions of phospholipids: A calorimetric, NMR, and x-ray diffraction study. Biophys. J. 2005, 88, 3368–3380. [Google Scholar] [CrossRef]
- Sot, J.; Goni, F.M.; Alonso, A. Molecular associations and surface-active properties of short- and long-N-acyl chain ceramides. Biochim. Biophys. Acta 2005, 1711, 12–19. [Google Scholar] [CrossRef]
- Kim, M.H.; Park, J.W.; Lee, E.J.; Kim, S.; Shin, S.H.; Ahn, J.H.; Jung, Y.; Park, I.; Park, W.J. C16-ceramide and sphingosine 1-phosphate/S1PR2 have opposite effects on cell growth through mTOR signaling pathway regulation. Oncol. Rep. 2018, 40, 2977–2987. [Google Scholar] [CrossRef] [PubMed]
- Pani, T.; Rajput, K.; Kar, A.; Sharma, H.; Basak, R.; Medatwal, N.; Saha, S.; Dev, G.; Kumar, S.; Gupta, S.; et al. Alternative splicing of ceramide synthase 2 alters levels of specific ceramides and modulates cancer cell proliferation and migration in Luminal B breast cancer subtype. Cell Death Dis. 2021, 12, 171. [Google Scholar] [CrossRef]
- Brovkovych, V.; Izhar, Y.; Danes, J.M.; Dubrovskyi, O.; Sakallioglu, I.T.; Morrow, L.M.; Atilla-Gokcumen, G.E.; Frasor, J. Fatostatin induces pro- and anti-apoptotic lipid accumulation in breast cancer. Oncogenesis 2018, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Guardiola-Serrano, F.; Beteta-Gobel, R.; Rodriguez-Lorca, R.; Ibarguren, M.; Lopez, D.J.; Teres, S.; Alonso-Sande, M.; Higuera, M.; Torres, M.; Busquets, X.; et al. The triacylglycerol, hydroxytriolein, inhibits triple negative mammary breast cancer cell proliferation through a mechanism dependent on dihydroceramide and Akt. Oncotarget 2019, 10, 2486–2507. [Google Scholar] [CrossRef] [PubMed]
- Sha, W.; Zhou, Y.; Ling, Z.Q.; Xie, G.; Pang, X.; Wang, P.; Gu, X. Antitumor properties of Salvianolic acid B against triple-negative and hormone receptor-positive breast cancer cells via ceramide-mediated apoptosis. Oncotarget 2018, 9, 36331–36343. [Google Scholar] [CrossRef]
- Aslan, M.; Afsar, E.; Kirimlioglu, E.; Ceker, T.; Yilmaz, C. Antiproliferative Effects of Thymoquinone in MCF-7 Breast and HepG2 Liver Cancer Cells: Possible Role of Ceramide and ER Stress. Nutr. Cancer 2021, 73, 460–472. [Google Scholar] [CrossRef]
- Cao, Q.; Chen, X.; Wu, X.; Liao, R.; Huang, P.; Tan, Y.; Wang, L.; Ren, G.; Huang, J.; Dong, C. Inhibition of UGT8 suppresses basal-like breast cancer progression by attenuating sulfatide-alphaVbeta5 axis. J. Exp. Med. 2018, 215, 1679–1692. [Google Scholar] [CrossRef]
- Bhadwal, P.; Dahiya, D.; Shinde, D.; Vaiphei, K.; Math, R.G.H.; Randhawa, V.; Agnihotri, N. LC-HRMS based approach to identify novel sphingolipid biomarkers in breast cancer patients. Sci. Rep. 2020, 10, 4668. [Google Scholar] [CrossRef]
- Moro, K.; Kawaguchi, T.; Tsuchida, J.; Gabriel, E.; Qi, Q.; Yan, L.; Wakai, T.; Takabe, K.; Nagahashi, M. Ceramide species are elevated in human breast cancer and are associated with less aggressiveness. Oncotarget 2018, 9, 19874–19890. [Google Scholar] [CrossRef]
- Nagahashi, M.; Tsuchida, J.; Moro, K.; Hasegawa, M.; Tatsuda, K.; Woelfel, I.A.; Takabe, K.; Wakai, T. High levels of sphingolipids in human breast cancer. J. Surg. Res. 2016, 204, 435–444. [Google Scholar] [CrossRef]
- Schiffmann, S.; Sandner, J.; Birod, K.; Wobst, I.; Angioni, C.; Ruckhaberle, E.; Kaufmann, M.; Ackermann, H.; Lotsch, J.; Schmidt, H.; et al. Ceramide synthases and ceramide levels are increased in breast cancer tissue. Carcinogenesis 2009, 30, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Erez-Roman, R.; Pienik, R.; Futerman, A.H. Increased ceramide synthase 2 and 6 mRNA levels in breast cancer tissues and correlation with sphingosine kinase expression. Biochem. Biophys. Res. Commun. 2010, 391, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Gupta, V.; Patwardhan, G.A.; Bhinge, K.; Zhao, Y.; Bao, J.; Mehendale, H.; Cabot, M.C.; Li, Y.T.; Jazwinski, S.M. Glucosylceramide synthase upregulates MDR1 expression in the regulation of cancer drug resistance through cSrc and beta-catenin signaling. Mol. Cancer 2010, 9, 145. [Google Scholar] [CrossRef]
- Kramer, R.; Bielawski, J.; Kistner-Griffin, E.; Othman, A.; Alecu, I.; Ernst, D.; Kornhauser, D.; Hornemann, T.; Spassieva, S. Neurotoxic 1-deoxysphingolipids and paclitaxel-induced peripheral neuropathy. FASEB J. 2015, 29, 4461–4472. [Google Scholar] [CrossRef]
- Brachtendorf, S.; El-Hindi, K.; Grosch, S. Ceramide synthases in cancer therapy and chemoresistance. Prog. Lipid Res. 2019, 74, 160–185. [Google Scholar] [CrossRef] [PubMed]
- Mesicek, J.; Lee, H.; Feldman, T.; Jiang, X.; Skobeleva, A.; Berdyshev, E.V.; Haimovitz-Friedman, A.; Fuks, Z.; Kolesnick, R. Ceramide synthases 2, 5, and 6 confer distinct roles in radiation-induced apoptosis in HeLa cells. Cell Signal 2010, 22, 1300–1307. [Google Scholar] [CrossRef]
- Morad, S.A.; Levin, J.C.; Shanmugavelandy, S.S.; Kester, M.; Fabrias, G.; Bedia, C.; Cabot, M.C. Ceramide--antiestrogen nanoliposomal combinations--novel impact of hormonal therapy in hormone-insensitive breast cancer. Mol. Cancer 2012, 11, 2352–2361. [Google Scholar] [CrossRef]
- Morad, S.A.; Tan, S.F.; Feith, D.J.; Kester, M.; Claxton, D.F.; Loughran, T.P., Jr.; Barth, B.M.; Fox, T.E.; Cabot, M.C. Modification of sphingolipid metabolism by tamoxifen and N-desmethyltamoxifen in acute myelogenous leukemia--Impact on enzyme activity and response to cytotoxics. Biochim. Biophys. Acta 2015, 1851, 919–928. [Google Scholar] [CrossRef]
- El Kaffas, A.; Al-Mahrouki, A.; Hashim, A.; Law, N.; Giles, A.; Czarnota, G.J. Role of Acid Sphingomyelinase and Ceramide in Mechano-Acoustic Enhancement of Tumor Radiation Responses. J. Natl. Cancer Inst. 2018, 110, 1009–1018. [Google Scholar] [CrossRef]
- Hait, N.C.; Maiti, A. The Role of Sphingosine-1-Phosphate and Ceramide-1-Phosphate in Inflammation and Cancer. Mediat. Inflamm. 2017, 2017, 4806541. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Lu, M.; Zou, X. Ceramide-1-phosphate and its transfer proteins in eukaryotes. Chem. Phys. Lipids 2021, 240, 105135. [Google Scholar] [CrossRef] [PubMed]
- Presa, N.; Gomez-Larrauri, A.; Dominguez-Herrera, A.; Trueba, M.; Gomez-Munoz, A. Novel signaling aspects of ceramide 1-phosphate. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158630. [Google Scholar] [CrossRef]
- Ruckhaberle, E.; Karn, T.; Rody, A.; Hanker, L.; Gatje, R.; Metzler, D.; Holtrich, U.; Kaufmann, M. Gene expression of ceramide kinase, galactosyl ceramide synthase and ganglioside GD3 synthase is associated with prognosis in breast cancer. J. Cancer Res. Clin Oncol 2009, 135, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Munoz, A. The Role of Ceramide 1-Phosphate in Tumor Cell Survival and Dissemination. Adv. Cancer Res. 2018, 140, 217–234. [Google Scholar] [CrossRef]
- Zhu, S.; Xu, Y.; Wang, L.; Liao, S.; Wang, Y.; Shi, M.; Tu, Y.; Zhou, Y.; Wei, W. Ceramide kinase mediates intrinsic resistance and inferior response to chemotherapy in triple-negative breast cancer by upregulating Ras/ERK and PI3K/Akt pathways. Cancer Cell Int. 2021, 21, 42. [Google Scholar] [CrossRef] [PubMed]
- Budczies, J.; Pfitzner, B.M.; Gyorffy, B.; Winzer, K.J.; Radke, C.; Dietel, M.; Fiehn, O.; Denkert, C. Glutamate enrichment as new diagnostic opportunity in breast cancer. Int. J. Cancer 2015, 136, 1619–1628. [Google Scholar] [CrossRef]
- Simpson, N.E.; Tryndyak, V.P.; Beland, F.A.; Pogribny, I.P. An in vitro investigation of metabolically sensitive biomarkers in breast cancer progression. Breast Cancer Res. Treat. 2012, 133, 959–968. [Google Scholar] [CrossRef]
- Schomel, N.; Hancock, S.E.; Gruber, L.; Olzomer, E.M.; Byrne, F.L.; Shah, D.; Hoehn, K.L.; Turner, N.; Grosch, S.; Geisslinger, G.; et al. UGCG influences glutamine metabolism of breast cancer cells. Sci. Rep. 2019, 9, 15665. [Google Scholar] [CrossRef]
- Cardoso, M.R.; Santos, J.C.; Ribeiro, M.L.; Talarico, M.C.R.; Viana, L.R.; Derchain, S.F.M. A Metabolomic Approach to Predict Breast Cancer Behavior and Chemotherapy Response. Int. J. Mol. Sci. 2018, 19, 617. [Google Scholar] [CrossRef]
- Shajahan-Haq, A.N.; Cheema, M.S.; Clarke, R. Application of metabolomics in drug resistant breast cancer research. Metabolites 2015, 5, 100–118. [Google Scholar] [CrossRef]
- Schomel, N.; Gruber, L.; Alexopoulos, S.J.; Trautmann, S.; Olzomer, E.M.; Byrne, F.L.; Hoehn, K.L.; Gurke, R.; Thomas, D.; Ferreiros, N.; et al. UGCG overexpression leads to increased glycolysis and increased oxidative phosphorylation of breast cancer cells. Sci. Rep. 2020, 10, 8182. [Google Scholar] [CrossRef] [PubMed]
- Izgordu, H.; Vejselova Sezer, C.; Comlekci, E.; Kutlu, H.M. Characteristics of apoptosis induction in human breast cancer cells treated with a ceramidase inhibitor. Cytotechnology 2020, 72, 907–919, Erratum in Cytotechnology 2021, 73, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Ruckhaberle, E.; Holtrich, U.; Engels, K.; Hanker, L.; Gatje, R.; Metzler, D.; Karn, T.; Kaufmann, M.; Rody, A. Acid ceramidase 1 expression correlates with a better prognosis in ER-positive breast cancer. Climacteric 2009, 12, 502–513. [Google Scholar] [CrossRef]
- Alshaker, H.; Thrower, H.; Pchejetski, D. Sphingosine Kinase 1 in Breast Cancer-A New Molecular Marker and a Therapy Target. Front. Oncol. 2020, 10, 289. [Google Scholar] [CrossRef]
- Fang, T.; Jiang, Y.X.; Chen, L.; Huang, L.; Tian, X.H.; Zhou, Y.D.; Nagle, D.G.; Zhang, D.D. Coix Seed Oil Exerts an Anti-Triple-Negative Breast Cancer Effect by Disrupting miR-205/S1PR1 Axis. Front. Pharm. 2020, 11, 529962. [Google Scholar] [CrossRef] [PubMed]
- Alshaker, H.; Srivats, S.; Monteil, D.; Wang, Q.; Low, C.M.R.; Pchejetski, D. Field template-based design and biological evaluation of new sphingosine kinase 1 inhibitors. Breast Cancer Res. Treat. 2018, 172, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Ochnik, A.M.; Baxter, R.C. Insulin-like growth factor receptor and sphingosine kinase are prognostic and therapeutic targets in breast cancer. BMC Cancer 2017, 17, 820. [Google Scholar] [CrossRef]
- Tsuchida, J.; Nagahashi, M.; Takabe, K.; Wakai, T. Clinical Impact of Sphingosine-1-Phosphate in Breast Cancer. Mediat. Inflamm. 2017, 2017, 2076239. [Google Scholar] [CrossRef]
- Marzec, K.A.; Baxter, R.C.; Martin, J.L. Targeting Insulin-Like Growth Factor Binding Protein-3 Signaling in Triple-Negative Breast Cancer. Biomed. Res. Int. 2015, 2015, 638526. [Google Scholar] [CrossRef]
- Hii, L.W.; Chung, F.F.; Mai, C.W.; Yee, Z.Y.; Chan, H.H.; Raja, V.J.; Dephoure, N.E.; Pyne, N.J.; Pyne, S.; Leong, C.O. Sphingosine Kinase 1 Regulates the Survival of Breast Cancer Stem Cells and Non-stem Breast Cancer Cells by Suppression of STAT1. Cells 2020, 9, 886. [Google Scholar] [CrossRef]
- Alshaker, H.; Wang, Q.; Srivats, S.; Chao, Y.; Cooper, C.; Pchejetski, D. New FTY720-docetaxel nanoparticle therapy overcomes FTY720-induced lymphopenia and inhibits metastatic breast tumour growth. Breast Cancer Res. Treat. 2017, 165, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Chen, Z.; Feng, H.; Chen, Y.; Zhang, C.; Yu, J.; Luo, Y.; Zhao, L.; Jiang, X.; Shi, F. Sphingomyelin synthase 2 promotes an aggressive breast cancer phenotype by disrupting the homoeostasis of ceramide and sphingomyelin. Cell Death Dis. 2019, 10, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shammout, O.D.A.; Ashmawy, N.S.; Shakartalla, S.B.; Altaie, A.M.; Semreen, M.H.; Omar, H.A.; Soliman, S.S.M. Comparative sphingolipidomic analysis reveals significant differences between doxorubicin-sensitive and -resistance MCF-7 cells. PLoS ONE 2021, 16, e0258363. [Google Scholar] [CrossRef]
- Alshaker, H.; Wang, Q.; Brewer, D.; Pchejetski, D. Transcriptome-Wide Effects of Sphingosine Kinases Knockdown in Metastatic Prostate and Breast Cancer Cells: Implications for Therapeutic Targeting. Front. Pharm. 2019, 10, 303. [Google Scholar] [CrossRef]
- Geffken, K.; Spiegel, S. Sphingosine kinase 1 in breast cancer. Adv. Biol. Regul. 2018, 67, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liu, B. Sphk1 participates in malignant progression of breast cancer by regulating epithelial-mesenchymal transition and stem cell characteristics. Tissue Cell 2020, 65, 101380. [Google Scholar] [CrossRef] [PubMed]
- Sakharkar, M.K.; Kaur Dhillon, S.; Chidambaram, S.B.; Essa, M.M.; Yang, J. Gene Pair Correlation Coefficients in Sphingolipid Metabolic Pathway as a Potential Prognostic Biomarker for Breast Cancer. Cancers 2020, 12, 1747. [Google Scholar] [CrossRef]
- Newcomb, B.; Rhein, C.; Mileva, I.; Ahmad, R.; Clarke, C.J.; Snider, J.; Obeid, L.M.; Hannun, Y.A. Identification of an acid sphingomyelinase ceramide kinase pathway in the regulation of the chemokine CCL5. J. Lipid. Res. 2018, 59, 1219–1229. [Google Scholar] [CrossRef]
- Schwalm, S.; Erhardt, M.; Romer, I.; Pfeilschifter, J.; Zangemeister-Wittke, U.; Huwiler, A. Ceramide Kinase Is Upregulated in Metastatic Breast Cancer Cells and Contributes to Migration and Invasion by Activation of PI 3-Kinase and Akt. Int. J. Mol. Sci. 2020, 21, 1396. [Google Scholar] [CrossRef]
- Pal, P.; Millner, A.; Semina, S.E.; Huggins, R.J.; Running, L.; Aga, D.S.; Tonetti, D.A.; Schiff, R.; Greene, G.L.; Atilla-Gokcumen, G.E.; et al. Endocrine Therapy-Resistant Breast Cancer Cells Are More Sensitive to Ceramide Kinase Inhibition and Elevated Ceramide Levels Than Therapy-Sensitive Breast Cancer Cells. Cancers 2022, 14, 2380. [Google Scholar] [CrossRef]
- Granado, M.H.; Gangoiti, P.; Ouro, A.; Arana, L.; Gomez-Munoz, A. Ceramide 1-phosphate inhibits serine palmitoyltransferase and blocks apoptosis in alveolar macrophages. Biochim. Biophys. Acta 2009, 1791, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Inwald, E.C.; Klinkhammer-Schalke, M.; Hofstadter, F.; Zeman, F.; Koller, M.; Gerstenhauer, M.; Ortmann, O. Ki-67 is a prognostic parameter in breast cancer patients: Results of a large population-based cohort of a cancer registry. Breast Cancer Res. Treat 2013, 139, 539–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kartal Yandim, M.; Apohan, E.; Baran, Y. Therapeutic potential of targeting ceramide/glucosylceramide pathway in cancer. Cancer Chemother. Pharm. 2013, 71, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, L.; Liu, D.; Wang, C. Ceramide glycosylation and related enzymes in cancer signaling and therapy. Biomed. Pharm. 2021, 139, 111565. [Google Scholar] [CrossRef]
- Zhang, X.; Li, J.; Qiu, Z.; Gao, P.; Wu, X.; Zhou, G. Co-suppression of MDR1 (multidrug resistance 1) and GCS (glucosylceramide synthase) restores sensitivity to multidrug resistance breast cancer cells by RNA interference (RNAi). Cancer Biol. 2009, 8, 1117–1121. [Google Scholar] [CrossRef] [PubMed]
- Wegner, M.S.; Schomel, N.; Gruber, L.; Ortel, S.B.; Kjellberg, M.A.; Mattjus, P.; Kurz, J.; Trautmann, S.; Peng, B.; Wegner, M.; et al. UDP-glucose ceramide glucosyltransferase activates AKT, promoted proliferation, and doxorubicin resistance in breast cancer cells. Cell Mol. Life Sci. 2018, 75, 3393–3410. [Google Scholar] [CrossRef]
- Jacobs, A.T.; Martinez Castaneda-Cruz, D.; Rose, M.M.; Connelly, L. Targeted therapy for breast cancer: An overview of drug classes and outcomes. Biochem. Pharm. 2022, 204, 115209. [Google Scholar] [CrossRef]
- Fisher, B.; Bauer, M.; Margolese, R.; Poisson, R.; Pilch, Y.; Redmond, C.; Fisher, E.; Wolmark, N.; Deutsch, M.; Montague, E.; et al. Five-year results of a randomized clinical trial comparing total mastectomy and segmental mastectomy with or without radiation in the treatment of breast cancer. N. Engl. J. Med. 1985, 312, 665–673. [Google Scholar] [CrossRef]
- Clifford, R.E.; Bowden, D.; Blower, E.; Kirwan, C.C.; Vimalachandran, D. Does tamoxifen have a therapeutic role outside of breast cancer? A systematic review of the evidence. Surg. Oncol. 2020, 33, 100–107. [Google Scholar] [CrossRef]
- Elojeimy, S.; Holman, D.H.; Liu, X.; El-Zawahry, A.; Villani, M.; Cheng, J.C.; Mahdy, A.; Zeidan, Y.; Bielwaska, A.; Hannun, Y.A.; et al. New insights on the use of desipramine as an inhibitor for acid ceramidase. FEBS Lett. 2006, 580, 4751–4756. [Google Scholar] [CrossRef]
- Santana, P.; Pena, L.A.; Haimovitz-Friedman, A.; Martin, S.; Green, D.; McLoughlin, M.; Cordon-Cardo, C.; Schuchman, E.H.; Fuks, Z.; Kolesnick, R. Acid sphingomyelinase-deficient human lymphoblasts and mice are defective in radiation-induced apoptosis. Cell 1996, 86, 189–199. [Google Scholar] [CrossRef]
- Zeidan, Y.H.; Jenkins, R.W.; Hannun, Y.A. Remodeling of cellular cytoskeleton by the acid sphingomyelinase/ceramide pathway. J. Cell Biol. 2008, 181, 335–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heakal, Y.; Kester, M. Nanoliposomal short-chain ceramide inhibits agonist-dependent translocation of neurotensin receptor 1 to structured membrane microdomains in breast cancer cells. Mol. Cancer Res. 2009, 7, 724–734. [Google Scholar] [CrossRef] [PubMed]
- Haakenson, J.K.; Khokhlatchev, A.V.; Choi, Y.J.; Linton, S.S.; Zhang, P.; Zaki, P.M.; Fu, C.; Cooper, T.K.; Manni, A.; Zhu, J.; et al. Lysosomal degradation of CD44 mediates ceramide nanoliposome-induced anoikis and diminished extravasation in metastatic carcinoma cells. J. Biol. Chem. 2015, 290, 8632–8643. [Google Scholar] [CrossRef]
- Jatoi, A.; Suman, V.J.; Schaefer, P.; Block, M.; Loprinzi, C.; Roche, P.; Garneau, S.; Morton, R.; Stella, P.J.; Alberts, S.R.; et al. A phase II study of topical ceramides for cutaneous breast cancer. Breast Cancer Res. Treat 2003, 80, 99–104. [Google Scholar] [CrossRef]
- Lerata, M.S.; D’Souza, S.; Sibuyi, N.R.S.; Dube, A.; Meyer, M.; Samaai, T.; Antunes, E.M.; Beukes, D.R. Encapsulation of Variabilin in Stearic Acid Solid Lipid Nanoparticles Enhances Its Anticancer Activity in Vitro. Molecules 2020, 25, 830. [Google Scholar] [CrossRef]
- Lee, S.Y.; Ko, S.H.; Shim, J.S.; Kim, D.D.; Cho, H.J. Tumor Targeting and Lipid Rafts Disrupting Hyaluronic Acid-Cyclodextrin-Based Nanoassembled Structure for Cancer Therapy. ACS Appl. Mater. Interfaces 2018, 10, 36628–36640. [Google Scholar] [CrossRef]
- Migotto, A.; Carvalho, V.F.M.; Salata, G.C.; da Silva, F.W.M.; Yan, C.Y.I.; Ishida, K.; Costa-Lotufo, L.V.; Steiner, A.A.; Lopes, L.B. Multifunctional nanoemulsions for intraductal delivery as a new platform for local treatment of breast cancer. Drug Deliv. 2018, 25, 654–667. [Google Scholar] [CrossRef]
- Overbye, A.; Holsaeter, A.M.; Markus, F.; Skalko-Basnet, N.; Iversen, T.G.; Torgersen, M.L.; Sonstevold, T.; Engebraaten, O.; Flatmark, K.; Maelandsmo, G.M.; et al. Ceramide-containing liposomes with doxorubicin: Time and cell-dependent effect of C6 and C12 ceramide. Oncotarget 2017, 8, 76921–76934. [Google Scholar] [CrossRef]
- Tonelli, F.; Lim, K.G.; Loveridge, C.; Long, J.; Pitson, S.M.; Tigyi, G.; Bittman, R.; Pyne, S.; Pyne, N.J. FTY720 and (S)-FTY720 vinylphosphonate inhibit sphingosine kinase 1 and promote its proteasomal degradation in human pulmonary artery smooth muscle, breast cancer and androgen-independent prostate cancer cells. Cell Signal 2010, 22, 1536–1542. [Google Scholar] [CrossRef]
- Azuma, H.; Horie, S.; Muto, S.; Otsuki, Y.; Matsumoto, K.; Morimoto, J.; Gotoh, R.; Okuyama, A.; Suzuki, S.; Katsuoka, Y.; et al. Selective cancer cell apoptosis induced by FTY720; evidence for a Bcl-dependent pathway and impairment in ERK activity. Anticancer Res 2003, 23, 3183–3193. [Google Scholar] [PubMed]
- Hait, N.C.; Avni, D.; Yamada, A.; Nagahashi, M.; Aoyagi, T.; Aoki, H.; Dumur, C.I.; Zelenko, Z.; Gallagher, E.J.; Leroith, D.; et al. The phosphorylated prodrug FTY720 is a histone deacetylase inhibitor that reactivates ERalpha expression and enhances hormonal therapy for breast cancer. Oncogenesis 2015, 4, e156. [Google Scholar] [CrossRef] [PubMed]
- Rupp, T.; Pelouin, O.; Genest, L.; Legrand, C.; Froget, G.; Castagne, V. Therapeutic potential of Fingolimod in triple negative breast cancer preclinical models. Transl. Oncol. 2021, 14, 100926. [Google Scholar] [CrossRef] [PubMed]
- McGowan, E.M.; Alling, N.; Jackson, E.A.; Yagoub, D.; Haass, N.K.; Allen, J.D.; Martinello-Wilks, R. Evaluation of cell cycle arrest in estrogen responsive MCF-7 breast cancer cells: Pitfalls of the MTS assay. PLoS ONE 2011, 6, e20623. [Google Scholar] [CrossRef]
- Mousseau, Y.; Mollard, S.; Faucher-Durand, K.; Richard, L.; Nizou, A.; Cook-Moreau, J.; Baaj, Y.; Qiu, H.; Plainard, X.; Fourcade, L.; et al. Fingolimod potentiates the effects of sunitinib malate in a rat breast cancer model. Breast Cancer Res. Treat 2012, 134, 31–40. [Google Scholar] [CrossRef]
- Marvaso, G.; Barone, A.; Amodio, N.; Raimondi, L.; Agosti, V.; Altomare, E.; Scotti, V.; Lombardi, A.; Bianco, R.; Bianco, C.; et al. Sphingosine analog fingolimod (FTY720) increases radiation sensitivity of human breast cancer cells in vitro. Cancer Biol. 2014, 15, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Katsuta, E.; Yan, L.; Nagahashi, M.; Raza, A.; Sturgill, J.L.; Lyon, D.E.; Rashid, O.M.; Hait, N.C.; Takabe, K. Doxorubicin effect is enhanced by sphingosine-1-phosphate signaling antagonist in breast cancer. J. Surg. Res. 2017, 219, 202–213. [Google Scholar] [CrossRef]
- Nagahashi, M.; Yamada, A.; Katsuta, E.; Aoyagi, T.; Huang, W.C.; Terracina, K.P.; Hait, N.C.; Allegood, J.C.; Tsuchida, J.; Yuza, K.; et al. Targeting the SphK1/S1P/S1PR1 Axis That Links Obesity, Chronic Inflammation, and Breast Cancer Metastasis. Cancer Res. 2018, 78, 1713–1725. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ding, Y.; Wang, C.; Gao, M.; Xu, Y.; Ma, X.; Ma, X.; Cui, H.; Li, L. Fenretinide-polyethylene glycol (PEG) conjugate with improved solubility enhanced cytotoxicity to cancer cell and potent in vivo efficacy. Pharm. Dev. Technol. 2020, 25, 962–970. [Google Scholar] [CrossRef]
- Decensi, A.; Bonanni, B.; Baglietto, L.; Guerrieri-Gonzaga, A.; Ramazzotto, F.; Johansson, H.; Robertson, C.; Marinucci, I.; Mariette, F.; Sandri, M.T.; et al. A two-by-two factorial trial comparing oral with transdermal estrogen therapy and fenretinide with placebo on breast cancer biomarkers. Clin. Cancer Res. 2004, 10, 4389–4397. [Google Scholar] [CrossRef]
- Formelli, F.; Camerini, T.; Cavadini, E.; Appierto, V.; Villani, M.G.; Costa, A.; De Palo, G.; Di Mauro, M.G.; Veronesi, U. Fenretinide breast cancer prevention trial: Drug and retinol plasma levels in relation to age and disease outcome. Cancer Epidemiol Biomark. Prev 2003, 12, 34–41. [Google Scholar]
- Veronesi, U.; Mariani, L.; Decensi, A.; Formelli, F.; Camerini, T.; Miceli, R.; Di Mauro, M.G.; Costa, A.; Marubini, E.; Sporn, M.B.; et al. Fifteen-year results of a randomized phase III trial of fenretinide to prevent second breast cancer. Ann. Oncol. 2006, 17, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Camerini, T.; Mariani, L.; De Palo, G.; Marubini, E.; Di Mauro, M.G.; Decensi, A.; Costa, A.; Veronesi, U. Safety of the synthetic retinoid fenretinide: Long-term results from a controlled clinical trial for the prevention of contralateral breast cancer. J. Clin. Oncol. 2001, 19, 1664–1670. [Google Scholar] [CrossRef] [PubMed]
- Johansson, H.; Gandini, S.; Guerrieri-Gonzaga, A.; Iodice, S.; Ruscica, M.; Bonanni, B.; Gulisano, M.; Magni, P.; Formelli, F.; Decensi, A. Effect of fenretinide and low-dose tamoxifen on insulin sensitivity in premenopausal women at high risk for breast cancer. Cancer Res. 2008, 68, 9512–9518. [Google Scholar] [CrossRef] [PubMed]
- Johansson, H.; Bonanni, B.; Gandini, S.; Guerrieri-Gonzaga, A.; Cazzaniga, M.; Serrano, D.; Macis, D.; Puccio, A.; Sandri, M.T.; Gulisano, M.; et al. Circulating hormones and breast cancer risk in premenopausal women: A randomized trial of low-dose tamoxifen and fenretinide. Breast Cancer Res. Treat. 2013, 142, 569–578. [Google Scholar] [CrossRef]
- Macis, D.; Gandini, S.; Guerrieri-Gonzaga, A.; Johansson, H.; Magni, P.; Ruscica, M.; Lazzeroni, M.; Serrano, D.; Cazzaniga, M.; Mora, S.; et al. Prognostic effect of circulating adiponectin in a randomized 2 x 2 trial of low-dose tamoxifen and fenretinide in premenopausal women at risk for breast cancer. J. Clin. Oncol. 2012, 30, 151–157. [Google Scholar] [CrossRef]
- Serrano, D.; Gandini, S.; Guerrieri-Gonzaga, A.; Feroce, I.; Johansson, H.; Macis, D.; Aristarco, V.; Bonanni, B.; DeCensi, A. Quality of Life in a Randomized Breast Cancer Prevention Trial of Low-Dose Tamoxifen and Fenretinide in Premenopausal Women. Cancer Prev. Res. 2018, 11, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Decensi, A.; Robertson, C.; Guerrieri-Gonzaga, A.; Serrano, D.; Cazzaniga, M.; Mora, S.; Gulisano, M.; Johansson, H.; Galimberti, V.; Cassano, E.; et al. Randomized double-blind 2 x 2 trial of low-dose tamoxifen and fenretinide for breast cancer prevention in high-risk premenopausal women. J. Clin. Oncol. 2009, 27, 3749–3756. [Google Scholar] [CrossRef]
- Acharya, S.; Yao, J.; Li, P.; Zhang, C.; Lowery, F.J.; Zhang, Q.; Guo, H.; Qu, J.; Yang, F.; Wistuba, I.I.; et al. Sphingosine Kinase 1 Signaling Promotes Metastasis of Triple-Negative Breast Cancer. Cancer Res. 2019, 79, 4211–4226. [Google Scholar] [CrossRef]
- Ling, L.U.; Tan, K.B.; Lin, H.; Chiu, G.N. The role of reactive oxygen species and autophagy in safingol-induced cell death. Cell Death Dis. 2011, 2, e129. [Google Scholar] [CrossRef]
- French, K.J.; Zhuang, Y.; Maines, L.W.; Gao, P.; Wang, W.; Beljanski, V.; Upson, J.J.; Green, C.L.; Keller, S.N.; Smith, C.D. Pharmacology and antitumor activity of ABC294640, a selective inhibitor of sphingosine kinase-2. J. Pharm. Exp. 2010, 333, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Antoon, J.W.; White, M.D.; Meacham, W.D.; Slaughter, E.M.; Muir, S.E.; Elliott, S.; Rhodes, L.V.; Ashe, H.B.; Wiese, T.E.; Smith, C.D.; et al. Antiestrogenic effects of the novel sphingosine kinase-2 inhibitor ABC294640. Endocrinology 2010, 151, 5124–5135. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Ma, D.; Cao, Y.; Hu, L.; Liu, S.; Yan, D.; Zhang, S.; Zhang, G.; Wang, Z.; Wu, J.; et al. SphK2/S1P Promotes Metastasis of Triple-Negative Breast Cancer Through the PAK1/LIMK1/Cofilin1 Signaling Pathway. Front. Mol. Biosci. 2021, 8, 598218. [Google Scholar] [CrossRef] [PubMed]
- Antoon, J.W.; Meacham, W.D.; Bratton, M.R.; Slaughter, E.M.; Rhodes, L.V.; Ashe, H.B.; Wiese, T.E.; Burow, M.E.; Beckman, B.S. Pharmacological inhibition of sphingosine kinase isoforms alters estrogen receptor signaling in human breast cancer. J. Mol. Endocrinol. 2011, 46, 205–216. [Google Scholar] [CrossRef]
- Gebremeskel, S.; Nelson, A.; Walker, B.; Oliphant, T.; Lobert, L.; Mahoney, D.; Johnston, B. Natural killer T cell immunotherapy combined with oncolytic vesicular stomatitis virus or reovirus treatments differentially increases survival in mouse models of ovarian and breast cancer metastasis. J. Immunother. Cancer Cancer 2021, 9, e002096. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Z.; Yu, F.; Li, M.; Zhu, H.; Wang, K.; Meng, M.; Zhao, W. The Adjuvant of alpha-Galactosylceramide Presented by Gold Nanoparticles Enhances Antitumor Immune Responses of MUC1 Antigen-Based Tumor Vaccines. Int. J. Nanomed. 2021, 16, 403–420. [Google Scholar] [CrossRef] [PubMed]
- Companioni, O.; Mir, C.; Garcia-Mayea, Y.; ME, L.L. Targeting Sphingolipids for Cancer Therapy. Front. Oncol. 2021, 11, 745092. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Enzyme Name (Abbreviation) | Gene Name(s) | Actions | Major Implication(s) in BC | Inhibitor | Citations |
|---|---|---|---|---|---|
| Serine palmitoyl transferase (SPT) | SPTLC1–3, SPTSSA-B | De novo ceramide synthesis | Enzyme activity increases in response to chemo- and radiotherapy | [55] | |
| Ceramide synthase | CERS1 | C18:0, C20:0 ceramide synthesis | Ceramide production under different stimulus | FB1 | [56] |
| CERS2 | C20:0, C22:0, C24:0, C26:0 ceramide synthesis | Long-chain ceramide production; alternative splicing drives aggressive luminal B phenotype | [43] | ||
| CERS3 | C16:0, C18:0, C22:0, C24:0 ceramide synthesis | Ceramide production under different stimulus | [44,45,46,47,48,49,56,57] | ||
| CERS4 | C18:0, C20:0, C22:0, C24:0, C26:0 ceramide synthesis | ||||
| CERS5 | C14:0, C16:0 C18:0, C18:1 ceramide synthesis | ||||
| CERS6 | C14:0, C16:0, C18:0 ceramide synthesis | Short-chain ceramide production; inhibits cell proliferation through mTOR pathway. | [42] | ||
| Sphingomyelinase (SMase) | SMPD2 | Ceramide production | Induce cell cycle arrest | GW4869 | [31,32] |
| SMPD1 | Ceramide production | Activity is required for chemo and radiotherapy | [58,59,60] | ||
| Ceramide kinase | CERK | C1P generation | Cell migration and metastasis | NVP-231 | [61,62,63,64,65,66] |
| UDP-glucose ceramide glucosyltransferase | UGCG | Glucosylceramide generation | Metabolic reprogramming, increased energy metabolism | [67,68,69,70,71,72] | |
| Acid Ceramidase | ASAH1 | Sphingosine production and subsequent S1P production | S1P generation for promoting BC growth | D-erythro-MAPP | [73,74] |
| Sphingosine kinase | SPHK1/2 | S1P generation | BC growth and proliferation | FTY720 | [75,76,77,78,79,80,81,82] |
| Sphingomyelin synthase | SGMS1/2 | SM generation | Promoting EMT, metastasis and chemoresistance | [83,84] |
| Drug/Compound Name | Target | Combination | Phase | Citations |
|---|---|---|---|---|
| Fingolimod (FTY720) | Structural analog of sphingosine, S1PR antagonist | Alone | Preclinical | [111,112,113,114,115] |
| Sunitinib malate | Preclinical | [116] | ||
| Radiation | Preclinical | [117] | ||
| Doxorubicin | Preclinical | [118] | ||
| Cisplatin | Preclinical | [119] | ||
| Fenretinide | Inhibit DEGS1/2 | Alone | Preclinical | [120] |
| Alone | Phase I/II | [121,122,123,124] | ||
| Tamoxifen | Phase I/II | [125,126,127,128,129] | ||
| Safingol | Inhibit SPHK1 | Alone | Preclinical | [130,131] |
| ABC294640 | Inhibit SPHK2 and DEGS1 | Alone | Preclinical | [132,133,134] |
| Ceramide- nanoliposomes (CNL) | Ceramide delivery | Alone | Preclinical | [104,105,106] |
| Tamoxifen | Preclinical | [58] | ||
| SKI-II | SPHK1/2 inhibitor | Alone | Preclinical | [78,135] |
| α-GalCer | Synthetic glycolipid α-galactosyl ceramide, a strong immunostimulant | Alone | Preclinical | [136,137] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pal, P.; Atilla-Gokcumen, G.E.; Frasor, J. Emerging Roles of Ceramides in Breast Cancer Biology and Therapy. Int. J. Mol. Sci. 2022, 23, 11178. https://doi.org/10.3390/ijms231911178
Pal P, Atilla-Gokcumen GE, Frasor J. Emerging Roles of Ceramides in Breast Cancer Biology and Therapy. International Journal of Molecular Sciences. 2022; 23(19):11178. https://doi.org/10.3390/ijms231911178
Chicago/Turabian StylePal, Purab, G. Ekin Atilla-Gokcumen, and Jonna Frasor. 2022. "Emerging Roles of Ceramides in Breast Cancer Biology and Therapy" International Journal of Molecular Sciences 23, no. 19: 11178. https://doi.org/10.3390/ijms231911178
APA StylePal, P., Atilla-Gokcumen, G. E., & Frasor, J. (2022). Emerging Roles of Ceramides in Breast Cancer Biology and Therapy. International Journal of Molecular Sciences, 23(19), 11178. https://doi.org/10.3390/ijms231911178