
Identification and Molecular Characterization of a Divergent Asian-like Canine Parvovirus Type 2b (CPV-2b) Strain in Southern Italy

 , ,
, ,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Clinical Case
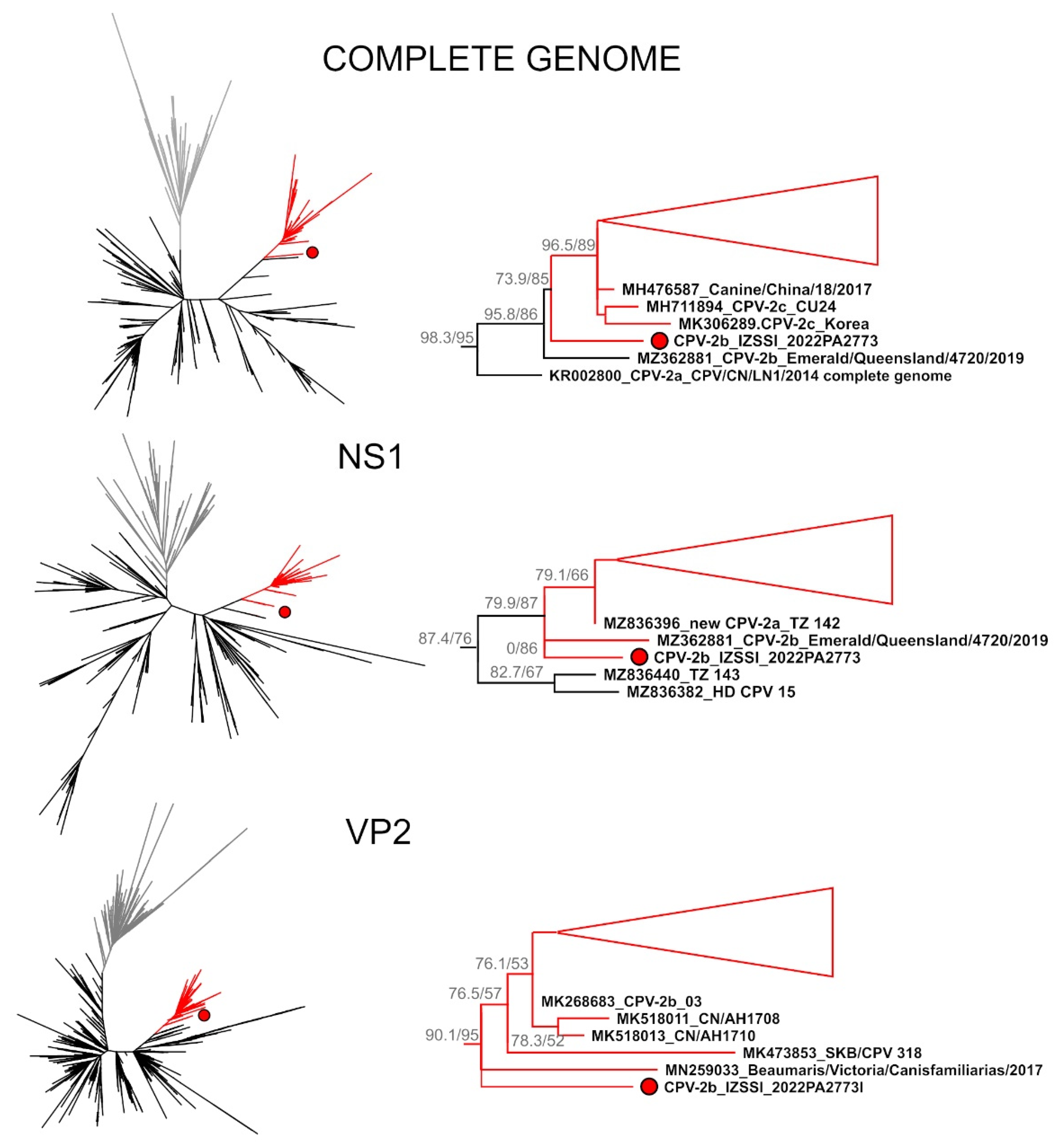
2.2. Identification, Genetic Characterization, and Phylogenetic Analysis of a Divergent Strain
3. Discussion
4. Materials and Methods
4.1. Nucleic Acid Extraction and Viral Screening
4.2. Sequence, Phylogenetic, and Recombination Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.-M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Pénzes, J.J.; et al. ICTV Virus Taxonomy Profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Buonavoglia, C. Canine Parvovirus—A Review of Epidemiological and Diagnostic Aspects, with Emphasis on Type 2c. Vet. Microbiol. 2012, 155, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Reed, A.P.; Jones, E.V.; Miller, T.J. Nucleotide Sequence and Genome Organization of Canine Parvovirus. J. Virol. 1988, 62, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, L.E. An Annotated Historical Account of Canine Parvovirus. J. Vet. Med. Ser. B Infect. Dis. Vet. Public Health 2005, 52, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Parrish, C.R.; Aquadro, C.F.; Strassheim, M.L.; Evermann, J.F.; Sgro, J.Y.; Mohammed, H.O. Rapid Antigenic-Type Replacement and DNA Sequence Evolution of Canine Parvovirus. J. Virol. 1991, 65, 6544–6552. [Google Scholar] [CrossRef]
- Buonavoglia, C.; Martella, V.; Pratelli, A.; Tempesta, M.; Cavalli, A.; Buonavoglia, D.; Bozzo, G.; Elia, G.; Decaro, N.; Carmichael, L. Evidence for Evolution of Canine Parvovirus Type 2 in Italy. J. Gen. Virol. 2001, 82, 3021–3025. [Google Scholar] [CrossRef]
- Mira, F.; Dowgier, G.; Purpari, G.; Vicari, D.; Di Bella, S.; Macaluso, G.; Gucciardi, F.; Randazzo, V.; Decaro, N.; Guercio, A. Molecular Typing of a Novel Canine Parvovirus Type 2a Mutant Circulating in Italy. Infect. Genet. Evol. 2018, 61, 67–73. [Google Scholar] [CrossRef]
- Voorhees, I.E.H.; Lee, H.; Allison, A.B.; Lopez-Astacio, R.; Goodman, L.B.; Oyesola, O.O.; Omobowale, O.; Fagbohun, O.; Dubovi, E.J.; Hafenstein, S.L.; et al. Limited Intrahost Diversity and Background Evolution Accompany 40 Years of Canine Parvovirus Host Adaptation and Spread. J. Virol. 2019, 94, e01162-19. [Google Scholar] [CrossRef]
- Mira, F.; Canuti, M.; Purpari, G.; Cannella, V.; Di Bella, S.; Occhiogrosso, L.; Schirò, G.; Chiaramonte, G.; Barreca, S.; Pisano, P.; et al. Molecular Characterization and Evolutionary Analyses of Carnivore Protoparvovirus 1 NS1 Gene. Viruses 2019, 11, 308. [Google Scholar] [CrossRef]
- Pratelli, A.; Cavalli, A.; Martella, V.; Tempesta, M.; Decaro, N.; Carmichael, L.E.; Buonavoglia, C. Canine Parvovirus (CPV) Vaccination: Comparison of Neutralizing Antibody Responses in Pups after Inoculation with CPV2 or CPV2b Modified Live Virus Vaccine. Clin. Diagn Lab. Immunol. 2001, 8, 612–615. [Google Scholar] [CrossRef] [Green Version]
- Battilani, M.; Modugno, F.; Mira, F.; Purpari, G.; Di Bella, S.; Guercio, A.; Balboni, A. Molecular Epidemiology of Canine Parvovirus Type 2 in Italy from 1994 to 2017: Recurrence of the CPV-2b Variant. BMC Vet. Res. 2019, 15, 393. [Google Scholar] [CrossRef] [PubMed]
- Ndiana, L.A.; Lanave, G.; Desario, C.; Berjaoui, S.; Alfano, F.; Puglia, I.; Fusco, G.; Colaianni, M.L.; Vincifori, G.; Camarda, A.; et al. Circulation of Diverse Protoparvoviruses in Wild Carnivores, Italy. Transbound. Emerg. Dis. 2021, 68, 2489–2502. [Google Scholar] [CrossRef] [PubMed]
- Mira, F.; Purpari, G.; Lorusso, E.; Di Bella, S.; Gucciardi, F.; Desario, C.; Macaluso, G.; Decaro, N.; Guercio, A. Introduction of Asian Canine Parvovirus in Europe through Dog Importation. Transbound. Emerg. Dis. 2018, 65, 16–21. [Google Scholar] [CrossRef]
- Mira, F.; Purpari, G.; Bella, S.D.; Colaianni, M.L.; Schirò, G.; Chiaramonte, G.; Gucciardi, F.; Pisano, P.; Lastra, A.; Decaro, N.; et al. Spreading of Canine Parvovirus Type 2c Mutants of Asian Origin in Southern Italy. Transbound. Emerg. Dis. 2019, 66, 2297. [Google Scholar] [CrossRef] [PubMed]
- Balboni, A.; Niculae, M.; Di Vito, S.; Urbani, L.; Terrusi, A.; Muresan, C.; Battilani, M. The Detection of Canine Parvovirus Type 2c of Asian Origin in Dogs in Romania Evidenced Its Progressive Worldwide Diffusion. BMC Vet. Res. 2021, 17, 206. [Google Scholar] [CrossRef] [PubMed]
- Ogbu, K.I.; Mira, F.; Purpari, G.; Nwosuh, C.; Loria, G.R.; Schirò, G.; Chiaramonte, G.; Tion, M.T.; Bella, S.D.; Ventriglia, G.; et al. Nearly Full-length Genome Characterization of Canine Parvovirus Strains Circulating in Nigeria. Transbound. Emerg. Dis. 2020, 67, 635. [Google Scholar] [CrossRef]
- Canuti, M.; Mira, F.; Sorensen, R.G.; Rodrigues, B.; Bouchard, É.; Walzthoni, N.; Hopson, M.; Gilroy, C.; Whitney, H.G.; Lang, A.S. Distribution and Diversity of Dog Parvoviruses in Wild, Free-Roaming and Domestic Canids of Newfoundland and Labrador, Canada. Transbound. Emerg. Dis. 2022. [Google Scholar] [CrossRef]
- Schirò, G.; Gambino, D.; Mira, F.; Vitale, M.; Guercio, A.; Purpari, G.; Antoci, F.; Licitra, F.; Chiaramonte, G.; La Giglia, M.; et al. Antimicrobial Resistance (AMR) of Bacteria Isolated from Dogs with Canine Parvovirus (CPV) Infection: The Need for a Rational Use of Antibiotics in Companion Animal Health. Antibiotics 2022, 11, 142. [Google Scholar] [CrossRef]
- Han, S.; Qi, B.; Zhang, X. A Retrospective Analysis on Phylogeny and Evolution of CPV Isolates in China. Asian J. Anim. Vet. Adv. 2011, 6, 1204–1213. [Google Scholar] [CrossRef]
- Yi, L.; Tong, M.; Cheng, Y.; Song, W.; Cheng, S. Phylogenetic Analysis of Canine Parvovirus VP2 Gene in China. Transbound. Emerg. Dis. 2016, 63, e262–e269. [Google Scholar] [CrossRef]
- Jiang, H.; Yu, Y.; Yang, R.; Zhang, S.; Wang, D.; Jiang, Y.; Yang, W.; Huang, H.; Shi, C.; Ye, L.; et al. Detection and Molecular Epidemiology of Canine Parvovirus Type 2 (CPV-2) Circulating in Jilin Province, Northeast China. Comp. Immunol. Microbiol. Infect. Dis. 2021, 74, 101602. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zhang, X.; Zhu, J.; Liao, L.; Bao, E. Molecular Epidemiological Survey of Canine Parvovirus Circulating in China from 2014 to 2019. Pathogens 2021, 10, 588. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, J.; Bi, Z.; Tan, Y.; Lv, L.; Zhao, H.; Xia, X.; Zhu, Y.; Wang, Y.; Qian, J. Molecular Epidemiology and Genetic Evolution of Canine Parvovirus in East China, during 2018–2020. Infect. Genet. Evol. 2021, 90, 104780. [Google Scholar] [CrossRef]
- Wang, J.; Lin, P.; Zhao, H.; Cheng, Y.; Jiang, Z.; Zhu, H.; Wu, H.; Cheng, S. Continuing Evolution of Canine Parvovirus in China: Isolation of Novel Variants with an Ala5Gly Mutation in the VP2 Protein. Infect. Genet. Evol. 2016, 38, 73–78. [Google Scholar] [CrossRef]
- Carrino, M.; Tassoni, L.; Campalto, M.; Cavicchio, L.; Mion, M.; Corrò, M.; Natale, A.; Beato, M.S. Molecular Investigation of Recent Canine Parvovirus-2 (CPV-2) in Italy Revealed Distinct Clustering. Viruses 2022, 14, 917. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.D.S.; Palermo, L.M.; Hafenstein, S.L.; Parrish, C.R. Different Mechanisms of Antibody-Mediated Neutralization of Parvoviruses Revealed Using the Fab Fragments of Monoclonal Antibodies. Virology 2007, 361, 283–293. [Google Scholar] [CrossRef]
- Ohshima, T.; Mochizuki, M. Evidence for Recombination between Feline Panleukopenia Virus and Canine Parvovirus Type 2. J. Vet. Med. Sci. 2009, 71, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yue, C.; Yang, Z.; Li, Y.; Zhang, D.; Zhang, J.; Yang, S.; Shen, Q.; Su, X.; Qi, D.; et al. Viral Metagenomics Unveiled Extensive Communications of Viruses within Giant Pandas and Their Associated Organisms in the Same Ecosystem. Sci. Total Environ. 2022, 820, 153317. [Google Scholar] [CrossRef]
- Li, X.; Wu, H.; Wang, L.; Spibey, N.; Liu, C.; Ding, H.; Liu, W.; Liu, Y.; Tian, K. Genetic Characterization of Parvoviruses in Domestic Cats in Henan Province, China. Transbound. Emerg. Dis. 2018, 65, 1429–1435. [Google Scholar] [CrossRef]
- Tang, Y.; Tang, N.; Zhu, J.; Wang, M.; Liu, Y.; Lyu, Y. Molecular Characteristics and Genetic Evolutionary Analyses of Circulating Parvoviruses Derived from Cats in Beijing. BMC Vet. Res. 2022, 18, 195. [Google Scholar] [CrossRef]
- Hoelzer, K.; Shackelton, L.A.; Parrish, C.R.; Holmes, E.C. Phylogenetic Analysis Reveals the Emergence, Evolution and Dispersal of Carnivore Parvoviruses. J. Gen. Virol. 2008, 89, 2280–2289. [Google Scholar] [CrossRef] [PubMed]
- Purpari, G.; Mira, F.; Di Bella, S.; Di Pietro, S.; Giudice, E.; Guercio, A. Investigation on Circulation in Dogs from Sicily (Italy) by Biomolecular Assay. Acta Vet. 2018, 68, 80–94. [Google Scholar] [CrossRef]
- Mira, F.; Purpari, G.; Di Bella, S.; Vicari, D.; Schirò, G.; Di Marco, P.; Macaluso, G.; Battilani, M.; Guercio, A. Update on Canine Distemper Virus (CDV) Strains of Arctic-like Lineage Detected in Dogs in Italy. Vet. Ital. 2018, 54, 225–236. [Google Scholar] [CrossRef]
- Pratelli, A.; Martella, V.; Elia, G.; Decaro, N.; Aliberti, A.; Buonavoglia, D.; Tempesta, M.; Buonavoglia, C. Variation of the Sequence in the Gene Encoding for Transmembrane Protein M of Canine Coronavirus (CCV). Mol. Cell. Probes 2001, 15, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.L.; Huang, G.; Qiu, W.; Zhong, Z.H.; Xia, X.Z.; Yin, Z. Detection and Differentiation of CAV-1 and CAV-2 by Polymerase Chain Reaction. Vet. Res. Commun. 2001, 25, 77–84. [Google Scholar] [CrossRef]
- Bodnar, L.; Lorusso, E.; Di Martino, B.; Catella, C.; Lanave, G.; Elia, G.; Bányai, K.; Buonavoglia, C.; Martella, V. Identification of a Novel Canine Norovirus. Infect. Genet. Evol. 2017, 52, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.M.; Kerin, T.; Hull, J.; McCaustland, K.; Gentsch, J. Enhancement of Detection and Quantification of Rotavirus in Stool Using a Modified Real-Time RT-PCR Assay. J. Med. Virol. 2008, 80, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- Pérez, R.; Calleros, L.; Marandino, A.; Sarute, N.; Iraola, G.; Grecco, S.; Blanc, H.; Vignuzzi, M.; Isakov, O.; Shomron, N.; et al. Phylogenetic and Genome-Wide Deep-Sequencing Analyses of Canine Parvovirus Reveal Co-Infection with Field Variants and Emergence of a Recent Recombinant Strain. PLoS ONE 2014, 9, e111779. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and Analysis of Recombination Patterns in Virus Genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| CPV-2 Sequence | NS1 | VP2 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variant | Strain | Country | Year | Acc. Nr. | 60 | 544 | 545 | 588 | 630 | 5 | 267 | 297 | 324 | 370 | 426 | 440 |
| CPV-2 1 | CPV-b | USA | 1978 | M38245 | I | Y | E | S | L | A | F | S | Y | Q | N | T |
| CPV-2a 1 | 43-91 | Italy 5 | 1997 | MF177224 | - | F | - | - | - | - | - | A | - | - | N | A |
| CPV-2b 1 | 1-00 | Italy 5 | 1999 | MF177226 | - | F | - | - | - | - | - | - | - | - | D | - |
| CPV-2b 1,2 | IZSSI_PA18546/18 | Italy 5 | 2018 | MT981023 | - | F | - | - | - | - | - | A | - | - | D | - |
| CPV-2c 1 | 485-09 | Italy 5 | 2009 | MF177228 | - | - | - | - | - | - | - | A | - | - | E | - |
| CPV-2a 1,3 | CPV/CN/LN1/2014 | China 6 | 2014 | KR002800 | V | F | V | - | P | - | Y | A | I | - | N | A |
| CPV-2b 1 | Canine/China/19/2017 | China 6 | 2017 | MH476588 | - | - | - | - | - | - | Y | A | I | - | D | A |
| CPV-2c 1 | Canine/China/12/2017 | China 6 | 2017 | MH476581 | V | F | V | - | P | G | Y | A | I | R | E | - |
| CPV-2b 4 | CPV-2b_IZSSI_2022PA2773 | Italy | 2022 | ON677437 | V | F | V | N | P | G | Y | A | I | R | D | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schirò, G.; Mira, F.; Canuti, M.; Vullo, S.; Purpari, G.; Chiaramonte, G.; Di Bella, S.; Cannella, V.; Randazzo, V.; Castronovo, C.; et al. Identification and Molecular Characterization of a Divergent Asian-like Canine Parvovirus Type 2b (CPV-2b) Strain in Southern Italy. Int. J. Mol. Sci. 2022, 23, 11240. https://doi.org/10.3390/ijms231911240
Schirò G, Mira F, Canuti M, Vullo S, Purpari G, Chiaramonte G, Di Bella S, Cannella V, Randazzo V, Castronovo C, et al. Identification and Molecular Characterization of a Divergent Asian-like Canine Parvovirus Type 2b (CPV-2b) Strain in Southern Italy. International Journal of Molecular Sciences. 2022; 23(19):11240. https://doi.org/10.3390/ijms231911240
Chicago/Turabian StyleSchirò, Giorgia, Francesco Mira, Marta Canuti, Stefano Vullo, Giuseppa Purpari, Gabriele Chiaramonte, Santina Di Bella, Vincenza Cannella, Vincenzo Randazzo, Calogero Castronovo, and et al. 2022. "Identification and Molecular Characterization of a Divergent Asian-like Canine Parvovirus Type 2b (CPV-2b) Strain in Southern Italy" International Journal of Molecular Sciences 23, no. 19: 11240. https://doi.org/10.3390/ijms231911240
APA StyleSchirò, G., Mira, F., Canuti, M., Vullo, S., Purpari, G., Chiaramonte, G., Di Bella, S., Cannella, V., Randazzo, V., Castronovo, C., Vicari, D., & Guercio, A. (2022). Identification and Molecular Characterization of a Divergent Asian-like Canine Parvovirus Type 2b (CPV-2b) Strain in Southern Italy. International Journal of Molecular Sciences, 23(19), 11240. https://doi.org/10.3390/ijms231911240