
Effects of Drought and Host on the Growth of Santalum album Seedlings in Pot Culture


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
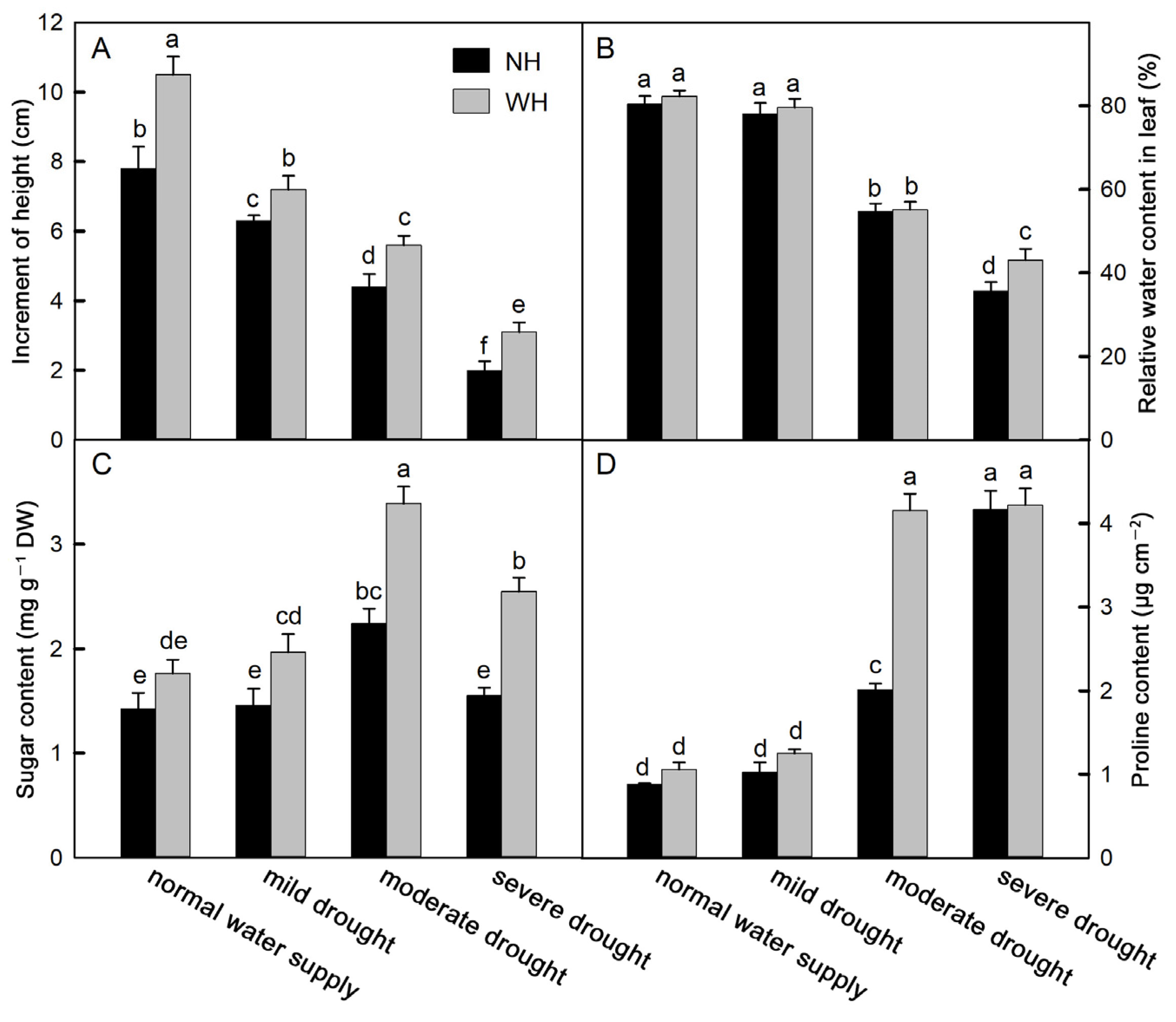
2.1. Increases in Plant Height and Proline, Soluble Sugar, and Leaf Relative Water Contents
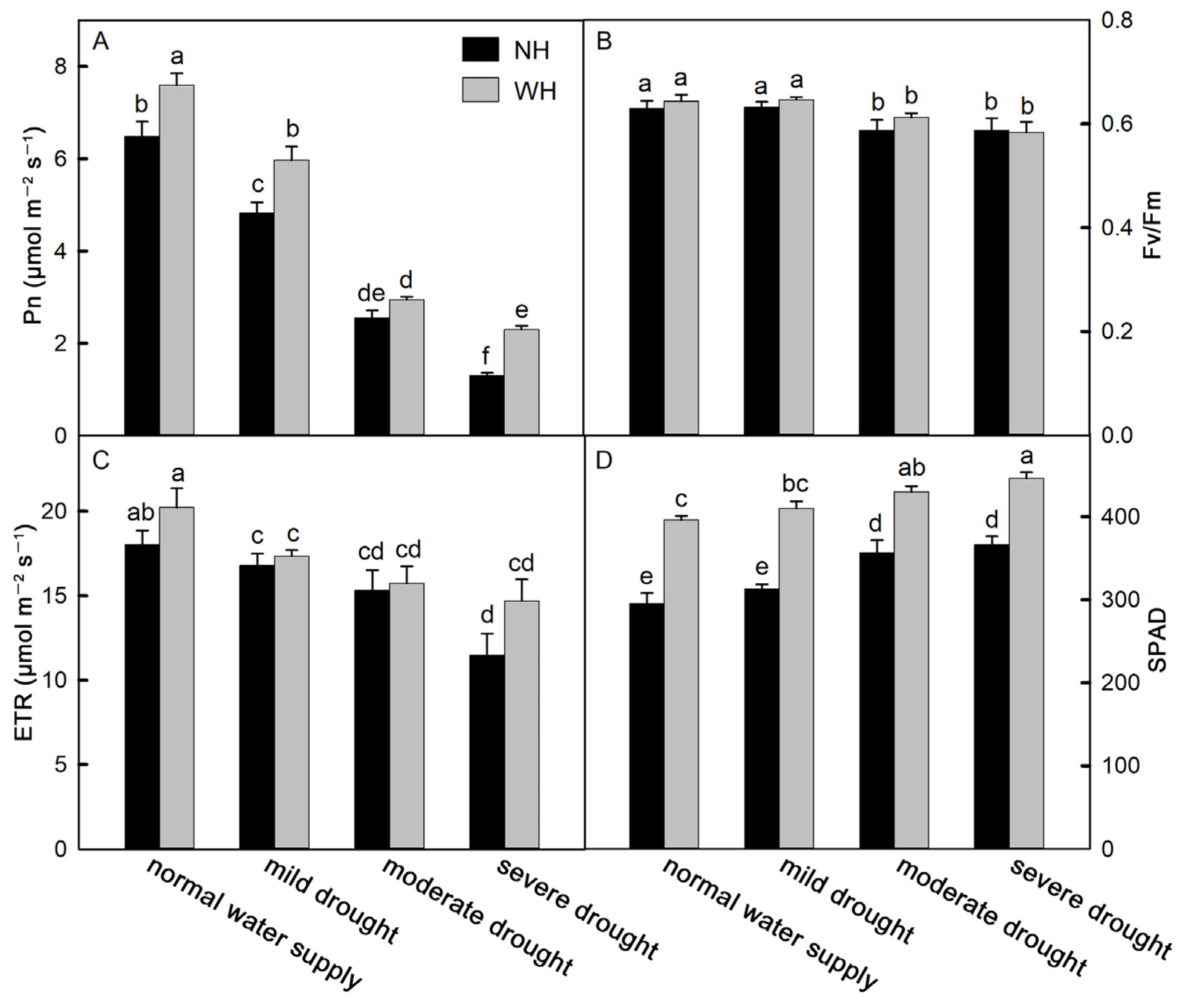
2.2. Photosynthetic Capacity
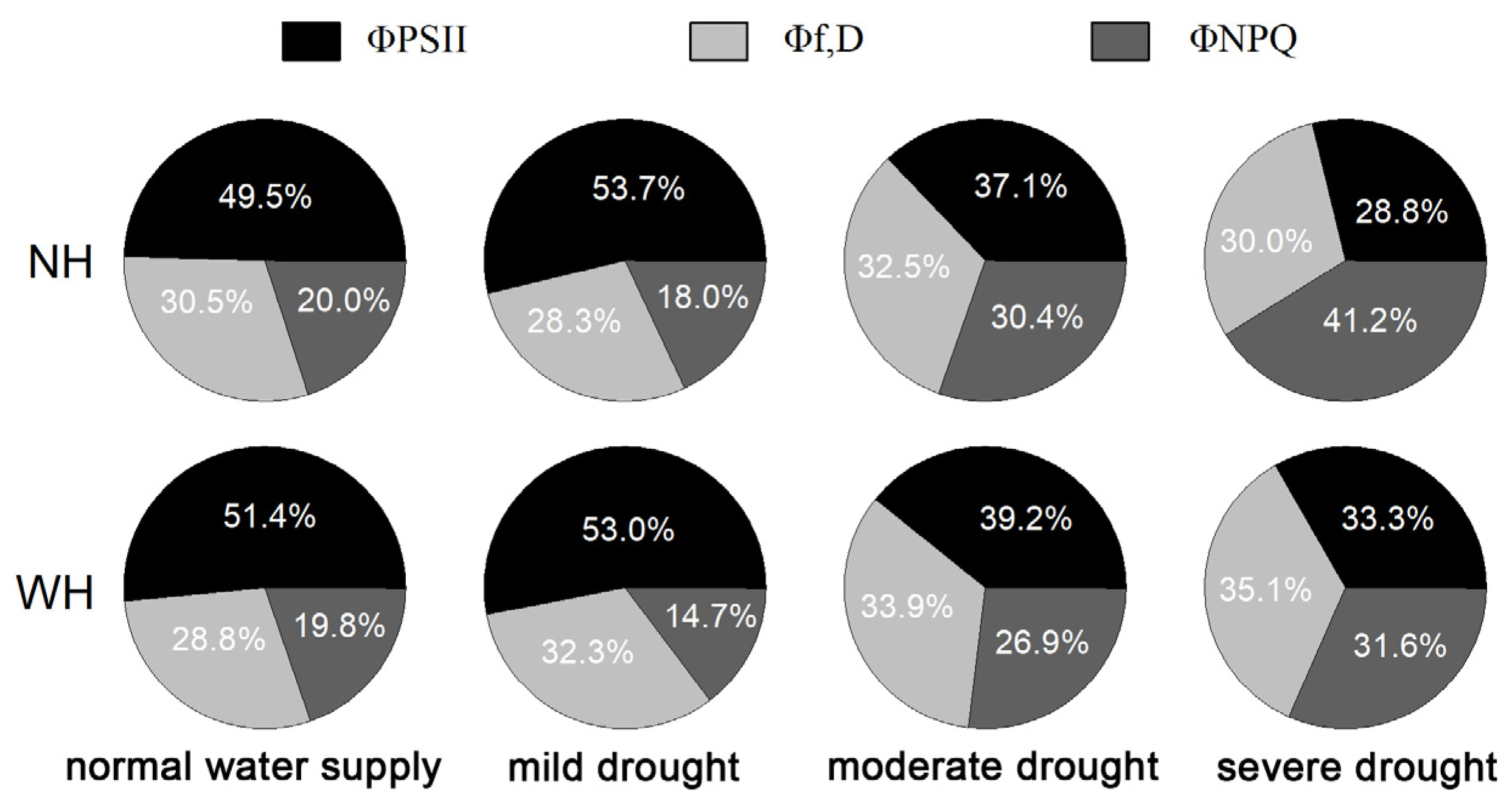
2.3. Light Energy Distribution
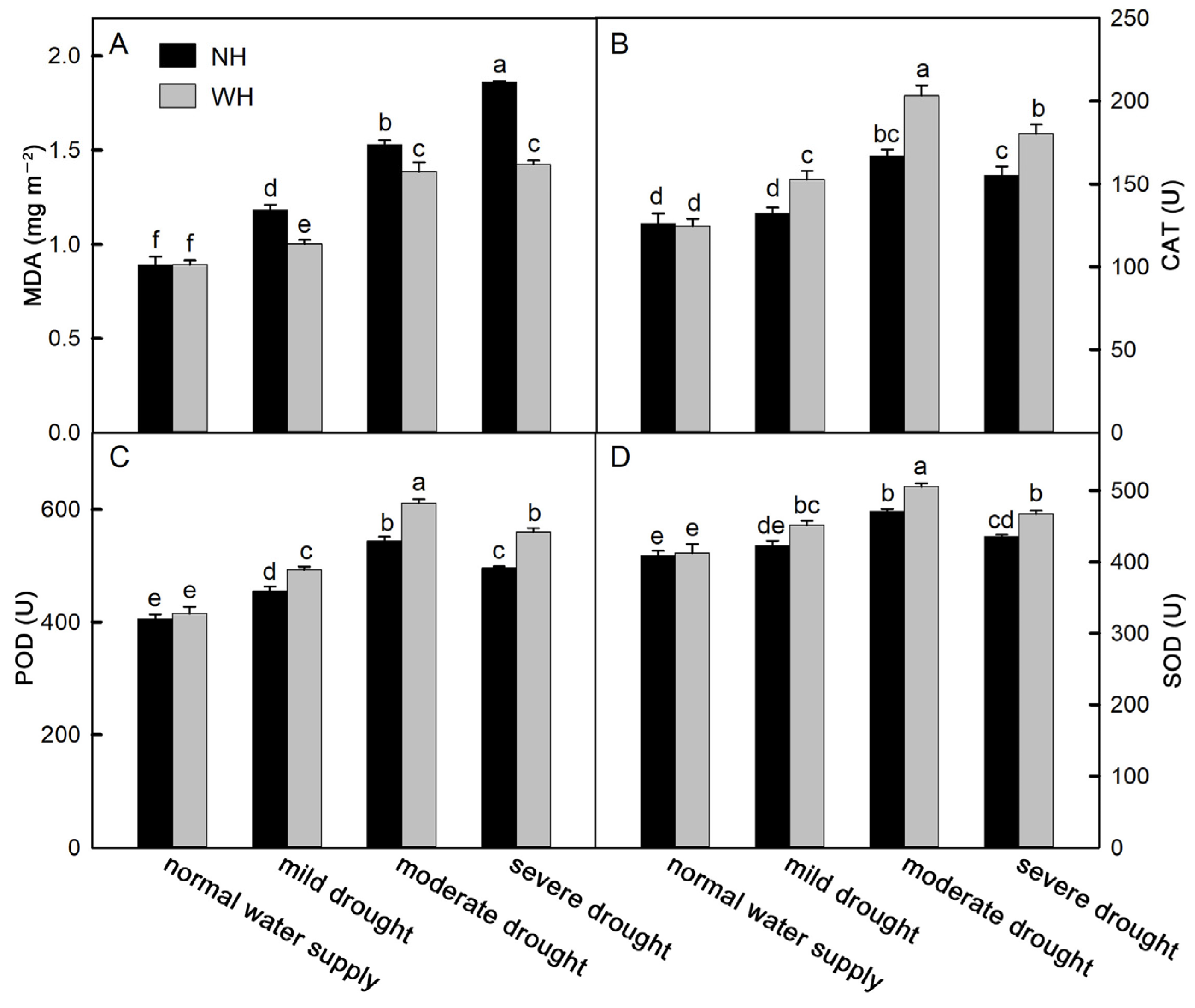
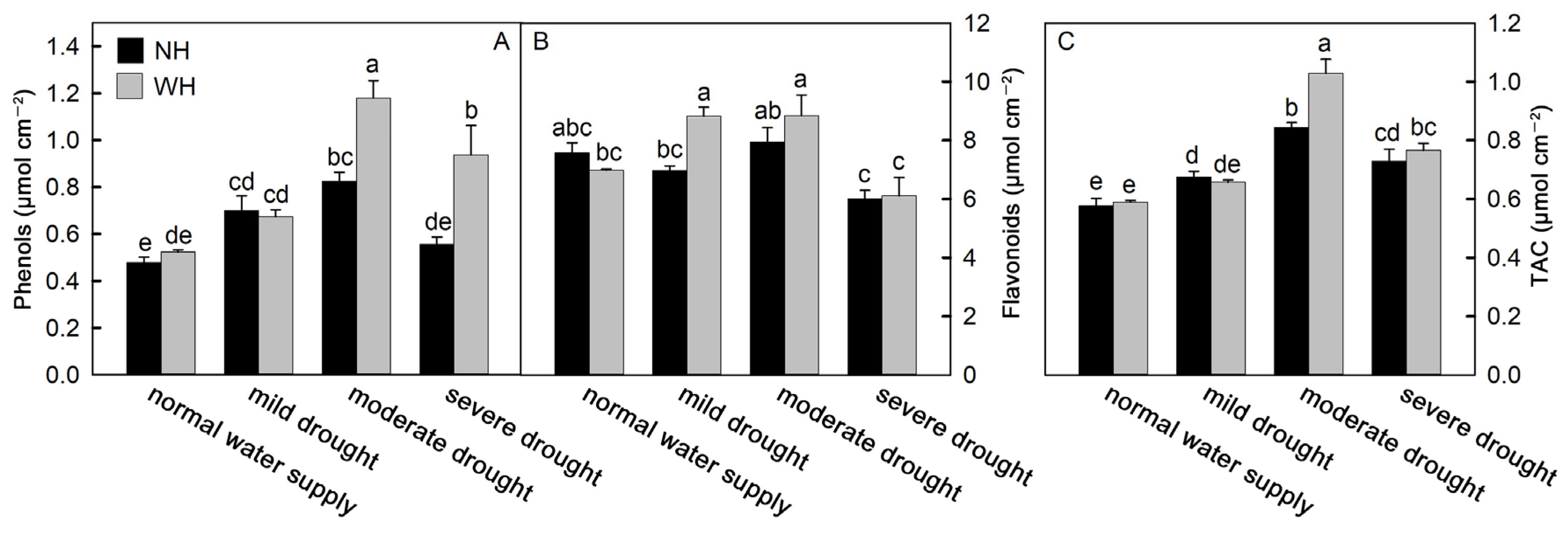
2.4. Antioxidant Capacity
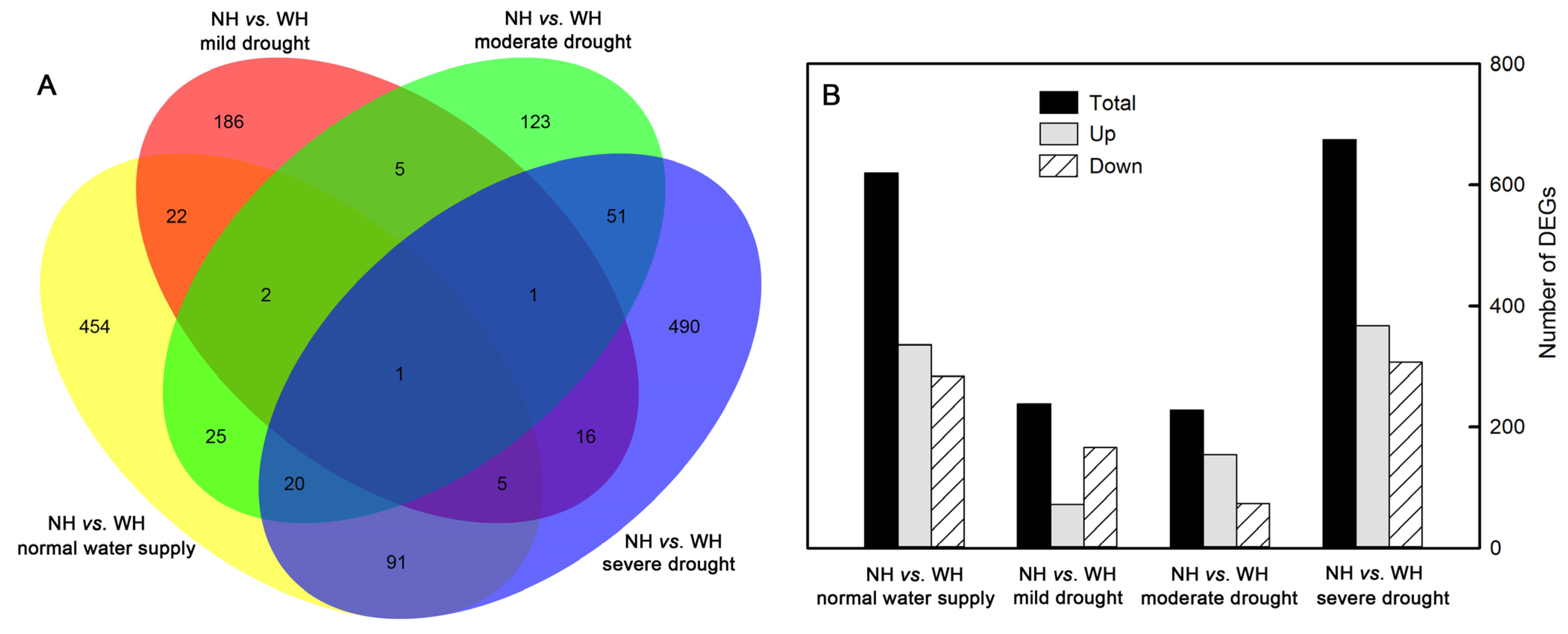
2.5. Number of Differentially Expressed Genes
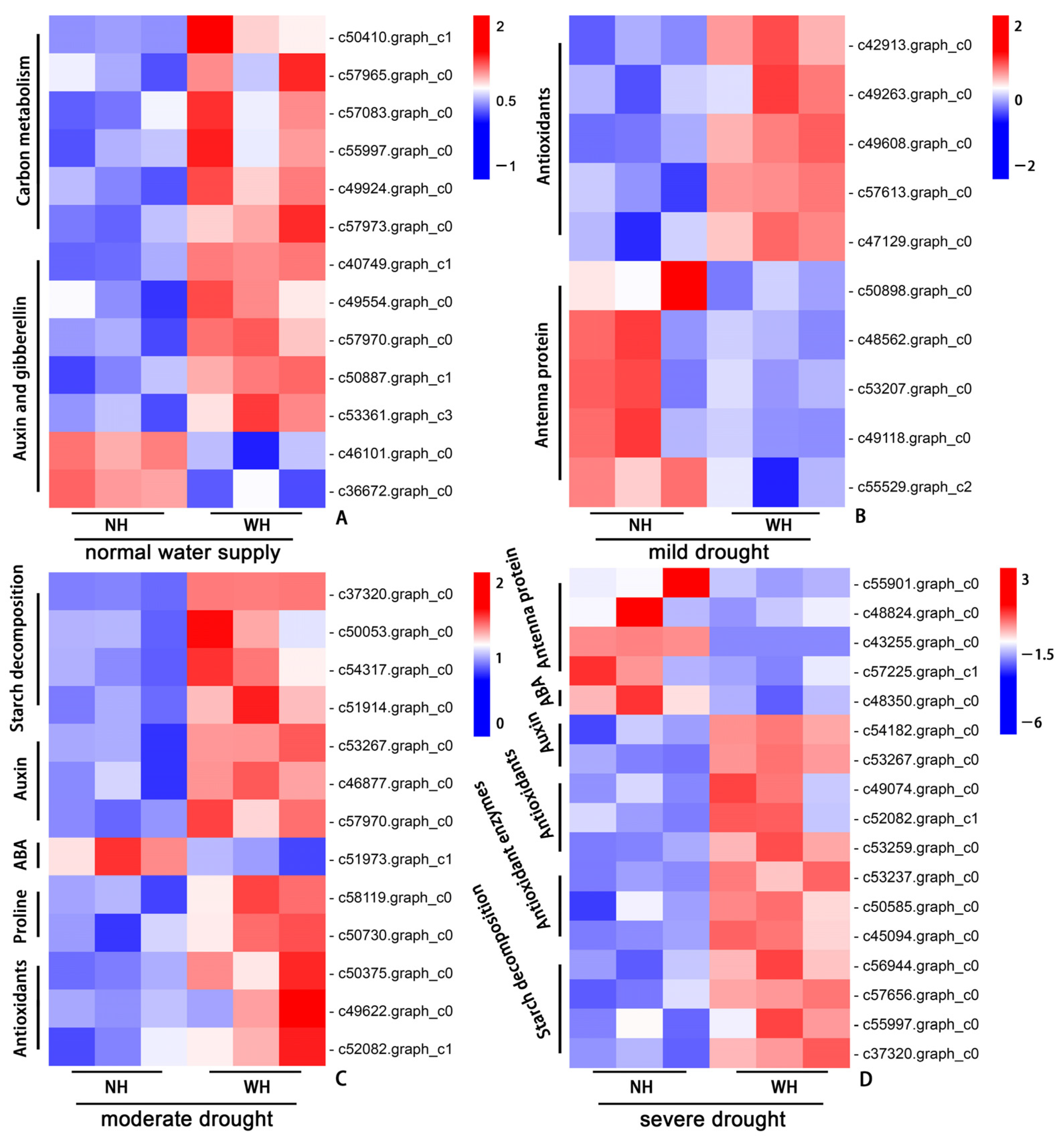
2.6. Gene Expressions between Plants Grown without Host and Those Grown with Host under Different Moisture Conditions
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Leaf Water Contents
4.3. Determinations of Net Photosynthetic Rates, Chlorophyll Fluorescence Parameters and Light Energy Distributions
4.4. Soluble Sugar and Proline Content
4.5. Activities of Catalase, Superoxide Dismutase and Peroxidase
4.6. Determinations of Malondialdehyde, Flavonoids, Total Phenols and Total Antioxidant Capacity
4.7. Total RNA Extraction and RNA Sequencing
4.8. Functional Annotation and Identification of Differentially Expressed Genes
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Ma, G.H.; Bunn, E.; Zhang, J.F.; Wu, G.J. Evidence of dichogamy in Santalum album L. J. Integr. Plant. Biol 2006, 48, 300–306. [Google Scholar] [CrossRef]
- Arasada, B.L.; Bommareddy, A.; Zhang, X.Y.; Bremmon, K.; Dwivedi, C. Effects of alpha-santalol on proapoptotic caspases and p53 expression in UVB irradiated mouse skin. Anticancer Res. 2008, 28, 129–132. [Google Scholar]
- Bhatia, S.P.; McGinty, D.; Letizia, C.S.; Api, A.M. Fragrance material review on alpha-santalol. Food Chem. Toxicol. 2008, 46, S267–S269. [Google Scholar] [CrossRef]
- Burdock, G.A.; Carabin, L.G. Safety assessment of sandalwood oil (Santalum album L.). Food Chem. Toxicol. 2008, 46, 421–432. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, X.; da Silva, J.A.T.; Liang, K.; Deng, R.; Ma, G. Ontogenesis of the collapsed layer during haustorium development in the root hemi-parasite Santalum album Linn. Plant. Biol. 2014, 16, 282–290. [Google Scholar] [CrossRef]
- Deepa, P.; Yusuf, A. Influence of different host associations on glutamine synthetase activity and ammonium transporter in Santalum album L. Physiol. Mol. Biol. Pla. 2016, 22, 331–340. [Google Scholar] [CrossRef]
- Radomiljac, A.M.; McComb, J.A.; Shea, S.R. Field establishment of Santalum album L.-the effect of the time of introduction of a pot host (Alternanthera nana R. Br.). Forest Ecol. Manag. 1998, 111, 107–118. [Google Scholar] [CrossRef]
- Balasubramanian, A.; Prasath, C.N.H.; Radhakrishnan, S.; Sivaprakash, M. Host-specific influence on early growth and physiological attributes of sandal (Santalum album) grown in farmlands. J. Environ. Biol. 2021, 42, 1162–1167. [Google Scholar] [CrossRef]
- Meng, S.; Ma, H.B.; Li, Z.S.; Yang, F.C.; Wang, S.K.; Lu, J.K. Impacts of nitrogen on physiological interactions of the hemiparasitic Santalum album and its N-2-fixing host Dalbergia odorifera. Trees-Struct. Funct. 2021, 35, 1039–1051. [Google Scholar] [CrossRef]
- Ramegowda, V.; Senthil-Kumar, M. The interactive effects of simultaneous biotic and abiotic stresses on plants: Mechanistic understanding from drought and pathogen combination. J. Plant. Physiol. 2015, 176, 47–54. [Google Scholar] [CrossRef]
- De Vries, F.T.; Griffiths, R.I.; Knight, C.G.; Nicolitch, O.; Williams, A. Harnessing rhizosphere microbiomes for drought-resilient crop production. Science 2020, 368, 270–274. [Google Scholar] [CrossRef]
- Soares-Cordeiro, A.S.; Carmo-Silva, A.E.; da Silva, A.; da Silva, J.; Keys, A.J.; Arrabaca, M.C. Effects of rapidly imposed water deficit on photosynthetic parameters of three C-4 grasses. Photosynthetica 2009, 47, 304–308. [Google Scholar] [CrossRef]
- Xu, X.; Peng, G.Q.; Wu, C.C.; Korpelainen, H.; Li, C.Y. Drought inhibits photosynthetic capacity more in females than in males of Populus cathayana. Tree Physiol. 2008, 28, 1751–1759. [Google Scholar] [CrossRef]
- Barber, J.; Andersson, B. Too Much of a Good Thing-Light Can Be Bad for Photosynthesis. Trends Biochem. Sci. 1992, 17, 61–66. [Google Scholar] [CrossRef]
- Murata, N.; Allakhverdiev, S.I.; Nishiyama, Y. The mechanism of photoinhibition in vivo: Re-evaluation of the roles of catalase, alpha-tocopherol, non-photochemical quenching, and electron transport. Bba-Bioenergetics 2012, 1817, 1127–1133. [Google Scholar] [CrossRef]
- Zhang, T.J.; Zheng, J.; Yu, Z.C.; Gu, X.Q.; Tian, X.S.; Peng, C.L.; Chow, W.S. Variations in photoprotective potential along gradients of leaf development and plant succession in subtropical forests under contrasting irradiances. Environ. Exp. Bot. 2018, 154, 23–32. [Google Scholar] [CrossRef]
- Da Silva, J.A.T.; Kher, M.M.; Soner, D.; Page, T.; Zhang, X.H.; Nataraj, M.; Ma, G.H. Sandalwood: Basic biology, tissue culture, and genetic transformation. Planta 2016, 243, 847–887. [Google Scholar] [CrossRef]
- Rashkow, E.D. Perfumed the axe that laid it low: The endangerment of sandalwood in southern India. Indian Econ. Soc. Hist. 2014, 51, 41–70. [Google Scholar] [CrossRef]
- Kaur, N.; Gupta, A.K. Signal transduction pathways under abiotic stresses in plants. Curr. Sci. India 2005, 88, 1771–1780. [Google Scholar]
- Zhang, Q.L.; Huang, J.D.; Ke, W.Q.; Cai, M.L.; Chen, G.X.; Peng, C.L. Responses of Sphagneticola trilobata, Sphagneticola calendulacea and their hybrid to drought stress. Int. J. Mol. Sci. 2021, 22, 11288. [Google Scholar] [CrossRef]
- Ouyang, Y.; Zhang, X.H.; Chen, Y.L.; da Silva, J.A.T.; Ma, G.H. Growth, photosynthesis and haustorial development of semiparasitic Santalum album L. penetrating into roots of three hosts: A comparative study. Trees-Struct. Funct. 2016, 30, 317–328. [Google Scholar] [CrossRef]
- Sadak, M.S.; Abdalla, A.M.; Abd Elhamid, E.M.; Ezzo, M.I. Role of melatonin in improving growth, yield quantity and quality of Moringa oleifera L. plant under drought stress. Bull. Natl. Res. Cent. 2020, 44, 1. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, F.; Suzuki, T.; Osakabe, Y.; Betsuyaku, S.; Kondo, Y.; Dohmae, N.; Fukuda, H.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A small peptide modulates stomatal control via abscisic acid in long-distance signalling. Nature 2018, 556, 235–238. [Google Scholar] [CrossRef]
- Guadagno, C.R.; Ewers, B.E.; Speckman, H.N.; Aston, T.L.; Huhn, B.J.; DeVore, S.B.; Ladwig, J.T.; Strawn, R.N.; Weinig, C. Dead or Alive? Using Membrane Failure and Chlorophyll a Fluorescence to Predict Plant Mortality from Drought. Plant Physiol. 2017, 175, 223–234. [Google Scholar] [CrossRef]
- Sun, L.Y.; Li, X.N.; Wang, Z.S.; Sun, Z.W.; Zhu, X.C.; Liu, S.Q.; Song, F.B.; Liu, F.L.; Wang, Y.J. Cold Priming Induced Tolerance to Subsequent Low Temperature Stress is Enhanced by Melatonin Application during Recovery in Wheat. Molecules 2018, 23, 1091. [Google Scholar] [CrossRef]
- Wu, H.; Fu, B.; Sun, P.P.; Xiao, C.; Liu, J.H. A NA.AC Transcription Factor Represses Putrescine Biosynthesis and Affects Drought Tolerance. Plant Physiol. 2016, 172, 1532–1547. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Zhou, G.S. Exploring the effects of water on vegetation change and net primary productivity along the IGBP Northeast China Transect. Environ. Earth Sci. 2011, 62, 1481–1490. [Google Scholar] [CrossRef]
- Bashir, W.; Anwar, S.; Zhao, Q.; Hussain, I.; Xie, F. Interactive effect of drought and cadmium stress on soybean root morphology and gene expression. Ecotox. Environ. Safe. 2019, 175, 90–101. [Google Scholar] [CrossRef]
- Khan, M.S.; Ahmad, D.; Khan, M.A. Utilization of genes encoding osmoprotectants in transgenic plants for enhanced abiotic stress tolerance. Electron. J. Biotechnol. 2015, 18, 257–266. [Google Scholar] [CrossRef]
- Alharby, H.F.; Fahad, S. Melatonin application enhances biochar efficiency for drought tolerance in maize varieties: Modifications in physio-biochemical machinery. Agron. J. 2020, 112, 2826–2847. [Google Scholar] [CrossRef]
- Yasmin, H.; Rashid, U.; Hassan, M.N.; Nosheen, A.; Naz, R.; Ilyas, N.; Sajjad, M.; Azmat, A.; Alyemeni, M.N. Volatile organic compounds produced by Pseudomonas pseudoalcaligenes alleviated drought stress by modulating defense system in maize (Zea mays L.). Physiol Plant. 2021, 172, 896–911. [Google Scholar] [CrossRef]
- Zhang, X.B.; Lei, L.; Lai, J.S.; Zhao, H.M.; Song, W.B. Effects of drought stress and water recovery on physiological responses and gene expression in maize seedlings. Bmc Plant. Biol. 2018, 18, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant. Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Barrero, J.M.; Rodriguez, P.L.; Quesada, V.; Piqueras, P.; Ponce, M.R.; Micol, J.L. Both abscisic acid (ABA)-dependent and ABA-independent pathways govern the induction of NCED3, AAO3 and ABA1 in response to salt stress. Plant. Cell Environ. 2006, 29, 2000–2008. [Google Scholar] [CrossRef]
- Liu, X.J.; Xu, D.P.; Yang, Z.J.; Zhang, N.N.; Pan, L.J. Investigation of Exogenous Benzyladenine on Growth, Biochemical Composition, Photosynthesis and Antioxidant Activity of Indian Sandalwood (Santalum album L.) Seedlings. J. Plant. Growth Regul. 2018, 37, 1148–1158. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant. Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef]
- Kuhlgert, S.; Austic, G.; Zegarac, R.; Osei-Bonsu, I.; Hoh, D.; Chilvers, M.I.; Roth, M.G.; Bi, K.; TerAvest, D.; Weebadde, P.; et al. MultispeQ Beta: A tool for large-scale plant phenotyping connected to the open PhotosynQ network. Roy. Soc. Open Sci. 2016, 3, 160592. [Google Scholar] [CrossRef]
- Yang, F.; Wang, Y.; Wang, J.; Deng, W.Q.; Liao, L.; Li, M. Different eco-physiological responses between male and female Populus deltoides clones to waterlogging stress. Forest Ecol. Manag. 2011, 262, 1963–1971. [Google Scholar] [CrossRef]
- Sun, Z.Y.; Chen, Y.Q.; Schaefer, V.; Liang, H.M.; Li, W.H.; Huang, S.Q.; Peng, C.L. Responses of the Hybrid between Sphagneticola trilobata and Sphagneticola calendulacea to Low Temperature and Weak Light Characteristic in South China. Sci. Rep. 2015, 5, 16906. [Google Scholar] [CrossRef]
- Heimler, D.; Vignolini, P.; Dini, M.G.; Romani, A. Rapid tests to assess the antioxidant activity of Phaseolus vulgaris L. dry beans. J. Agr. Food Chem. 2005, 53, 3053–3056. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Saha, M.R.; Hasan, S.M.R.; Akter, R.; Hossain, M.M.; Alam, M.S.; Alam, M.A.; Mazumder, M.E.H. In vitro free radical scavenging activity of methanol extract of the leaves of Mimusops elengi Linn. Bangladesh J. Vet. Med. 2008, 6, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.J.; Zheng, J.; Yu, Z.C.; Huang, X.D.; Zhang, Q.L.; Tian, X.S.; Peng, C.L. Functional characteristics of phenolic compounds accumulated in young leaves of two subtropical forest tree species of different successional stages. Tree Physiol. 2018, 38, 1486–1501. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Liu, X.; Xu, D.; Hong, Z.; Zhang, N.; Cui, Z. Effects of Drought and Host on the Growth of Santalum album Seedlings in Pot Culture. Int. J. Mol. Sci. 2022, 23, 11241. https://doi.org/10.3390/ijms231911241
Zhang Q, Liu X, Xu D, Hong Z, Zhang N, Cui Z. Effects of Drought and Host on the Growth of Santalum album Seedlings in Pot Culture. International Journal of Molecular Sciences. 2022; 23(19):11241. https://doi.org/10.3390/ijms231911241
Chicago/Turabian StyleZhang, Qilei, Xiaojin Liu, Daping Xu, Zhou Hong, Ningnan Zhang, and Zhiyi Cui. 2022. "Effects of Drought and Host on the Growth of Santalum album Seedlings in Pot Culture" International Journal of Molecular Sciences 23, no. 19: 11241. https://doi.org/10.3390/ijms231911241
APA StyleZhang, Q., Liu, X., Xu, D., Hong, Z., Zhang, N., & Cui, Z. (2022). Effects of Drought and Host on the Growth of Santalum album Seedlings in Pot Culture. International Journal of Molecular Sciences, 23(19), 11241. https://doi.org/10.3390/ijms231911241