
Roles of Exosomes in Chronic Rhinosinusitis: A Systematic Review



Abstract
:1. Introduction
2. Methods
2.1. Methods Literature Retrieval
2.2. Data Extraction
2.3. Assessment of Quality of Studies
3. Results and Discussion
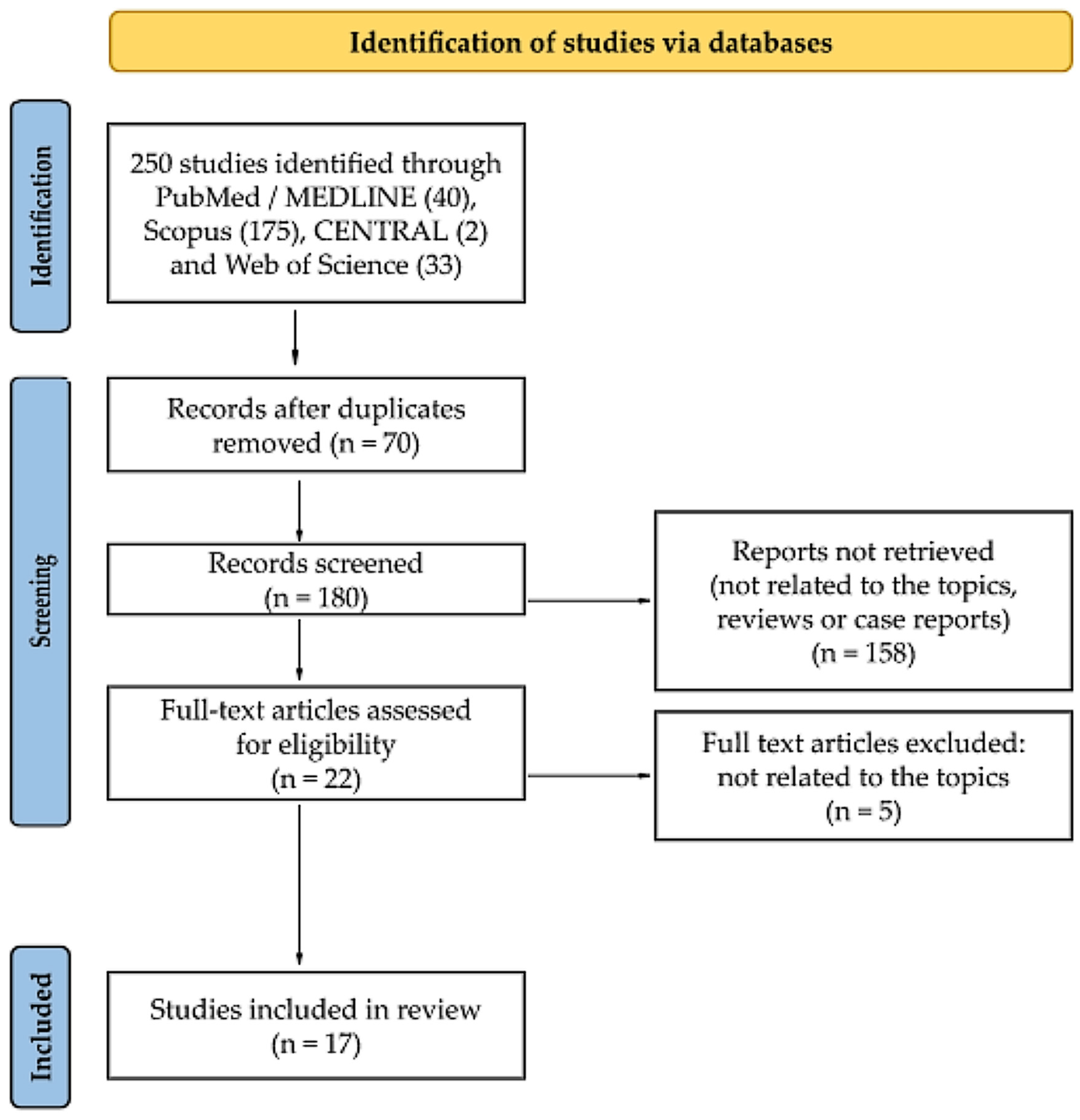
3.1. Search Results
3.2. Study Characteristics and Study Quality
3.3. Method of Mucus and NLF Collection
3.4. Overview of EV Isolation Methodologies and Characterisation

{kind=link}
{kind=link}
| Study | Isolation Method | Exosome Characterization | Exosome Markers | |
|---|---|---|---|---|
| Positive | Negative | |||
| Lasser (2016) [8] | UC described by Lasser [10] 1. Centrifugation at 300× g for 10 min at 4 °C. 2. Storage at −80 °C. 3. Thawing and transferring to ultracentrifuge tubes filled with PBS. 4. Centrifugation at 16,500× g for 20 min at 4 °C. 5. Filtration 0.2-µM filter. 6. Ultracentrifugation at 120,000× g for 70 min at 4 °C. | FCM, WB | CD9, CD14, CD63, TSG101, NOS2, S100A8 | Calnexin |
| Nocera (2017) [19] | UC described by Thery [9] 1. Centrifugation at 1500× g for 30 min at 4 °C. 2. Dilution in 150 µL of PBS with Protease Inhibitor Cocktail. 3. Centrifugation at 12,000× g for 45 min at 4 °C. 4. Suspending in 4.5 mL of PBS in polypropylene tubes. 5. Ultracentrifugation at 110,000× g for 2 h at 4 °C. 6. Resuspend in 4.5 mL of PBS. 7. Filtration 0.22-µM filter. 8. Centrifugation at 110,000× g for 70 min at 4 °C. | ELISA, TEM | CD9, CD63 | NR |
| Zhang (2018) [25] | UC described by Valadi [33] and Romancino [34] 1. Centrifugation at 6000× g for 30 min at 4 °C. 2. Centrifugation at 10,000× g for 60 min at 4 °C. 3. Filtration 0.2-µM filter and qEV size-exclusion columns. 4. Ultracentrifugation at 100,000× g for 60 min at 4 °C. | TEM, NTA, WB | CD9, CD63, β-actin | NR |
| Mueller (2018) [17] | UC described by Thery [9] (see above) | SOMAscan | NR | NR |
| Mueller (2019) [15] | UC described by Thery [9] (see above) | WB, SOMAscan | GAPDH | NR |
| Mueller (2019) [16] | UC described by Thery [9] (see above) | WB | GAPDH | NR |
| Miyake (2019) [14] | UC described by Thery [9] (see above) | ELISA | NR | NR |
| Zhou (2020) [27] | 1. Centrifugation at 1000× g for 10 min. 2. Centrifugation at 16,500× g for 30 min at 4 °C. 3. Ultracentrifugation at 100,000× g for 2 h at 4 °C. 4. Resuspend in 4.5 mL of PBS. 5. Filtration 0.2-µM filter. | NTA, TEM, WB | CD9, TSG101, GAPDH | NR |
| Workman (2020) [24] | UC described by Thery [9] (see above) | SOMAscan | NR | NR |
| Mueller (2020) [18] | UC described by Thery [9] (see above) | SOMAscan | NR | NR |
| Shin (2020) [22] | 1. Centrifugation at 200× g for 10 min at 4 °C. 2. Centrifugation at 2000× g for 20 min at 4 °C. 3. Centrifugation at 10,000× g for 30 min at 4 °C. 4. Filtration 0.22-µM filter 5. Centrifugation at 100,000× g for 70 min. 6. Washing twice in PBS by centrifugation at 100,000× g for 70 min. | NR | NR | NR |
| Zhang (2020) [26] | UC described by Valadi [33] and Romancino [34] (see above) | WB, TEM, NTA | CD9, CD63, ALIX, TSG101 | GM130 |
| Cha (2021) [12] | 1. Centrifugation at 300× g for 10 min at 4 °C. 2. Centrifugation at 2000× g for 20 min. 3. Centrifugation at 16,500× g for 50 min. 4. Centrifugation at 120,000× g for 60 min. 5. Vortexing with 1 mL of ice-cold PBS. | FCM, TEM | CD9, CD63 | NR |
| Wang (2021) [23] | UC described by Lasser [10] with minor modification (see above) | NTA, WB, TEM | CD9, CD63, Annexin V, TSG101 | NR |
| Shimizu (2022) [21] | 1. Centrifugation at 1500× g for 5 min. 2. Centrifugation at 14,000× g for 5 min. 3. Centrifugation at 100,000× g for 60 min. | ELISA, NTA, TEM | CD9, CD63 | NR |
| He (2022) [13] | 1. Centrifugation at 5000× g for 20 min below 4 °C. 2. Filtration 0.45-µM membrane. 3. Using Exosome Isolation Kit (Abace Biotechnology, Beijing, China). | NTA, WB, TEM | CD9, CD63, TSG101, ALIX | NR |
| Nocera (2022) [20] | UC described by Thery [9] (see above) | WB, SOMAscan | GAPDH | NR |
3.5. Abundance and Morphology of CRS-Derived EVs
3.6. Differential Expression of CRS-Derived EV Surface and Cargo Proteins
3.7. Directed MicroRNA Cargo of CRS-Derived EVs
3.8. EV-Mediated Fibroblast Interactions in CRS
3.9. Immune Influence of CRS-Derived EV
3.10. CRS-Derived Evs Promote Angiogenesis and Vascular Permeability
3.11. Exosomal Coagulation Pathway Derangement in CRSwNP
3.12. Effect of the Diseased Epithelial Exosomes on the Proliferation of hNECs
3.13. Exosomal Biomarkers Predicting Presence, Phenotype, and Disease Severity in CRS
3.14. Therapeutic Influence on CRS-EVs
3.15. Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bachert, C.; Zhang, N.; Hellings, P.W.; Bousquet, J. Endotype-driven care pathways in patients with chronic rhinosinusitis. J. Allergy Clin. Immunol. 2018, 141, 1543–1551. [Google Scholar] [CrossRef]
- Fokkens, W.J.; Lund, V.J.; Hopkins, C.; Hellings, P.W.; Kern, R.; Reitsma, S.; Toppila-Salmi, S.; Bernal-Sprekelsen, M.; Mullol, J.; Alobid, I.; et al. European Position Paper on Rhinosinusitis and Nasal Polyps 2020. Rhinology 2020, 58, 1–464. [Google Scholar] [CrossRef] [PubMed]
- Stevens, W.W.; Peters, A.T.; Tan, B.K.; Klingler, A.I.; Poposki, J.A.; Hulse, K.E.; Grammer, L.C.; Welch, K.C.; Smith, S.S.; Conley, D.B.; et al. Associations Between Inflammatory Endotypes and Clinical Presentations in Chronic Rhinosinusitis. J. Allergy Clin. Immunol. Pract. 2019, 7, 2812–2820.e3. [Google Scholar] [CrossRef] [PubMed]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Lässer, C.; O’Neil, S.E.; Shelke, G.V.; Sihlbom, C.; Hansson, S.F.; Gho, Y.S.; Lundbäck, B.; Lötvall, J. Exosomes in the nose induce immune cell trafficking and harbour an altered protein cargo in chronic airway inflammation. J. Transl. Med. 2016, 14, 181. [Google Scholar] [CrossRef]
- Thery, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr. Protoc. Cell Biol. 2006, 30, 3–22. [Google Scholar] [CrossRef]
- Lasser, C.; O’Neil, S.E.; Ekerljung, L.; Ekstrom, K.; Sjostrand, M.; Lotvall, J. RNA-containing exosomes in human nasal secretions. Am. J. Rhinol. Allergy 2011, 25, 89–93. [Google Scholar] [CrossRef]
- Page, M.J.; Moher, D.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. PRISMA 2020 explanation and elaboration: Updated guidance and exemplars for reporting systematic reviews. BMJ 2021, 372, n160. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.; Seo, E.H.; Lee, S.H.; Kim, K.S.; Oh, C.S.; Moon, J.S.; Kim, J.K. MicroRNA Expression in Extracellular Vesicles from Nasal Lavage Fluid in Chronic Rhinosinusitis. Biomedicines 2021, 9, 471. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Wu, J.; Han, D.; Li, Y.; Wang, T.; Wei, H.; Pan, Y.; Zang, H. Differential expression profile of plasma exosomal microRNAs in chronic rhinosinusitis with nasal polyps. Exp. Biol. Med. 2022, 247, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.M.; Workman, A.D.; Nocera, A.L.; Wu, D.; Mueller, S.K.; Finn, K.; Amiji, M.M.; Bleier, B.S. Discriminant analysis followed by unsupervised cluster analysis including exosomal cystatins predict presence of chronic rhinosinusitis, phenotype, and disease severity. Int. Forum Allergy Rhinol. 2019, 9, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.K.; Nocera, A.L.; Dillon, S.T.; Gu, X.; Wendler, O.; Otu, H.H.; Libermann, T.A.; Bleier, B.S. Noninvasive exosomal proteomic biosignatures, including cystatin SN, peroxiredoxin-5, and glycoprotein VI, accurately predict chronic rhinosinusitis with nasal polyps. Int. Forum Allergy Rhinol. 2019, 9, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.K.; Nocera, A.L.; Dillon, S.T.; Libermann, T.A.; Wendler, O.; Bleier, B.S. Tissue and Exosomal Serine Protease Inhibitors Are Significantly Overexpressed in Chronic Rhinosinusitis With Nasal Polyps. Am. J. Rhinol. Allergy 2019, 33, 359–368. [Google Scholar] [CrossRef]
- Mueller, S.K.; Nocera, A.L.; Dillon, S.T.; Wu, D.; Libermann, T.A.; Bleier, B.S. Highly multiplexed proteomic analysis reveals significant tissue and exosomal coagulation pathway derangement in chronic rhinosinusitis with nasal polyps. Int. Forum Allergy Rhinol. 2018, 8, 1438–1444. [Google Scholar] [CrossRef]
- Mueller, S.K.; Nocera, A.L.; Workman, A.; Libermann, T.; Dillon, S.T.; Stegmann, A.; Wurm, J.; Iro, H.; Wendler, O.; Bleier, B.S. Significant polyomic and functional upregulation of the PAPP-A/IGFBP-4/5/IGF-1 axis in chronic rhinosinusitis with nasal polyps. Int. Forum Allergy Rhinol. 2020, 10, 546–555. [Google Scholar] [CrossRef]
- Nocera, A.L.; Miyake, M.M.; Seifert, P.; Han, X.; Bleier, B.S. Exosomes mediate interepithelial transfer of functional P-glycoprotein in chronic rhinosinusitis with nasal polyps. Laryngoscope 2017, 127, E295–E300. [Google Scholar] [CrossRef]
- Nocera, A.L.; Mueller, S.K.; Workman, A.D.; Wu, D.; McDonnell, K.; Sadow, P.M.; Amiji, M.M.; Bleier, B.S. Cystatin SN is a potent upstream initiator of epithelial-derived type 2 inflammation in chronic rhinosinusitis. J. Allergy Clin. Immunol. 2022. [Google Scholar] [CrossRef]
- Shimizu, S.; Tojima, I.; Nakamura, K.; Arai, H.; Kouzaki, H.; Shimizu, T. Nasal polyp fibroblasts (NPFs)-derived exosomes are important for the release of vascular endothelial growth factor from cocultured eosinophils and NPFs. Auris Nasus Larynx 2022, 49, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.H.; Byun, J.; Lee, K.; Kim, B.; Noh, Y.K.; Tran, N.L.; Park, K.; Kim, S.H.; Kim, T.H.; Oh, S.J. Exosomal miRNA-19a and miRNA-614 Induced by Air Pollutants Promote Proinflammatory M1 Macrophage Polarization via Regulation of RORα Expression in Human Respiratory Mucosal Microenvironment. J. Immunol. 2020, 205, 3179–3190. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.F.; Lee, C.H.; Liang, S.S.; Hung, C.C.; Wu, Y.R.; Chien, C.Y.; Lee, C.H.; Chen, J.Y. Mucin 5AC is significantly upregulated in exosomes from the nasal lavage fluid and may promote the expression of COX-2, VEGF and MMP-9: An implication in nasal polyp pathogenesis. Rhinology 2021, 59, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Workman, A.D.; Miyake, M.M.; Nocera, A.L.; Mueller, S.K.; Finn, K.; Otu, H.H.; Libermann, T.A.; Bleier, B.S. Unexpected effects of systemic steroids on the CRSwNP proteome: Is protein upregulation more important than inhibition? Int. Forum Allergy Rhinol. 2020, 10, 334–342. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, J.; Cheng, L.; Ni, H.; You, B.; Shan, Y.; Bao, L.; Wu, D.; Zhang, T.; Yue, H.; et al. A disintegrin and metalloprotease 10-containing exosomes derived from nasal polyps promote angiogenesis and vascular permeability. Mol. Med. Rep. 2018, 17, 5921–5927. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, T.; Yan, Y.; Zhang, J.; Zhou, Y.; Pei, Y.; Yao, L.; You, B.; Chen, J. Exosomal miR-22-3p Derived from Chronic Rhinosinusitis with Nasal Polyps Regulates Vascular Permeability by Targeting VE-Cadherin. Biomed. Res. Int. 2020, 2020, 1237678. [Google Scholar] [CrossRef]
- Zhou, M.; Tan, K.S.; Guan, W.J.; Jiang, L.J.; Deng, J.; Gao, W.X.; Lee, Y.M.; Xu, Z.F.; Luo, X.; Liu, C.; et al. Proteomics profiling of epithelium-derived exosomes from nasal polyps revealed signaling functions affecting cellular proliferation. Respir. Med. 2020, 162, 105871. [Google Scholar] [CrossRef]
- Nocera, A.L.; Meurer, A.T.; Miyake, M.M.; Sadow, P.M.; Han, X.; Bleier, B.S. Secreted P-glycoprotein is a noninvasive biomarker of chronic rhinosinusitis. Laryngoscope 2017, 127, E1–E4. [Google Scholar] [CrossRef]
- Wang, S.W.; Lan, T.; Sheng, H.; Zheng, F.; Lei, M.K.; Wang, L.X.; Chen, H.F.; Xu, C.Y.; Zhang, F. Nobiletin Alleviates Non-alcoholic Steatohepatitis in MCD-Induced Mice by Regulating Macrophage Polarization. Front. Physiol. 2021, 12, 687744. [Google Scholar] [CrossRef]
- Howarth, P.H.; Persson, C.G.; Meltzer, E.O.; Jacobson, M.R.; Durham, S.R.; Silkoff, P.E. Objective monitoring of nasal airway inflammation in rhinitis. J. Allergy Clin. Immunol. 2005, 115, S414–S441. [Google Scholar] [CrossRef]
- Kurian, T.K.; Banik, S.; Gopal, D.; Chakrabarti, S.; Mazumder, N. Elucidating Methods for Isolation and Quantification of Exosomes: A Review. Mol. Biotechnol. 2021, 63, 249–266. [Google Scholar] [CrossRef] [PubMed]
- An, M.; Wu, J.; Zhu, J.; Lubman, D.M. Comparison of an Optimized Ultracentrifugation Method versus Size-Exclusion Chromatography for Isolation of Exosomes from Human Serum. J. Proteome Res. 2018, 17, 3599–3605. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Romancino, D.P.; Paterniti, G.; Campos, Y.; De Luca, A.; Di Felice, V.; d’Azzo, A.; Bongiovanni, A. Identification and characterization of the nano-sized vesicles released by muscle cells. FEBS Lett. 2013, 587, 1379–1384. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Su, M.; Wang, X.; Xie, C. Exosomal microRNA-22-3p alleviates cerebral ischemic injury by modulating KDM6B/BMP2/BMF axis. Stem Cell Res. Ther. 2021, 12, 111. [Google Scholar] [CrossRef]
- Jiang, H.; Zhao, H.; Zhang, M.; He, Y.; Li, X.; Xu, Y.; Liu, X. Hypoxia Induced Changes of Exosome Cargo and Subsequent Biological Effects. Front. Immunol. 2022, 13, 824188. [Google Scholar] [CrossRef]
- Boyle, E.I.; Weng, S.; Gollub, J.; Jin, H.; Botstein, D.; Cherry, J.M.; Sherlock, G. GO::TermFinder--open source software for accessing Gene Ontology information and finding significantly enriched Gene Ontology terms associated with a list of genes. Bioinformatics 2004, 20, 3710–3715. [Google Scholar] [CrossRef]
- Lin, T.M.; Galbert, S.P.; Kiefer, D.; Spellacy, W.N.; Gall, S. Characterization of four human pregnancy-associated plasma proteins. Am. J. Obstet. Gynecol. 1974, 118, 223–236. [Google Scholar] [CrossRef]
- Conover, C.A.; Oxvig, C. PAPP-A and cancer. J. Mol. Endocrinol. 2018, 61, T1–T10. [Google Scholar] [CrossRef]
- Espelund, U.S.; Bjerre, M.; Hjortebjerg, R.; Rasmussen, T.R.; Lundby, A.; Hoeflich, A.; Folkersen, B.H.; Oxvig, C.; Frystyk, J. Insulin-Like Growth Factor Bioactivity, Stanniocalcin-2, Pregnancy-Associated Plasma Protein-A, and IGF-Binding Protein-4 in Pleural Fluid and Serum From Patients With Pulmonary Disease. J. Clin. Endocrinol. Metab. 2017, 102, 3526–3534. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Jiang, J.; Qu, C.; Wang, C.; Sun, Z. Predictive value of serum pregnancy-associated plasma protein A for patients with ischemic cerebrovascular disease. J. Clin. Lab. Anal. 2017, 31, e22091. [Google Scholar] [CrossRef] [PubMed]
- Bulut, I.; Ozseker, Z.F.; Coskun, A.; Serteser, M.; Unsal, I. Pregnancy-associated plasma protein-A (PAPP-A) levels in patients with severe allergic asthma are reduced by omalizumab. J. Asthma 2018, 55, 1116–1121. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, H.; Zhou, L.; Wang, Z.; Hua, B. Pregnancy-Associated Plasma Protein A Induces Inflammatory Cytokine Expression by Activating IGF-I/PI3K/Akt Pathways. Mediat. Inflamm. 2019, 2019, 8436985. [Google Scholar] [CrossRef] [PubMed]
- Monget, P.; Oxvig, C. PAPP-A and the IGF system. Ann. Endocrinol. 2016, 77, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Li, X.; Chang, L.; Chen, X.; Huang, Z.; Bao, H.; Huang, J.; Yang, L.; Wu, X.; Wang, Z.; et al. IL-19 Up-Regulates Mucin 5AC Production in Patients With Chronic Rhinosinusitis via STAT3 Pathway. Front. Immunol. 2019, 10, 1682. [Google Scholar] [CrossRef]
- Xia, W.; Bai, J.; Wu, X.; Wei, Y.; Feng, S.; Li, L.; Zhang, J.; Xiong, G.; Fan, Y.; Shi, J.; et al. Interleukin-17A promotes MUC5AC expression and goblet cell hyperplasia in nasal polyps via the Act1-mediated pathway. PLoS ONE 2014, 9, e98915. [Google Scholar] [CrossRef]
- Jiao, J.; Zhang, T.; Zhang, Y.; Li, J.; Wang, M.; Wang, M.; Li, Y.; Wang, X.; Zhang, L. Epidermal growth factor upregulates expression of MUC5AC via TMEM16A, in chronic rhinosinusitis with nasal polyps. Allergy Asthma Clin. Immunol. 2020, 16, 40. [Google Scholar] [CrossRef]
- Jiao, J.; Hu, P.; Li, Y.; Cai, C.; Wang, X.; Zhang, L. PM2.5 Upregulates the Expression of MUC5AC via the EGFR-PI3K Pathway in Human Sinonasal Epithelial Cells. Int. Arch. Allergy Immunol. 2022, 183, 361–374. [Google Scholar] [CrossRef]
- Kato, K.; Chang, E.H.; Chen, Y.; Lu, W.; Kim, M.M.; Niihori, M.; Hecker, L.; Kim, K.C. MUC1 contributes to goblet cell metaplasia and MUC5AC expression in response to cigarette smoke in vivo. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 319, L82–L90. [Google Scholar] [CrossRef]
- Hirschberger, S.; Hinske, L.C.; Kreth, S. MiRNAs: Dynamic regulators of immune cell functions in inflammation and cancer. Cancer Lett. 2018, 431, 11–21. [Google Scholar] [CrossRef]
- Xuan, L.; Luan, G.; Wang, Y.; Lan, F.; Zhang, X.; Hao, Y.; Zheng, M.; Wang, X.; Zhang, L. MicroRNAs regulating mucin type O-glycan biosynthesis and transforming growth factor beta signaling pathways in nasal mucosa of patients with chronic rhinosinusitis with nasal polyps in Northern China. Int. Forum Allergy Rhinol. 2019, 9, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Xia, G.; Bao, L.; Gao, W.; Liu, S.; Ji, K.; Li, J. Differentially Expressed miRNA in Inflammatory Mucosa of Chronic Rhinosinusitis. J. Nanosci. Nanotechnol. 2015, 15, 2132–2139. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Zhang, Y.N.; Li, H.B.; Hu, C.Y.; Wang, N.; Cao, P.P.; Liao, B.; Lu, X.; Cui, Y.H.; Liu, Z. Overexpression of miR-125b, a novel regulator of innate immunity, in eosinophilic chronic rhinosinusitis with nasal polyps. Am. J. Respir. Crit. Care Med. 2012, 185, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Korde, A.; Ahangari, F.; Haslip, M.; Zhang, X.; Liu, Q.; Cohn, L.; Gomez, J.L.; Chupp, G.; Pober, J.S.; Gonzalez, A.; et al. An endothelial microRNA-1-regulated network controls eosinophil trafficking in asthma and chronic rhinosinusitis. J. Allergy Clin. Immunol. 2020, 145, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Panganiban, R.P.; Lambert, K.A.; Hsu, M.H.; Laryea, Z.; Ishmael, F.T. Isolation and profiling of plasma microRNAs: Biomarkers for asthma and allergic rhinitis. Methods 2019, 152, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Panganiban, R.P.; Wang, Y.; Howrylak, J.; Chinchilli, V.M.; Craig, T.J.; August, A.; Ishmael, F.T. Circulating microRNAs as biomarkers in patients with allergic rhinitis and asthma. J. Allergy Clin. Immunol. 2016, 137, 1423–1432. [Google Scholar] [CrossRef]
- Panganiban, R.P.; Pinkerton, M.H.; Maru, S.Y.; Jefferson, S.J.; Roff, A.N.; Ishmael, F.T. Differential microRNA epression in asthma and the role of miR-1248 in regulation of IL-5. Am. J. Clin. Exp. Immunol. 2012, 1, 154–165. [Google Scholar]
- Wu, G.; Yang, G.; Zhang, R.; Xu, G.; Zhang, L.; Wen, W.; Lu, J.; Liu, J.; Yu, Y. Altered microRNA Expression Profiles of Extracellular Vesicles in Nasal Mucus From Patients With Allergic Rhinitis. Allergy Asthma Immunol. Res. 2015, 7, 449–457. [Google Scholar] [CrossRef]
- Guzman-Aranguez, A.; Argueso, P. Structure and biological roles of mucin-type O-glycans at the ocular surface. Ocul. Surf. 2010, 8, 8–17. [Google Scholar] [CrossRef]
- Schmidt-Weber, C.B.; Blaser, K. The role of TGF-beta in allergic inflammation. Immunol. Allergy Clin. N. Am. 2006, 26, 233–244. [Google Scholar] [CrossRef]
- Zhang, Y.E. Non-Smad Signaling Pathways of the TGF-beta Family. Cold Spring Harb. Perspect. Biol. 2017, 9, a022129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, J.; Wang, M.; Duan, S.; Meng, Y.; Meng, N.; Li, Y.; Fan, E.; Akdis, C.A.; Zhang, L. Transforming growth factor-beta1 decreases epithelial tight junction integrity in chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2018, 141, 1160–1163.e9. [Google Scholar] [CrossRef]
- Tubita, V.; Callejas-Diaz, B.; Roca-Ferrer, J.; Marin, C.; Liu, Z.; Wang, Y.; Mullol, J. Role of microRNAs in inflammatory upper airway diseases. Allergy 2021, 76, 1967–1980. [Google Scholar] [CrossRef] [PubMed]
- Galagali, H.; Kim, J.K. The multifaceted roles of microRNAs in differentiation. Curr. Opin. Cell Biol. 2020, 67, 118–140. [Google Scholar] [CrossRef] [PubMed]
- Melincovici, C.S.; Bosca, A.B.; Susman, S.; Marginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Wittekindt, C.; Hess, A.; Bloch, W.; Sultanie, S.; Michel, O. Immunohistochemical expression of VEGF and VEGF receptors in nasal polyps as compared to normal turbinate mucosa. Eur. Arch. Otorhinolaryngol. 2002, 259, 294–298. [Google Scholar] [CrossRef]
- Cavaliere, C.; Masieri, S.; Greco, A.; Lambiase, A.; Segatto, M. Nasal expression of the vascular endothelial growth factor and its receptors is reduced by mepolizumab in chronic rhinosinusitis with nasal polyposis. Ann. Allergy Asthma Immunol. 2021, 126, 442–443. [Google Scholar] [CrossRef]
- Sun, Z.; Luo, D.; Wang, S.; Huang, R.; Dong, P. Salvianolic acid B inhibits myofibroblast differentiation and extracellular matrix accumulation in nasal polyp fibroblasts via the TGF-beta1 signaling pathway. Mol. Med. Rep. 2021, 23, 478. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, S.; Tojima, I.; Takezawa, K.; Matsumoto, K.; Kouzaki, H.; Shimizu, T. Thrombin and activated coagulation factor X stimulate the release of cytokines and fibronectin from nasal polyp fibroblasts via protease-activated receptors. Am. J. Rhinol. Allergy 2017, 31, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Ball, S.L.; Mann, D.A.; Wilson, J.A.; Fisher, A.J. The Role of the Fibroblast in Inflammatory Upper Airway Conditions. Am. J. Pathol. 2016, 186, 225–233. [Google Scholar] [CrossRef]
- Azizzadeh Delshad, A.; Jalali Nadoushan, M.; Davati, A.; Rostami, A. Expression of Vascular Endothelial Growth Factor in Nasal Polyp and Chronic Rhinosinusitis. Iran. J. Pathol. 2016, 11, 231–237. [Google Scholar] [PubMed]
- Xu, M.; Chen, D.; Zhou, H.; Zhang, W.; Xu, J.; Chen, L. The Role of Periostin in the Occurrence and Progression of Eosinophilic Chronic Sinusitis with Nasal Polyps. Sci. Rep. 2017, 7, 9479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Q.; Zhang, T.; Zhang, J. Correlation analysis of STAT3 and VEGF expression and eosinophil infiltration in nasal polyps. Eur. Arch. Otorhinolaryngol. 2015, 272, 1955–1960. [Google Scholar] [CrossRef]
- Muluk, N.B.; Atasoy, P.; Arikan, O.K.; Koc, C. Role of vascular endothelial growth factor in the pathogenesis of nasal polyps. J. Otolaryngol. 2007, 36, 357–366. [Google Scholar] [PubMed]
- Matsuwaki, Y.; Ookushi, T.; Asaka, D.; Mori, E.; Nakajima, T.; Yoshida, T.; Kojima, J.; Chiba, S.; Ootori, N.; Moriyama, H. Chronic rhinosinusitis: Risk factors for the recurrence of chronic rhinosinusitis based on 5-year follow-up after endoscopic sinus surgery. Int. Arch. Allergy Immunol. 2008, 146 (Suppl. 1), 77–81. [Google Scholar] [CrossRef]
- Shimizu, S.; Kouzaki, H.; Ogawa, T.; Takezawa, K.; Tojima, I.; Shimizu, T. Eosinophil-epithelial cell interactions stimulate the production of MUC5AC mucin and profibrotic cytokines involved in airway tissue remodeling. Am. J. Rhinol. Allergy 2014, 28, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.W.; Park, J.H.; Jo, M.S.; Shin, J.M.; Kim, D.W.; Park, I.H. Eosinophil-Derived Osteopontin Induces the Expression of Pro-Inflammatory Mediators and Stimulates Extracellular Matrix Production in Nasal Fibroblasts: The Role of Osteopontin in Eosinophilic Chronic Rhinosinusitis. Front. Immunol. 2022, 13, 777928. [Google Scholar] [CrossRef]
- Kahveci, O.K.; Derekoy, F.S.; Yilmaz, M.; Serteser, M.; Altuntas, A. The role of MMP-9 and TIMP-1 in nasal polyp formation. Swiss Med. Wkly. 2008, 138, 684–688. [Google Scholar]
- Li, X.; Tao, Y.; Li, X. Expression of MMP-9/TIMP-2 in nasal polyps and its functional implications. Int. J. Clin. Exp. Pathol. 2015, 8, 14556–14561. [Google Scholar] [PubMed]
- Li, X.; Huang, J.; Chen, X.; Lai, X.; Huang, Z.; Li, Y.; Li, S.; Chang, L.; Zhang, G. IL-19 induced by IL-13/IL-17A in the nasal epithelium of patients with chronic rhinosinusitis upregulates MMP-9 expression via ERK/NF-kappaB signaling pathway. Clin. Transl. Allergy 2021, 11, e12003. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chang, L.; Li, X.; Huang, J.; Yang, L.; Lai, X.; Huang, Z.; Wang, Z.; Wu, X.; Zhao, J.; et al. Tc17/IL-17A Up-Regulated the Expression of MMP-9 via NF-kappaB Pathway in Nasal Epithelial Cells of Patients With Chronic Rhinosinusitis. Front. Immunol. 2018, 9, 2121. [Google Scholar] [CrossRef] [PubMed]
- De Borja Callejas, F.; Picado, C.; Martinez-Anton, A.; Alobid, I.; Pujols, L.; Valero, A.; Roca-Ferrer, J.; Mullol, J. Differential expression of remodeling markers by tissue structure in nasal polyposis. Am. J. Rhinol. Allergy 2013, 27, e69–e74. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Wang, C.H.; Wu, P.W.; He, J.R.; Huang, C.C.; Chang, P.H.; Fu, C.H.; Lee, T.J. Increased nasal matrix metalloproteinase-1 and -9 expression in smokers with chronic rhinosinusitis and asthma. Sci. Rep. 2019, 9, 15357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumsion, J.S.; Pulsipher, A.; Alt, J.A. Differential expression and role of S100 proteins in chronic rhinosinusitis. Curr. Opin. Allergy Clin. Immunol. 2020, 20, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Tieu, D.D.; Peters, A.T.; Carter, R.G.; Suh, L.; Conley, D.B.; Chandra, R.; Norton, J.; Grammer, L.C.; Harris, K.E.; Kato, A.; et al. Evidence for diminished levels of epithelial psoriasin and calprotectin in chronic rhinosinusitis. J. Allergy Clin. Immunol. 2010, 125, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Van Crombruggen, K.; Vogl, T.; Perez-Novo, C.; Holtappels, G.; Bachert, C. Differential release and deposition of S100A8/A9 proteins in inflamed upper airway tissue. Eur. Respir. J. 2016, 47, 264–274. [Google Scholar] [CrossRef]
- Cho, S.H.; Kim, D.W.; Lee, S.H.; Kolliputi, N.; Hong, S.J.; Suh, L.; Norton, J.; Hulse, K.E.; Seshadri, S.; Conley, D.B.; et al. Age-related increased prevalence of asthma and nasal polyps in chronic rhinosinusitis and its association with altered IL-6 trans-signaling. Am. J. Respir. Cell Mol. Biol. 2015, 53, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Pulsipher, A.; Davis, B.M.; Smith, K.A.; Ashby, S.; Qin, X.; Firpo, M.; Orlandi, R.R.; Alt, J.A. Calgranulin C (S100A12) Is Differentially Expressed in Subtypes of Chronic Rhinosinusitis. Am. J. Rhinol. Allergy 2018, 32, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Gorur, A.; Bayraktar, R.; Ivan, C.; Mokhlis, H.A.; Bayraktar, E.; Kahraman, N.; Karakas, D.; Karamil, S.; Kabil, N.N.; Kanlikilicer, P.; et al. ncRNA therapy with miRNA-22-3p suppresses the growth of triple-negative breast cancer. Mol. Ther. Nucleic Acids 2021, 23, 930–943. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, X.; Wang, H.; Guo, X.; Xue, J.; Wang, X.; Ni, J. MicroRNA-22-3p and MicroRNA-149-5p Inhibit Human Hepatocellular Carcinoma Cell Growth and Metastasis Properties by Regulating Methylenetetrahydrofolate Reductase. Curr. Issues Mol. Biol. 2022, 44, 63. [Google Scholar] [CrossRef]
- Jia, X.; Yang, M.; Hu, W.; Cai, S. Overexpression of miRNA-22-3p attenuates osteoporosis by targeting MAPK14. Exp. Ther. Med. 2021, 22, 692. [Google Scholar] [CrossRef] [PubMed]
- Korma, W.; Mihret, A.; Tarekegn, A.; Chang, Y.; Hwang, D.; Tessema, T.S.; Lee, H. Identification of Circulating miR-22-3p and miR-93-5p as Stable Endogenous Control in Tuberculosis Study. Diagnostics 2020, 10, 868. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.H.; Chiu, C.T.; Lee, C.; Chu, Y.Y.; Cheng, H.T.; Hsu, J.T.; Wu, R.C.; Yeh, T.S.; Lin, K.H. Circulating microRNA-22-3p Predicts the Malignant Progression of Precancerous Gastric Lesions from Intestinal Metaplasia to Early Adenocarcinoma. Dig. Dis. Sci. 2018, 63, 2301–2308. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Zhan, H.; Zhou, X.Y.; Yao, L.; Yan, M.; Chen, A.; Liu, J.; Ren, X.; Zhang, X.; Liu, J.X.; et al. MicroRNA-22 regulates inflammation and angiogenesis via targeting VE-cadherin. FEBS Lett. 2017, 591, 513–526. [Google Scholar] [CrossRef]
- Rastegar-Moghaddam, S.H.; Ebrahimzadeh-Bideskan, A.; Shahba, S.; Malvandi, A.M.; Mohammadipour, A. MicroRNA-22: A Novel and Potent Biological Therapeutics in Neurological Disorders. Mol. Neurobiol. 2022, 59, 2694–2701. [Google Scholar] [CrossRef]
- Giannotta, M.; Trani, M.; Dejana, E. VE-cadherin and endothelial adherens junctions: Active guardians of vascular integrity. Dev. Cell 2013, 26, 441–454. [Google Scholar] [CrossRef]
- Yukitatsu, Y.; Hata, M.; Yamanegi, K.; Yamada, N.; Ohyama, H.; Nakasho, K.; Kojima, Y.; Oka, H.; Tsuzuki, K.; Sakagami, M.; et al. Decreased expression of VE-cadherin and claudin-5 and increased phosphorylation of VE-cadherin in vascular endothelium in nasal polyps. Cell Tissue Res. 2013, 352, 647–657. [Google Scholar] [CrossRef]
- Dreymueller, D.; Pruessmeyer, J.; Groth, E.; Ludwig, A. The role of ADAM-mediated shedding in vascular biology. Eur. J. Cell Biol. 2012, 91, 472–485. [Google Scholar] [CrossRef] [PubMed]
- Isozaki, T.; Rabquer, B.J.; Ruth, J.H.; Haines, G.K., 3rd; Koch, A.E. ADAM-10 is overexpressed in rheumatoid arthritis synovial tissue and mediates angiogenesis. Arthritis Rheum. 2013, 65, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Donners, M.M.; Wolfs, I.M.; Olieslagers, S.; Mohammadi-Motahhari, Z.; Tchaikovski, V.; Heeneman, S.; van Buul, J.D.; Caolo, V.; Molin, D.G.; Post, M.J.; et al. A disintegrin and metalloprotease 10 is a novel mediator of vascular endothelial growth factor-induced endothelial cell function in angiogenesis and is associated with atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2188–2195. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Pruessmeyer, J.; Maretzky, T.; Ludwig, A.; Blobel, C.P.; Saftig, P.; Reiss, K. ADAM10 regulates endothelial permeability and T-Cell transmigration by proteolysis of vascular endothelial cadherin. Circ. Res. 2008, 102, 1192–1201. [Google Scholar] [CrossRef]
- Bae, W.Y.; Park, S.K.; Kim, D.H.; Koh, T.K.; Hur, D.Y.; Chueh, H.W. Expression of ADAM17 and ADAM10 in nasal polyps. Int. Forum Allergy Rhinol. 2016, 6, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Van Crombruggen, K.; Holtappels, G.; De Ruyck, N.; Derycke, L.; Tomassen, P.; Bachert, C. RAGE processing in chronic airway conditions: Involvement of Staphylococcus aureus and ECP. J. Allergy Clin. Immunol. 2012, 129, 1515–1521.e8. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.P.; Wang, B.F.; Norton, J.E.; Suh, L.A.; Carter, R.G.; Stevens, W.W.; Staudacher, A.G.; Huang, J.H.; Hulse, K.E.; Peters, A.T.; et al. Studies on activation and regulation of the coagulation cascade in chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2022, 150, 467–476.e1. [Google Scholar] [CrossRef]
- Shimizu, S.; Ogawa, T.; Takezawa, K.; Tojima, I.; Kouzaki, H.; Shimizu, T. Tissue factor and tissue factor pathway inhibitor in nasal mucosa and nasal secretions of chronic rhinosinusitis with nasal polyp. Am. J. Rhinol. Allergy 2015, 29, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Takabayashi, T.; Kato, A.; Peters, A.T.; Hulse, K.E.; Suh, L.A.; Carter, R.; Norton, J.; Grammer, L.C.; Cho, S.H.; Tan, B.K.; et al. Excessive fibrin deposition in nasal polyps caused by fibrinolytic impairment through reduction of tissue plasminogen activator expression. Am. J. Respir. Crit. Care Med. 2013, 187, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Takabayashi, T.; Kato, A.; Peters, A.T.; Hulse, K.E.; Suh, L.A.; Carter, R.; Norton, J.; Grammer, L.C.; Tan, B.K.; Chandra, R.K.; et al. Increased expression of factor XIII-A in patients with chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2013, 132, 584–592.e4. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, S.; Gabazza, E.C.; Ogawa, T.; Tojima, I.; Hoshi, E.; Kouzaki, H.; Shimizu, T. Role of thrombin in chronic rhinosinusitis-associated tissue remodeling. Am. J. Rhinol. Allergy 2011, 25, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.Y.; Zhou, M.; Lin, M.Q.; Liang, S.T.; Yan, Y.; Wang, S.M.; Fang, C.S.; Li, D.; Ruan, Y. Research Progress on the SERPINE1 Protein and Chronic Inflammatory Diseases of the Upper Respiratory Tract: A Literature Review. Int. Arch. Allergy Immunol. 2021, 182, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Imoto, Y.; Kato, A.; Takabayashi, T.; Stevens, W.; Norton, J.E.; Suh, L.A.; Carter, R.G.; Weibman, A.R.; Hulse, K.E.; Harris, K.E.; et al. Increased thrombin-activatable fibrinolysis inhibitor levels in patients with chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2019, 144, 1566–1574.e6. [Google Scholar] [CrossRef] [PubMed]
- Poncin, M.A.; Van Meerbeeck, P.; Simpson, J.D.; Clippe, A.; Tyckaert, F.; Bouillenne, F.; Degand, H.; Matagne, A.; Morsomme, P.; Knoops, B.; et al. Role of the Redox State of Human Peroxiredoxin-5 on Its TLR4-Activating DAMP Function. Antioxidants 2021, 10, 1902. [Google Scholar] [CrossRef] [PubMed]
- Knoops, B.; Becker, S.; Poncin, M.A.; Glibert, J.; Derclaye, S.; Clippe, A.; Alsteens, D. Specific Interactions Measured by AFM on Living Cells between Peroxiredoxin-5 and TLR4: Relevance for Mechanisms of Innate Immunity. Cell Chem. Biol. 2018, 25, 550–559.e3. [Google Scholar] [CrossRef] [PubMed]
- Millien, V.O.; Lu, W.; Shaw, J.; Yuan, X.; Mak, G.; Roberts, L.; Song, L.Z.; Knight, J.M.; Creighton, C.J.; Luong, A.; et al. Cleavage of fibrinogen by proteinases elicits allergic responses through Toll-like receptor 4. Science 2013, 341, 792–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borst, O.; Gawaz, M. Glycoprotein VI—Novel target in antiplatelet medication. Pharmacol. Ther. 2021, 217, 107630. [Google Scholar] [CrossRef] [PubMed]
- Claushuis, T.A.M.; de Vos, A.F.; Nieswandt, B.; Boon, L.; Roelofs, J.; de Boer, O.J.; van ‘t Veer, C.; van der Poll, T. Platelet glycoprotein VI aids in local immunity during pneumonia-derived sepsis caused by gram-negative bacteria. Blood 2018, 131, 864–876. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.M.; Nocera, A.; Miyake, M.M. P-glycoprotein and chronic rhinosinusitis. World J. Otorhinolaryngol. Head Neck Surg. 2018, 4, 169–174. [Google Scholar] [CrossRef]
- Bleier, B.S.; Nocera, A.L.; Iqbal, H.; Hoang, J.D.; Alvarez, U.; Feldman, R.E.; Han, X. P-glycoprotein promotes epithelial T helper 2-associated cytokine secretion in chronic sinusitis with nasal polyps. Int. Forum Allergy Rhinol. 2014, 4, 488–494. [Google Scholar] [CrossRef]
- Leopoldo, M.; Nardulli, P.; Contino, M.; Leonetti, F.; Luurtsema, G.; Colabufo, N.A. An updated patent review on P-glycoprotein inhibitors (2011–2018). Expert Opin. Ther. Pat. 2019, 29, 455–461. [Google Scholar] [CrossRef]
- Workman, A.D.; Mueller, S.K.; McDonnell, K.; Goldfarb, J.W.; Bleier, B.S. Phase I safety and tolerability study of topical verapamil HCl in chronic rhinosinusitis with nasal polyps. Int. Forum Allergy Rhinol. 2022, 12, 1071–1074. [Google Scholar] [CrossRef]
- Taha, M.S.; Nocera, A.; Workman, A.; Amiji, M.M.; Bleier, B.S. P-glycoprotein inhibition with verapamil overcomes mometasone resistance in Chronic Sinusitis with Nasal Polyps. Rhinology 2021, 59, 205–211. [Google Scholar] [CrossRef]
- Bleier, B.S.; Kocharyan, A.; Singleton, A.; Han, X. Verapamil modulates interleukin-5 and interleukin-6 secretion in organotypic human sinonasal polyp explants. Int. Forum Allergy Rhinol. 2015, 5, 10–13. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.M.; Nocera, A.; Levesque, P.; Guo, R.; Finn, C.A.; Goldfarb, J.; Gray, S.; Holbrook, E.; Busaba, N.; Dolci, J.E.L.; et al. Double-blind placebo-controlled randomized clinical trial of verapamil for chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2017, 140, 271–273. [Google Scholar] [CrossRef]
- Kato, Y.; Takabayashi, T.; Sakashita, M.; Imoto, Y.; Tokunaga, T.; Ninomiya, T.; Morikawa, T.; Yoshida, K.; Noguchi, E.; Fujieda, S. Expression and Functional Analysis of CST1 in Intractable Nasal Polyps. Am. J. Respir. Cell Mol. Biol. 2018, 59, 448–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, B.; Lou, H.; Wang, Y.; Li, Y.; Meng, Y.; Qi, S.; Wang, M.; Xiao, L.; Wang, C.; Zhang, L. Epithelium-derived cystatin SN enhances eosinophil activation and infiltration through IL-5 in patients with chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2019, 144, 455–469. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Yan, B.; Wang, Y.; Wang, C.; Zhang, L. Prognostic and pharmacologic value of cystatin SN for chronic rhinosinusitis with nasal polyps. J. Allergy Clin. Immunol. 2021, 148, 450–460. [Google Scholar] [CrossRef]
- Mueller, S.K.; Wendler, O.; Nocera, A.; Grundtner, P.; Schlegel, P.; Agaimy, A.; Iro, H.; Bleier, B.S. Escalation in mucus cystatin 2, pappalysin-A, and periostin levels over time predict need for recurrent surgery in chronic rhinosinusitis with nasal polyps. Int. Forum Allergy Rhinol. 2019, 9, 1212–1219. [Google Scholar] [CrossRef]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and intercellular transfer of microRNAs in living cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef]
- Savina, A.; Furlan, M.; Vidal, M.; Colombo, M.I. Exosome release is regulated by a calcium-dependent mechanism in K562 cells. J. Biol. Chem. 2003, 278, 20083–20090. [Google Scholar] [CrossRef]
- Kulshreshtha, A.; Ahmad, T.; Agrawal, A.; Ghosh, B. Proinflammatory role of epithelial cell-derived exosomes in allergic airway inflammation. J. Allergy Clin. Immunol. 2013, 131, 1194–1203. [Google Scholar] [CrossRef]
- Workman, A.D.; Nocera, A.L.; Mueller, S.K.; Otu, H.H.; Libermann, T.A.; Bleier, B.S. Translating transcription: Proteomics in chronic rhinosinusitis with nasal polyps reveals significant discordance with messenger RNA expression. Int. Forum Allergy Rhinol. 2019, 9, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Al Obaidi, A.H. Role of airway lactoperoxidase in scavenging of hydrogen peroxide damage in asthma. Ann. Thorac. Med. 2007, 2, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Seshadri, S.; Lin, D.C.; Rosati, M.; Carter, R.G.; Norton, J.E.; Suh, L.; Kato, A.; Chandra, R.K.; Harris, K.E.; Chu, H.W.; et al. Reduced expression of antimicrobial PLUNC proteins in nasal polyp tissues of patients with chronic rhinosinusitis. Allergy 2012, 67, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.T.; Johnstone, R.M. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: Selective externalization of the receptor. Cell 1983, 33, 967–978. [Google Scholar] [CrossRef]
- Harding, C.; Stahl, P. Transferrin recycling in reticulocytes: pH and iron are important determinants of ligand binding and processing. Biochem. Biophys. Res. Commun. 1983, 113, 650–658. [Google Scholar] [CrossRef]
- Lucchetti, D.; Calapa, F.; Palmieri, V.; Fanali, C.; Carbone, F.; Papa, A.; De Maria, R.; De Spirito, M.; Sgambato, A. Differentiation Affects the Release of Exosomes from Colon Cancer Cells and Their Ability to Modulate the Behavior of Recipient Cells. Am. J. Pathol. 2017, 187, 1633–1647. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.K. The Role of Exosomes in the Pathophysiology of Chronic Rhinosinusitis. Front. Cell. Infect. Microbiol. 2021, 11, 812920. [Google Scholar] [CrossRef] [PubMed]
- Parra-Ferro, M.; Justice, J.M.; Lobo, B.C.; Munger, S.D.; Schlosser, R.J.; Mulligan, J.K. Utilization of Nasal Mucus to Investigate the Pathophysiology of Chronic Rhinosinusitis. Am. J. Rhinol. Allergy 2022, 19458924221111830. [Google Scholar] [CrossRef]
- Tu, Y.; Liu, J.; Li, T.; Zhou, X.; Tan, K.S.; Ong, H.H.; Sun, K.; Ouyang, Y.; Liang, X.; Ong, Y.K.; et al. Mucus composition abnormalities in sinonasal mucosa of chronic rhinosinusitis with and without nasal polyps. Inflammation 2021, 44, 1937–1948. [Google Scholar] [CrossRef]
- Kao, S.S.; Bassiouni, A.; Ramezanpour, M.; Finnie, J.; Chegeni, N.; Colella, A.D.; Chataway, T.K.; Wormald, P.J.; Vreugde, S.; Psaltis, A.J. Proteomic analysis of nasal mucus samples of healthy patients and patients with chronic rhinosinusitis. J. Allergy Clin. Immunol. 2021, 147, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Biswas, K.; Chang, A.; Hoggard, M.; Radcliff, F.J.; Jiang, Y.; Taylor, M.W.; Darveau, R.; Douglas, R.G. Toll-like receptor activation by sino-nasal mucus in chronic rhinosinusitis. Rhinology 2017, 55, 59–69. [Google Scholar] [CrossRef]
- Al Badaai, Y.; DiFalco, M.R.; Tewfik, M.A.; Samaha, M. Quantitative proteomics of nasal mucus in chronic sinusitis with nasal polyposis. J. Otolaryngol. Head Neck Surg. 2009, 38, 381–389. [Google Scholar]
- Stevens, W.W.; Ocampo, C.J.; Berdnikovs, S.; Sakashita, M.; Mahdavinia, M.; Suh, L.; Takabayashi, T.; Norton, J.E.; Hulse, K.E.; Conley, D.B.; et al. Cytokines in Chronic Rhinosinusitis. Role in Eosinophilia and Aspirin-exacerbated Respiratory Disease. Am. J. Respir. Crit. Care Med. 2015, 192, 682–694. [Google Scholar] [CrossRef]
- Nocera, A.L.; Mueller, S.K.; Stephan, J.R.; Hing, L.; Seifert, P.; Han, X.; Lin, D.T.; Amiji, M.M.; Libermann, T.; Bleier, B.S. Exosome swarms eliminate airway pathogens and provide passive epithelial immunoprotection through nitric oxide. J. Allergy Clin. Immunol. 2019, 143, 1525–1535.e1. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S.; Takanashi, M.; Sudo, K.; Ueda, S.; Ishikawa, A.; Matsuyama, N.; Fujita, K.; Mizutani, T.; Ohgi, T.; Ochiya, T.; et al. Systemically injected exosomes targeted to EGFR deliver antitumor microRNA to breast cancer cells. Mol. Ther. 2013, 21, 185–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, I.A.; Hur, J.Y.; Kim, H.J.; Kim, W.S.; Lee, K.Y. Extracellular Vesicle-Based Bronchoalveolar Lavage Fluid Liquid Biopsy for EGFR Mutation Testing in Advanced Non-Squamous NSCLC. Cancers 2022, 14, 2744. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.B.; Hong, S.W.; Kim, D.K.; Jeon, S.G.; Kim, K.R.; Cho, S.H.; Gho, Y.S.; Jee, Y.K.; Kim, Y.K. Decreased diversity of nasal microbiota and their secreted extracellular vesicles in patients with chronic rhinosinusitis based on a metagenomic analysis. Allergy 2014, 69, 517–526. [Google Scholar] [CrossRef]
- Kim, M.R.; Hong, S.W.; Choi, E.B.; Lee, W.H.; Kim, Y.S.; Jeon, S.G.; Jang, M.H.; Gho, Y.S.; Kim, Y.K. Staphylococcus aureus-derived extracellular vesicles induce neutrophilic pulmonary inflammation via both Th1 and Th17 cell responses. Allergy 2012, 67, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, S.H.; Lim, J.Y.; Kim, D.; Jeong, I.S.; Lee, D.K.; Jang, Y.J. Association between the sinus microbiota with eosinophilic inflammation and prognosis in chronic rhinosinusitis with nasal polyps. Exp. Mol. Med. 2020, 52, 978–987. [Google Scholar] [CrossRef] [PubMed]


| Inclusion Criteria | Exclusion Criteria | |
|---|---|---|
| Study status | Completed, Published | Unfinished, Unpublished |
| Study type | Original article |
Reviews Book chapters Expert opinions Conference papers Case reports |
| Language | English | Other than English |
| Sample origin | Exosomes from human cells | Bacteria-derived exosomes |
| Topic | Studies concerning CRS | Studies not related to the CRS topic |
| Quality | Good quality of research studies | Poor quality of research studies |
| Study | The Source of Exosomes | Study Group | Control Group | Exclusion Criteria | Main Results |
|---|---|---|---|---|---|
| Laesser et al. (2016) [8] | • NLF | • asthma (n = 15) • asthma with coexisting CRS (n = 13) | • HS (n = 14) | • Exposition to antihistamines for 72 h, LABA for 24 h, SABA for 8 h and Spiriva for 24 h | • Nasal exosomes induced the migration of several immune cells (monocytes, neutrophils, and NK cells). • 604 proteins were identified in nasal exosomes. • The nasal exosomal proteome showed strong associations with immune-related functions, such as immune cell trafficking. • In the exosomes from patients with respiratory diseases serum-associated proteins and mucins were upregulated. In contrast, proteins with antimicrobial functions and barrier-related proteins were downregulated relative to the control group. |
| Nocera et al. (2017) [19] | • NM | • CRSwNP (n = 10) | • HS (n = 10) | • Exposition to antibiotics or steroids in last 4 weeks, • Ciliary dysfunction, autoimmune disease, CF, immunodeficiency, smoking | • Exosomal P-gp was significantly enriched among the CRSwNP relative to the control group. • Exosomes were absorbed by cultured sinonasal epithelial cells within 10 min leading to a significant increase in P-gp activity in CRSwNP patients in comparison to the control group. |
| Zhang et al. (2018) [25] | • NLF | • CRSwNP (n = 25) | • HS (n = 15) | • Exposition to topical /systemic steroids in last 3 weeks, • Allergic rhinitis, asthma, aspirin sensitivity | • Incubation of fluorescent NLF-derived exosomes resulted in transferring of fluorescence to HUVECs. • NLF-derived exosomes from CRSwNP patients stimulated tube formation, proliferation, and permeability of HUVECs. • ADAM10 was enriched in NLF-derived exosomes from CRSwNP patients relative to the HS group. |
| Mueller et al. (2018) [17] | • NM | • CRSwNP (n = 10) | • HS (n = 10) | • Exposition to antibiotics or any topical /systemic steroids in last 4 weeks • Ciliary dysfunction, autoimmune disease, CF, immunodeficiency • Among the controls additionally allergy or asthma | • Among all protein pathways, the coagulation cascade was most significantly associated with CRSwNP. • The correlation between tissue- and NM-derived exosomal fibrinolysis and coagulation protein expression was strong, inverse, and highly significant. • Novel tissue proteome findings: overexpression of plasma kallikrein and vitamin K-dependent protein S and the downregulation of coagulation factor IXab in CRSwNP. |
| Mueller et al. (2019) [15] | • NM | • CRSwNP (n = 20) | • HS (n = 20) | • Exposition to antibiotics or any topical /systemic steroids in last 4 weeks • Ciliary dysfunction, autoimmune disease, CF, immunodeficiency • Among the controls additionally allergy or asthma | • The exosomal proteome demonstrated 123 significantly differentially regulated proteins in CRSwNP relative to HS. • 80 exosomal proteins overlapped with the matched CRSwNP tissue proteome, whereas 4 proteins overlapped with the matched whole mucus samples. • 43 significantly dysregulated pathway networks overlapped between the exosomal and tissue proteome in CRSwNP, whereas only 3 among them matched withwhole mucus samples. |
| Mueller et al. (2019) [16] | • NM | • CRSwNP (n = 16) | • HS (n = 16) | • Exposition to antibiotics or any topical /systemic steroids in the last 4 weeks • Ciliary dysfunction, autoimmune disease, CF, immunodeficiency, • Among the controls additionally allergic rhinitis, AERD and asthma | • Exosomal and tissue expression of serpinB2, serpinE1, serpinF2 and serpinG1 was higher in CRSwNP patients than in HS group. • There was a strong and significant correlation among the serpinB2, serpinE1, serpinF2, and serpinG1 genes for tissue and exosomes. |
| Miyake et al. (2019) [14] | • NM | • CRSwNP (n = 40) • CRSsNP (n = 33) | • HS (n = 32) | • Ciliary dysfunction, autoimmune disease, CF, immunodeficiency • Among the controls additionally current smoking or asthma | • Expression of exosomal CST-1 was significantly higher in CRSwNP and control group than in CRSsNP group. • Expression of exosomal CST-2 was significantly higher in CRSwNP than in both the CRsNP and control groups. • Seven clusters were identified among patients based on cystatin expression, diagnosis, demographic, and biological variables. CST-2 levels trended linearly with phenotype and clinical severity parameters. |
| Zhou et al. (2020) [27] | • NLF • hNECs from NM | • CRSwNP without coexisting asthma (n = 3) • CRSwNP with coexisting asthma (n = 3) | • HS (n = 3) | • Exposition to leukotrienes and antibiotics • Atopy, aspirin and nonsteroidal anti-inflammatory drug intolerance • Among the controls, there is additional exposition to topical /systemic steroids | • The hNECs-derived exosomes from patients with CRSwNP with and without asthma contained differentially expressed proteins that were mainly involved in epithelial remodeling via pathways such as p53. • Epithelial-derived exosomes from CRSwNP patients (with and without coexisting asthma) significantly reduced the proliferation rate of control hNECs at an effective concentration of ≥10 µg/mL. |
| Workman et al. (2020) [24] | • NM | • CRSwNP (n = 12) | - | • Exposition to oral steroids or antibiotics in last 30 days • Ciliary dysfunction, autoimmune disease, CF, immunodeficiency | • 18 proteins were identified to be highly underexpressed in CRSwNP, of which 16 increased after steroid treatment, including lactoperoxidase and platelet factor 4. • 53 proteins were highly overexpressed in CRSwNP, of which 22 decreased in quantity after steroid treatment. • Blood coagulation and fibrinolysis regulation pathways of the NM were significantly upregulated in exosomes from CRSwNP patients and decreased after the oral prednisone course. |
| Mueller et al. (2020) [18] | • NM | • CRSwNP (n = 46) | • HS (n = 46) | • Exposition to antibiotics or any topical /systemic steroids in last 4 weeks • Ciliary dysfunction, autoimmune disease, CF, immunodeficiency • Among the controls additionally allergy or asthma | • Significant upregulation of tissue and exosomal PAPP-A on a proteomic, transcriptomic and functional level in CRSwNP compared to HS group. • The transcriptomic data using qPCR demonstrated a significant upregulation of PAPP-A in CRSwNP, a significant downregulation of the inhibitor stanniocalcin-1 (STC-1), STC-2 and insulin-like growth factor binding protein–5 (IGFBP-5) and no differences for IGFBP-4 and insulin-like growth factor-1 (IGF-1) between the CRSwNP and HS group. |
| Shin et al. (2020) [22] | • cultured human nasal epithelial cell line RPMI 2650 | • NECRSwNP (n = 9) • RPMI 2650 cell culture treated with PM for 48 h • PM-treated hNECs | • HS (n = 9) • RPMI 2650 cell culture not treated with PM • control hNECs | • Exposition to oral or topical medication (including steroids, antihistamines, antibiotics) in last 3 months | • The induction of exosomal miRNAs from hNECs upon airborne PM exposure promotes proinflammatory M1 macrophage polarization via downregulation of RORα expression in the human respiratory mucosal microenvironment. • Exosomal miRNA-19a and miRNA-614 directly bind to the 3′-untranslated region of RORα mRNA and downregulate RORα expression, which leads to inflammation due to inflammatory cytokine upregulation and M1/M2 macrophage polarization. • Enhanced expression of miRNA-19a and miRNA-614 and reduced RORα expression were observed in tissue from CRS patients relative to HS. |
| Zhang et al. (2020) [26] | • NLF | • CRSwNP (n = 26) | • HS (n = 10) | • Exposition to topical /systemic steroids in last 3 weeks • Ciliary dysfunction, CF, autoimmune disease, immunodeficiency | • MiRNA-22-3p was upregulated in NLF-EVs from CRSwNP relative to the HS group. • Exosomal miRNA-22-3p derived from CRSwNP enhanced the tubule permeability of HUVECs. • VE-cadherin was determined as a direct target of miRNA-22-3p. • MiRNA-22-3p regulated vascular permeability by targeting VE-cadherin in HUVECs. |
| Cha et al. (2021) [12] | • NLF | • CRSwNP (n = 7) • CRSsNP (n = 8) | • HS (n = 7) | NR | • The expression of exosomal miRNA was significantly increased in NLF of CRS patients relative to HS group. • 12 miRNAs were differentially expressed in exosomes from CRS patients relative to HS individuals, including seven upregulated miRNAs and five downregulated miRNAs. • 8 miRNAs were differentially expressed in the NLF-derived exosomes of CRSwNP relative to CRSsNP patients. • The mucin-type O-glycan biosynthesis was a high-ranked predicted pathway in the presence of CRS, while TGF- β signaling pathway was a high-ranked predicted pathway in CRSwNP relative to CRSsNP patients. |
| Wang et al. (2021) [23] | • NLF | • CRSwNP (n = 20) • CRSsNP -derived fibroblasts treated with the mucin 5AC-enriched exosomes | • CRSsNP (n = 22) • CRSsNP -derived fibroblasts treated with the mucin 5AC-deificent exosomes | • Exposition to topical /systemic steroids or antibiotics in last 2 weeks • Malignancies, asthma, upper respiratory tract infection within 4 weeks • Previous nasal surgery | • Mucin 5AC was significantly upregulated in NLF-derived exosomes of CRSwNP patients. • The expression of mucin 5AC was increased in the tissue specimens of the CRSwNP patients. • CRSsNP-derived fibroblasts treated with the mucin 5AC-enriched exosomes had a significantly increased level of VEGF, COX-2, and MMP-9 but not MMP-2 in comparison to those treated with mucin 5AC-deficient exosomes. |
| Shimizu et al. (2022) [21] | • NPFs cultured from nasal polyp tissue specimens | • ECRSwNP (n = 3) • NPFs incubated with GW4869 and DMSO • NPFs incubated with DMA and DMSO | • NPFs incubated with DMSO | NR | • Interaction of NPFs and peripheral blood eosinophils or EoL-1 cells stimulated the release of exosomes and VEGF. • NPF-derived EVs significantly stimulated VEGF release from EoL-1 cells, whereas cultured NPF-derived EVs alone did not produce VEGF for 24 h. • Pretreatment of NPFs with GW4869 or DMA attenuated the release of exosomes and VEGF from cocultured EoL-1 cells and NPFs. |
| He et al. (2022) [13] | • plasma | • CRSwNP (n = 5) | • HS (n = 5) | • Exposition to antibiotics, systemic /topical steroids or antileukotrienes in last 3 months • Among the controls additionally allergy or asthma | • 1692 known miRNAs and 1068 novel miRNAs were identified in plasma-derived exosomes. • 159 plasma exosomal miRNAs were differentially expressed (93 upregulated and 66 downregulated) by miRNA sequencing in CRSwNP relative to the HS group. • The top three upregulated miRNAs were novel_miRNA_677, novel_miRNA_1037, and novel_miRNA_79. • The top three downregulated miRNAs were novel_miRNA_192, novel_miRNA_1022, and novel_miRNA_4. • Corresponding differentially expressed target genes in the GO and KEGG analyses revealed: axon guidance, extracellular matrix (ECM)-receptor interaction, protein digestion and absorption, the calcium, the Hippo, the Notch, the ErbB, the cAMP signaling pathway, and focal adhesion. |
| Nocera et al. (2022) [20] | • NM | • CRSwNP (n = 10) • the recombinant CST-1 mouse model with ACSC1a knockdown (1 mg/kg or 0.1 mg/kg) | • HS (n = 10) • the recombinant CST-1 mouse model without ABCB1a knockdown | • Exposition to antibiotics or any topical /systemic steroids in last 4 weeks • Ciliary dysfunction, autoimmune disease, CF, immunodeficiency • Among the controls additionally the presence of environmental allergy or asthma | • CST-1 and CST-2 are among the most overexpressed protease inhibitors in tissue, mucus-derived exosome and mucus samples in CRSwNP patients in relation to HS. • Exosomal CST-1 and CST-2 were strongly and significantly correlated with tissue eosinophils and Lund-Mackay scores. • Exposure to CST-1 induced type 2 cytokine secretion and was abrogated by epithelial knockdown of ABCB1a, which encodes P-glycoprotein. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dżaman, K.; Czerwaty, K. Roles of Exosomes in Chronic Rhinosinusitis: A Systematic Review. Int. J. Mol. Sci. 2022, 23, 11284. https://doi.org/10.3390/ijms231911284
Dżaman K, Czerwaty K. Roles of Exosomes in Chronic Rhinosinusitis: A Systematic Review. International Journal of Molecular Sciences. 2022; 23(19):11284. https://doi.org/10.3390/ijms231911284
Chicago/Turabian StyleDżaman, Karolina, and Katarzyna Czerwaty. 2022. "Roles of Exosomes in Chronic Rhinosinusitis: A Systematic Review" International Journal of Molecular Sciences 23, no. 19: 11284. https://doi.org/10.3390/ijms231911284
APA StyleDżaman, K., & Czerwaty, K. (2022). Roles of Exosomes in Chronic Rhinosinusitis: A Systematic Review. International Journal of Molecular Sciences, 23(19), 11284. https://doi.org/10.3390/ijms231911284