
Nobiletin Prevents D-Galactose-Induced C2C12 Cell Aging by Improving Mitochondrial Function
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
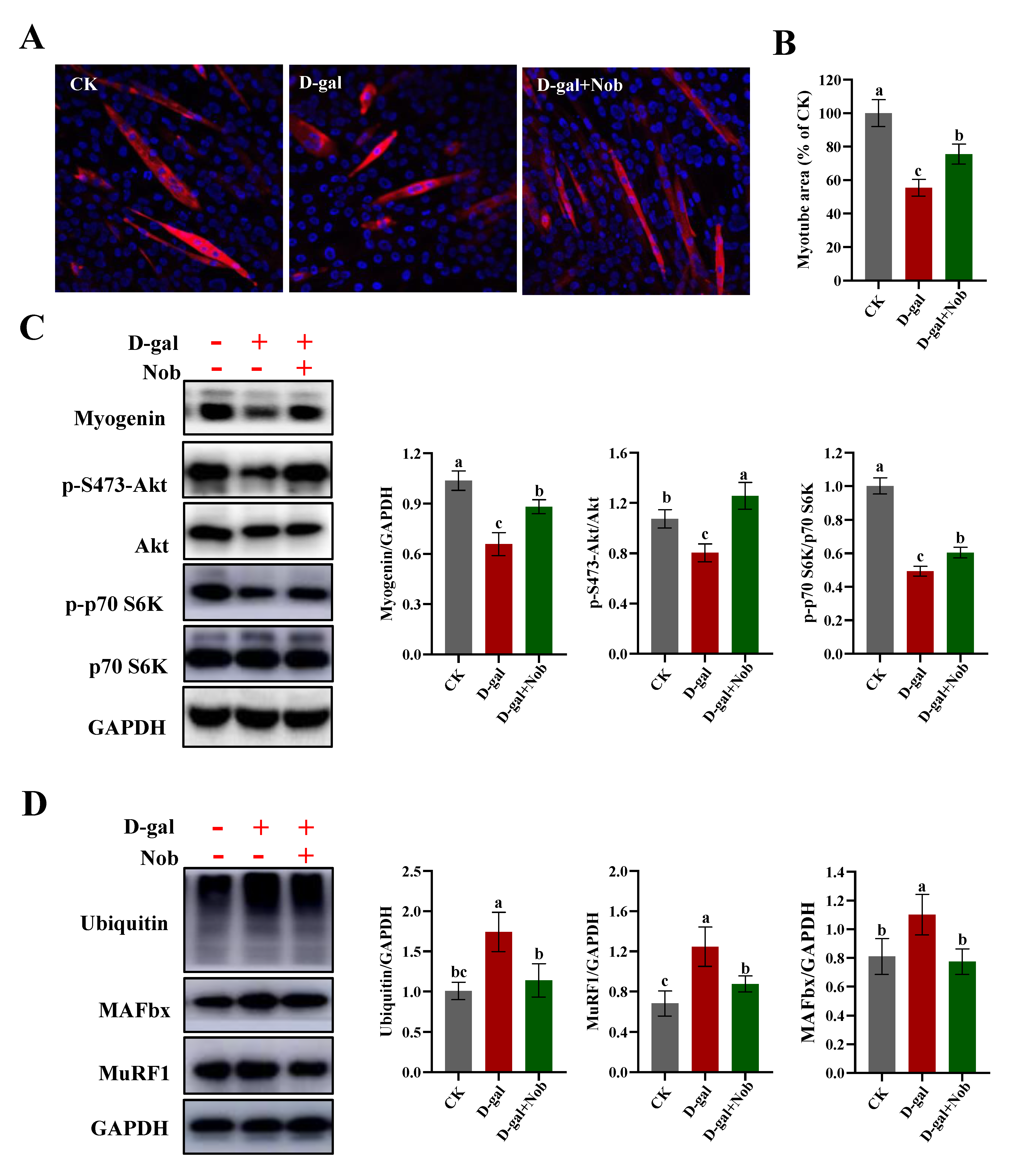
2.1. Effect of Nobiletin (Nob) on D-Galactose-Induced (D-Gal-Induced) Atrophy of Skeletal Muscle
2.2. Effect of Nob on Senescent Cells and Senescence Markers in D-Gal-Induced C2C12 Cells
2.3. Effect of Nob on ROS and Inflammation in D-Gal-Induced C2C12 Cells
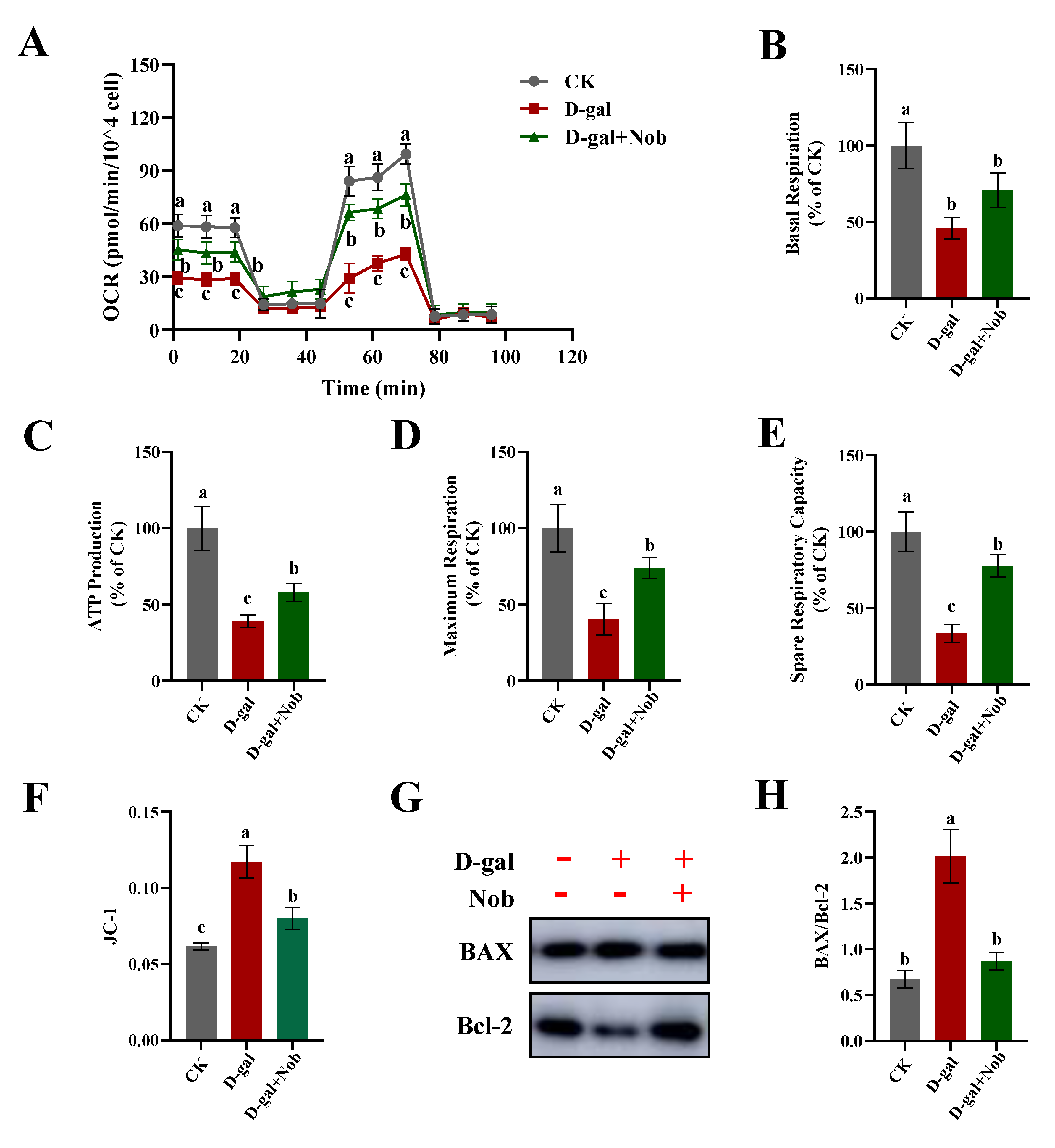
2.4. Effect of Nob on Mitochondrial Function in D-Gal-Induced C2C12 Cells
2.5. Effect of Nob on Autophagy in D-Gal-Induced C2C12 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Treatment
4.3. Cell Viability Assay
4.4. Senescence-Associated β-Galactosidase (SA-β-Gal) Activity
4.5. Immunofluorescence Staining
4.6. RT-qPCR: RNA Extraction, cDNA Synthesis, and PCR
4.7. Western Blotting
4.8. ROS Measurements
4.9. Seahorse Mitochondrial Oxygen Consumption Rate Measurements
4.10. JC-1 Measurements
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rafi, S.T.M.; Sambandam, Y.; Sittadjody, S.; Pathak, S.; Ramachandran, I.; Kumaran, R.I. Chapter 9—Skeletal muscle cell aging and stem cells. In Stem Cells and Aging; Pathak, S., Banerjee, A., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 125–145. [Google Scholar]
- Chang, Y.C.; Liu, H.W.; Chen, Y.T.; Chen, Y.A.; Chen, Y.J.; Chang, S.J. Resveratrol protects muscle cells against palmitate-induced cellular senescence and insulin resistance through ameliorating autophagic flux. J. Food Drug Anal. 2018, 26, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Nohara, K.; Mallampalli, V.; Nemkov, T.; Wirianto, M.; Yang, J.; Ye, Y.Q.; Sun, Y.X.; Han, L.; Esser, K.A.; Mileykovskaya, E.; et al. Nobiletin fortifies mitochondrial respiration in skeletal muscle to promote healthy aging against metabolic challenge. Nat. Commun. 2019, 10, 3923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.C.; Cheng, N.X.; Qiao, B.K.; Zhang, F.; Wu, J.; Liu, C.; Li, Y.L.; Du, J. Age-related decline of interferon-gamma responses in macrophage impairs satellite cell proliferation and regeneration. J. Cachexia Sarcopeni 2020, 11, 1291–1305. [Google Scholar] [CrossRef] [PubMed]
- Paris, M.T.; Bell, K.E.; Avrutin, E.; Mourtzakis, M. Associations between skeletal muscle echo intensity and thickness in relation to glucose homeostasis in healthy and glucose impaired older males. Exp. Gerontol. 2021, 154, 111547. [Google Scholar] [CrossRef] [PubMed]
- Romanello, V.; Sandri, M. Mitochondrial Quality Control and Muscle Mass Maintenance. Front. Physiol. 2016, 6, 422. [Google Scholar] [CrossRef]
- Xu, Q.X.; Fu, Q.; Li, Z.; Liu, H.X.; Wang, Y.; Lin, X.; He, R.K.; Zhang, X.G.; Ju, Z.Y.; Campisi, J.; et al. The flavonoid procyanidin C1 has senotherapeutic activity and increases lifespan in mice. Nat. Metab. 2021, 3, 1706–1726. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [Green Version]
- He, S.H.; Sharpless, N.E. Senescence in Health and Disease. Cell 2017, 169, 1000–1011. [Google Scholar] [CrossRef] [Green Version]
- Chapman, J.; Fielder, E.; Passos, J.F. Mitochondrial dysfunction and cell senescence: Deciphering a complex relationship. Febs. Lett. 2019, 593, 1566–1579. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, L.H.; Wang, Z.G.; Liu, J.P. Roles of Telomere Biology in Cell Senescence, Replicative and Chronological Ageing. Cells 2019, 8, 54. [Google Scholar] [CrossRef]
- Davalli, P.; Mitic, T.; Caporali, A.; Lauriola, A.; D’Arca, D. ROS, Cell Senescence, and Novel Molecular Mechanisms in Aging and Age-Related Diseases. Oxid. Med. Cell Longev. 2016, 2016, 3565127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasa, S.M.M.; Annunziata, F.; Krepelova, A.; Nunna, S.; Omrani, O.; Gebert, N.; Adam, L.; Käppel, S.; Höhn, S.; Donati, G.; et al. Inflammaging is driven by upregulation of innate immune receptors and systemic interferon signaling and is ameliorated by dietary restriction. Cell Rep. 2022, 39, 111017. [Google Scholar] [CrossRef]
- von Zglinicki, T. Oxidative stress shortens telomeres. Trends Biochem. Sci. 2002, 27, 339–344. [Google Scholar] [CrossRef]
- Morrow, N.M.; Trzaskalski, N.A.; Hanson, A.A.; Fadzeyeva, E.; Telford, D.E.; Chhoker, S.S.; Sutherland, B.G.; Edwards, J.Y.; Huff, M.W.; Mulvihill, E.E. Nobiletin Prevents High-Fat Diet-Induced Dysregulation of Intestinal Lipid Metabolism and Attenuates Postprandial Lipemia. Arter. Thromb. Vasc. Biol. 2022, 42, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Bunbupha, S.; Pakdeechote, P.; Maneesai, P.; Prasarttong, P. Nobiletin alleviates high-fat diet-induced nonalcoholic fatty liver disease by modulating AdipoR1 and gp91(phox) expression in rats. J. Nutr. Biochem. 2021, 87, 108526. [Google Scholar] [CrossRef]
- Wang, H.D.; Guo, Y.J.; Qiao, Y.; Zhang, J.; Jiang, P. Nobiletin Ameliorates NLRP3 Inflammasome-Mediated Inflammation Through Promoting Autophagy via the AMPK Pathway. Mol. Neurobiol. 2020, 57, 5056–5068. [Google Scholar] [CrossRef]
- Azman, K.F.; Zakaria, R. D-Galactose-induced accelerated aging model: An overview. Biogerontology 2019, 20, 763–782. [Google Scholar] [CrossRef]
- Kou, X.; Li, J.; Liu, X.; Yang, X.; Fan, J.; Chen, N. Ampelopsin attenuates the atrophy of skeletal muscle from d-gal-induced aging rats through activating AMPK/SIRT1/PGC-1α signaling cascade. Biomed. Pharmacother. 2017, 90, 311–320. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Chen, Y.F.; Xu, X.B.; Xiang, H.Y.; Shi, Y.Z.; Gao, Y.; Wang, X.W.; Jiang, X.F.; Li, N.; Pan, J.P. Autophagy inhibits the mesenchymal stem cell aging induced by D-galactose through ROS/JNK/p38 signalling. Clin. Exp. Pharm. Physiol. 2020, 47, 466–477. [Google Scholar] [CrossRef]
- Chen, Q.N.; Fan, Z.; Lyu, A.K.; Wu, J.; Guo, A.; Yang, Y.F.; Chen, J.L.; Xiao, Q. Effect of sarcolipin-mediated cell transdifferentiation in sarcopenia-associated skeletal muscle fibrosis. Exp. Cell Res. 2020, 389, 111890. [Google Scholar] [CrossRef]
- Yin, H.; Price, F.; Rudnicki, M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013, 93, 23–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.H.; Song, Y.Y.; Liu, Z.Q.; Li, M.H.; Zhang, L.; Yu, Q.L.; Guo, Z.B.; Wei, J.M. Effects of iron-catalyzed and metmyoglobin oxidizing systems on biochemical properties of yak muscle myofibrillar protein. Meat Sci. 2020, 166, 108041. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.W.; Liu, J.; Sun, Z.C.; Yin, Y.J.; Mao, Y.; Xu, D.Q.; Liu, L.; Xu, Z.S.; Guo, Q.Q.; Ding, C.Y.; et al. AMPK-dependent and -independent coordination of mitochondrial function and muscle fiber type by FNIP1. PLoS Genet. 2021, 17, e1009488. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.H.; Chen, B.H.; Chiang, H.S.; Chen, C.W.; Chen, M.F.; Ke, C.C.; Wang, Y.Y.; Lin, W.N.; Wang, C.C.; Lin, Y.H. Optimizing a Male Reproductive Aging Mouse Model by D-Galactose Injection. Int. J. Mol. Sci. 2016, 17, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passos, J.F.; Saretzki, G.; Ahmed, S.; Nelson, G.; Richter, T.; Peters, H.; Wappler, I.; Birket, M.J.; Harold, G.; Schaeuble, K.; et al. Mitochondrial dysfunction accounts for the stochastic heterogeneity in telomere-dependent senescence. PLoS Biol. 2007, 5, 1138–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Wang, J.H.; Jiang, J.Q.; Zhang, C.M.; Zhao, M.; Chen, Z.; Wang, N.; Hu, D.D.; Liu, X.Y.; Peng, H.S.; et al. Sonodynamic therapy in atherosclerosis by curcumin nanosuspensions: Preparation design, efficacy evaluation, and mechanisms analysis. Eur. J. Pharm. Biopharm. 2020, 146, 101–110. [Google Scholar] [CrossRef]
- Feng, W.J.; Liu, J.Y.; Wang, S.; Hu, Y.; Pan, H.; Hu, T.; Guan, H.S.; Zhang, D.F.; Mao, Y.J. Alginate oligosaccharide alleviates D-galactose-induced cardiac ageing via regulating myocardial mitochondria function and integrity in mice. J. Cell Mol. Med. 2021, 25, 7157–7168. [Google Scholar] [CrossRef]
- El-Far, A.H.; Lebda, M.A.; Noreldin, A.E.; Atta, M.S.; Elewa, Y.H.A.; Elfeky, M.; Mousa, S.A. Quercetin Attenuates Pancreatic and Renal D-Galactose-Induced Aging-Related Oxidative Alterations in Rats. Int. J. Mol. Sci. 2020, 21, 4348. [Google Scholar] [CrossRef]
- Denison, H.J.; Cooper, C.; Sayer, A.A.; Robinson, S.M. Prevention and optimal management of sarcopenia: A review of combined exercise and nutrition interventions to improve muscle outcomes in older people. Clin. Interv. Aging 2015, 10, 859–869. [Google Scholar]
- Kwak, J.Y.; Kwon, K.-S. Pharmacological Interventions for Treatment of Sarcopenia: Current Status of Drug Development for Sarcopenia. Ann. Geriatr. Med. Res. 2019, 23, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Pal, R.; Palmieri, M.; Loehr, J.A.; Li, S.; Abo-Zahrah, R.; Monroe, T.O.; Thakur, P.B.; Sardiello, M.; Rodney, G.G. Src-dependent impairment of autophagy by oxidative stress in a mouse model of Duchenne muscular dystrophy. Nat. Commun. 2014, 5, 4425. [Google Scholar] [CrossRef] [PubMed]
- Bechara, L.R.G.; Moreira, J.B.N.; Jannig, P.R.; Voltarelli, V.A.; Dourado, P.M.; Vasconcelos, A.R.; Scavone, C.; Ramires, P.R.; Brum, P.C. NADPH oxidase hyperactivity induces plantaris atrophy in heart failure rats. Int. J. Cardiol. 2014, 175, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Jeyapalan, J.C.; Sedivy, J.M. Cellular senescence and organismal aging. Mech. Ageing Dev. 2008, 129, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Di Leonardo, A.; Linke, S.P.; Clarkin, K.; Wahl, G.M. DNA damage triggers a prolonged p53-dependent G1 arrest and long-term induction of Cip1 in normal human fibroblasts. Genes Dev. 1994, 8, 2540–2551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passos, J.F.; Nelson, G.; Wang, C.F.; Richter, T.; Simillion, C.; Proctor, C.J.; Miwa, S.; Olijslagers, S.; Hallinan, J.; Wipat, A.; et al. Feedback between p21 and reactive oxygen production is necessary for cell senescence. Mol. Syst. Biol. 2010, 6, 347. [Google Scholar] [CrossRef]
- Chinnery, P.F.; Samuels, D.C.; Elson, J.; Turnbull, D.M. Accumulation of mitochondrial DNA mutations in ageing, cancer, and mitochondrial disease: Is there a common mechanism? Lancet 2002, 360, 1323–1325. [Google Scholar] [CrossRef]
- Garcia-Prat, L.; Martinez-Vicente, M.; Perdiguero, E.; Ortet, L.; Rodriguez-Ubreva, J.; Rebollo, E.; Ruiz-Bonilla, V.; Gutarra, S.; Ballestar, E.; Serrano, A.L.; et al. Autophagy maintains stemness by preventing senescence. Nature 2016, 529, 37–42. [Google Scholar] [CrossRef]
- Masiero, E.; Agatea, L.; Mammucari, C.; Blaauw, B.; Loro, E.; Komatsu, M.; Metzger, D.; Reggiani, C.; Schiaffino, S.; Sandri, M. Autophagy Is Required to Maintain Muscle Mass. Cell Metab. 2009, 10, 507–515. [Google Scholar] [CrossRef]
- Raben, N.; Hill, V.; Shea, L.; Takikita, S.; Baum, R.; Mizushima, N.; Ralston, E.; Plotz, P. Suppression of autophagy in skeletal muscle uncovers the accumulation of ubiquitinated proteins and their potential role in muscle damage in Pompe disease. Hum. Mol. Genet. 2008, 17, 3897–3908. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, R.A.; Thomas, A. Mitophagy and Mitochondrial Quality Control Mechanisms in the Heart. Curr. Pathobiol. Rep. 2017, 5, 161–169. [Google Scholar] [CrossRef]
- Sun, Y.N.; Yang, Z.X.; Ren, F.Z.; Fang, B. FGF19 alleviates palmitate-induced atrophy in C2C12 cells by inhibiting mitochondrial overload and insulin resistance. Int. J. Biol. Macromol. 2020, 158, 401–407. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.-H.; Sun, Y.-N.; Qu, T.-Q.; Sang, X.-Q.; Zhou, L.-M.; Li, Y.-X.; Ren, F.-Z. Nobiletin Prevents D-Galactose-Induced C2C12 Cell Aging by Improving Mitochondrial Function. Int. J. Mol. Sci. 2022, 23, 11963. https://doi.org/10.3390/ijms231911963
Wang H-H, Sun Y-N, Qu T-Q, Sang X-Q, Zhou L-M, Li Y-X, Ren F-Z. Nobiletin Prevents D-Galactose-Induced C2C12 Cell Aging by Improving Mitochondrial Function. International Journal of Molecular Sciences. 2022; 23(19):11963. https://doi.org/10.3390/ijms231911963
Chicago/Turabian StyleWang, Hui-Hui, Ya-Nan Sun, Tai-Qi Qu, Xue-Qin Sang, Li-Mian Zhou, Yi-Xuan Li, and Fa-Zheng Ren. 2022. "Nobiletin Prevents D-Galactose-Induced C2C12 Cell Aging by Improving Mitochondrial Function" International Journal of Molecular Sciences 23, no. 19: 11963. https://doi.org/10.3390/ijms231911963
APA StyleWang, H. -H., Sun, Y. -N., Qu, T. -Q., Sang, X. -Q., Zhou, L. -M., Li, Y. -X., & Ren, F. -Z. (2022). Nobiletin Prevents D-Galactose-Induced C2C12 Cell Aging by Improving Mitochondrial Function. International Journal of Molecular Sciences, 23(19), 11963. https://doi.org/10.3390/ijms231911963