
Melatonin Promotes Antler Growth by Accelerating MT1-Mediated Mesenchymal Cell Differentiation and Inhibiting VEGF-Induced Degeneration of Chondrocytes


Abstract
:1. Introduction
2. Results
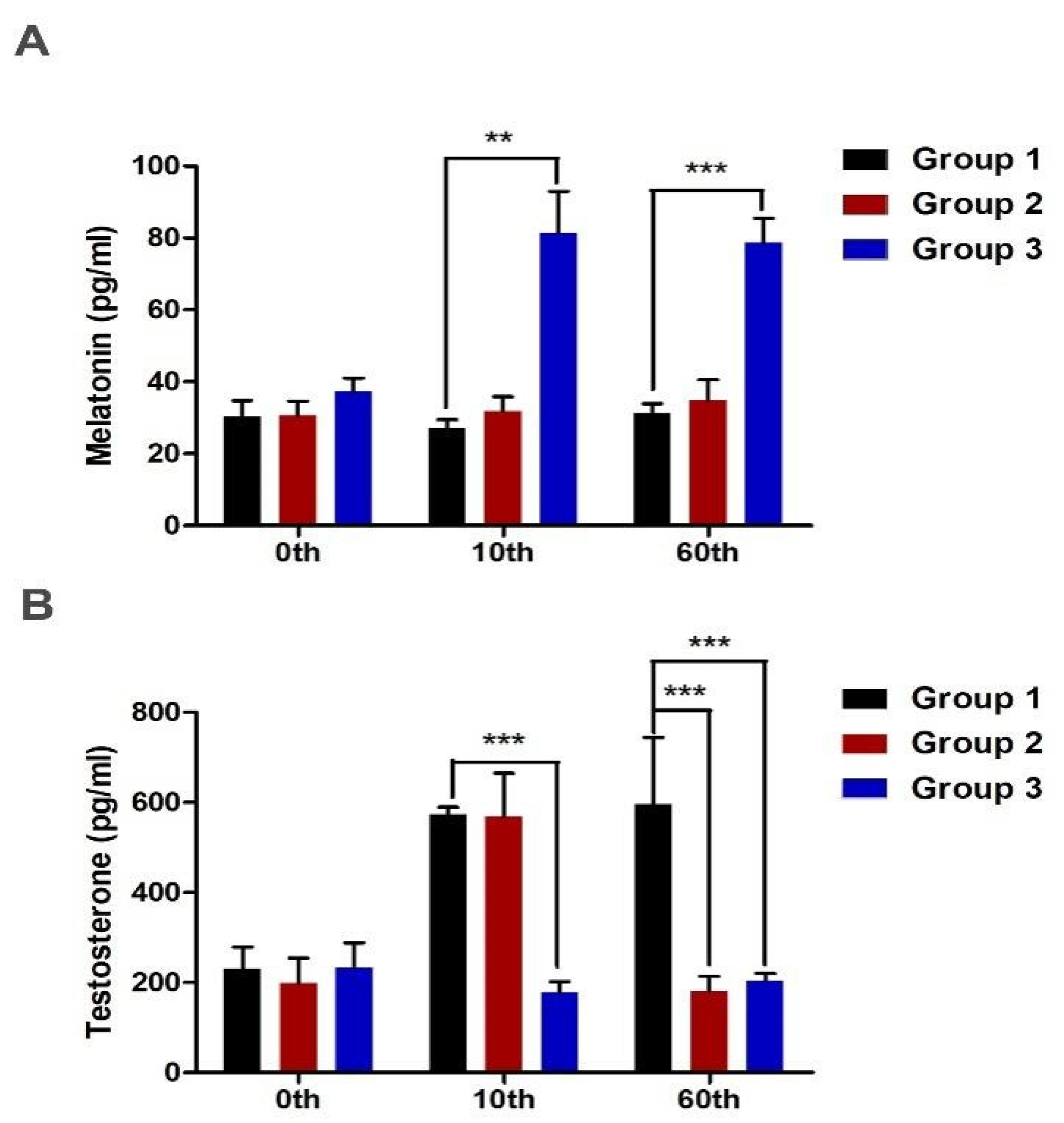
2.1. Subcutaneous Implantation of Melatonin and Endocrinological Profiles
2.2. Effect of Melatonin Implantation on Antler Growth
2.3. MLT Accelerates Antler Mesenchymal Cells Differentiation In Vitro
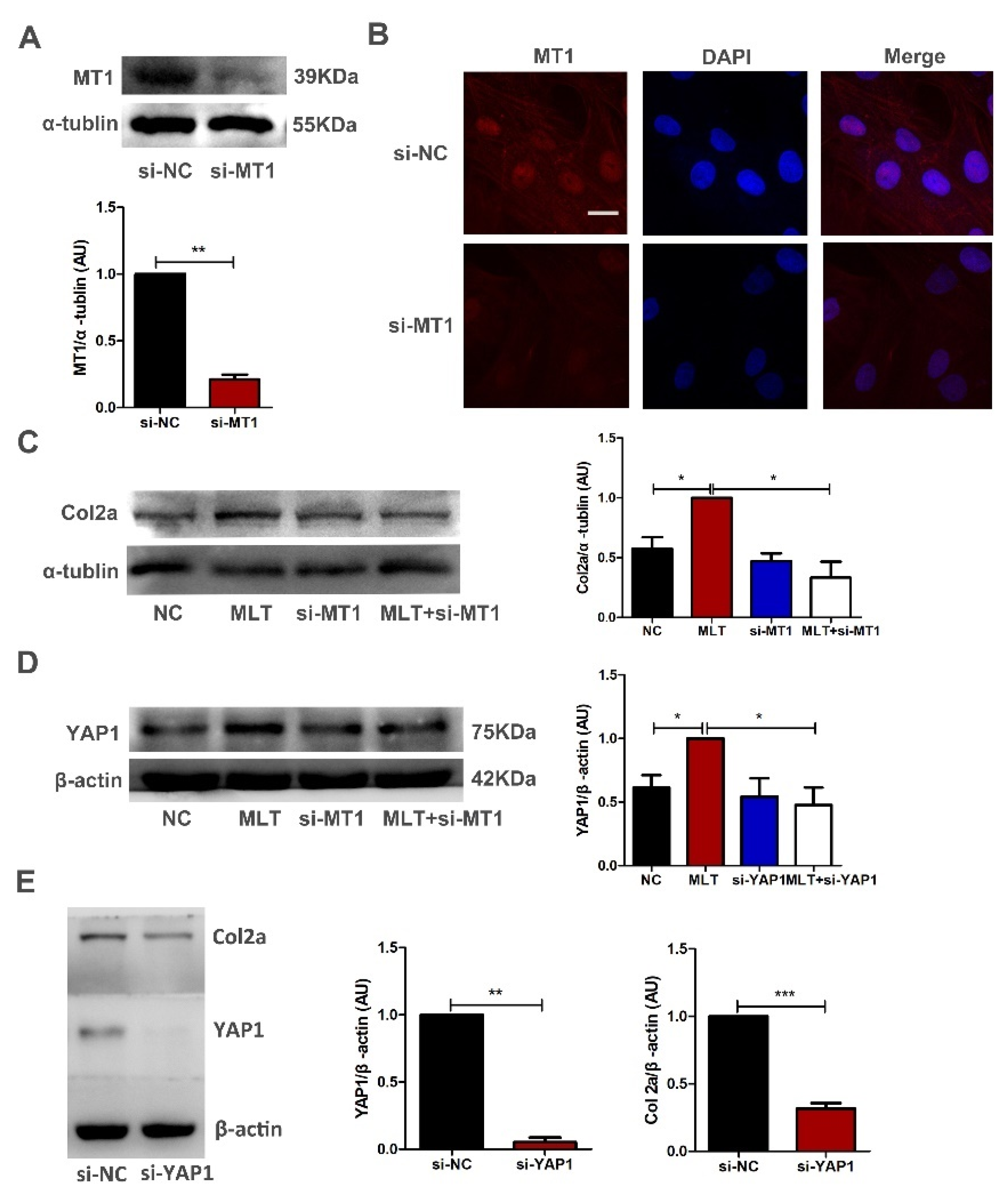
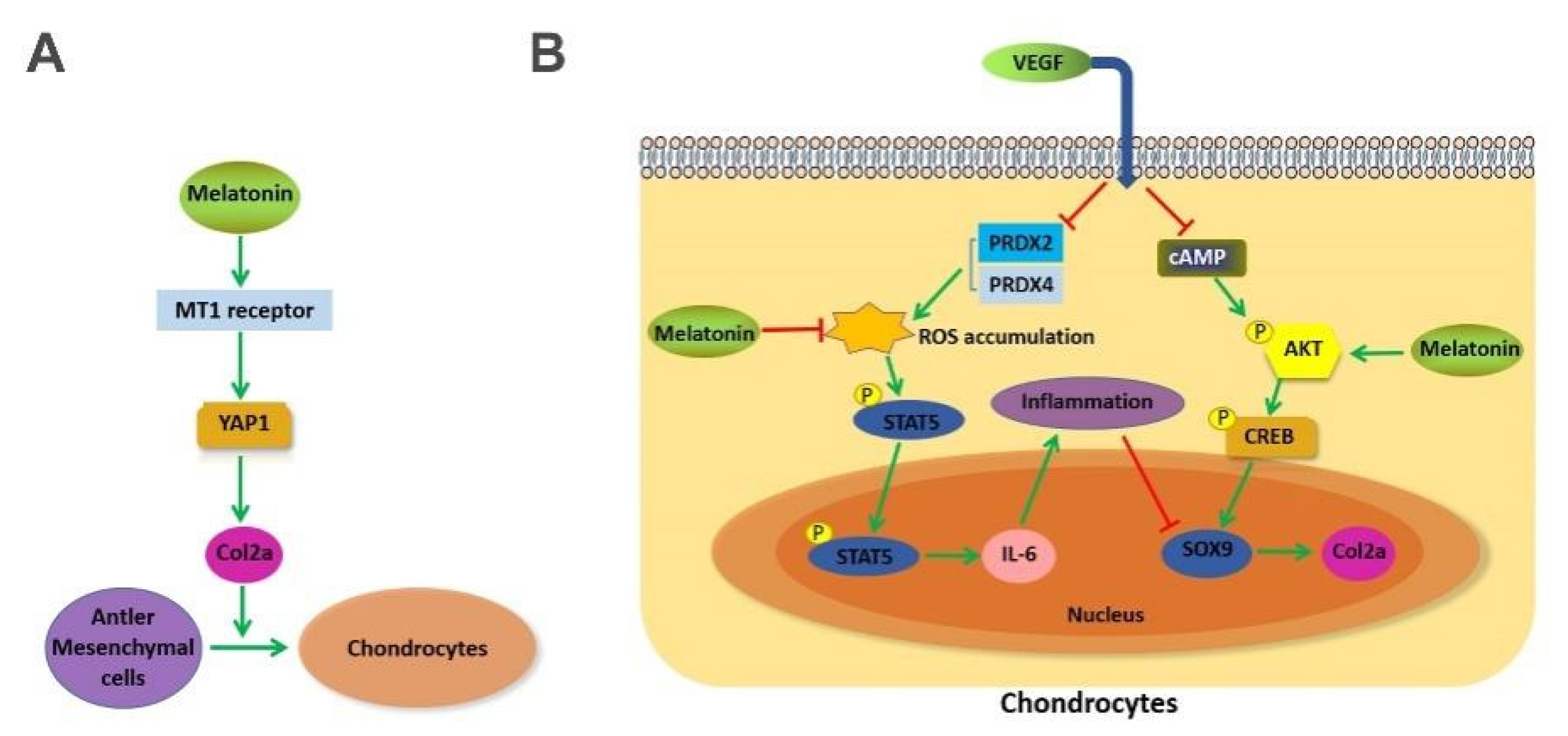
2.4. MLT Regulates the Expression of Col2a through the MT1 Binding Mediated YAP1 Pathway in Antler Mesenchymal Cells
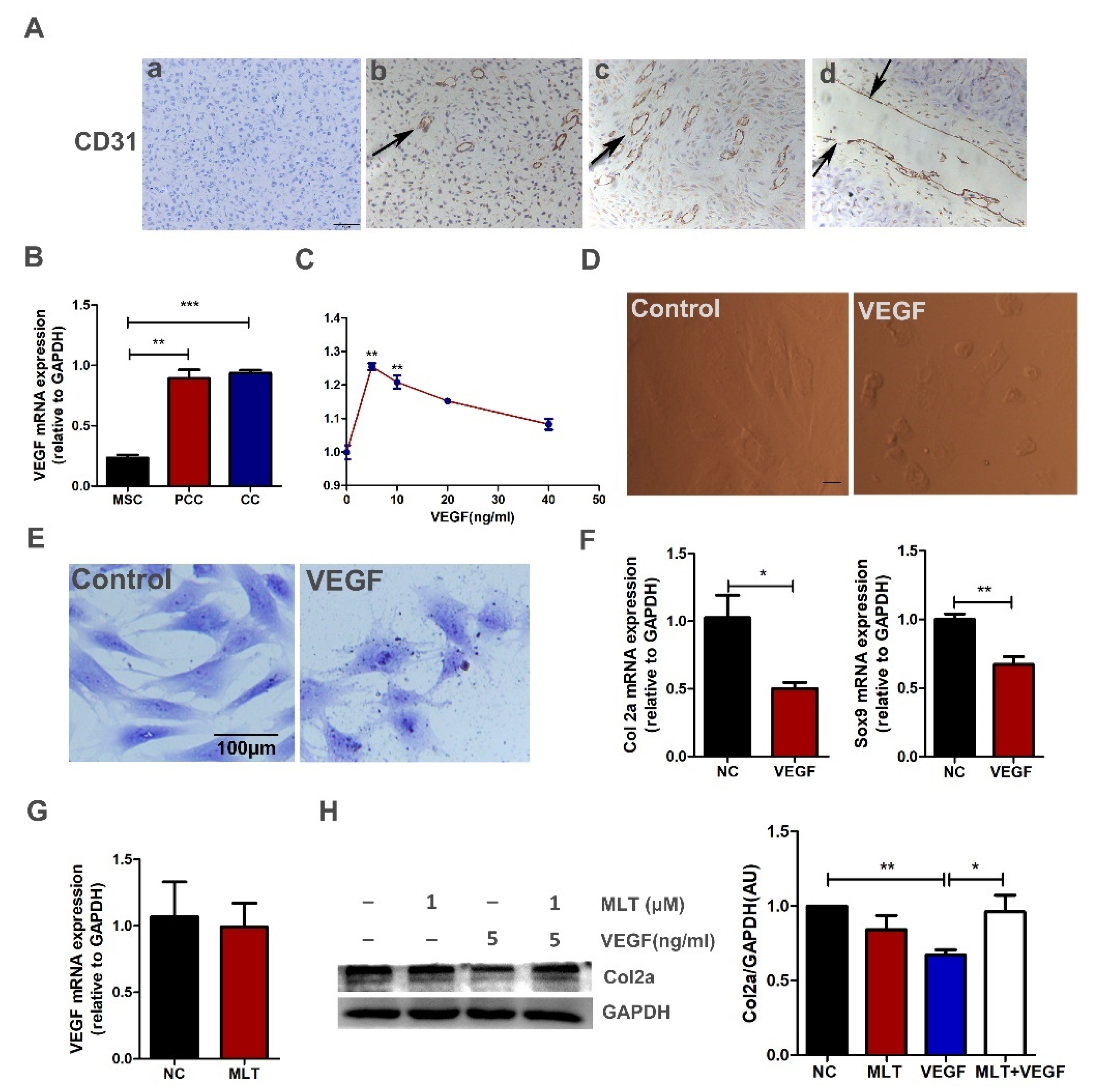
2.5. MLT Inhibits VEGF-Induced Degeneration of Antler Chondrocytes
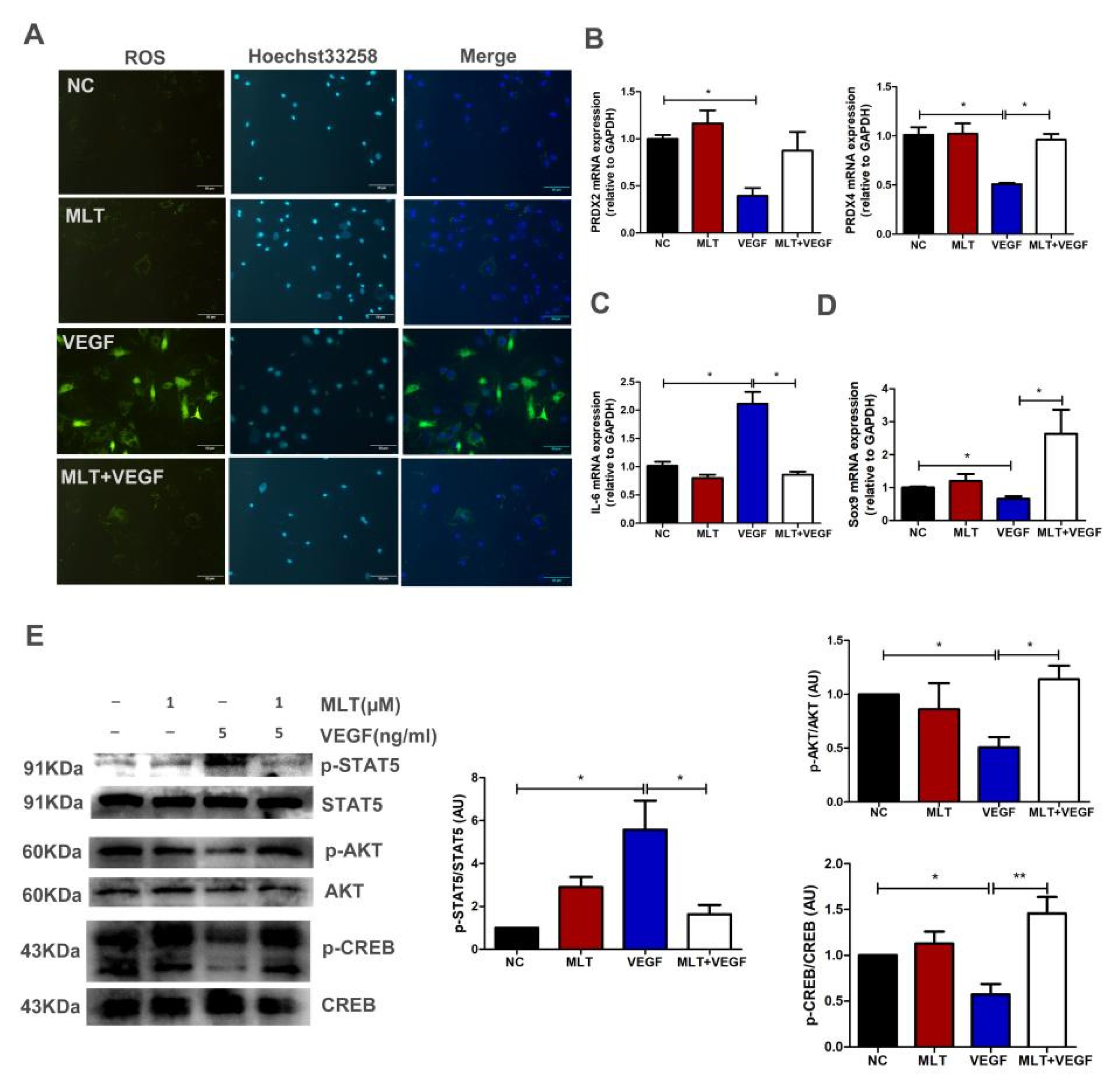
2.6. Effects of MLT on VEGF-Induced Chondrocytes Degeneration through Inhibition of p-STAT5/IL-6 and Activation of p-AKT/p-CREB Dependent on Sox9 Expression
3. Discussion
4. Materials and Methods
4.1. Subcutaneous Implantation of Melatonin
4.2. Detection of Hormone Level in Plasma
4.3. Cell Separation and Culture
4.4. Immunohistochemistry Staining
4.5. RNA Interference
4.6. Cell Viability Test
4.7. Total RNA and Quantitative Real-Time PCR (q-PCR)
4.8. Western Blot Assay
4.9. Reactive Oxygen Species
4.10. Immunofluorescence Assay
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Price, J.; Allen, S. Exploring the mechanisms regulating regeneration of deer antlers. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2004, 359, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhao, H.; Liu, Z.; McMahon, C. Deer antler—A novel model for studying organ regeneration in mammals. Int. J. Biochem. Cell Biol. 2014, 56, 111–122. [Google Scholar] [CrossRef]
- Wang, D.; Berg, D.; Ba, H.; Sun, H.; Wang, Z.; Li, C. Deer antler stem cells are a novel type of cells that sustain full regeneration of a mammalian organ-deer antler. Cell Death Dis. 2019, 10, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, C.; Wang, N.; Li, Z.; Heller, R.; Liu, R.; Zhao, Y.; Han, J.; Pan, X.; Zheng, Z.; et al. Genetic basis of ruminant headgear and rapid antler regeneration. Science 2019, 364, 6335. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; He, C.; Sun, X.; Wang, J.; Luo, C.; Liu, G.; Yang, L.; Xiong, J.; Huo, L. The Regulatory Mechanism of MLT/MT1 Signaling on the Growth of Antler Mesenchymal Cells. Molecules 2017, 22, 1793. [Google Scholar] [CrossRef] [Green Version]
- Skarlis, C.; Anagnostouli, M. The role of melatonin in Multiple Sclerosis. Neurol. Sci. Off. J. Ital. Neurol. Soc. Ital. Soc. Clin. Neurophysiol. 2020, 41, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Gan, L.; Luo, D.; Sun, C. Melatonin promotes circadian rhythm-induced proliferation through Clock/histone deacetylase 3/c-Myc interaction in mouse adipose tissue. J. Pineal Res. 2017, 62, e12383. [Google Scholar] [CrossRef]
- Zeman, M.; Herichova, I. Melatonin and clock genes expression in the cardiovascular system. Front. Biosci. 2013, 5, 743–753. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Patel, K.K.; Dehari, D.; Agrawal, A.K.; Singh, S. Melatonin and its ubiquitous anticancer effects. Mol. Cell. Biochem. 2019, 462, 133–155. [Google Scholar] [CrossRef]
- Luo, C.; Yang, Q.; Liu, Y.; Zhou, S.; Jiang, J.; Reiter, R.J.; Bhattacharya, P.; Cui, Y.; Yang, H.; Ma, H.; et al. The multiple protective roles and molecular mechanisms of melatonin and its precursor N-acetylserotonin in targeting brain injury and liver damage and in maintaining bone health. Free. Radic. Biol. Med. 2019, 130, 215–233. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Esteban-Zubero, E.; Zhou, Z.; Reiter, R.J. Melatonin as a Potent and Inducible Endogenous Antioxidant: Synthesis and Metabolism. Molecules 2015, 20, 18886–18906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, R.J.; Tan, D.X.; Rosales-Corral, S.; Galano, A.; Zhou, X.J.; Xu, B. Mitochondria: Central Organelles for Melatonin’s Antioxidant and Anti-Aging Actions. Molecules 2018, 23, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fathizadeh, H.; Mirzaei, H.; Asemi, Z. Melatonin: An anti-tumor agent for osteosarcoma. Cancer Cell Int. 2019, 19, 319. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.F.; Huo, L.J.; Yang, L.G.; Riaz, H.; Xiong, L.R.; Chen, J.G.; Zhang, S.J.; Xiong, J.J. Association between melatonin receptor 1A (MTNR1A) gene single-nucleotide polymorphisms and the velvet antler yield of Sika deer. Mol. Biol. Rep. 2014, 41, 1901–1906. [Google Scholar] [CrossRef] [PubMed]
- Malpaux, B.; Migaud, M.; Tricoire, H.; Chemineau, P. Biology of mammalian photoperiodism and the critical role of the pineal gland and melatonin. J. Biol. Rhythm. 2001, 16, 336–347. [Google Scholar] [CrossRef]
- Weis, S.M.; Cheresh, D.A. Tumor angiogenesis: Molecular pathways and therapeutic targets. Nat. Med. 2011, 17, 1359–1370. [Google Scholar] [CrossRef]
- Li, L.; Liu, F.; Huang, W.; Wang, J.; Yin, Z.J.B. Ricolinostat (ACY-1215) inhibits VEGF expression via PI3K/AKT pathway and promotes apoptosis in osteoarthritic osteoblasts. Biomed. Pharmacother. 2019, 118, 109357. [Google Scholar] [CrossRef]
- Murata, M.; Yudoh, K.; Masuko, K. The potential role of vascular endothelial growth factor (VEGF) in cartilage: How the angiogenic factor could be involved in the pathogenesis of osteoarthritis? Osteoarthr. Cartil. 2008, 16, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Nagao, M.; Hamilton, J.L.; Kc, R.; Berendsen, A.D.; Duan, X.; Cheong, C.W.; Li, X.; Im, H.J.; Olsen, B.R. Vascular Endothelial Growth Factor in Cartilage Development and Osteoarthritis. Sci. Rep. 2017, 7, 13027. [Google Scholar] [CrossRef]
- Xu, Y.; Lu, X.; Hu, Y.; Yang, B.; Tsui, C.K.; Yu, S.; Lu, L.; Liang, X. Melatonin attenuated retinal neovascularization and neuroglial dysfunction by inhibition of HIF-1α-VEGF pathway in oxygen-induced retinopathy mice. J. Pineal Res. 2018, 64, e12473. [Google Scholar] [CrossRef]
- Cheng, J.; Yang, H.L.; Gu, C.J.; Liu, Y.K.; Shao, J.; Zhu, R.; He, Y.Y.; Zhu, X.Y.; Li, M.Q. Melatonin restricts the viability and angiogenesis of vascular endothelial cells by suppressing HIF-1α/ROS/VEGF. Int. J. Mol. Med. 2019, 43, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Zhang, Y.; Ma, Y.; Du, J.; Gu, L.; Zheng, L.; Zhang, X. Effect of melatonin on EGF- and VEGF-induced monolayer permeability of HUVECs. Am. J. Physiology. Heart Circ. Physiol. 2019, 316, H1178–H1191. [Google Scholar] [CrossRef]
- Alcantara, M.B.; Dass, C.R. Pigment epithelium-derived factor as a natural matrix metalloproteinase inhibitor: A comparison with classical matrix metalloproteinase inhibitors used for cancer treatment. J. Pharm. Pharmacol. 2014, 66, 895–902. [Google Scholar] [CrossRef]
- Dupont, S. Role of YAP/TAZ in cell-matrix adhesion-mediated signalling and mechanotransduction. Exp. Cell Res. 2016, 343, 42–53. [Google Scholar] [CrossRef]
- Juhász, T.; Matta, C.; Somogyi, C.; Katona, É.; Takács, R.; Soha, R.F.; Szabó, I.A.; Cserháti, C.; Sződy, R.; Karácsonyi, Z.; et al. Mechanical loading stimulates chondrogenesis via the PKA/CREB-Sox9 and PP2A pathways in chicken micromass cultures. Cell. Signal. 2014, 26, 468–482. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, T.; Ogiwara, K. Roles of melatonin in the teleost ovary: A review of the current status. Comp. Biochem. Physiology. Part A Mol. Integr. Physiol. 2021, 254, 110907. [Google Scholar] [CrossRef]
- Zisapel, N. New perspectives on the role of melatonin in human sleep, circadian rhythms and their regulation. Br. J. Pharmacol. 2018, 175, 3190–3199. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Liu, Q.; Chen, Y.; Wang, X.; Ran, Z.; Fang, F.; Xiong, J.; Liu, G.; Li, X.; Yang, L.; et al. Melatonin delays ovarian aging in mice by slowing down the exhaustion of ovarian reserve. Commun. Biol. 2021, 4, 534. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Jozaki, M.; Tanabe, M.; Shirafuta, Y.; Mihara, Y.; Shinagawa, M.; Tamura, I.; Maekawa, R.; Sato, S.; Taketani, T.; et al. Importance of Melatonin in Assisted Reproductive Technology and Ovarian Aging. Int. J. Mol. Sci. 2020, 21, 1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, P.H.; Tung, Y.T.; Chen, H.Y.; Chiang, Y.F.; Hong, H.C.; Huang, K.C.; Hsu, S.P.; Huang, T.C.; Hsia, S.M. Melatonin activates cell death programs for the suppression of uterine leiomyoma cell proliferation. J. Pineal Res. 2020, 68, e12620. [Google Scholar] [CrossRef] [PubMed]
- Koçak, N.; Dönmez, H.; Yildirim, İ.H. Effects of melatonin on apoptosis and cell differentiation in MCF-7 derived cancer stem cells. Cell. Mol. Biol. 2018, 64, 56–61. [Google Scholar] [CrossRef]
- Kierdorf, U.; Kierdorf, H. Deer antlers-a model of mammalian appendage regeneration: An extensive review. Gerontology 2011, 57, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Molnár, A.; Gyurján, I.; Korpos, E.; Borsy, A.; Stéger, V.; Buzás, Z.; Kiss, I.; Zomborszky, Z.; Papp, P.; Deák, F.; et al. Identification of differentially expressed genes in the developing antler of red deer Cervus elaphus. Mol. Genet. Genom. MGG 2007, 277, 237–248. [Google Scholar] [CrossRef]
- Wang, B.; Wen, H.; Smith, W.; Hao, D.; He, B.; Kong, L. Regulation effects of melatonin on bone marrow mesenchymal stem cell differentiation. J. Cell. Physiol. 2019, 234, 1008–1015. [Google Scholar] [CrossRef]
- Dong, P.; Gu, X.; Zhu, G.; Li, M.; Ma, B.; Zi, Y. Melatonin Induces Osteoblastic Differentiation of Mesenchymal Stem Cells and Promotes Fracture Healing in a Rat Model of Femoral Fracture via Neuropeptide Y/Neuropeptide Y Receptor Y1 Signaling. Pharmacology 2018, 102, 272–280. [Google Scholar] [CrossRef]
- Zhu, G.; Ma, B.; Dong, P.; Shang, J.; Gu, X.; Zi, Y. Melatonin promotes osteoblastic differentiation and regulates PDGF/AKT signaling pathway. Cell Biol. Int. 2020, 44, 402–411. [Google Scholar] [CrossRef]
- Zhong, W.; Tian, K.; Zheng, X.; Li, L.; Zhang, W.; Wang, S.; Qin, J. Mesenchymal stem cell and chondrocyte fates in a multishear microdevice are regulated by Yes-associated protein. Stem Cells Dev. 2013, 22, 2083–2093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Cai, T.; Chen, K.; Wang, R.; Wang, J.; Cui, C.; Yuan, J.; Zhang, K.; Liu, Z.; Deng, Y.; et al. Kindlin-2 regulates mesenchymal stem cell differentiation through control of YAP1/TAZ. J. Cell Biol. 2018, 217, 1431–1451. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.Y.; Song, R.J.; Xu, B.B.; Xu, Y.; Wang, X.X.; Sun, H.Y.; Li, S.N.; Liu, S.Z.; Yu, M.X.; Yang, F.; et al. Melatonin promotes cardiomyocyte proliferation and heart repair in mice with myocardial infarction via miR-143-3p/Yap/Ctnnd1 signaling pathway. Acta Pharmacol. Sin. 2021, 42, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Kierdorf, U.; Li, C.; Price, J.S. Improbable appendages: Deer antler renewal as a unique case of mammalian regeneration. Semin. Cell Dev. Biol. 2009, 20, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M.Y.; Ahmad, N.; Haqqi, T.M. Oxidative stress and inflammation in osteoarthritis pathogenesis: Role of polyphenols. Biomed. Pharmacother. 2020, 129, 110452. [Google Scholar] [CrossRef]
- Lepetsos, P.; Papavassiliou, A.G. ROS/oxidative stress signaling in osteoarthritis. Biochim. Biophys. Acta 2016, 1862, 576–591. [Google Scholar] [CrossRef]
- Zhang, Z.; Lin, J.; Tian, N.; Wu, Y.; Zhou, Y.; Wang, C.; Wang, Q.; Jin, H.; Chen, T.; Nisar, M.; et al. Melatonin protects vertebral endplate chondrocytes against apoptosis and calcification via the Sirt1-autophagy pathway. J. Cell. Mol. Med. 2019, 23, 177–193. [Google Scholar] [CrossRef]
- Feng, K.; Chen, Z.; Pengcheng, L.; Zhang, S.; Wang, X. Quercetin attenuates oxidative stress-induced apoptosis via SIRT1/AMPK-mediated inhibition of ER stress in rat chondrocytes and prevents the progression of osteoarthritis in a rat model. J. Cell. Physiol. 2019, 234, 18192–18205. [Google Scholar] [CrossRef] [PubMed]
- Fei, J.; Liang, B.; Jiang, C.; Ni, H.; Wang, L. Luteolin inhibits IL-1β-induced inflammation in rat chondrocytes and attenuates osteoarthritis progression in a rat model. Biomed. Pharmacother. 2019, 109, 1586–1592. [Google Scholar] [CrossRef]
- Kang, N.J.; Han, S.C.; Kang, G.J.; Koo, D.H.; Koh, Y.S.; Hyun, J.W.; Lee, N.H.; Ko, M.H.; Kang, H.K.; Yoo, E.S. Diphlorethohydroxycarmalol inhibits interleukin-6 production by regulating NF-κB, STAT5 and SOCS1 in lipopolysaccharide-stimulated RAW264.7 cells. Mar. Drugs 2015, 13, 2141–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, A.; Naka, T.; Muta, T.; Takeuchi, O.; Akira, S.; Kawase, I.; Kishimoto, T. Suppressor of cytokine signaling-1 selectively inhibits LPS-induced IL-6 production by regulating JAK-STAT. Proc. Natl. Acad. Sci. USA 2005, 102, 17089–17094. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Shen, N.; Yuan, K.; Li, J.; Wu, H.; Zeng, Y.; Fox, J., III; Bansal, A.K.; Singh, B.B.; Gao, H.; et al. Caveolin-1 plays a critical role in host immunity against Klebsiella pneumoniae by regulating STAT5 and Akt activity. Eur. J. Immunol. 2012, 42, 1500–1511. [Google Scholar] [CrossRef]
- Krylatov, A.V.; Maslov, L.N.; Voronkov, N.S.; Boshchenko, A.A.; Popov, S.V.; Gomez, L.; Wang, H.; Jaggi, A.S.; Downey, J.M. Reactive Oxygen Species as Intracellular Signaling Molecules in the Cardiovascular System. Curr. Cardiol. Rev. 2018, 14, 290–300. [Google Scholar] [CrossRef]
- Lefebvre, V.; Dvir-Ginzberg, M. SOX9 and the many facets of its regulation in the chondrocyte lineage. Connect. Tissue Res. 2017, 58, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Kurakazu, I.; Akasaki, Y.; Hayashida, M.; Tsushima, H.; Goto, N.; Sueishi, T.; Toya, M.; Kuwahara, M.; Okazaki, K.; Duffy, T.; et al. FOXO1 transcription factor regulates chondrogenic differentiation through transforming growth factor β1 signaling. J. Biol. Chem. 2019, 294, 17555–17569. [Google Scholar] [CrossRef]
- Piera-Velazquez, S.; Hawkins, D.F.; Whitecavage, M.K.; Colter, D.C.; Stokes, D.G.; Jimenez, S.A. Regulation of the human SOX9 promoter by Sp1 and CREB. Exp. Cell Res. 2007, 313, 1069–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, H.; Tu, Y.; Ma, T.; Liu, X.; Wen, T.; Cai, M.; Xia, Z.; Mei, J. Lactoferrin Inhibits IL-1β-Induced Chondrocyte Apoptosis Through AKT1-Induced CREB1 Activation. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2015, 36, 2456–2465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Clark, D.E.; Lord, E.A.; Stanton, J.A.; Suttie, J.M. Sampling technique to discriminate the different tissue layers of growing antler tips for gene discovery. Anat. Rec. 2002, 268, 125–130. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number | Antler Weight (Kg) | Relative Weight | Weight Gain |
|---|---|---|---|---|
| Group 1 (0 mg/per) | 7 | 1.13 ± 0.05 b | 1 ± 0.04 b | - |
| Group 2 (40 mg/per) | 7 | 1.68 ± 0.19 a | 1.49 ± 0.16 a | 49% |
| Group 3 (80 mg/per) | 7 | 1.79 ± 0.51 a | 1.59 ± 0.45 a | 59% |
| Number | Group 1 (0 mg/per) | Group 2 (40 mg/per) | Group 3 (80 mg/per) |
|---|---|---|---|
| 1 | 0.54 ± 0.05 | 0.56 ± 0.05 | 0.58 ± 0.01 |
| 2 | 0.23 ± 0.05 | 0.26 ± 0.05 | 0.39 ± 0.11 |
| 3 | 0.32 ± 0.11 | 0.37 ± 0.07 | 0.36 ± 0.16 |
| 4 | 0.16 ± 0.03 | 0.23 ± 0.15 | 0.27 ± 0.09 |
| 5 | 0.15 ± 0.05 | 0.16 ± 0.07 | 0.22 ± 0.09 |
| 6 | 0.07 ± 0.03 | 0.1 ± 0.05 | 0.13 ± 0.08 |
| 7 | 0.15 ± 0.06 | 0.13 ± 0.04 | 0.18 ± 0.07 |
| Parameters | Group 1 | Group 2 | Group 3 |
|---|---|---|---|
| Number | 7 | 7 | 7 |
| Melatonin dosage (mg/per) | 0 | 40 | 80 |
| Gene | Primer Sequence (5′-3′) | Product Size |
|---|---|---|
| PRDX2 | F: ACATTCCCCTGCTGGCTGAT | 267 bp |
| R: CGTCCACATTGGGCTTGATT | ||
| PRDX4 | F: TGATTCACAGTTCACCCATTTG | 214 bp |
| R:CACGGGAAGGTCATTCAGAGTA | ||
| IL6 | F: GCATTCCCTCCTCTGGTCA | 228 bp |
| R: AAAACATTCAAGCCGCACA | ||
| Col2a | F: GAGGCAGCCGGCAACCTGAG | 118 bp |
| R: TGCGAGCTGGGTTCTTGCGG | ||
| Sox-9 | F: ACGCAGATTCCCAAGACACTA | 143 bp |
| R: ACGCTCGCTTTGAAGGTTT | ||
| VEGF-A | F: AATGACGAAAGTCTGGAGTG | 120 bp |
| R: TTTGTTATGCTGTAGGAAGC | ||
| GAPDH | F: GAAGGGTGGCGCCAAGAGGG | 142 bp |
| R: GGGGGCCAAGCAGTTGGTGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Gu, X.; Li, K.; Li, M.; Peng, J.; Zhang, X.; Yang, L.; Xiong, J. Melatonin Promotes Antler Growth by Accelerating MT1-Mediated Mesenchymal Cell Differentiation and Inhibiting VEGF-Induced Degeneration of Chondrocytes. Int. J. Mol. Sci. 2022, 23, 759. https://doi.org/10.3390/ijms23020759
Sun X, Gu X, Li K, Li M, Peng J, Zhang X, Yang L, Xiong J. Melatonin Promotes Antler Growth by Accelerating MT1-Mediated Mesenchymal Cell Differentiation and Inhibiting VEGF-Induced Degeneration of Chondrocytes. International Journal of Molecular Sciences. 2022; 23(2):759. https://doi.org/10.3390/ijms23020759
Chicago/Turabian StyleSun, Xuyang, Xiaoying Gu, Keke Li, Mengqi Li, Jingna Peng, Xinxin Zhang, Liguo Yang, and Jiajun Xiong. 2022. "Melatonin Promotes Antler Growth by Accelerating MT1-Mediated Mesenchymal Cell Differentiation and Inhibiting VEGF-Induced Degeneration of Chondrocytes" International Journal of Molecular Sciences 23, no. 2: 759. https://doi.org/10.3390/ijms23020759
APA StyleSun, X., Gu, X., Li, K., Li, M., Peng, J., Zhang, X., Yang, L., & Xiong, J. (2022). Melatonin Promotes Antler Growth by Accelerating MT1-Mediated Mesenchymal Cell Differentiation and Inhibiting VEGF-Induced Degeneration of Chondrocytes. International Journal of Molecular Sciences, 23(2), 759. https://doi.org/10.3390/ijms23020759