
Accumulation of Cytochrome b558 at the Plasma Membrane: Hallmark of Oxidative Stress in Phagocytic Cells

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effect of Gamma Irradiation on PLB Cell Survival
2.2. ROS Production by Irradiated PLB-985 Cells
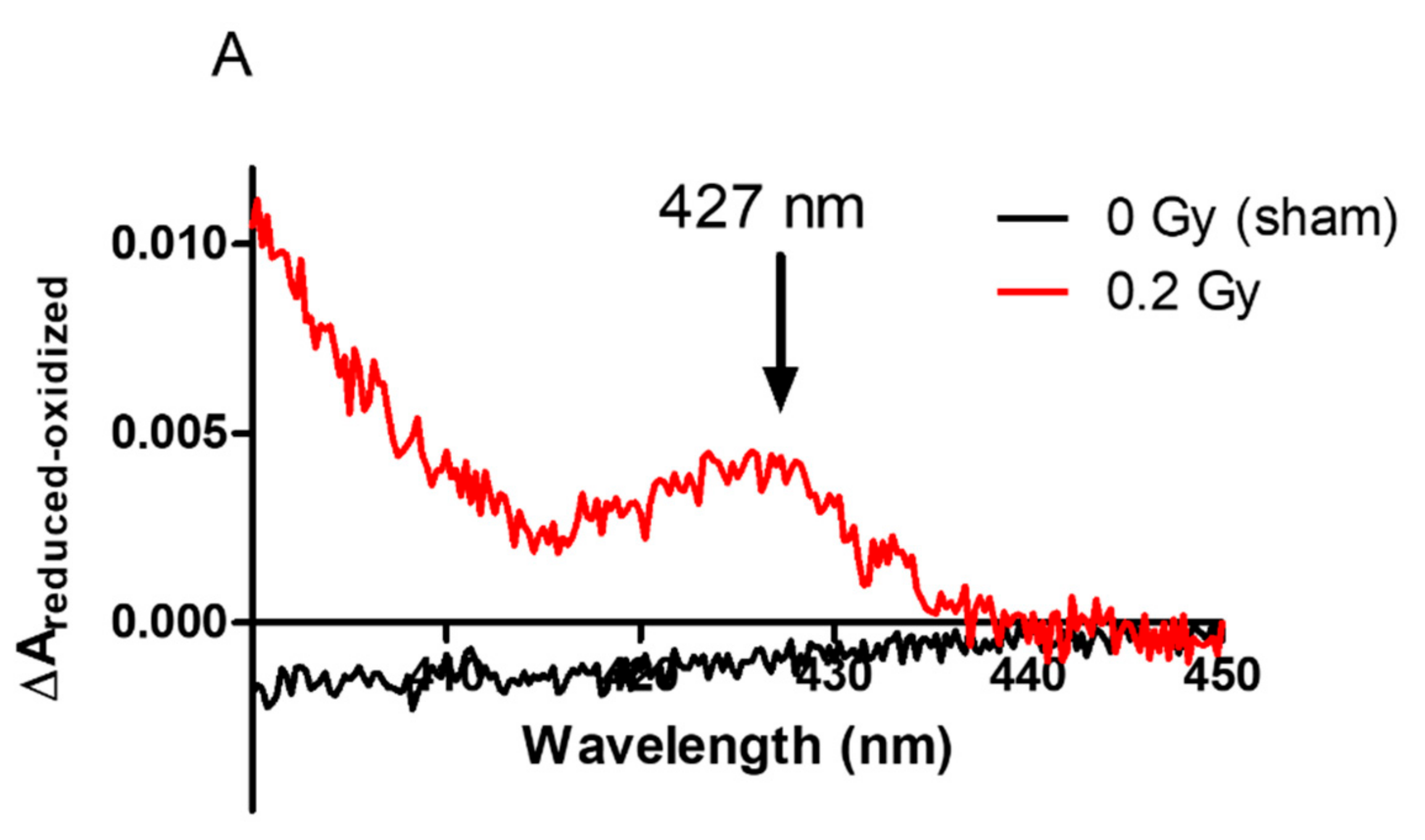
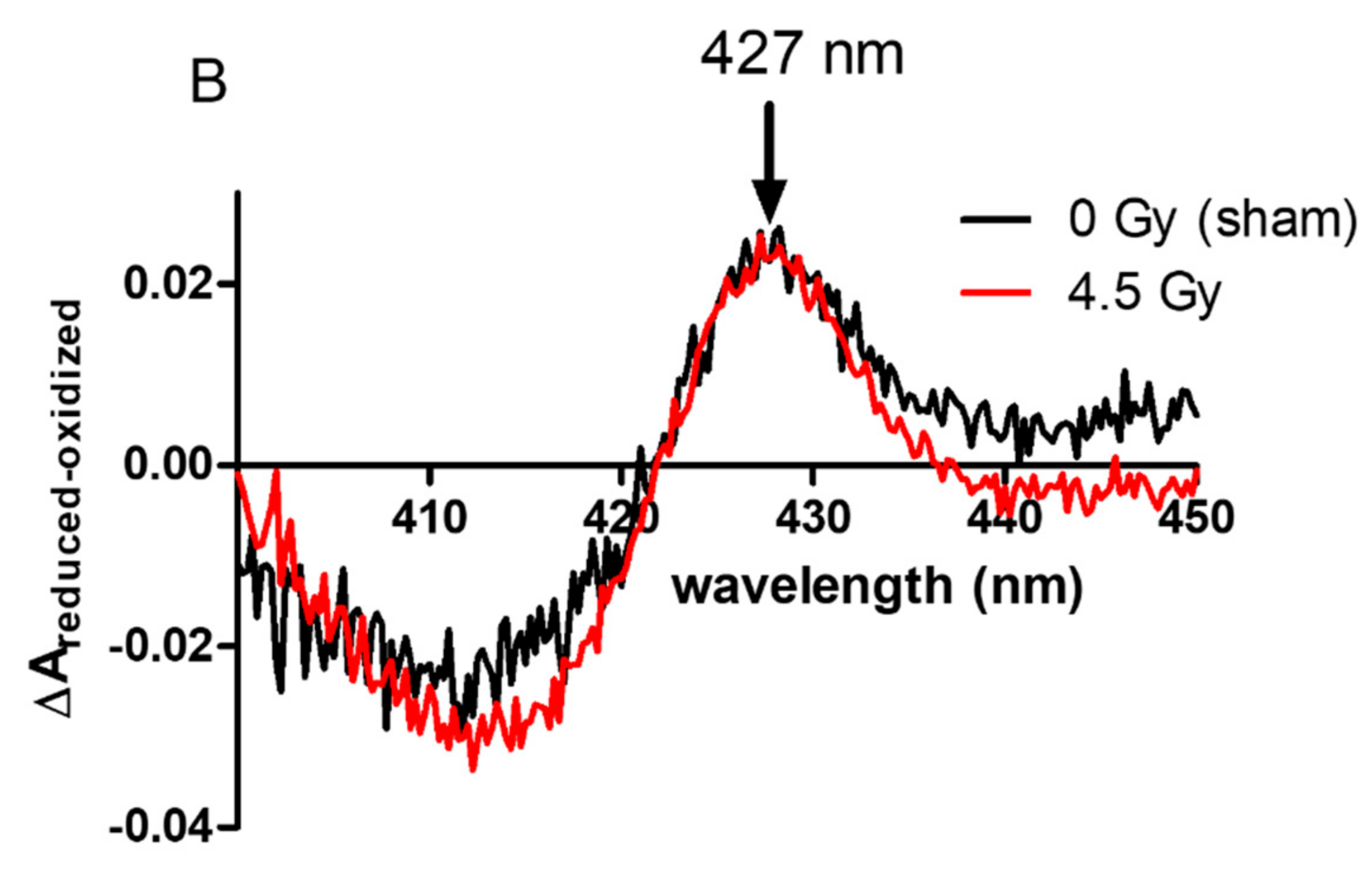
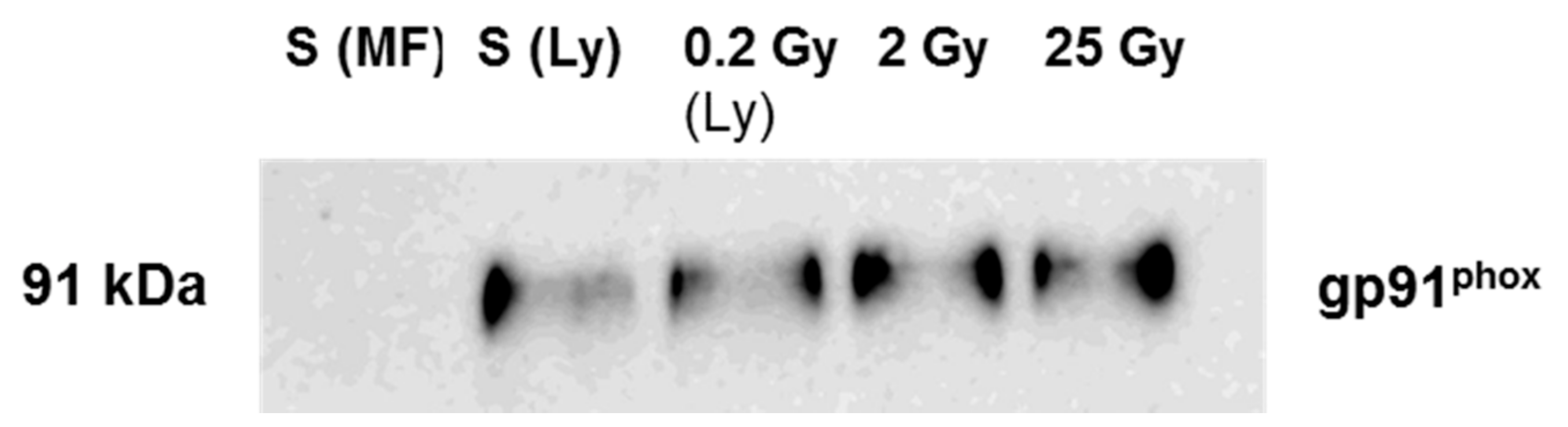
2.3. Localization of NADPH Oxidase in Irradiated PLB-985 Cells
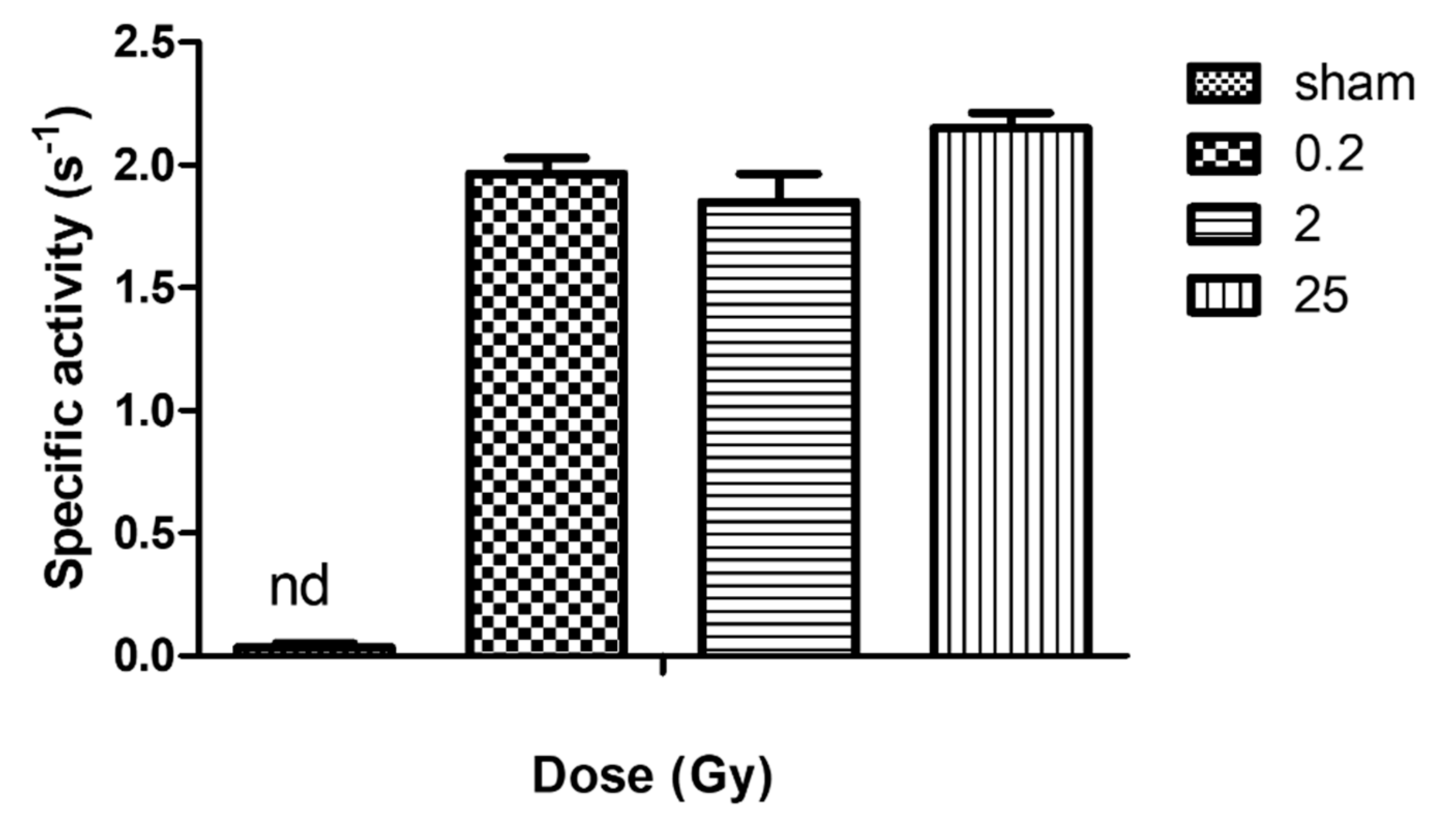
2.4. Activity of NADPH Oxidase in Irradiated PLB-985 Cells
3. Discussion and Conclusions
4. Materials and Methods
4.1. PLB Cell Culture and Irradiation
4.2. Zymosan Preparation
4.3. Phagocytosis Assay
4.4. In Cellullo ROS Detections
4.5. Purification of Plasma Membrane from PLB-985 Cells
4.6. The In Vitro NADPH Oxidase Activity
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ponti, G.; Maccaferri, M.; Ruini, C.; Tomasi, A.; Ozben, T. Biomarkers associated with COVID-19 disease progression. Crit. Rev. Clin. Lab. Sci. 2020, 57, 389–399. [Google Scholar] [CrossRef]
- Pan, F.; Yang, L.; Li, Y.C.; Liang, B.; Li, L.; Ye, T.H.; Li, L.L.; Liu, D.H.; Gui, S.; Hu, Y.; et al. Factors associated with death outcome in patients with severe coronavirus disease-19 (COVID-19): A case-control study. Int. J. Med. Sci. 2020, 17, 1281–1292. [Google Scholar] [CrossRef]
- Button, L.N.; Dewolf, W.C.; Newburger, P.E.; Jacobson, M.S.; Kevy, S.V. The Effects of Irradiation on Blood Components. Transfusion 1981, 21, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Buescher, E.S.; Gallin, J.T. Quality-Control of White Blood-Cell Transfusions—Radiation Effects on Human-Leukocytes. Clin. Res. 1982, 30, A312. [Google Scholar]
- Valerius, N.H.; Johansen, K.S.; Nielsen, O.S.; Platz, P.; Rosenkvist, J.; Sorensen, H. Effect of Invitro X-Irradiation on Lymphocyte and Granulocyte Function. Scand. J. Haematol. 1981, 27, 9–18. [Google Scholar] [CrossRef]
- Schwarzenberg, L.; Mathe, G.; Amiel, J.L.; Cattan, A.; Schneider, M.; Schlumberger, J.R. Study of Factors Determining Usefulness and Complications of Leukocyte Transfusions. Am. J. Med. 1967, 43, 206–213. [Google Scholar] [CrossRef]
- Holley, T.R.; Van Epps, D.E.; Harvey, R.L.; Anderson, R.E.; Williams, R.C., Jr. Effect of high doses of radiation on human neutrophil chemotaxis, phagocytosis and morphology. Am. J. Pathol. 1974, 75, 61–72. [Google Scholar] [PubMed]
- Owusu, S.B.; Hudik, E.; Ferard, C.; Dupre-Crochet, S.; Addison, E.C.D.K.; Preko, K.; Bizouarn, T.; Houee-Levin, C.; Baciou, L. Radiation-induced reactive oxygen species partially assemble neutrophil NADPH oxidase. Free. Radic. Biol. Med. 2021, 164, 76–84. [Google Scholar] [CrossRef]
- Ostuni, M.A.; Bizouarn, T.; Baciou, L.; Houee-Levin, C. Modulation of the Activity of the Nadph Oxidase System by Reactive Oxygen Species: Influence of Catalase. Radiat. Prot. Dosim. 2011, 143, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Ostuni, M.A.; Gelinotte, M.; Bizouarn, T.; Baciou, L.; Houee-Levin, C. Targeting NADPH-oxidase by reactive oxygen species reveals an initial sensitive step in the assembly process. Free Radic. Biol. Med. 2010, 49, 900–907. [Google Scholar] [CrossRef]
- Maduro, J.H.; Pras, E.; Willemse, P.H.B.; de Vries, E.G.E. Acute and long-term toxicity following radiotherapy alone or in combination with chemotherapy for locally advanced cervical cancer. Cancer Treat. Rev. 2003, 29, 471–488. [Google Scholar] [CrossRef]
- Collen, E.B.; Mayer, M.N. Acute effects of radiation treatment: Skin reactions. Can. Vet. J. Rev. Vet. Can. 2006, 47, 931. [Google Scholar]
- Sonveaux, P. ROS and radiotherapy: More we care. Oncotarget 2017, 8, 35482–35483. [Google Scholar] [CrossRef] [PubMed]
- Romero-Weaver, A.L.; Wan, X.S.; Diffenderfer, E.S.; Lin, L.; Kennedy, A.R. Effect of SPE-like Proton or Photon Radiation on the Kinetics of Mouse Peripheral Blood Cells and Radiation Biological Effectiveness Determinations. Astrobiology 2013, 13, 570–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, K.A.; Lilly, M.B.; Heck, L.; Rado, T.A. Characterization of a New Human-Diploid Myeloid-Leukemia Cell-Line (Plb-985) with Granulocytic and Monocytic Differentiating Capacity. Blood 1987, 70, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Volk, A.P.D.; Barber, B.M.; Goss, K.L.; Ruff, J.G.; Heise, C.K.; Hook, J.S.; Moreland, J.G. Priming of Neutrophils and Differentiated PLB-985 Cells by Pathophysiological Concentrations of TNF-alpha Is Partially Oxygen Dependent. J. Innate Immun. 2011, 3, 298–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedruzzi, E.; Fay, M.; Elbim, C.; Gaudry, M.; Gougerot-Pocidalo, M.A. Differentiation of PLB-985 myeloid cells into mature neutrophils, shown by degranulation of terminally differentiated compartments in response to N-formyl peptide and priming of superoxide anion production by granulocyte-macrophage colony-stimulating factor. Br. J. Haematol. 2002, 117, 719–726. [Google Scholar] [CrossRef] [PubMed]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic Transfer of Proteins from Polyacrylamide Gels to Nitrocellulose Sheets - Procedure and Some Applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaumel, S.; Stasia, M.J. The X-CGD PLB-985 Cell Model for NOX2 Structure-Function Analysis. Nadph Oxidases Methods Protoc. 2019, 1982, 153–171. [Google Scholar] [CrossRef]
- Yamauchi, A.; Yu, L.X.; Potgens, A.J.G.; Kuribayashi, F.; Nunoi, H.; Kanegasaki, S.; Roos, D.; Malech, H.L.; Dinauer, M.C.; Nakamura, M. Location of the epitope for 7D5, a monoclonal antibody raised against human flavocytochrome b(558), to the extracellular peptide portion of primate gp91(phox). Microbiol. Immunol. 2001, 45, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Pivot-Pajot, C.; Chouinard, F.C.; El Azreq, M.A.; Harbour, D.; Bourgoin, S.G. Characterisation of degranulation and phagocytic capacity of a human neutrophitic cellular model, PLB-985 cells. Immunobiology 2010, 215, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.J.; Lee, S.S.; Lee, S.T.; Lin, W.W. Signaling mechanisms of enhanced neutrophil phagocytosis and chemotaxis by the polysaccharide purified from Ganoderma lucidum. Br. J. Pharmacol. 2003, 139, 289–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Gou, S.J.; Huang, J.; Hao, J.; Chen, M.; Zhao, M.H. C5a and its receptors in human anti-neutrophil cytoplasmic antibody (ANCA)-associated vasculitis. Arthritis. Res. Ther. 2012, 14, R140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, K.L.; Summers, C.; Chilvers, E.R.; Condliffe, A.M. Priming and de-priming of neutrophil responses in vitro and in vivo. Eur. J. Clin. Investig. 2018, 48 (Suppl. 2), e12967. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dose (Gy) | Phagocytic Activity |
|---|---|
| Sham | 2011 ± 395 |
| 0.2 | 2123 ± 128 |
| 2 | 3082 ± 452 |
| 25 | 2467± 334 |
| Dose (Gy) | Total Membrane Proteins (mg/mL) | [cytb558] (µg/mL) | Total Number of Cells Broken (×106) | mg cytb558/mg Total Membrane Proteins/Cells (×10−11) |
|---|---|---|---|---|
| 0 | 4.01 ± 0.04 | nd | 17.5 ± 0.1 | nd |
| 0.2 | 4.90 ± 0.01 | 8.0 ± 1.0 | 17.6 ± 0.1 | 9.27 ± 0.79 |
| 2 | 5.09 ± 0.13 | 7.0 ± 0.0 | 13.9 ± 0.6 | 9.89 ± 0.82 |
| 25 | 5.2 ± 0.12 | 5.0 ± 0.0 | 11.3 ± 0.5 | 8.51 ± 0.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Owusu, S.B.; Dupré-Crochet, S.; Bizouarn, T.; Houée-Levin, C.; Baciou, L. Accumulation of Cytochrome b558 at the Plasma Membrane: Hallmark of Oxidative Stress in Phagocytic Cells. Int. J. Mol. Sci. 2022, 23, 767. https://doi.org/10.3390/ijms23020767
Owusu SB, Dupré-Crochet S, Bizouarn T, Houée-Levin C, Baciou L. Accumulation of Cytochrome b558 at the Plasma Membrane: Hallmark of Oxidative Stress in Phagocytic Cells. International Journal of Molecular Sciences. 2022; 23(2):767. https://doi.org/10.3390/ijms23020767
Chicago/Turabian StyleOwusu, Stephenson B., Sophie Dupré-Crochet, Tania Bizouarn, Chantal Houée-Levin, and Laura Baciou. 2022. "Accumulation of Cytochrome b558 at the Plasma Membrane: Hallmark of Oxidative Stress in Phagocytic Cells" International Journal of Molecular Sciences 23, no. 2: 767. https://doi.org/10.3390/ijms23020767
APA StyleOwusu, S. B., Dupré-Crochet, S., Bizouarn, T., Houée-Levin, C., & Baciou, L. (2022). Accumulation of Cytochrome b558 at the Plasma Membrane: Hallmark of Oxidative Stress in Phagocytic Cells. International Journal of Molecular Sciences, 23(2), 767. https://doi.org/10.3390/ijms23020767