

Chi-Circ_0006511 Positively Regulates the Differentiation of Goat Intramuscular Adipocytes via Novel-miR-87/CD36 Axis
 ,
,
Abstract
:1. Introduction
2. Results
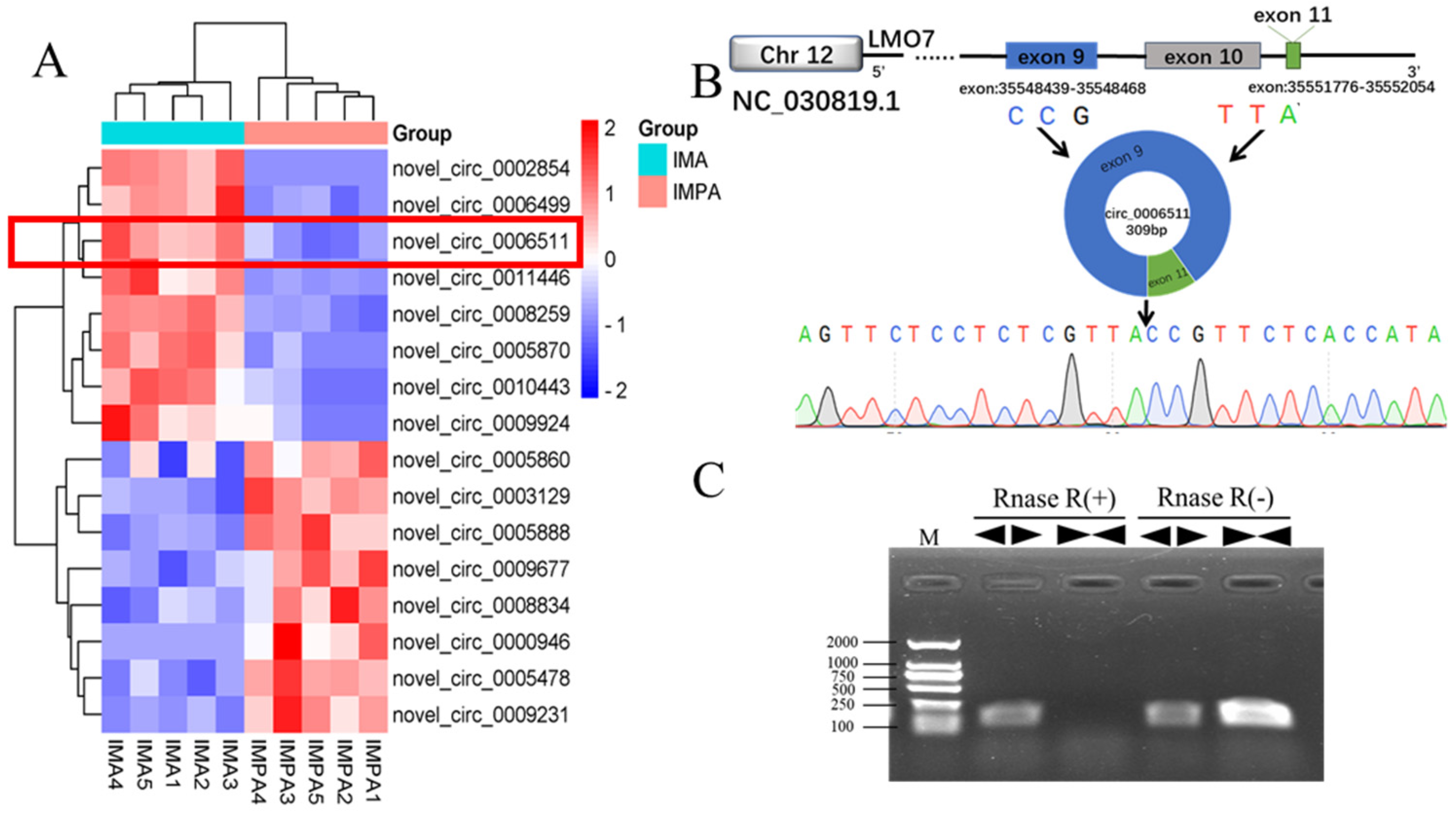
2.1. Chi-Circ_0006511 Identification
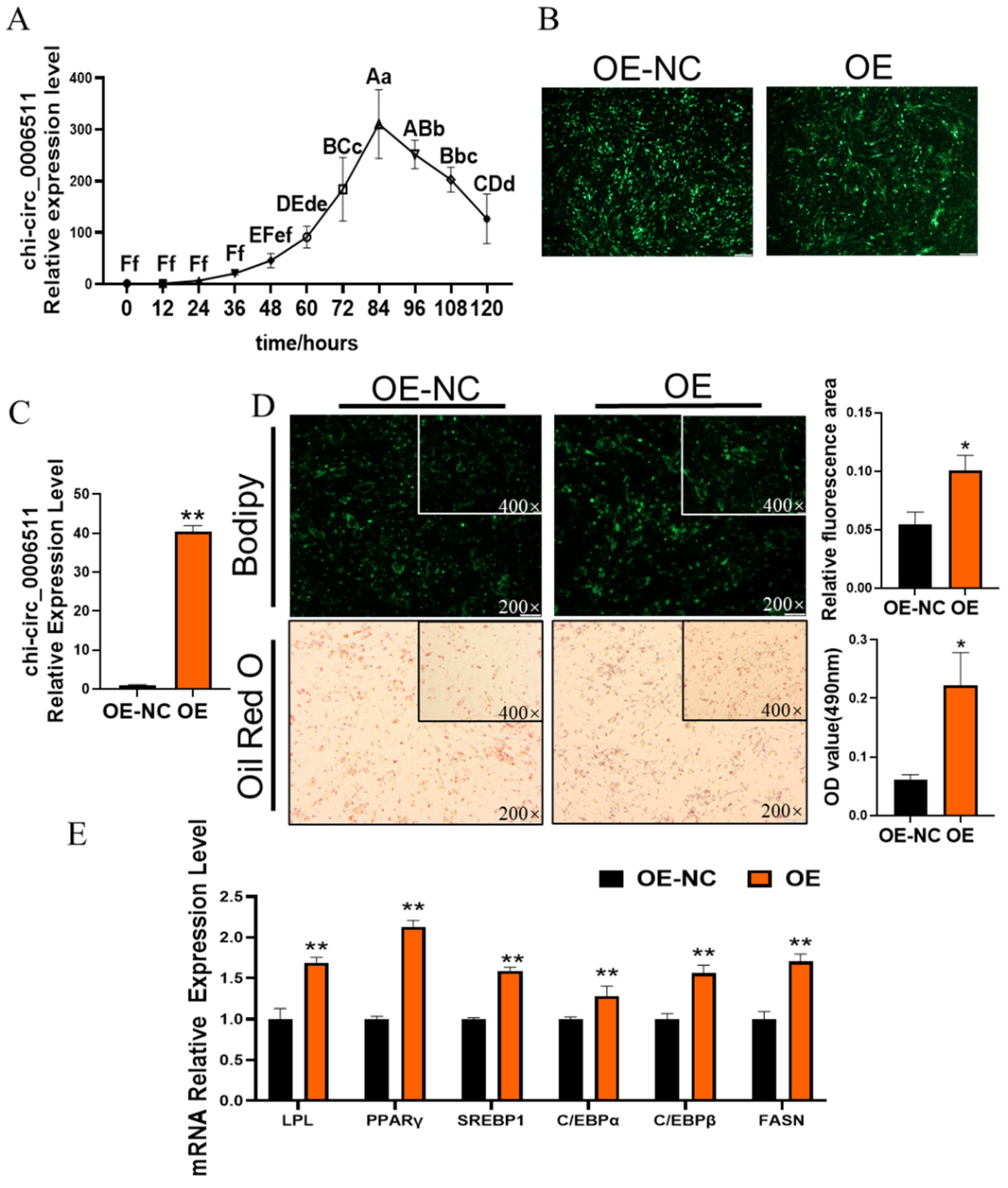
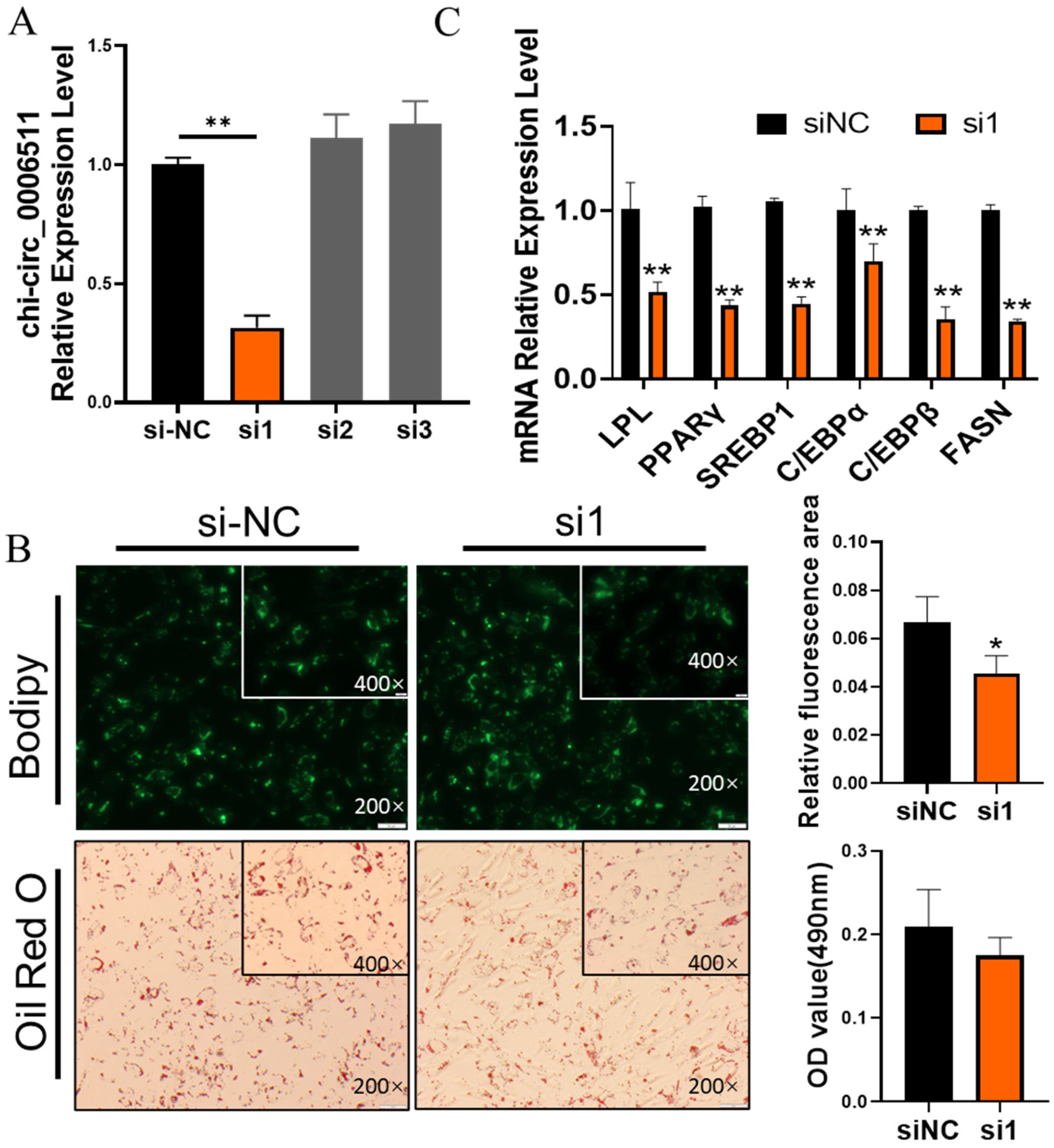
2.2. Chi-Circ_0006511 Promotes Gimpa Differentiation
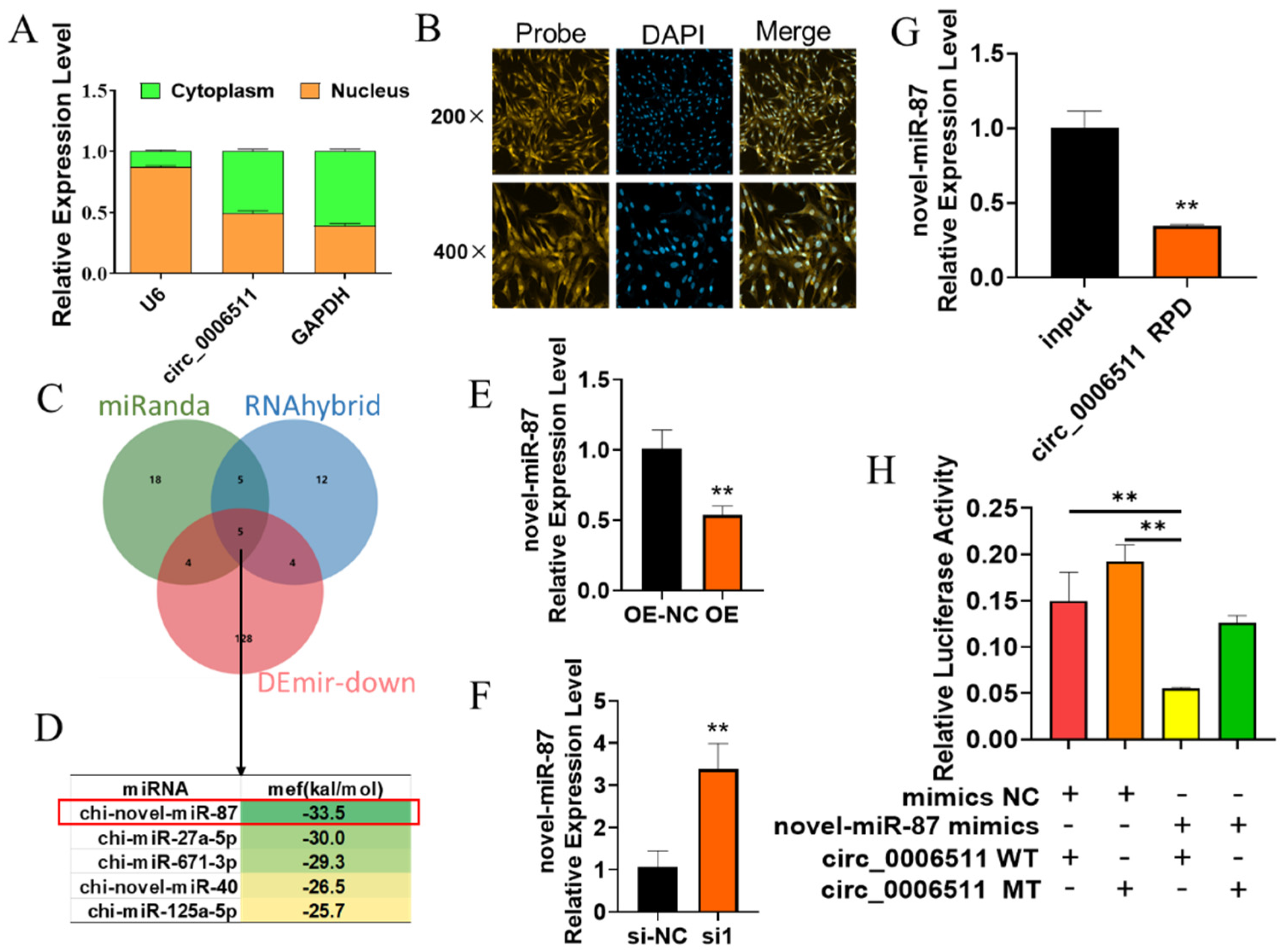
2.3. Chi-Circ_0006511 Sponge Novel-miR-87 as a ceRNA
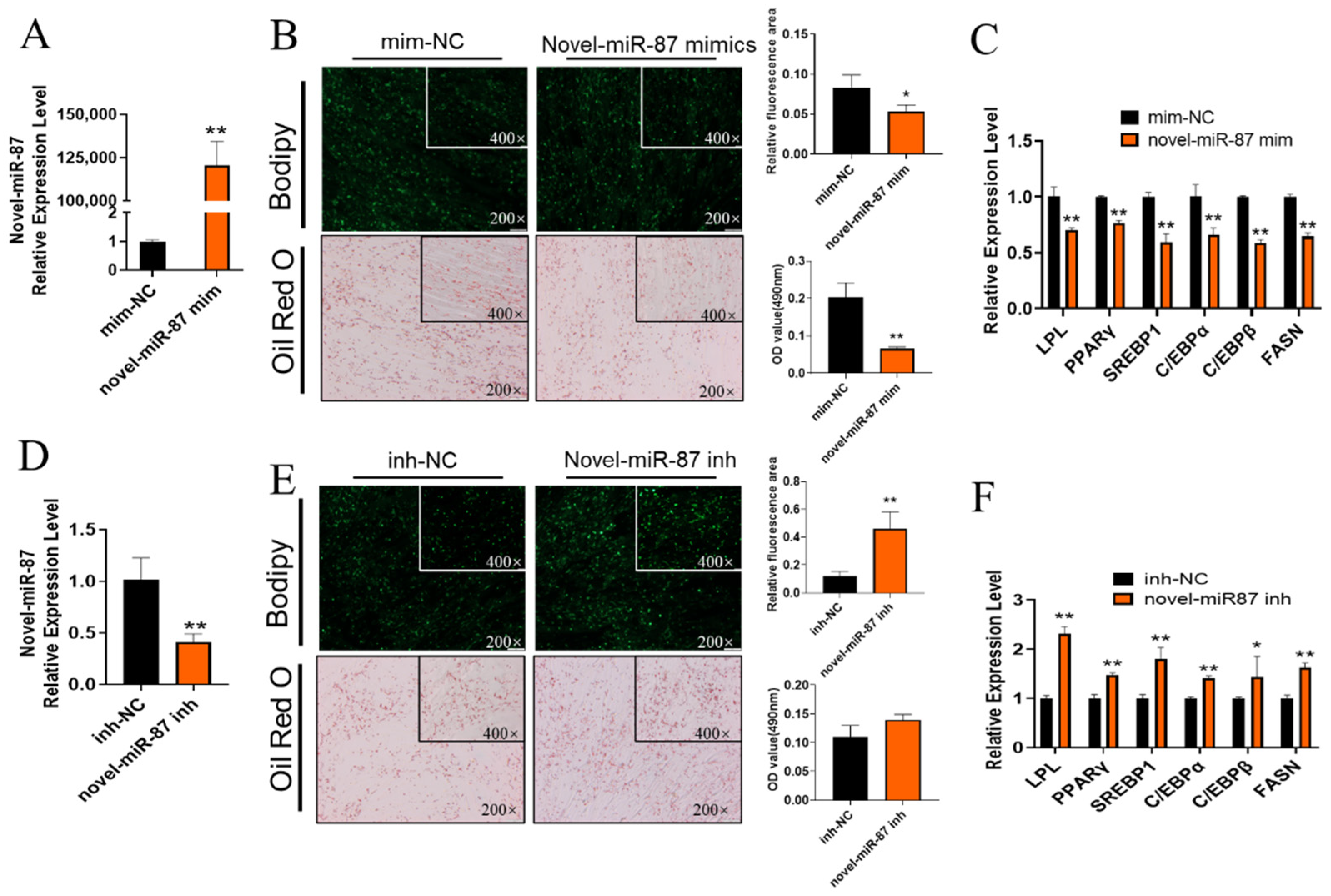
2.4. Novel-miR-87 Inhibits GIMPA Differentiation
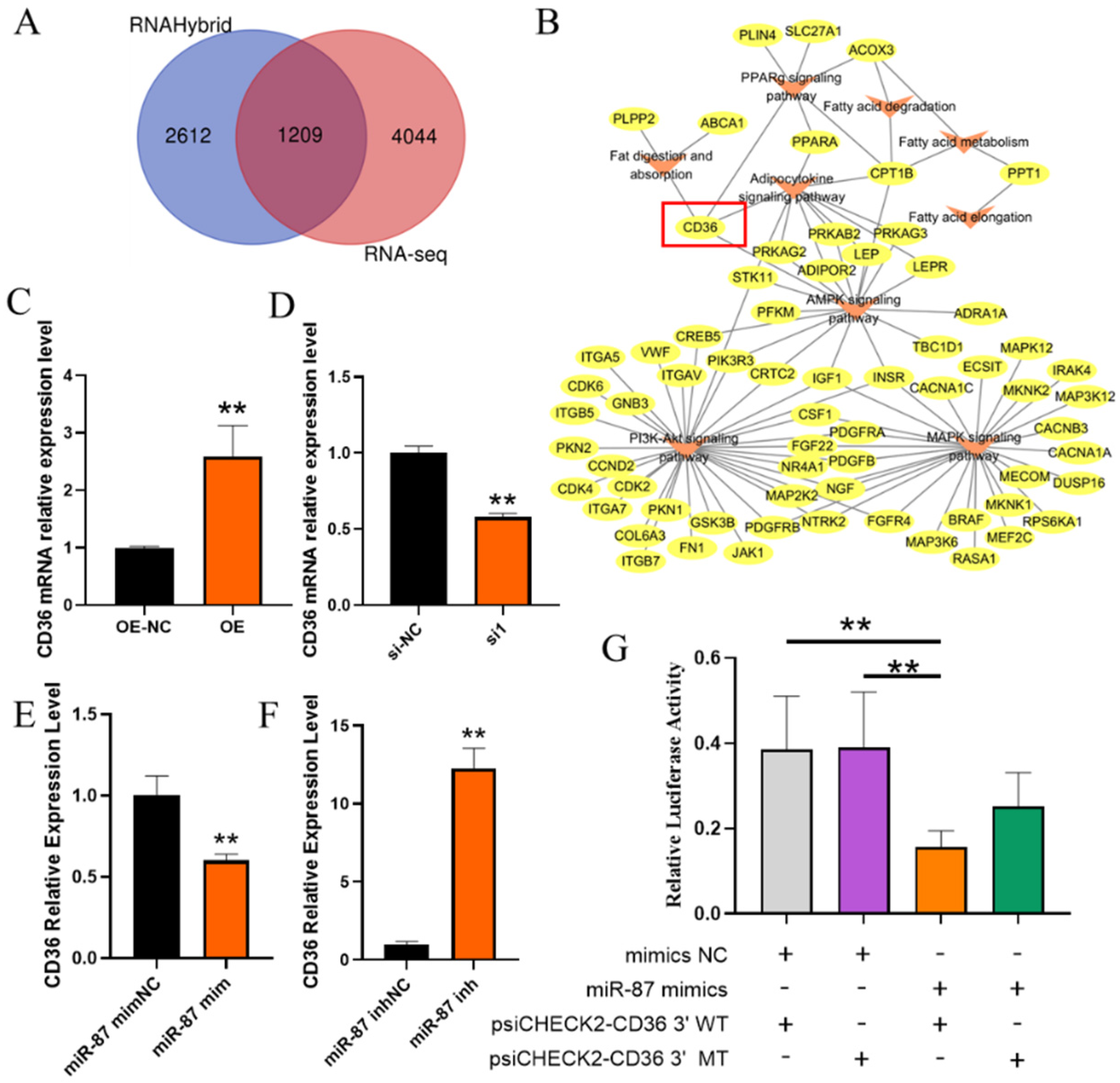
2.5. Identification of Target Genes of Novel-miR-87
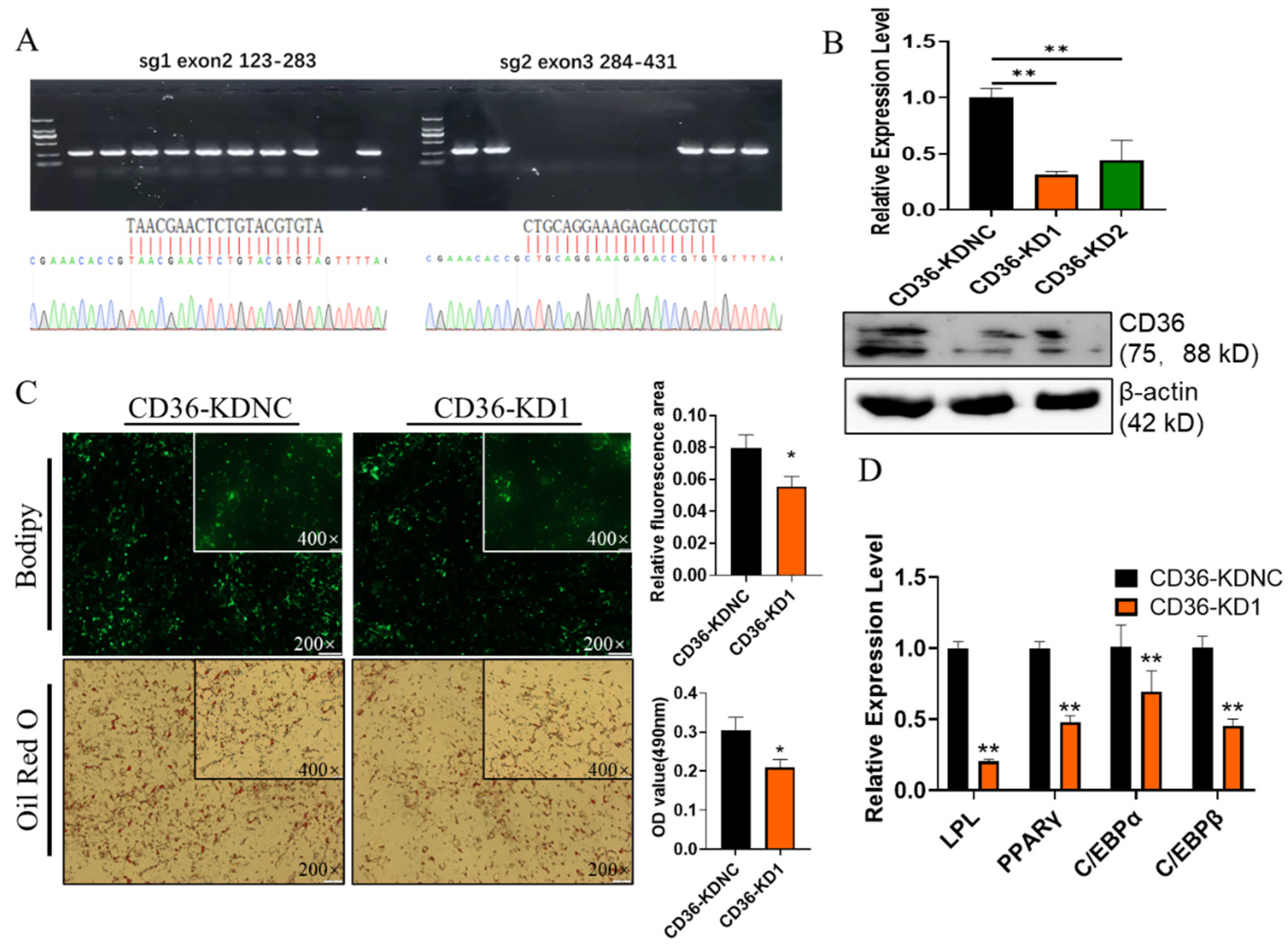
2.6. Knockdown of CD36 Inhibits GIMPA Differentiation
3. Discussion
4. Materials and Methods
4.1. Cell Isolation, Culture and Induction of Differentiation
4.2. Total RNA Extraction, Rnase R Treatment and cDNA Synthesis
4.3. Real-Time Quantitative PCR
4.4. Fluorescence In Situ Hybridization (FISH)
4.5. Nuclear and Cytoplasmic Isolation
4.6. Vector Construction
4.7. Small RNA Synthesis and Cell Transfection
4.8. Dual-Luciferase Reporter Assay
4.9. Bodipy Staining and Oil Red O Staining
4.10. Western Blot
4.11. RNA Pull Down
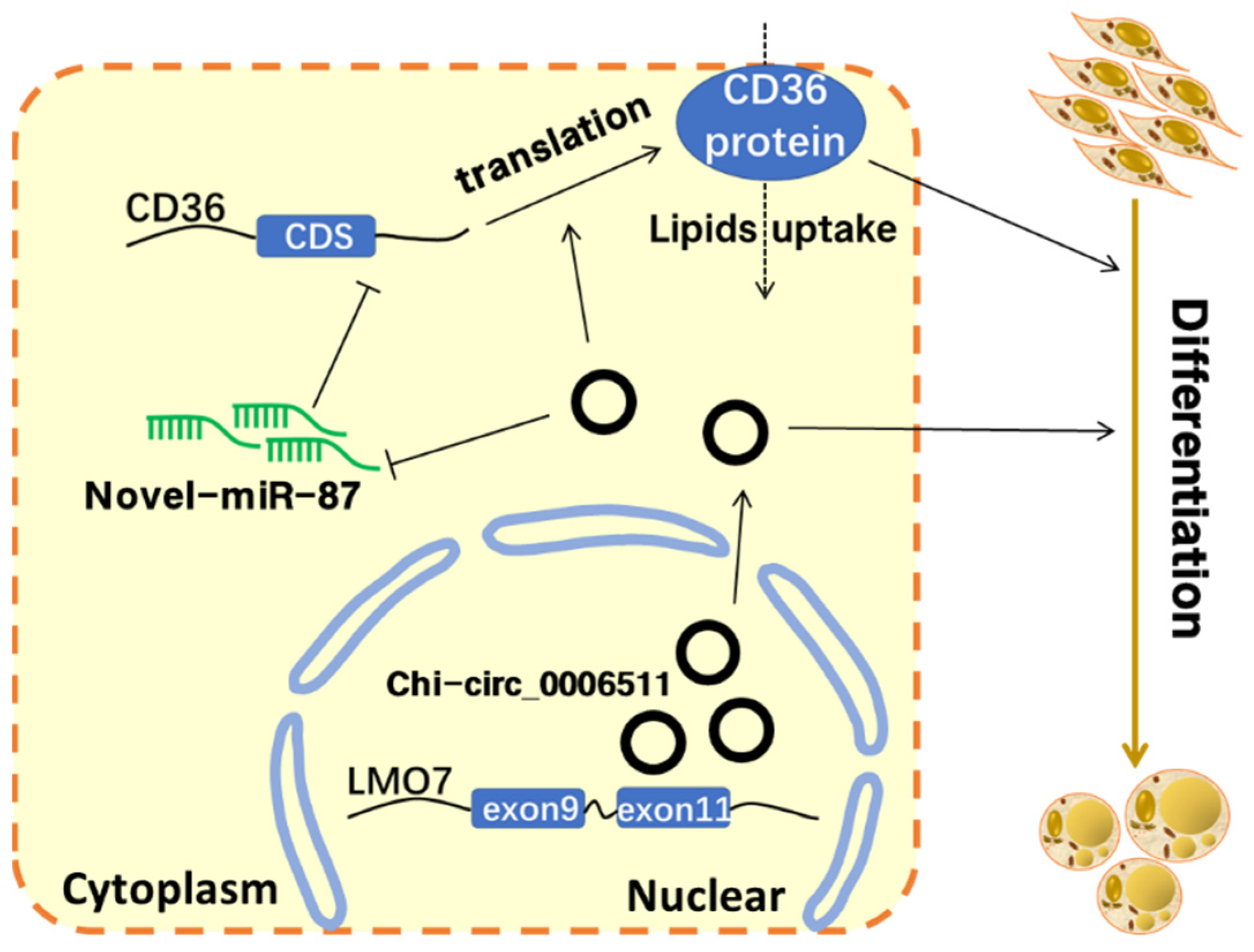
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Tefiel, H.; Ata, N.; Chahbar, M.; Benyarou, M.; Fantazi, K.; Yilmaz, O.; Cemal, I.; Karaca, O.; Boudouma, D.; Gaouar, S.B.S. Genetic characterization of four Algerian goat breeds assessed by microsatellite markers. Small Rumin. Res. 2018, 160, 65–71. [Google Scholar] [CrossRef]
- Stanišić, N.; Žujović, M.; Tomić, Z.; Maksimović, N.; Bijelić, Z.; Ivanović, S.; Memiši, N. The Effects of Crossing Balkan and Saanen Goat Breeds on Carcass Traits and Certain Quality Parameters of Kid Meat. Ann. Anim. Sci. 2012, 12, 53–62. [Google Scholar] [CrossRef]
- Ivanović, S.; Pavlović, M.; Pavlović, I.; Tasić, A.; Janjić, J.; Baltić, M.Ž. Influence of breed on selected quality parameters of fresh goat meat. Arch. Anim. Breed. 2020, 63, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Madruga, M.S.; Torres, T.S.; Carvalho, F.F.; Queiroga, R.C.; Narain, N.; Garrutti, D.; Neto, M.A.S.; Mattos, C.W.; Costa, R.G. Meat quality of Moxotó and Canindé goats as affected by two levels of feeding. Meat Sci. 2008, 80, 1019–1023. [Google Scholar] [CrossRef] [PubMed]
- Silva Mariniello, T.; Oliveira Lopes, R.; Barbosa Pires, L.; Neto, G.; Froes, A.; Bagaldo, A.R.; Lanna, D.P.D.; Alves de Silva, M.C.; Brito de Jesus, I. Preliminary Study on Meat Quality of Goats Fed Levels of Licury Oil in the Diet. Asian-Australas. J. Anim. Sci. 2011, 24, 1112–1119. [Google Scholar] [CrossRef]
- Thomas, E.M.; Roden, J.A.; Haresign, W.; Richardson, R.I.; Lambe, N.R.; Clelland, N.; Gardner, G.E.; Scollan, N.D. Meat eating and nutritional quality of lambs sired by high and low muscle density rams. Animal 2021, 15, 100136. [Google Scholar] [CrossRef]
- Zhang, H.; Guan, W. The response of gene expression associated with intramuscular fat deposition in the longissimus dorsi muscle of Simmental x Yellow breed cattle to different energy levels of diets. Anim. Sci. J. 2019, 90, 493–503. [Google Scholar] [CrossRef]
- Haczeyni, F.; Bell-Anderson, K.S.; Farrell, G.C. Causes and mechanisms of adipocyte enlargement and adipose expansion. Obes. Rev. 2018, 19, 406–420. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, X.; Deng, Y.; Cui, Q.; Zhu, J.; Ren, H.; Liu, Y.; Hu, X.; Zuo, J.; Peng, Y. Regulatory roles of circRNAs in adipogenesis and lipid metabolism: Emerging insights into lipid-related diseases. FEBS J. 2021, 288, 3663–3682. [Google Scholar] [CrossRef] [PubMed]
- de Sa, P.M.; Richard, A.J.; Hang, H.; Stephens, J.M. Transcriptional Regulation of Adipogenesis. Compr. Physiol. 2017, 7, 635–674. [Google Scholar] [CrossRef]
- Xu, S.; Chen, P.; Sun, L. Regulatory networks of non-coding RNAs in brown/beige adipogenesis. Biosci. Rep. 2015, 35, e00262. [Google Scholar] [CrossRef]
- Zhang, J.; Luo, Y.; Wang, Y.; He, L.; Li, M.; Wang, X. MicroRNA regulates animal adipocyte differentiation. Yichuan 2015, 37, 1175–1184. [Google Scholar] [CrossRef]
- Lefterova, M.I.; Zhang, Y.; Steger, D.J.; Schupp, M.; Schug, J.; Cristancho, A.; Feng, D.; Zhuo, D.; Stoeckert, C.J., Jr.; Liu, X.S.; et al. PPAR gamma and C/EBP factors orchestrate adipocyte biology via adjacent binding on a genome-wide scale. Genes Dev. 2008, 22, 2941–2952. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; Hsu, C.H.; Wang, X.Z.; Sakai, S.; Freeman, M.W.; Gonzalez, F.J.; Spiegelman, B.M. C/EBP alpha induces adipogenesis through PPAR gamma: A unified pathway. Genes Dev. 2002, 16, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Cirilli, M.; Bereshchenko, O.; Ermakova, O.; Nerlov, C. Insights into specificity, redundancy and new cellular functions of C/EBPa and C/EBPb transcription factors through interactome network analysis. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 467–476. [Google Scholar] [CrossRef]
- Tanaka, T.; Yoshida, N.; Kishimoto, T.; Akira, S. Defective adipocyte differentiation in mice lacking the C/EBPbeta and/or C/EBPdelta gene. EMBO Eur. Mol. Biol. Organ. J. 1997, 16, 7432–7443. [Google Scholar] [CrossRef] [Green Version]
- Hamm, J.K.; Park, B.H.; Farmer, S.R. A role for C/EBP beta in regulating peroxisome proliferator-activated receptor gamma activity during adipogenesis in 3T3-L1 preadipocytes. J. Biol. Chem. 2001, 276, 18464–18471. [Google Scholar] [CrossRef] [Green Version]
- Salma, N.; Xiao, H.G.; Mueller, E.; Imbalzano, A.N. Temporal recruitment of transcription factors and SWI/SNF chromatin-remodeling enzymes during adipogenic induction of the peroxisome proliferator-activated receptor gamma nuclear hormone receptor. Mol. Cell. Biol. 2004, 24, 4651–4663. [Google Scholar] [CrossRef] [Green Version]
- Salma, N.; Xiao, H.; Imbalzano, A.N. Temporal recruitment of CCAAT/enhancer-binding proteins to early and late adipogenic promoters in vivo. J. Mol. Endocrinol. 2006, 36, 139–151. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, S.G.; Kang, Y.J.; Kwon, T.K.; Nam, J.-O. Kahweol inhibits adipogenesis of 3T3-L1 adipocytes through downregulation of PPAR. Nat. Prod. Res. 2018, 32, 1216–1219. [Google Scholar] [CrossRef]
- Engin, A.B. MicroRNA and Adipogenesis. In Obesity and Lipotoxicity; Engin, A.B., Engin, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 489–509. [Google Scholar]
- Wang, Z.; Luo, Z.; Dai, Z.; Zhong, Y.; Liu, X.; Zuo, C. Long non-coding RNA lnc-OAD is required for adipocyte differentiation in 3T3-L1 preadipocytes. Biochem. Biophys. Res. Commun. 2019, 511, 753–758. [Google Scholar] [CrossRef]
- Zhu, Y.; Gui, W.; Lin, X.; Li, H. Knock-down of circular RNA H19 induces human adipose-derived stem cells adipogenic differentiation via a mechanism involving the polypyrimidine tract-binding protein 1. Exp. Cell Res. 2020, 387, 111753. [Google Scholar] [CrossRef]
- Liu, H.; Li, B.; Qiao, L.; Liu, J.; Ren, D.; Liu, W. miR-340-5p inhibits sheep adipocyte differentiation by targeting ATF7. Anim. Sci. J. 2020, 91, e13462. [Google Scholar] [CrossRef]
- Du, Y.; Zhao, Y.; Wang, Y.; Meng, Q.; Zhu, J.; Lin, Y. MiR-25-3p regulates the differentiation of intramuscular preadipocytes in goat via targeting KLF4. Arch. Anim. Breed. 2021, 64, 17–25. [Google Scholar] [CrossRef]
- He, C.; Wang, Y.; Zhu, J.; Li, Y.; Chen, J.; Lin, Y. Integrative Analysis of lncRNA-miRNA-mRNA Regulatory Network Reveals the Key lncRNAs Implicated Potentially in the Differentiation of Adipocyte in Goats. Front. Physiol. 2022, 13, 900179. [Google Scholar] [CrossRef]
- Kang, Z.; Zhang, S.; Jiang, E.; Wang, X.; Wang, Z.; Chen, H.; Lan, X. circFLT1 and lncCCPG1 Sponges miR-93 to Regulate the Proliferation and Differentiation of Adipocytes by Promoting lncSLC30A9 Expression. Mol. Ther. Nucleic Acids 2020, 22, 484–499. [Google Scholar] [CrossRef]
- Liu, X.; Bai, Y.; Cui, R.; He, S.; Zhao, X.; Wu, K.; Fang, M. Sus_circPAPPA2 Regulates Fat Deposition in Castrated Pigs through the miR-2366/GK Pathway. Biomolecules 2022, 12, 753. [Google Scholar] [CrossRef]
- Laidi, W.; Wenshuang, L.; Shasha, W.; Zhixiu, W.; Hao, B.; Yong, J.; Yulin, B.; Guohong, C.; Guobin, C. Circular RNA expression profiling reveals that circ-PLXNA1 functions in duck adipocyte differentiation. PLoS ONE 2020, 15, e0236069. [Google Scholar]
- Dong, R.; Ma, X.-K.; Chen, L.-L.; Yang, L. Increased complexity of circRNA expression during species evolution. RNA Biol. 2017, 14, 1064–1074. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhong, Y.; Wang, X.; Shen, J.; An, W. Advances in Circular RNA and Its Applications. Int. J. Med. Sci. 2022, 19, 975–985. [Google Scholar] [CrossRef]
- Pamudurti, N.R.; Bartok, O.; Jens, M.; Ashwal-Fluss, R.; Stottmeister, C.; Ruhe, L.; Hanan, M.; Wyler, E.; Perez-Hernandez, D.; Ramberger, E.; et al. Translation of CircRNAs. Mol. Cell 2017, 66, 9–21.e27. [Google Scholar] [CrossRef]
- Chekulaeva, M.; Rajewsky, N. Roles of Long Noncoding RNAs and Circular RNAs in Translation. Cold Spring Harb. Perspect. Biol. 2019, 11, a032680. [Google Scholar] [CrossRef]
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell. 2013, 51, 792–806. [Google Scholar] [CrossRef] [Green Version]
- Zganiacz, D.; Milanowski, R. Characteristics of circular ribonucleic acid molecules (circRNA). Postep. Biochem. 2017, 63, 221–232. [Google Scholar]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Qiao, L.; Li, C.G.; Liu, D. CircRNA_0048211 protects postmenopausal osteoporosis through targeting miRNA-93-5p to regulate BMP2. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 3459–3466. [Google Scholar]
- Wang, X.; Li, H.; Lu, Y.; Cheng, L. Regulatory Effects of Circular RNAs on Host Genes in Human Cancer. Front. Oncol. 2021, 10, 586163. [Google Scholar] [CrossRef]
- Chen, C.-K.; Cheng, R.; Demeter, J.; Chen, J.; Weingarten-Gabbay, S.; Jiang, L.; Snyder, M.P.; Weissman, J.S.; Segal, E.; Jackson, P.K.; et al. Structured elements drive extensive circular RNA translation. Mol. Cell 2021, 81, 4300–4318. [Google Scholar] [CrossRef]
- Zhao, H.; Zhou, Q.; Li, X. Protein Bait Hypothesis: CircRNA-Encoded Proteins Competitively Inhibit Cognate Functional Isoforms. Trends Genet. 2021, 37, 616–624. [Google Scholar] [CrossRef]
- Zhu, B.; Kang, Z.; Zhu, S.; Zhang, Y.; Lai, X.; Zhou, L.; Huang, H.; Gao, X.; Jiang, C.; Zeng, J. Multi-Omics Characterization of Circular RNA-Encoded Novel Proteins Associated with Bladder Outlet Obstruction. Front. Cell Dev. Biol. 2022, 9, 772534. [Google Scholar] [CrossRef]
- Ma, H.B.; Yao, Y.N.; Yu, J.J.; Chen, X.X.; Li, H.F. Extensive profiling of circular RNAs and the potential regulatory role of circRNA-000284 in cell proliferation and invasion of cervical cancer via sponging miR-506. Am. J. Transl. Res. 2018, 10, 592–604. [Google Scholar]
- Zhang, D.D.; Ni, N.; Wang, Y.Y.; Tang, Z.M.; Gao, H.Q.; Ju, Y.H.; Sun, N.; He, X.Y.; Gu, P.; Fan, X.N. CircRNA-vgll3 promotes osteogenic differentiation of adipose-derived mesenchymal stem cells via modulating miRNA-dependent integrin alpha 5 expression. Cell Death Differ. 2021, 28, 283–302. [Google Scholar] [CrossRef]
- Jiang, R.; Li, H.; Yang, J.M.; Shen, X.M.; Song, C.C.; Yang, Z.X.; Wang, X.G.; Huang, Y.Z.; Lan, X.Y.; Lei, C.Z.; et al. circRNA Profiling Reveals an Abundant circFUT10 that Promotes Adipocyte Proliferation and Inhibits Adipocyte Differentiation via Sponging let-7. Mol. Ther. Nucleic Acids 2020, 20, 491–501. [Google Scholar] [CrossRef]
- Wang, H.B.; Niu, X.B.; Mao, F.; Liu, X.Z.; Zhong, B.; Jiang, H.S.; Fu, G.B. Hsa_circRNA_100146 Acts as a Sponge of miR-149-5p in Promoting Bladder Cancer Progression via Regulating RNF2. Oncotargets Ther. 2020, 13, 11007–11017. [Google Scholar] [CrossRef]
- Wang, Y.H.; Li, M.L.; Wang, Y.H.; Liu, J.; Zhang, M.L.; Fang, X.T.; Chen, H.; Zhang, C.L. A Zfp609 circular RNA regulates myoblast differentiation by sponging miR-194-5p. Int. J. Biol. Macromol. 2019, 121, 1308–1313. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Zhu, L.; Bai, M.; Liu, Y.; Zhan, Y.; Deng, T.; Yang, H.O.; Sun, W.; Wang, X.Y.; Zhu, K.G.; et al. Exosomal circRNA derived from gastric tumor promotes white adipose browning by targeting the miR-133/PRDM16 pathway. Int. J. Cancer 2019, 144, 2501–2515. [Google Scholar] [CrossRef]
- Jiyao, W.; Shaoli, Z.; Binglin, Y.; Sihuan, Z.; Enhui, J.; Hong, C.; Xianyong, L. CircRNA Profiling Reveals CircPPARγ Modulates Adipogenic Differentiation via Sponging miR-92a-3p. J. Agric. Food Chem. 2022, 70, 6698–6708. [Google Scholar]
- Li, B.J.; He, Y.; Wu, W.J.; Tan, X.F.; Wang, Z.C.H.; Irwin, D.M.; Wang, Z.; Zhang, S.Y. Circular RNA Profiling Identifies Novel circPPARA that Promotes Intramuscular Fat Deposition in Pigs. J. Agric. Food Chem. 2022, 70, 4123–4137. [Google Scholar] [CrossRef]
- Li, X.; Yang, L.; Chen, L.-L. The Biogenesis, Functions, and Challenges of Circular RNAs. Mol. Cell 2018, 71, 428–442. [Google Scholar] [CrossRef] [Green Version]
- Gregory, R.I.; Yan, K.P.; Amuthan, G.; Chendrimada, T.; Doratotaj, B.; Cooch, N.; Shiekhattar, R. The Microprocessor complex mediates the genesis of microRNAs. Nature 2004, 432, 235–240. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Najafi, M.; Mohammadinejad, R.; Farkhondeh, T.; Samarghandian, S. Flaming the fight against cancer cells: The role of microRNA-93. Cancer Cell Int. 2020, 20, 227. [Google Scholar] [CrossRef]
- Abeyrathna, P.; Su, Y. The critical role of Akt in cardiovascular function. Vasc. Pharmacol. 2015, 74, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef] [Green Version]
- Shackelford, D.B.; Shaw, R.J. The LKB1-AMPK pathway: Metabolism and growth control in tumour suppression. Nat. Rev. Cancer 2009, 9, 563–575. [Google Scholar] [CrossRef]
- Cui, Y.; Hosui, A.; Sun, R.; Shen, K.; Gavrilova, O.; Chen, W.; Cam, M.C.; Gao, B.; Robinson, G.W.; Hennighausen, L. Loss of signal transducer and activator of transcription 5 leads to hepatosteatosis and impaired liver regeneration. Hepatology 2007, 46, 504–513. [Google Scholar] [CrossRef]
- Li, P.; Shan, K.; Liu, Y.; Zhang, Y.; Xu, L.; Xu, L. CircScd1 Promotes Fatty Liver Disease via the Janus Kinase 2/Signal Transducer and Activator of Transcription 5 Pathway. Dig. Dis. Sci. 2019, 64, 113–122. [Google Scholar] [CrossRef]
- Sos, B.C.; Harris, C.; Nordstrom, S.M.; Tran, J.L.; Balazs, M.; Caplazi, P.; Febbraio, M.; Applegate, M.A.B.; Wagner, K.-U.; Weiss, E.J. Abrogation of growth hormone secretion rescues fatty liver in mice with hepatocyte-specific deletion of JAK2. J. Clin. Investig. 2011, 121, 1412–1423. [Google Scholar] [CrossRef]
- Beigneux, A.P.; Davies, B.S.J.; Gin, P.; Weinstein, M.M.; Farber, E.; Qiao, X.; Peale, F.; Bunting, S.; Walzem, R.L.; Wong, J.S.; et al. Glycosylphosphatidylinositol-anchored high-density lipoprotein-binding protein 1 plays a critical role in the lipolytic processing of chylomicrons. Cell Metab. 2007, 5, 279–291. [Google Scholar] [CrossRef] [Green Version]
- Korn, E.D. Clearing factor, a heparin-activated lipoprotein lipase. I. Isolation and characterization of the enzyme from normal rat heart. J. Biol. Chem. 1955, 215, 1–14. [Google Scholar] [CrossRef]
- Weinstock, P.H.; Bisgaier, C.L.; Aalto-Setala, K.; Radner, H.; Ramakrishnan, R.; Levak-Frank, S.; Essenburg, A.D.; Zechner, R.; Breslow, J.L. Severe hypertriglyceridemia, reduced high density lipoprotein, and neonatal death in lipoprotein lipase knockout mice: Mild hypertriglyceridemia with impaired very low density lipoprotein clearance in heterozygotes. J. Clin. Investig. 1995, 96, 2555–2568. [Google Scholar] [CrossRef]
- Kersten, S. Physiological regulation of lipoprotein lipase. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2014, 1841, 919–933. [Google Scholar] [CrossRef] [PubMed]
- Stern, J.H.; Rutkowski, J.M.; Scherer, P.E. Adiponectin, Leptin, and Fatty Acids in the Maintenance of Metabolic Homeostasis through Adipose Tissue Crosstalk. Cell Metab. 2016, 23, 770–784. [Google Scholar] [CrossRef] [Green Version]
- Pepino, M.Y.; Kuda, O.; Samovski, D.; Abumrad, N.A. Structure-Function of CD36 and Importance of Fatty Acid Signal Transduction in Fat Metabolism. Annu. Rev. Nutr. 2014, 34, 281–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putri, M.; Syamsunarno, M.; Iso, T.; Yamaguchi, A.; Hanaoka, H.; Sunaga, H.; Koitabashi, N.; Matsui, H.; Yamazaki, C.; Kameo, S.; et al. CD36 is indispensable for thermogenesis under conditions of fasting and cold stress. Biochem. Biophys. Res. Commun. 2015, 457, 520–525. [Google Scholar] [CrossRef]
- Nahle, Z.; Hsieh, M.; Pietka, T.; Coburn, C.T.; Grimaldi, P.A.; Zhang, M.Q.; Das, D.; Abumrad, N.A. CD36-dependent regulation of muscle FoxO1 and PDK4 in the PPAR delta/beta-mediated adaptation to metabolic stress. J. Biol. Chem. 2008, 283, 14317–14326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samovski, D.; Su, X.; Xu, Y.; Abumrad, N.A.; Stahl, P.D. Insulin and AMPK regulate FA translocase/CD36 plasma membrane recruitment in cardiomyocytes via Rab GAP AS160 and Rab8a Rab GTPase. J. Lipid Res. 2012, 53, 709–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thampi, P.; Stewart, B.W.; Joseph, L.; Melnyk, S.B.; Henningse, L.J.; Nagarajana, S. Dietary homocysteine promotes atherosclerosis in apoE-deficient mice by inducing scavenger receptors expression. Atherosclerosis 2008, 197, 620–629. [Google Scholar] [CrossRef]
 ” indicates Divergent primer, “
” indicates Divergent primer, “ ” indicates Convergent primer.
” indicates Divergent primer, “” indicates Convergent primer.
” indicates Convergent primer.
” indicates Divergent primer, “” indicates Convergent primer.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence | Purpose | Products Length | TM/°C |
|---|---|---|---|---|
| chi_circ_0006511 Divergent | GGATCCCAGAGGATGTGAACTGGA | qPCR | 235 | 60 |
| TCATCCTTTACAGTGTTGGGAACGC | ||||
| chi_circ_0006511 Convergent | GGGCACGAACGTGGAGA | PCR | 147 | 60 |
| GGGCTTGGCATGACTGG | ||||
| Novel-miR-87 | ACTAATGGGCTTGGGGAGCCT | Novel-miR-87qPCR | 64 | |
| A-TY | ATCCAGTGCAGGGTCCGAGG | |||
| CD36 | AGAATCCAGACGAAGTGGCA | qPCR | 214 | 60 |
| ACAGCCAGGTTGAGAATGGT | ||||
| SREBP1 | AAGTGGTGGGCCTCTCTGA | qPCR | 127 | 58 |
| GCAGGGGTTTCTCGGACT | ||||
| FASN | TGTGCAACTGTGCCCTAG | qPCR | 111 | 57 |
| GTCCTCTGAGCAGCGTGT | ||||
| PPARγ | AAGCGTCAGGGTTCCACTATG | qPCR | 197 | 60 |
| GAACCTGATGGCGTTATGAGAC | ||||
| C/EBPα | CCGTGGACAAGAACAGCAAC | qPCR | 142 | 58 |
| AGGCGGTCATTGTCACTGGT | ||||
| C/EBPβ | CAAGAAGACGGTGGACAAGC | qPCR | 204 | 66 |
| AACAAGTTCCGCAGGGTG | ||||
| LPL | TCCTGGAGTGACGGAATCTGT | qPCR | 174 | 60 |
| GACAGCCAGTCCACCACGAT | ||||
| UXT | GCAAGTGGATTTGGGCTGTAAC | qPCR | 180 | 60 |
| ATGGAGTCCTTGGTGAGGTTGT | ||||
| U6 | TGGAACGCTTCACGAATTTGCG | qPCR | 60 | |
| GGAACGATACAGAGAAGATTAGC | ||||
| px459-U6 | GGCCTATTTCCCATGATTCCT | bacterial liquid PCR | 267 | 60 |
| Name | Sequence 5′-3′ |
|---|---|
| Chi-circ_0006511-si1 | S: GGUGAGAACGGUAACGAGATT |
| A: UCUCCUCUCGUUACCGUUCTT | |
| Chi-circ_0006511-si2 | S: GAGAACGGUAACGAGAGGATT |
| A: UCCUCUCGUUACCGUUCUCTT | |
| Chi-circ_0006511-si3 | S: GAACGGUAACGAGAGGAGATT |
| A: UCUCCUCUCGUUACCGUUCTT | |
| siNC | S: UUCUCCGAACGUGUCACGUTT |
| A: ACGUGACACGUUCGGAGAATT |
| Name | Sequence 5′-3′ |
|---|---|
| miR-87 mimNC | S: UUCUCCGAACGUGUCACGUTT |
| A: ACGUGACACGUUCGGAGAATT | |
| miR-87 mim | S: UGGGCUUGGGGAGCCUGGGACU |
| A: UCCCAGGCUCCCCAAGCCCAUU | |
| miR-87 inhNC | CAGUACUUUUGUGUAGUACAA |
| miR-87 inh | AGUCCCAGGCUCCCCAAGCCCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhang, H.; Wang, Y.; Li, Y.; Wang, Y.; Zhu, J.; Lin, Y. Chi-Circ_0006511 Positively Regulates the Differentiation of Goat Intramuscular Adipocytes via Novel-miR-87/CD36 Axis. Int. J. Mol. Sci. 2022, 23, 12295. https://doi.org/10.3390/ijms232012295
Li X, Zhang H, Wang Y, Li Y, Wang Y, Zhu J, Lin Y. Chi-Circ_0006511 Positively Regulates the Differentiation of Goat Intramuscular Adipocytes via Novel-miR-87/CD36 Axis. International Journal of Molecular Sciences. 2022; 23(20):12295. https://doi.org/10.3390/ijms232012295
Chicago/Turabian StyleLi, Xin, Hao Zhang, Yong Wang, Yanyan Li, Youli Wang, Jiangjiang Zhu, and Yaqiu Lin. 2022. "Chi-Circ_0006511 Positively Regulates the Differentiation of Goat Intramuscular Adipocytes via Novel-miR-87/CD36 Axis" International Journal of Molecular Sciences 23, no. 20: 12295. https://doi.org/10.3390/ijms232012295
APA StyleLi, X., Zhang, H., Wang, Y., Li, Y., Wang, Y., Zhu, J., & Lin, Y. (2022). Chi-Circ_0006511 Positively Regulates the Differentiation of Goat Intramuscular Adipocytes via Novel-miR-87/CD36 Axis. International Journal of Molecular Sciences, 23(20), 12295. https://doi.org/10.3390/ijms232012295