

Integrin Alpha v Beta 6 (αvβ6) and Its Implications in Cancer Treatment

Abstract
:1. Introduction
2. Biology of the αvβ6 Integrin
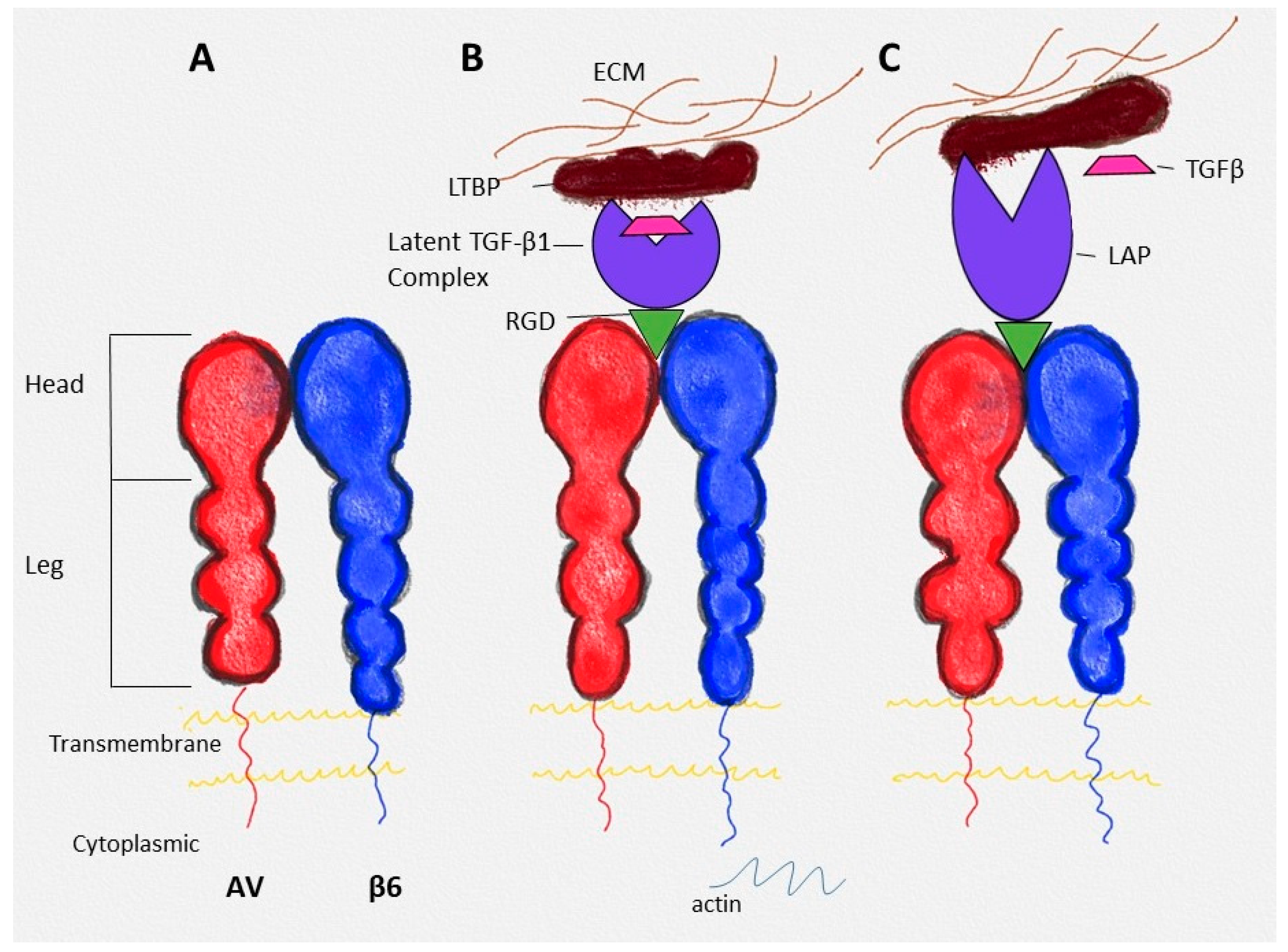
2.1. Structure
2.2. Ligands
2.3. Expression
2.4. Regulation of Expression
3. Crosstalk with Growth Factors in General and Specifically with TGF-β
4. Implication of INGB6 in Cancer and Its Progression
5. Integrin αvβ6 as a Target for Imaging and Therapy
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Campbell, I.D.; Humphries, M.J. Integrin Structure, Activation, and Interactions. Cold Spring Harb. Perspect. Biol. 2011, 3, a004994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef] [Green Version]
- Breuss, J.; Gallo, J.; DeLisser, H.; Klimanskaya, I.; Folkesson, H.; Pittet, J.; Nishimura, S.; Aldape, K.; Landers, D.; Carpenter, W. Expression of the beta 6 integrin subunit in development, neoplasia and tissue repair suggests a role in epithelial remodeling. J. Cell Sci. 1995, 108, 2241–2251. [Google Scholar] [CrossRef]
- Busk, M.; Pytela, R.; Sheppard, D. Characterization of the integrin alpha v beta 6 as a fibronectin-binding protein. J. Biol. Chem. 1992, 267, 5790–5796. [Google Scholar] [CrossRef]
- Fernández-Ruiz, E.; Sánchez-Madrid, F. Regional localization of the human integrin β6 gene (ITGB6) to chromosome 2q24-q31. Genomics 1994, 21, 638–640. [Google Scholar] [CrossRef] [PubMed]
- Takada, Y.; Ye, X.; Simon, S. The integrins. Genome Biol. 2007, 8, 215. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Hudson, N.; Lu, C.; Lin, F.Y.; Li, J.; Xie, Y.; Anderson, J.R.; Bergal, H.T.; Jiang, Y.; Yang, D.; et al. Structural determinants of integrin β-subunit specificity for latent TGF-β. Nat. Struct. Mol. Biol. 2014, 21, 1091–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, E.R.; Humphries, J.D.; James, J.; Jones, M.C.; Askari, J.A.; Humphries, M.J. The integrin adhesome network at a glance. J. Cell Sci. 2016, 129, 4159–4163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.; Lee, C.; Lee, S.; Siu, A.; Ramos, D.M. The Cytoplasmic Extension of the Integrin β6 Subunit Regulates Epithelial–to–Mesenchymal Transition. Anticancer Res. 2014, 34, 659–664. [Google Scholar] [PubMed]
- Morgan, M.R.; Thomas, G.J.; Russell, A.; Hart, I.R.; Marshall, J.F. The integrin cytoplasmic-tail motif EKQKVDLSTDC is sufficient to promote tumor cell invasion mediated by matrix metalloproteinase (MMP)-2 or MMP-9. J. Biol. Chem. 2004, 279, 26533–26539. [Google Scholar] [CrossRef] [PubMed]
- Larjava, H.; Koivisto, L.; Häkkinen, L.; Heino, J. Epithelial integrins with special reference to oral epithelia. J. Dent. Res. 2011, 90, 1367–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Zhao, B.; Iacob, R.E.; Zhu, J.; Koksal, A.C.; Lu, C.; Engen, J.R.; Springer, T.A. Force interacts with macromolecular structure in activation of TGF-β. Nature 2017, 542, 55–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koivisto, L.; Bi, J.; Häkkinen, L.; Larjava, H. Integrin αvβ6: Structure, function and role in health and disease. Int. J. Biochem. Cell Biol. 2018, 99, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Lian, P.-L.; Liu, Z.; Yang, G.-Y.; Zhao, R.; Zhang, Z.-Y.; Chen, Y.-G.; Zhuang, Z.-N.; Xu, K.-S. Integrin αvβ6 and matrix metalloproteinase 9 correlate with survival in gastric cancer. World J. Gastroenterol. 2016, 22, 3852. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Wang, J.; Muhammad, S.; Niu, W.; Liu, E.; Peng, C.; Liang, B.; Sun, Q.; Obo, S.; He, Z. Protein expression of eIF4E and integrin αvβ6 in colon cancer can predict clinical significance, reveal their correlation and imply possible mechanism of interaction. Cell Biosci. 2014, 4, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, C.; Gao, H.; Niu, Z.; Wang, B.; Tan, Z.; Niu, W.; Liu, E.; Wang, J.; Sun, J.; Shahbaz, M. Integrin αvβ6 and transcriptional factor Ets-1 act as prognostic indicators in colorectal cancer. Cell Biosci. 2014, 4, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, N.; Pansino, F.; Baker, M.; Rice, G.; Quinn, M. Association between αvβ6 integrin expression, elevated p42/44 kDa MAPK, and plasminogen-dependent matrix degradation in ovarian cancer. J. Cell Biochem. 2002, 84, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Berghoff, A.S.; Kovanda, A.K.; Melchardt, T.; Bartsch, R.; Hainfellner, J.A.; Sipos, B.; Schittenhelm, J.; Zielinski, C.C.; Widhalm, G.; Dieckmann, K. αvβ3, αvβ5 and αvβ6 integrins in brain metastases of lung cancer. Clin. Exp. Metastasis 2014, 31, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Böger, C.; Kalthoff, H.; Goodman, S.L.; Behrens, H.-M.; Röcken, C. Integrins and their ligands are expressed in non-small cell lung cancer but not correlated with parameters of disease progression. Virchows Arch. 2014, 464, 69–78. [Google Scholar] [CrossRef]
- Ahmed, N.; Riley, C.; Rice, G.E.; Quinn, M.A.; Baker, M.S. αvβ6 integrin-A marker for the malignant potential of epithelial ovarian cancer. J. Histochem. Cytochem. 2002, 50, 1371–1379. [Google Scholar] [CrossRef]
- Xu, M.; Chen, X.; Yin, H.; Yin, L.; Liu, F.; Fu, Y.; Yao, J.; Deng, X. Cloning and characterization of the human integrin β6 gene promoter. PLoS ONE 2015, 10, e0121439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, B.P.; Kassel, K.M.; Manley, S.; Baker, A.K.; Luyendyk, J.P. Regulation of transforming growth factor-β1-dependent integrin β6 expression by p38 mitogen-activated protein kinase in bile duct epithelial cells. J. Pharmacol. Exp. Ther. 2011, 337, 471–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azare, J.; Leslie, K.; Al-Ahmadie, H.; Gerald, W.; Weinreb, P.H.; Violette, S.M.; Bromberg, J. Constitutively activated Stat3 induces tumorigenesis and enhances cell motility of prostate epithelial cells through integrin β6. Mol. Cell. Biol. 2007, 2, 4444–4453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, R.C.; Bellovin, D.I.; Brown, C.; Maynard, E.; Wu, B.; Kawakatsu, H.; Sheppard, D.; Oettgen, P.; Mercurio, A.M. Transcriptional activation of integrin β6 during the epithelial-mesenchymal transition defines a novel prognostic indicator of aggressive colon carcinoma. J. Clin. Investig. 2005, 115, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Massagué, J.; Blain, S.W.; Lo, R.S. TGFbeta signaling in growth control, cancer, and heritable disorders. Cell 2000, 103, 295–309. [Google Scholar] [CrossRef] [Green Version]
- Derynck, R.; Lindquist, P.; Lee, A.; Wen, D.; Tamm, J.; Graycar, J.; Rhee, L.; Mason, A.; Miller, D.; Coffey, R. A new type of transforming growth factor-beta, TGF-beta 3. EMBO J. 1988, 7, 3737–3743. [Google Scholar] [CrossRef]
- López-Casillas, F.; Wrana, J.L.; Massagué, J. Betaglycan presents ligand to the TGFβ signaling receptor. Cell 1993, 73, 1435–1444. [Google Scholar] [CrossRef]
- Niu, J.; Li, Z. The roles of integrin αvβ6 in cancer. Cancer Lett. 2017, 403, 128–137. [Google Scholar] [CrossRef]
- Bretones, G.; Delgado, M.D.; León, J. Myc and cell cycle control. Biochim. Biophys. Acta 2015, 1849, 506–516. [Google Scholar] [CrossRef]
- Ludlow, A.; Yee, K.O.; Lipman, R.; Bronson, R.; Weinreb, P.; Huang, X.; Sheppard, D.; Lawler, J. Characterization of integrin β6 and thrombospondin-1 double-null mice. J. Cell. Mol. Med. 2005, 9, 421–437. [Google Scholar] [CrossRef]
- Bi, J.; Koivisto, L.; Pang, A.; Li, M.; Jiang, G.; Aurora, S.; Wang, Z.; Owen, G.R.; Dai, J.; Shen, Y. Suppression of αvβ6 integrin expression by polymicrobial oral biofilms in gingival epithelial cells. Sci. Rep. 2017, 7, 4411. [Google Scholar] [CrossRef]
- Zhao, R.; Liu, X.-Q.; Wu, X.-P.; Liu, Y.-F.; Zhang, Z.-Y.; Yang, G.-Y.; Guo, S.; Niu, J.; Wang, J.-Y.; Xu, K.-S. Vascular endothelial growth factor (VEGF) enhances gastric carcinoma invasiveness via integrin alpha(v)beta6. Cancer Lett. 2010, 287, 150–156. [Google Scholar] [CrossRef]
- Bandyopadhyay, A.; Raghavan, S. Defining the role of integrin αvβ6 in cancer. Cur. Drug Targ. 2009, 10, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Eberlein, C.; Kendrew, J.; McDaid, K.; Alfred, A.; Kang, J.; Jacobs, V.; Ross, S.; Rooney, C.; Smith, N.; Rinkenberger, J. A human monoclonal antibody 264RAD targeting αvβ6 integrin reduces tumour growth and metastasis, and modulates key biomarkers in vivo. Oncogene 2013, 32, 4406–4416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Principe, D.R.; Doll, J.A.; Bauer, J.; Jung, B.; Munshi, H.G.; Bartholin, L.; Pasche, B.; Lee, C.; Grippo, P.J. TGF-β: Duality of function between tumor prevention and carcinogenesis. J. Nat. Cancer Inst. 2014, 106, djt369. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, T.; Ohba, M.; Ohmori, T. Molecular-targeted therapies for epidermal growth factor receptor and its resistance mechanisms. Int. J. Mol. Sci. 2017, 18, 2420. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Yu, Y.; Sun, C.; Liu, T.; Liang, T.; Zhan, L.; Lin, X.; Feng, X.-H. STAT3 selectively interacts with Smad3 to antagonize TGF-β. Oncogene 2016, 35, 4388–4398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantor, D.I.; Cheruku, H.R.; Nice, E.C.; Baker, M.S. Integrin αvβ6 sets the stage for colorectal cancer metastasis. Cancer Metastasis Rev. 2015, 34, 715–734. [Google Scholar] [CrossRef]
- Santibanez, J.F. Transforming growth factor-Beta and urokinase-type plasminogen activator: Dangerous partners in tumorigenesis—Implications in skin cancer. ISRN Dermatol. 2013, 2013, 597927. [Google Scholar] [CrossRef] [Green Version]
- Mittal, V. Epithelial mesenchymal transition in tumor metastasis. Annu. Rev. Pathol. Mech. Dis. 2018, 13, 395–412. [Google Scholar] [CrossRef] [PubMed]
- Hecht, J.L.; Dolinski, B.M.; Gardner, H.A.; Violette, S.M.; Weinreb, P.H. Overexpression of the alphavbeta6 integrin in endometrial cancer. Appl. Immunohistochem. Mol. Morphol. 2008, 16, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Marsh, D.; Dickinson, S.; Neill, G.W.; Marshall, J.F.; Hart, I.R.; Thomas, G.J. Alpha vbeta 6 Integrin promotesthe invasion of morphoeic basal cell carcinoma through stromal modulation. Cancer Res. 2008, 68, 3295–3303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.Y.; Xu, K.S.; Pan, Z.Q.; Zhang, Z.Y.; Mi, Y.T.; Wang, J.S.; Chen, R.; Niu, J. Integrin alpha v beta 6 mediates thepotential for colon cancer cells to colonize in and metastasize to the liver. Cancer Sci. 2008, 99, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Chen, Y.; Prijatelj, P.; Sakai, T.; Fässler, R.; Sakai, L.Y.; Rifkin, D.B. Fibronectin is required for integrin al-phavbeta6-mediated activation of latent TGF-beta complexes containing LTBP-1. FASEB J. 2005, 19, 1798–1808. [Google Scholar] [CrossRef] [PubMed]
- Munger, J.S.; Huang, X.; Kawakatsu, H.; Griffiths, M.J.; Dalton, S.L.; Wu, J.; Pittet, J.F.; Kaminski, N.; Garat, C.; Matthay, M.A.; et al. The integrin alpha v beta 6 binds and activates latent TGF beta 1: A mech anism for regulating pulmonary inflammation and fibrosis. Cell 1999, 96, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Yokosaki, Y.; Tanaka, K.; Higashikawa, K.; Yamashita, K.; Eboshida, A. Distinct structural requirements for binding of the integrins alphavbeta6, alphavbeta3, alphavbeta5, alpha5beta1 and alpha9beta1 to osteopontin. Matrix Biol. J. Int. Soc. Matrix Biol. 2005, 24, 418–427. [Google Scholar] [CrossRef]
- Annes, J.P.; Rifkin, D.B.; Munger, J.S. The integrin alphaVbeta6 binds and ac tivates latent TGFbeta3. FEBS Lett. 2002, 511, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Weinacker, A.; Chen, A.; Agrez, M.; Cone, R.I.; Nishimura, S.; Wayner, E.; Pytela, R.; Sheppard, D. Role of the integrin alpha v beta 6 in cell attachment to fibronectin. Heter ologous expression of intact and secreted forms of the receptor. J. Biol. Chem. 1994, 269, 6940–6948. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Xu, K.S.; Wang, J.S.; Yang, G.Y.; Wang, W.; Wang, J.Y.; Niu, W.-B.; Liu, E.-Y.; Mi, Y.-T.; Niu, J. Integrin alphanvbeta6 acts as aprognostic indicator in gastric carcinoma. Clin. Oncol. 2008, 20, 61–66. [Google Scholar] [CrossRef]
- Hazelbag, S.; Kenter, G.G.; Gorter, A.; Dreef, E.J.; Koopman, L.A.; Violette, S.M.; Weinreb, P.H.; Fleuren, G.J. Overexpression ofthe alpha v beta 6 integrin in cervical squamous cell carcinoma is a prognostic factor for decreased survival. J. Pathol. 2007, 212, 316–324. [Google Scholar] [CrossRef]
- Jones, J.; Watt, F.M.; Speight, P.M. Changes in the expression of alpha v integrins in oral squamous cellcarcinomas. J. Oral Pathol. Med. 1997, 26, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Hamidi, S.; Salo, T.; Kainulainen, T.; Epstein, J.; Lerner, K.; Larjava, H. Expression of alpha(v)beta6 integrinin oral leukoplakia. Br. J. Cancer 2000, 82, 1433–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, G.J.; Nyström, M.L.; Marshall, J.F. Alphavbeta6 integrin in wound healing and cancer of the oral cavity. J. Oral Pathol. Med. 2006, 35, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sipos, B.; Hahn, D.; Carceller, A.; Piulats, J.; Hedderich, J.; Kalthoff, H.; Goodman, S.L.; Kosmahl, M.; Klöppel, G. Immunohistochemicalscreening for beta6-integrin subunit expression in adenocarcinomas using a novel monoclonal antibody reveals strong up-regulation in pancreatic ductal adenocarcinomas in vivo and in vitro. Histopathology 2004, 45, 226–236. [Google Scholar] [CrossRef]
- Murphy, P.A.; Begum, S.; Hynes, R.O. Tumor angiogenesis in the absence of fibronectin or its cognate integrin receptors. PLoS ONE 2015, 10, e0120872. [Google Scholar] [CrossRef] [Green Version]
- Arihiro, K.; Kaneko, M.; Fujii, S.; Inai, K.; Yokosaki, Y. Significance of alpha 9 beta 1 and alpha v beta 6 integrin expression in breast carcinoma. Breast Cancer 2000, 7, 19–26. [Google Scholar] [CrossRef]
- Katoh, D.; Nagaharu, K.; Shimojo, N.; Hanamura, N.; Yamashita, M.; Kozuka, Y.; Imanaka-Yoshida, K.; Yoshida, T. Binding of alphavbeta1 and alphavbeta6 integrins to tenascin-C induces epithelial-mesenchymal transition-like change of breast cancer cells. Oncogenesis 2013, 2, e65. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, N.; Pansino, F.; Clyde, R.; Murthi, P.; Quinn, M.A.; Rice, G.E.; Agrez, M.V.; Mok, S.; Baker, M.S. Overexpression of alpha(v)beta6 integrin in serous epithelial ovarian cancer regulates extracellular matrix degradation via the plasminogen activation cascade. Carcinogenesis 2002, 23, 237–244. [Google Scholar] [CrossRef]
- Ahn, S.B.; Mohamedali, A.; Anand, S.; Cheruku, H.R.; Birch, D.; Sowmya, G.; Cantor, D.; Ranganathan, S.; Inglis, D.W.; Frank, R.; et al. Characterization of the interaction between heterodimeric alphavbeta6 integrin and urokinase plasminogen activator receptor (uPAR) using functional proteomics. J. Proteome Res. 2014, 13, 5956–5964. [Google Scholar] [CrossRef]
- Wang, J.P.; Hielscher, A. Fibronectin: How its aberrant expression in tumors may improve therapeutic targeting. J. Cancer 2017, 8, 674. [Google Scholar] [CrossRef]
- Liu, H.; Wu, Y.; Wang, F.; Liu, Z. Molecular imaging of integrin αvβ6 expression in living subjects. Am. J. Nucl. Med. Mol. Imaging 2014, 4, 333. [Google Scholar] [PubMed]
- Weinreb, P.H.; Simon, K.J.; Rayhorn, P.; Yang, W.J.; Leone, D.R.; Dolinski, B.M.; Pearse, B.R.; Yokota, Y.; Kawakatsu, H.; Atakilit, A.; et al. Function-blocking Integrin αvβ6Monoclonal Antibodies. J. Biol. Chem. 2004, 279, 17875–17887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koopman Van Aarsen, L.A.; Leone, D.R.; Ho, S.; Dolinski, B.M.; McCoon, P.E.; LePage, D.J.; Kelly, R.; Heaney, G.; Rayhorn, P.; Reid, C.; et al. Antibody-Mediated Blockade of Integrin v 6 Inhibits Tumor Progression In vivo by a Transforming Growth Factor- Regulated Mechanism. Cancer Res. 2008, 68, 561–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olof Olsson, P.; Gustafsson, R.; Salnikov, A.V.; Göthe, M.; Zeller, K.S.; Friman, T.; Baldetorp, B.; Koopman, L.A.; Weinreb, P.H.; Violette, S.M.; et al. Inhibition of integrin αVβ6 changes fibril thickness of stromal collagen in experimental carcinomas. Cell Commun. Signal. 2018, 16, 36. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Y.; Xu, K.-S.; He, Q.-S.; Niu, W.-B.; Wang, J.-Y.; Mi, Y.-T.; Wang, J.-S.; Qiang, G.-Q.; Yang, G.-Y.; Niu, J. Signaling and regulatory mechanisms of integrin ανβ6 on the apoptosis of colon cancer cells. Cancer Lett. 2008, 66, 209–215. [Google Scholar] [CrossRef]
- Kogelberg, H.; Tolner, B.; Thomas, G.J.; Di Cara, D.; Minogue, S.; Ramesh, B.; Sodha, S.; Marsh, D.; Lowdell, M.W.; Meyer, T.; et al. Engineering a Single-Chain Fv Antibody to αvβ6 Integrin Using the Specificity-Determining Loop of a Foot-and-Mouth Disease Virus. J. Mol. Biol. 2008, 382, 385–401. [Google Scholar] [CrossRef] [Green Version]
- Reader, C.S.; Vallath, S.; Steele, C.W.; Haider, S.; Brentnall, A.; Desai, A.; Moore, K.M.; Jamieson, N.B.; Chang, D.; Bailey, P.; et al. The integrin αvβ6 drives pancreatic cancer through diverse mechanisms and represents an effective target for therapy. J. Pathol. 2019, 249, 332–342. [Google Scholar] [CrossRef] [Green Version]
- Kogelberg, H.; Miranda, E.; Burnet, J.; Ellison, D.; Tolner, B.; Foster, J.; Picón, C.; Thomas, G.J.; Meyer, T.; Marshall, J.F. Generation and characterization of a diabody targeting the αvβ6 integrin. PLoS ONE 2013, 8, e73260. [Google Scholar] [CrossRef] [Green Version]
- Moore, K.M.; Desai, A.; Delgado, B.d.L.; Trabulo, S.M.D.; Reader, C.; Brown, N.F.; Murray, E.R.; Brentnall, A.; Howard, P.; Masterson, L.; et al. Integrin αvβ6-specific therapy for pancreatic cancer developed from foot-and-mouth-disease virus. Theranostics 2020, 10, 2930–2942. [Google Scholar] [CrossRef]
- Busenhart, P.; Montalban-Arques, A.; Katkeviciute, E.; Morsy, Y.; van Passen, C.; Hering, L.; Atrott, K.; Lang, S.; Garzon, J.F.G.; Naschberger, E.; et al. Inhibition of integrin αvβ6 sparks T-cell antitumor response and enhances immune checkpoint blockade therapy in colorectal cancer. J. Immunother. Cancer 2022, 10, e003465. [Google Scholar] [CrossRef]
- Bagati, A.; Kumar, S.; Jiang, P.; Pyrdol, J.; Zou, A.E.; Godicelj, A.; Mathewson, N.D.; Cartwright, A.N.R.; Cejas, P.; Brown, M.; et al. Integrin αvβ6-TGFβ-SOX4 pathway drives immune evasion in triple-negative breast cancer. Cancer Cell. 2021, 39, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Saleem, A.; Helo, Y.; Win, Z.; Dale, R.; Cook, J.; Searle, G.E.; Wells, P. Integrin αvβ6 Positron Emission Tomography Imaging in Lung Cancer Patients Treated with Pulmonary Radiation Therapy. Int. J. Radiat. Oncol. Biol. Phys. 2020, 107, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Hausner, S.H.; Bold, R.J.; Cheuy, L.Y.; Chew, H.K.; Daly, M.E.; Davis, R.A.; Foster, C.C.; Kim, E.J.; Sutcliffe, J.L. Preclinical Development and First-in-Human Imaging of the Integrin αvβ6 with [18F]αvβ6-Binding Peptide in Metastatic Carcinoma. Clin. Cancer Res. 2019, 25, 1206–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.B.; Hu, L.Y.; Boucher, D.L.; Sutcliffe, J.L. ImmunoPET Imaging of αvβ6 Expression Using an Engineered Anti-αvβ6 Cys-diabody Site-Specifically Radiolabeled with Cu-64: Considerations for Optimal Imaging with Antibody Fragments. Mol. Imaging Biol. 2017, 20, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Meecham, A.; Marshall, J. Harnessing the power of foot-and-mouth-disease virus for targeting integrin alpha-v beta-6 for the therapy of cancer. Exp. Opin. Drug Disc. 2021, 16, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Search of: αvβ6—List Results—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/results?cond=&term=%CE%B1v%CE%B26+&cntry=&state=&city=&dist= (accessed on 10 October 2022).




{kind=link}
{kind=link}
{kind=link}
| Cancer | Functions |
|---|---|
| Endometrial | Promotes invasiveness [41] |
| Basal Cell | Promotes invasiveness [42] |
| Liver | Poor prognosis [43] |
| Colon Cancer | Involved in cell adhesion and migration, activation of TGF-B, regulation of extracellular proteases, poor prognosis marker and implications in cancer metastasis to liver [43,44,45,46,47,48] |
| Gastric | Prognostic marker [49] |
| Cervical Squamous | Unfavourable marker [50] |
| Oral SCC | Increased expression in invasive stage of the cancer, regulation of migration and adhesion of cancer cells [51,52,53] |
| Pancreatic | Enhanced expression, regulation of tumour angiogenesis [54,55] |
| Breast | EMT induction and invasiveness [56,57] |
| Ovary | Enhanced expression leads to promotion of metastasis [58,59] |
| Company | Target | Molecule ID/Name | Molecule Type | Drug Disease | Highest Status Reached | Summary |
|---|---|---|---|---|---|---|
| AstraZeneca | integrin subunit alpha V integrin subunit beta 6 | 264RAD | Antibody | Cancer, pancreatic | Preclinical, no active development reported. | 264RAD is a human therapeutic antibody targeting alphavβ6 integrin, which was under development by AstraZeneca for the treatment of pancreatic cancer. |
| Aura Biosciences Cancer Research Technology | integrin subunit alpha V integrin subunit beta 6 | [19F] FAB-A20FMDV2; A20FMDV2; | Synthetic peptide | Cancer, pancreatic diagnosis, cancer | Preclinical, no active development reported. | A20FMDV2 is a lead integrin alphaVβ6 binding peptides, which was under development by Cancer Research Technology (CRT) for use in targeted delivery of anticancer compounds and for cancer imaging. |
| Merck KGaA | integrin subunit alpha V integrin subunit beta 1 integrin subunit beta 3 integrin subunit beta 5 integrin subunit beta 6 integrin subunit beta 8 | Abituzumab | Antibody | Cancer, solid | Phase II, no active development reported. | Abituzumab is an investigational integrin inhibiting monoclonal antibody with activity against avβ1, 3, 5, 6, and 8 integrin heterodimers, which was under development by Merck KGaA for the treatment of colorectal cancer. It was previously developed for systemic sclerosis. |
| Venn Therapeutics | integrin subunit alpha V integrin subunit beta 6 integrin subunit beta 8 transmembrane protein 173 | Ad-VCA0848; AdVCA0848; VTX 002; | Antibody | Cancer, unspecified. Chronic obstructive pulmonary disease, idiopathic pulmonary fibrosis | Pre-clinical, no active development reported. | Ad-VCA-0848 (VTX-002) is a novel, immunotherapeutic off-the-shelf humanized monoclonal antibody STING agonist which recombined a potent diguanylate cyclase gene, VCA-0848, into a nonreplicating adenovirus serotype 5 targeting both alphavbeta8 and alphavbeta6, which was under development by Venn Therapeutics for the treatment of pulmonary fibrosis and COPD. It was previously under development for the treatment of cancer. |
| GlaxoSmithKline | integrin subunit beta 6 | alpha v integrin antagonists, GlaxoSmithKline; avβ6 antagonists; avβ6 integrin anatagonists; integrin avβ6 anatagonists | Small molecule | Idiopathic pulmonary fibrosis | Preclinical, no active development reported. | GlaxoSmithKline was developing alpha v beta 6 integrin antagonists for the treatment of pulmonary fibrosis. |
| DiCE Therapeutics | integrin subunit alpha V integrin subunit beta 1 integrin subunit beta 6 | alphaVbeta1/alphaVbeta6 integrin antagonist, DiCE Therapeutics; aVβ1/aVβ6 integrin antagonist, DiCE Therapeutics | Small molecule | Idiopathic pulmonary fibrosis | Preclinical, development active | DiCE Therapeutics (previously Dice Molecules) is developing an orally available alphaVbeta1/alphaVbeta6 (aVβ1/aVβ6) integrin antagonist using its DELSCAPE platform, for the treatment of idiopathic pulmonary fibrosis. |
| Seagen | integrin subunit alpha V integrin subunit beta 6 | anti-alpha-V-beta6 ADC, Seattle; anti-avβ6 ADC, Seattle | Small molecule | Cancer, unspecified | Preclinical, no active development reported. | Seattle was developing a MAb against Integrin alpha-v-beta6, using its antibody–drug conjugate (ADC) technology, for the treatment of lung, pancreatic, and head and neck cancers. |
| Cancer Research Technology Cancer Research Technology | integrin subunit alpha V integrin subunit beta 6 | anti-alphavβ6 antibodies, CRT; anti-integrin avss6 antibodies, CRT | Antibody | Cancer, unspecified; fibrosis, unspecified | Preclinical, no active development reported. | Cancer Research Technology (CRT) was developing anti-alphavβ6 antibodies for the treatment of cancer and fibrotic diseases. |
| Biogen | integrin subunit alpha V integrin subunit beta 6 | alphav beta 6 integrin inhibitor, Biogen Idec; BG 00011; BG-00011; BG00011; STX-100; STX100 | Antibody | Cancer, unspecified; pulmonary fibrosis, nephropathy | Phase II clinical trials, no active development reported. | Biogen Idec (Stromedix) has discontinued development of BG-00011 (STX-100), a humanized mAb targeting the alpha v beta 6 integrin for the treatment of idiopathic pulmonary fibrosis (IPF), due to safety concerns. |
| Panorama Research Corbus Pharmaceuticals | integrin subunit alpha V integrin subunit beta 6 integrin subunit beta 8 | CRB 602; CRB-602; CRB602 | Antibody | Cancer, unspecified; fibrosis, unspecified | Preclinical, development active | CRB-602 is an anti-alphavbeta6 and alphavbeta8 (avβ6/8) monoclonal antibody under development by Panorama Research in collaboration with Corbus Pharmaceuticals for the treatment of fibrosis and cancer. |
| GlaxoSmithKline | integrin subunit alpha V integrin subunit beta 6 | alpha V beta 6 integrin antagonists, GlaxoSmithKline; GSK 3008348; GSK-3008348; GSK3008348 | Small molecule | Idiopathic pulmonary fibrosis | Phase I clinical trial, no active development reported. | GlaxoSmithKline has discontinued development of GSK-3008348, an alphaVbeta 6 integrin antagonist for the treatment of idiopathic pulmonary fibrosis, due to a lack of confidence in developability and portfolio considerations following an interim analysis of Phase Ib data. It is an inhaled formulation. |
| Indalo Therapeutics | integrin subunit alpha V integrin subunit beta 1 integrin subunit beta 3 integrin subunit beta 6 | Fibrotic disease therapy, Indalo Therapeutics; IDL 2965; IDL-2965; IDL2965 | Small molecule | Chronic renal failure, idiopathic pulmonary fibrosis, non-alcoholic steatohepatitis | Phase I clinical trial, no active development reported. | IDL-2965 is an oral, selective RGD-binding integrin (alphaVbeta1, alphaVbeta3 and alphaVbeta6) antagonist, which was under development by Indalo Therapeutics for the treatment of idiopathic pulmonary fibrosis and non-alcoholic steatohepatitis. RGD-binding integrin inhibits the activation of TGF-β as well as the ability of stiff extracellular matrix to promote fibroblast migration and survival. |
| Centocor | integrin subunit alpha V integrin subunit beta 3 integrin subunit beta 5 integrin subunit beta 6 | BGB-101; BGB101; CNTO 95; CNTO-095; CNTO-95; intetumumab | Antibody | Cancer, solid | Phase II clinical trials, no active development reported. | Intetumumab is a high-affinity fully-human MAb, which was under development by Centocor (Johnson & Johnson (J&J)) for the treatment of cancer. It binds and inhibits integrins alphavβ1, alphavβ3, alphavβ5, alphavβ6 and alphavβ8, and has antiangiogenic activity It was generated using Medarex’s (now Bristol-Myers Squibb’s (BMS)) UltiMAb technology. |
| AbbVie/Morphic Therapeutic | integrin subunit alpha V integrin subunit beta 6 | alphaVbeta6 small molecule inhibitor, Morphic Therapeutic; integrin therapies, Morphic Therapeutics; MORF 627; MORF beta6; MORF-627; MORF-beta6; MORF627; MORFbeta6 | Small molecule | Non-alcoholic steatohepatitis, Primary sclerosing cholangitis | Preclinical, no active development reported. | AbbVie has discontinued development of MORF-627 (MORF-beta6), an oral selective alphaVbeta6 small-molecule inhibitor licensed from Morphic Therapeutic, for the treatment of primary sclerosing cholangitis due to safety concerns. It was previously under development for non-alcoholic steatohepatitis. |
| Morphic Therapeutic/AbbVie | integrin subunit alpha V integrin subunit beta 6 | MORF 720; MORF-720; MORF720 | Small molecule | Fibrosis, pulmonary, idiopathic | Preclinical, no active development reported. | AbbVie has discontinued development of MORF-720, an oral selective alphaVbeta6 small-molecule inhibitor, licensed from Morphic Therapeutic using its MlnT platform, for the treatment of idiopathic pulmonary fibrosis. |
| Pliant Therapeutics | integrin subunit alpha V integrin subunit beta 1 integrin subunit beta 6 | avβ6/avβ1 dual integrin inhibitor, Pliant Therapeutics; PLN 1561; PLN-1561; PLN1561 | Small molecule | Primary sclerosing cholangitis | Preclinical, no active development reported. | PLN-1561 is an avβ6/aVβ1 dual integrin inhibitor, which was under development by Pliant Therapeutics for the treatment of primary sclerosing cholangitis. |
| Pliant Therapeutics | integrin subunit alpha V integrin subunit beta 6 | aVβ6 inhibitor, Pliant Therapeutics; PLN 1705; PLN-1705; PLN1705 | Small molecule | Primary sclerosing cholangitis | Preclinical, no active development reported. | PLN-1705 is an aVβ6 selective inhibitor, which was under development by Pliant Therapeutics for the treatment of primary sclerosing cholangitis. |
| Pliant Therapeutics | integrin subunit alpha V integrin subunit beta 1 integrin subunit beta 6 | avβ1/avβ6 dual integrin inhibitor, Pliant; avβ1/avβ6 integrin inhibitor, Pliant; PLN 74809; PLN-74809; PLN74809 | Small Molecule | Idiopathic pulmonary fibrosis, primary sclerosing cholangitis, COVID-19 complications, acute respiratory distress syndrome | Phase II clinical trial, active development reported. | PLN-74809 is an oral, small-molecule avbeta1/avbeta 6 dual integrin inhibitor under development by Pliant Therapeutics for the treatment of idiopathic pulmonary fibrosis and primary sclerosing cholangitis. The avβ1/avβ6 integrin blocks the activation of TGF-β in a tissue-specific manner, preventing the growth of fibrotic tissue within the lung. It was previously under development for acute respiratory distress syndrome (ARDS) associated with COVID-19. |
| OcuTerra Therapeutics | integrin subunit alpha V integrin subunit beta 3 integrin subunit beta 6 integrin subunit beta 8 | OTT 166; OTT-166; OTT166; SF 0166; SF-0166; SF0166 | Small molecule | Retinopathy, diabetic macular degeneration, age-related, wet oedema, macular, diabetic | Phase II clinical trial, development active. | SF-0166 is a fluorinated, selective small-molecule inhibitor of integrin alphaVbeta3, alphaVbeta6 and alphaVbeta8, which is under development by SciFluor Life Sciences (now OcuTerra Therapeutics) for the treatment of diabetic retinopathy. It was previously in development for retinal disease, including wet age-related macular degeneration (AMD) and diabetic macular oedema (DME). It is administered topically to the eye. |
| Seagen | integrin subunit beta 6 secretory leukocyte peptidase inhibitor | SGN B6A; SGN-B6A; SGNB6A | Small molecule | Cancer, solid, unspecified | Phase I clinical trial, development active. | SGN-B6A is an antibody-drug conjugate (ADC) targeting integrin beta-6, which is under development by Seagen (formerly Seattle Genetics) for the treatment of solid tumours. |
| Ligand Pharmaceuticals | integrin subunit alpha 5 integrin subunit alpha V integrin subunit beta 1 integrin subunit beta 3 integrin subunit beta 5 integrin subunit beta 6 | XC 201; XC-201; XC201 | Recombinant peptide | Cancer, unspecified | Preclinical, active development reported. | XC-201 is a peptide-Fc fusion (mAb-lite) targeting avβ1, avβ3, avβ5, avβ6, and a5β1 integrins, which was under development by xCella Biosciences for the treatment of cancer. A unique engineered integrin-binding peptide is created, and it is genetically fused to an antibody Fc domain. It is a universal tumour targeting agent that binds a broad panel of integrins (av and a5) expressed at high levels on tumours. |
| Title | Status | Conditions | Interventions | Locations | Phase | Reference |
|---|---|---|---|---|---|---|
| First-in-Human Positron Emission Tomography Study Using the 18FαVβ6-Binding-Peptide | Recruiting | Breast carcinoma|Colorectal carcinoma|Lung carcinoma|Metastatic malignant neoplasm in the breast|Metastatic malignant neoplasm in the colon|Metastatic malignant neoplasm in the lung|Metastatic malignant neoplasm in the rectum|Pancreatic carcinoma | Drug: 18F-αVβ6-BP | University of California Davis Comprehensive Cancer Center, Sacramento, California, United States | Early Phase 1 | https://ClinicalTrials.gov/show/NCT03164486, accessed on 10 October 2022 |
| Phase 2a Evaluation of PLN-74809 on αVβ6 Receptor Occupancy Using PET Imaging in Participants With IPF/ | Recruiting | Idiopathic pulmonary fibrosis | Drug: PLN-74809|Radiation: Knottin tracer | Stanford Medical Center, Palo Alto, California, United States | Phase 2 | https://ClinicalTrials.gov/show/NCT04072315, accessed on 10 October 2022 |
| PET/CT Imaging in COVID-19 Patients | Enrolling by invitation | COVID-19|SARS-CoV-2 Infection | Drug: 18F-αVβ6-BP | University of California Davis, Sacramento, California, United States | Early Phase 1 | https://ClinicalTrials.gov/show/NCT04376593, accessed on 10 October 2022 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brzozowska, E.; Deshmukh, S. Integrin Alpha v Beta 6 (αvβ6) and Its Implications in Cancer Treatment. Int. J. Mol. Sci. 2022, 23, 12346. https://doi.org/10.3390/ijms232012346
Brzozowska E, Deshmukh S. Integrin Alpha v Beta 6 (αvβ6) and Its Implications in Cancer Treatment. International Journal of Molecular Sciences. 2022; 23(20):12346. https://doi.org/10.3390/ijms232012346
Chicago/Turabian StyleBrzozowska, Ewa, and Sameer Deshmukh. 2022. "Integrin Alpha v Beta 6 (αvβ6) and Its Implications in Cancer Treatment" International Journal of Molecular Sciences 23, no. 20: 12346. https://doi.org/10.3390/ijms232012346
APA StyleBrzozowska, E., & Deshmukh, S. (2022). Integrin Alpha v Beta 6 (αvβ6) and Its Implications in Cancer Treatment. International Journal of Molecular Sciences, 23(20), 12346. https://doi.org/10.3390/ijms232012346