

Insights into Carbapenem Resistance in Vibrio Species: Current Status and Future Perspectives

 ,
,  , , , ,
, , , ,  ,
,  ,
,  and
and
Abstract
:1. Introduction
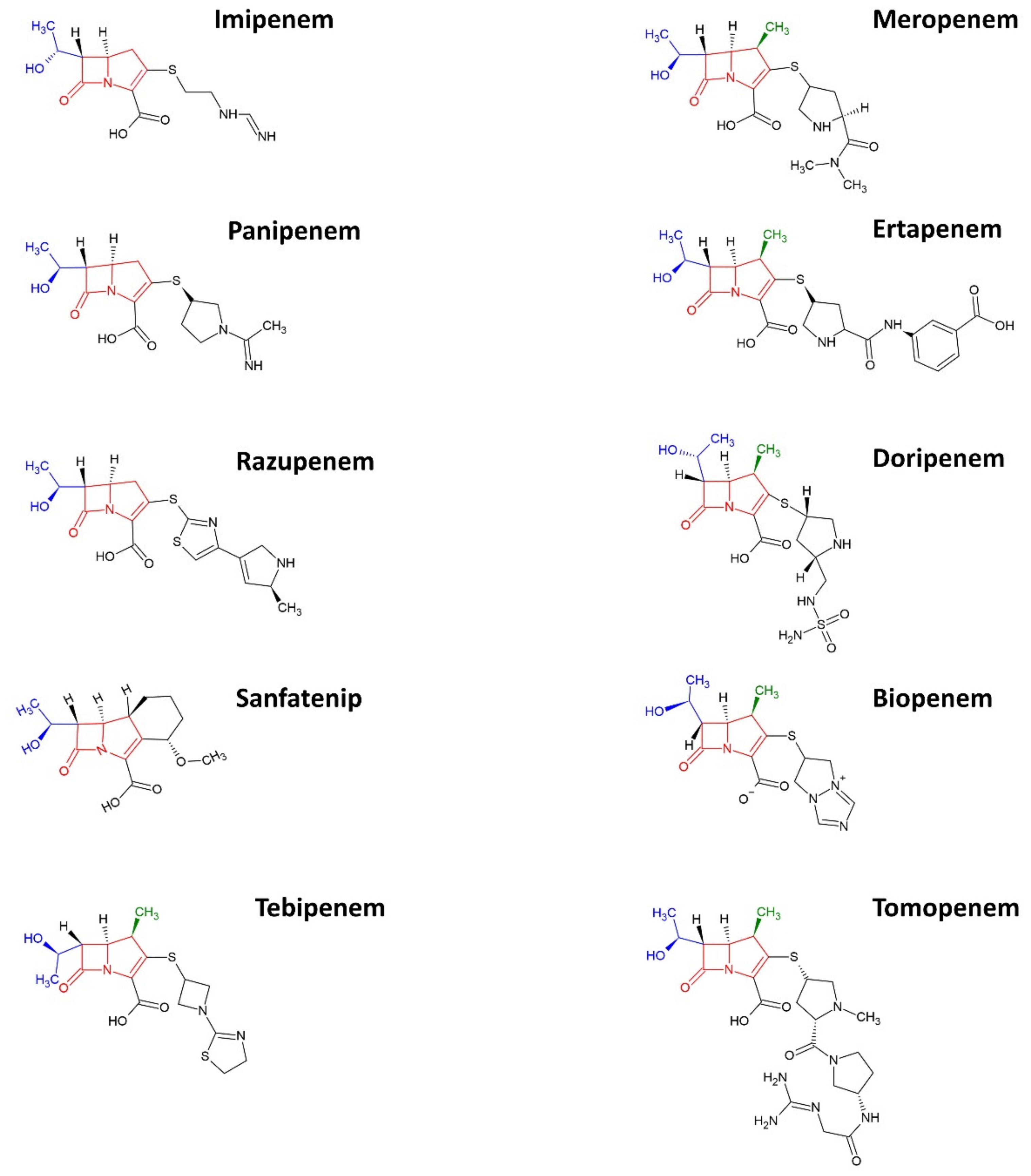
2. Carbapenems
3. Vibrio spp.
3.1. Vibrio spp. as the Indicator Strain of Carbapenem Resistance in the Environment
4. Prevalence of Carbapenem Resistance
5. Mechanism to Carbapenem Resistance Development
5.1. Limiting the Intracellular Concentration of Drug
5.2. Carbapenemase
5.3. Resistance Gene Transfer
5.4. Antimicrobial Tolerance
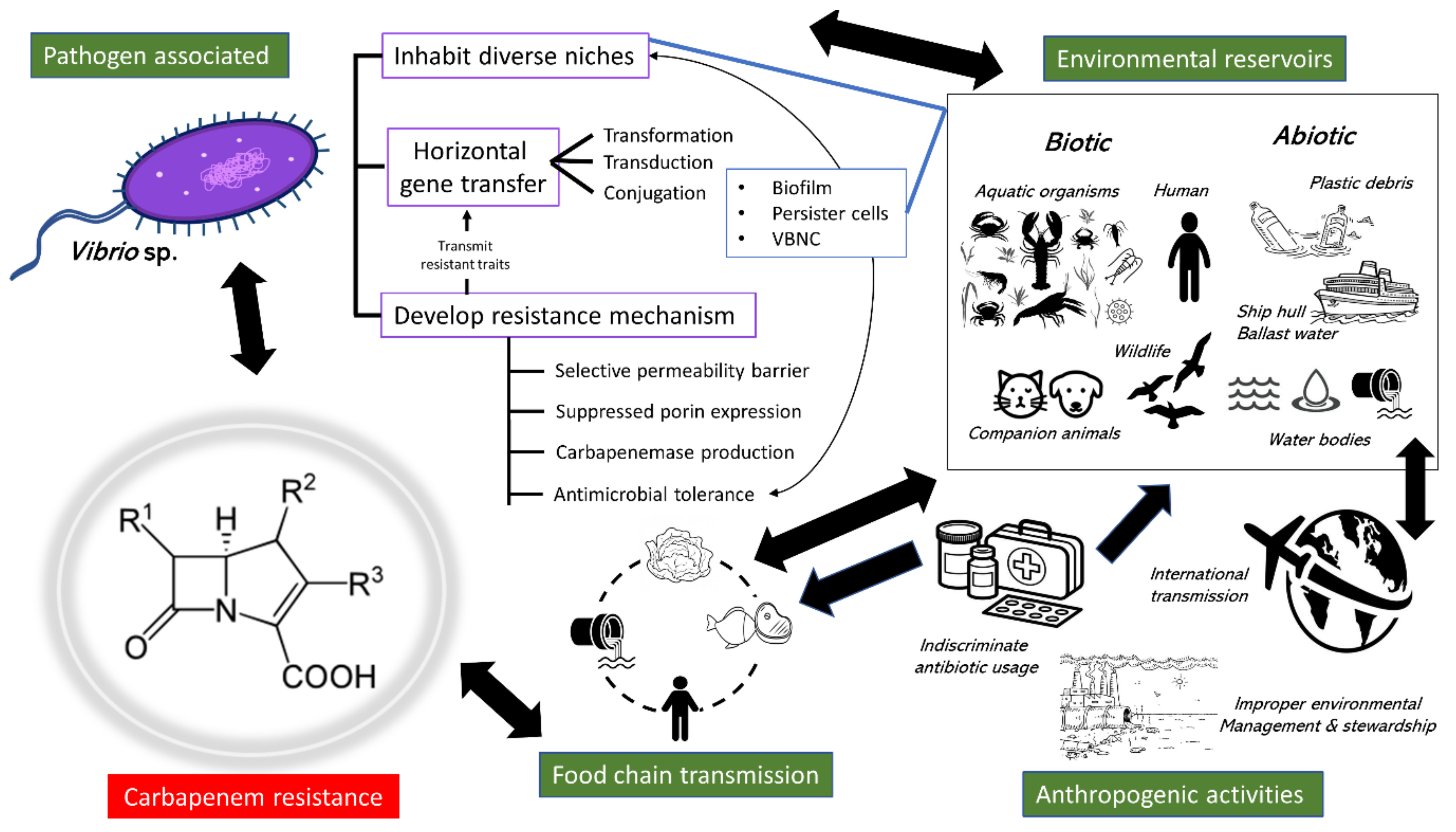
6. Dissemination Pathways
6.1. Excessive Clinical Usage
6.2. Transmission via the Food Chain
6.3. Environmental Reservoirs
6.4. Anthropogenic Factors
7. Future Research Perspectives
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Huang, W.; Qiao, F.; Zhang, Y.; Huang, J.; Deng, Y.; Li, J.; Zong, Z. In-hospital Medical Costs of Infections Caused by Carbapenem-resistant Klebsiella pneumoniae. Clin. Infect. Dis. 2018, 67, S225–S230. [Google Scholar] [CrossRef] [PubMed]
- Hofer, U. The cost of antimicrobial resistance. Nat. Rev. Genet. 2019, 17, 3. [Google Scholar] [CrossRef] [PubMed]
- Vasoo, S.; Barreto, J.N.; Tosh, P.K. Emerging issues in gram-negative bacterial resistance: An update for the practicing clinician. In Mayo Clinic Proceedings; Elsevier: Amsterdam, The Netherlands, 2015; pp. 395–403. [Google Scholar]
- Patel, G.; Bonomo, R. Stormy waters ahead: Global emergence of carbapenemases. Front. Microbiol. 2013, 4, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, A.; Fahrbach, K.; Zhao, Q.; Lodise, T. Association Between Carbapenem Resistance and Mortality Among Adult, Hospitalized Patients with Serious Infections Due to Enterobacteriaceae: Results of a Systematic Literature Review and Meta-analysis. Open Forum Infect. Dis. 2018, 5, ofy150. [Google Scholar] [CrossRef]
- World Health Organization. Critically important antimicrobials for human medicine: Categorization for the development of risk management strategies to contain antimicrobial resistance due to non-human antimicrobial use. In Proceedings of the Report of the Second WHO Expert Meeting, Copenhagen, Denmark, 29–31 May 2007. [Google Scholar]
- Tzouvelekis, L.S.; Markogiannakis, A.; Psichogiou, M.; Tassios, P.T.; Daikos, G.L. Carbapenemases in Klebsiella pneumoniae and Other Enterobacteriaceae: An Evolving Crisis of Global Dimensions. Clin. Microbiol. Rev. 2012, 25, 682–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottesman, T.; Agmon, O.; Shwartz, O.; Dan, M. Household Transmission of Carbapenemase-producing Klebsiella pneumoniae. Emerg. Infect. Dis. 2008, 14, 859–860. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L. The difficult-to-control spread of carbapenemase producers among Enterobacteriaceae worldwide. Clin. Microbiol. Infect. 2014, 20, 821–830. [Google Scholar] [CrossRef] [Green Version]
- Won, S.Y.; Munoz-Price, L.S.; Lolans, K.; Hota, B.; Weinstein, R.A.; Hayden, M.K.; Centers for Disease Control and Prevention Epicenter Program. Emergence and Rapid Regional Spread of Klebsiella pneumoniae Carbapenemase-Producing Enterobacteriaceae. Clin. Infect. Dis. 2011, 53, 532–540. [Google Scholar] [CrossRef] [Green Version]
- Snitkin, E.S.; Zelazny, A.M.; Thomas, P.J.; Stock, F.; Comparative Sequencing Program, N.I.S.C.; Henderson, D.K.; Palmore, T.N.; Segre, J.A. Tracking a Hospital Outbreak of Carbapenem-Resistant Klebsiella pneumoniae with Whole-Genome Sequencing. Sci. Transl. Med. 2012, 4, 148ra116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhen, X.; Chen, Y.; Hu, X.; Dong, P.; Gu, S.; Sheng, Y.Y.; Dong, H. The difference in medical costs between carbapenem-resistant Acinetobacter baumannii and non-resistant groups: A case study from a hospital in Zhejiang province, China. Eur. J. Clin. Microbiol. 2017, 36, 1989–1994. [Google Scholar] [CrossRef]
- Falagas, M.E.; Tansarli, G.S.; Karageorgopoulos, D.E.; Vardakas, K.Z. Deaths Attributable to Carbapenem-Resistant Entero bacteriaceae Infections. Emerg. Infect. Dis. 2014, 20, 1170–1175. [Google Scholar] [CrossRef] [PubMed]
- Galler, H.; Feierl, G.; Petternel, C.; Reinthaler, F.F.; Haas, D.; Grisold, A.J.; Luxner, J.; Zarfel, G. KPC-2 and OXA-48 carbapenemase-harbouring Enterobacteriaceae detected in an Austrian wastewater treatment plant. Clin. Microbiol. Infect. 2014, 20, O132–O134. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Huang, L.; Li, L.; Yang, Y.; Mao, D.; Luo, Y. Discharge of KPC-2 genes from the WWTPs contributed to their enriched abundance in the receiving river. Sci. Total Environ. 2017, 581–582, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Fernando, D.M.; Tun, H.M.; Poole, J.; Patidar, R.; Li, R.; Mi, R.; Amarawansha, G.E.; Fernando, W.D.; Khafipour, E.; Farenhorst, A.; et al. Detection of Antibiotic Resistance Genes in Source and Drinking Water Samples from a First Nations Community in Canada. Appl. Environ. Microbiol. 2016, 82, 4767–4775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacão, M.; Correia, A.; Henriques, I.S. Low Prevalence of Carbapenem-Resistant Bacteria in River Water: Resistance Is Mostly Related to Intrinsic Mechanisms. Microb. Drug Resist. 2015, 21, 497–506. [Google Scholar] [CrossRef]
- Tafoukt, R.; Touati, A.; Leangapichart, T.; Bakour, S.; Rolain, J.-M. Characterization of OXA-48-like-producing Enterobacteriaceae isolated from river water in Algeria. Water Res. 2017, 120, 185–189. [Google Scholar] [CrossRef]
- Nascimento, T.; Cantamessa, R.; Melo, L.; Fernandes, M.R.; Fraga, E.; Dropa, M.; Sato, M.I.; Cerdeira, L.; Lincopan, N. International high-risk clones of Klebsiella pneumoniae KPC-2/CC258 and Escherichia coli CTX-M-15/CC10 in urban lake waters. Sci. Total Environ. 2017, 598, 910–915. [Google Scholar] [CrossRef]
- Canellas, A.L.; Lopes, I.R.; Mello, M.P.; Paranhos, R.; de Oliveira, B.F.; Laport, M.S. Vibrio species in an urban tropical estuary: Antimicrobial susceptibility, interaction with environmental parameters, and possible public health outcomes. Microorganisms 2021, 9, 1007. [Google Scholar] [CrossRef]
- Bier, N.; Schwartz, K.; Guerra, B.; Strauch, E. Survey on antimicrobial resistance patterns in Vibrio vulnificus and Vibrio cholerae non-O1/non-O139 in Germany reveals carbapenemase-producing Vibrio cholerae in coastal waters. Front. Microbiol. 2015, 6, 1179. [Google Scholar] [CrossRef] [Green Version]
- Dewi, D.A.P.R.; Götz, B.; Thomas, T. Diversity and Genetic Basis for Carbapenem Resistance in a Coastal Marine Environment. Appl. Environ. Microbiol. 2020, 86, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Xin, R.; Zhang, K.; Wu, N.; Zhang, Y.; Niu, Z. The pollution level of the blaOXA-58 carbapenemase gene in coastal water and its host bacteria characteristics. Environ. Pollut. 2019, 244, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Fischer, J.; Helmuth, R. An emerging public health problem: Acquired carbapenemase-producing microorganisms are present in food-producing animals, their environment, companion animals and wild birds. Vet. Microbiol. 2014, 171, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Zurfluh, K.; Poirel, L.; Nordmann, P.; Klumpp, J.; Stephan, R. First detection of Klebsiella variicola producing OXA-181 carbapenemase in fresh vegetable imported from Asia to Switzerland. Antimicrob. Resist. Infect. Control 2015, 4, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, B.J.; Rubin, J.E. Carbapenemase Producing Bacteria in the Food Supply Escaping Detection. PLoS ONE 2015, 10, e0126717. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.-H.; Ab Mutalib, N.-S.; Law, J.W.-F.; Wong, S.H.; Letchumanan, V. Discovery on Antibiotic Resistance Patterns of Vibrio parahaemolyticus in Selangor Reveals Carbapenemase Producing Vibrio parahaemolyticus in Marine and Freshwater Fish. Front. Microbiol. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Letchumanan, V.; Ab Mutalib, N.-S.; Wong, S.H.; Chan, K.-G.; Lee, L.-H. Determination of antibiotic resistance patterns of Vibrio parahaemolyticus from shrimp and shellfish in Selangor, Malaysia. Prog. Microbes Mol. Biol. 2019, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Letchumanan, V.; Pusparajah, P.; Tan, L.T.-H.; Yin, W.-F.; Lee, L.-H.; Chan, K.-G. Occurrence and Antibiotic Resistance of Vibrio parahaemolyticus from Shellfish in Selangor, Malaysia. Front. Microbiol. 2015, 6, 1417. [Google Scholar] [CrossRef] [Green Version]
- Letchumanan, V.; Yin, W.-F.; Lee, L.-H.; Chan, K.-G. Prevalence and antimicrobial susceptibility of Vibrio parahaemolyticus isolated from retail shrimps in Malaysia. Front. Microbiol. 2015, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Mangat, C.S.; Boyd, D.; Janecko, N.; Martz, S.-L.; Desruisseau, A.; Carpenter, M.; Reid-Smith, R.J.; Mulvey, M.R. Characterization of VCC-1, a Novel Ambler Class A Carbapenemase from Vibrio cholerae Isolated from Imported Retail Shrimp Sold in Canada. Antimicrob. Agents Chemother. 2016, 60, 1819–1825. [Google Scholar] [CrossRef] [Green Version]
- Venggadasamy, V.; Tan, L.T.-H.; Law, J.W.F.; Ser, H.-L.; Letchumanan, V.; Pusparajah, P. Incidence, Antibiotic Susceptibility and Characterization of Vibrio parahaemolyticus Isolated from Seafood in Selangor, Malaysia. Prog. Microbes Mol. Biol. 2021, 4, 1–34. [Google Scholar] [CrossRef]
- Baker-Austin, C.; McArthur, J.V.; Lindell, A.H.; Wright, M.S.; Tuckfield, R.C.; Gooch, J.; Warner, L.; Oliver, J.; Stepanauskas, R. Multi-site Analysis Reveals Widespread Antibiotic Resistance in the Marine Pathogen Vibrio vulnificus. Microb. Ecol. 2009, 57, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.V.; Joseph, T.C.; Peeralil, S.; Koombankallil, R.; Vaiyapuri, M.; Mothadaka, M.P.; Lalitha, K.V. Tropical shrimp aquaculture farms harbour pathogenic Vibrio parahaemolyticus with high genetic diversity and Carbapenam resistance. Mar. Pollut. Bull. 2020, 160, 111551. [Google Scholar] [CrossRef]
- Scotta, C.; Juan, C.; Cabot, G.; Oliver, A.; Lalucat, J.; Bennasar, A.; Albertí, S. Environmental Microbiota Represents a Natural Reservoir for Dissemination of Clinically Relevant Metallo-beta-Lactamases. Antimicrob. Agents Chemother. 2011, 55, 5376–5379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galal, A.M.; Gul, W.; Noreddin, A.M.; Slade, D. An update on the synthesis and antibacterial effects of carbapenems. Recent Patents Anti-Infective Drug Discov. 2010, 5, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, M.; Mmohammadi-Sichani, M.; Karbasizadeh, V. Prevalence of SHV, TEM, CTX-M and OXA-48 β-Lactamase genes in clinical isolates of Pseudomonas aeruginosa in Bandar-Abbas, Iran. Avicenna J. Clin. Microbiol. Infect. 2018, 5, 86–90. [Google Scholar] [CrossRef]
- El-Gamal, M.I.; Brahim, I.; Hisham, N.; Aladdin, R.; Mohammed, H.; Bahaaeldin, A. Recent updates of carbapenem antibiotics. Eur. J. Med. Chem. 2017, 131, 185–195. [Google Scholar] [CrossRef]
- Maveyraud, L.; Mourey, L.; Kotra, L.P.; Pedelacq, J.-D.; Guillet, V.; Mobashery, S.; Samama, J.-P. Structural Basis for Clinical Longevity of Carbapenem Antibiotics in the Face of Challenge by the Common Class A β-Lactamases from the Antibiotic-Resistant Bacteria. J. Am. Chem. Soc. 1998, 120, 9748–9752. [Google Scholar] [CrossRef]
- Zhanel, G.G.; Johanson, C.; Embil, J.M.; Noreddin, A.; Gin, A.; Vercaigne, L.; Hoban, D.J. Ertapenem: Review of a new carbapenem. Expert Rev. Anti-Infect. Ther. 2005, 3, 23–39. [Google Scholar] [CrossRef]
- Bassetti, M.; Nicolini, L.; Esposito, S.; Righi, E.; Viscoli, C. Current Status of Newer Carbapenems. Curr. Med. Chem. 2009, 16, 564–575. [Google Scholar] [CrossRef]
- Papp-Wallace, K.M.; Endimiani, A.; Taracila, M.A.; Bonomo, R.A. Carbapenems: Past, present, and future. Antimicrob. Agents Chemother. 2011, 55, 4943–4960. [Google Scholar] [CrossRef]
- Livermore, D.M.; Sefton, A.M.; Scott, G.M. Properties and potential of ertapenem. J. Antimicrob. Chemother. 2003, 52, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Kattan, J.; Villegas, M.; Quinn, J. New developments in carbapenems. Clin. Microbiol. Infect. 2008, 14, 1102–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Top, J.; Willems, R.; Bonten, M. Emergence of CC17 Enterococcus faecium: From commensal to hospital-adapted pathogen. FEMS Immunol. Med Microbiol. 2008, 52, 297–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicodemo, A.; Paez, J. Antimicrobial therapy for Stenotrophomonas maltophilia infections. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Meletis, G. Carbapenem resistance: Overview of the problem and future perspectives. Ther. Adv. Infect. Dis. 2015, 3, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Granowitz, E.V.; Brown, R.B. Antibiotic Adverse Reactions and Drug Interactions. Crit. Care Clin. 2008, 24, 421–442. [Google Scholar] [CrossRef]
- Hashizume, T.; Ishino, F.; Nakagawa, J.-I.; Tamaki, S.; Matsuhashi, M. Studies on the mechanism of action of imipenem (N-formimidoylthienamycin) in vitro: Binding to the penicillin-binding proteins (PBPs) in Escherichia coli and Pseudomonas aeruginosa, and inhibition of enzyme activities due to the PBPs in E. coli. J. Antibiot. 1984, 37, 394–400. [Google Scholar] [CrossRef] [Green Version]
- Bonfiglio, G.; Russo, G.; Nicoletti, G. Recent developments in carbapenems. Expert Opin. Investig. Drugs 2002, 11, 529–544. [Google Scholar]
- Clark, R.B. Antibiotic susceptibilities of the Vibrionaceae to meropenem and other antimicrobial agents. Diagn. Microbiol. Infect. Dis. 1992, 15, 453–455. [Google Scholar] [CrossRef]
- Wang, H.; Chen, M.; Ni, Y.; Liu, Y.; Sun, H.; Yu, Y.; Yu, X.; Mei, Y.; Liu, M.; Sun, Z.; et al. Antimicrobial resistance among clinical isolates from the Chinese Meropenem Surveillance Study (CMSS), 2003–2008. Int. J. Antimicrob. Agents 2010, 35, 227–234. [Google Scholar] [CrossRef]
- Urban, C.; Mariano, N.; Rahal, J.J. In vitro double and triple bactericidal activities of doripenem, polymyxin B, and rifampin against multidrug-resistant Acinetobacter baumannii, Pseudomonas aeruginosa, Klebsiella pneumoniae, and Escherichia coli. Antimicrob. Agents Chemother. 2010, 54, 2732–2734. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, K.; Ip, M.; Ker, H.; Mendoza, M.; Hsu, L.; Kiratisin, P.; Chongthaleong, A.; Redjeki, I.; Quintana, A.; Flamm, R.; et al. In vitro activity of doripenem and other carbapenems against contemporary Gram-negative pathogens isolated from hospitalised patients in the Asia-Pacific region: Results of the COMPACT Asia-Pacific Study. Int. J. Antimicrob. Agents 2010, 36, 501–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, S.E.; Miller, L.; Needleman, J.; Uslan, D.Z.; Bell, D.; Watson, K.; Humphries, R.; McKinnell, J.A. Risk Factors for Development of Carbapenem Resistance Among Gram-Negative Rods. Open Forum Infect. Dis. 2019, 6, ofz027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doi, Y. Treatment Options for Carbapenem-resistant Gram-negative Bacterial Infections. Clin. Infect. Dis. 2019, 69 (Suppl. S7), S565–S575. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, G.; Pazhani, G.P.; Sarkar, A.; Rajendran, K.; Mukhopadhyay, A.K.; Bhattacharya, M.K.; Ghosh, A.; Ramamurthy, T. Carbapenem Resistance in Clonally Distinct Clinical Strains of Vibrio fluvialis Isolated from Diarrheal Samples. Emerg. Infect. Dis. 2016, 22, 1754–1761. [Google Scholar] [CrossRef] [Green Version]
- Woodford, N.; Wareham, D.W.; Guerra, B.; Teale, C. Carbapenemase-producing Enterobacteriaceae and non-Enterobacteriaceae from animals and the environment: An emerging public health risk of our own making? J. Antimicrob. Chemother. 2014, 69, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Verma, J.; Bag, S.; Saha, B.; Kumar, P.; Ghosh, T.S.; Dayal, M.; Senapati, T.; Mehra, S.; Dey, P.; Desigamani, A.; et al. Genomic plasticity associated with antimicrobial resistance in Vibrio cholerae. Proc. Natl. Acad. Sci. USA 2019, 116, 6226–6231. [Google Scholar] [CrossRef] [Green Version]
- Oyelade, A.A.; Adelowo, O.O.; Fagade, O.E. blaNDM-1-producing Vibrio parahaemolyticus and V. vulnificus isolated from recreational beaches in Lagos, Nigeria. Environ. Sci. Pollut. Res. 2018, 25, 33538–33547. [Google Scholar] [CrossRef]
- Le Thanh Dong, H.V.E.; Espinoza, J.L. Emerging superbugs: The threat of carbapenem resistant entero bacteriaceae. AIMS Microbiol. 2020, 6, 176. [Google Scholar] [CrossRef]
- Lee, L.-H.; Raghunath, P. Vibrionaceae Diversity, Multidrug Resistance and Management. Front. Microbiol. 2018, 9, 563. [Google Scholar] [CrossRef] [Green Version]
- Thompson, F.; Gomez-Gil, B. International Committee on Systematics of Prokaryotes Subcommittee on the taxonomy of Aeromonadaceae, Vibrionaceae and related organisms Minutes of the meeting, 13 November 2017, Chicago, USA. Int. J. Syst. Evol. Microbiol. 2018, 68, 2111–2112. [Google Scholar] [CrossRef] [PubMed]
- Roig, F.J.; González-Candelas, F.; Sanjuán, E.; Fouz, B.; Feil, E.J.; Llorens, C.; Baker-Austin, C.; Oliver, J.D.; Danin-Poleg, Y.; Gibas, C.J.; et al. Phylogeny of Vibrio vulnificus from the Analysis of the Core-Genome: Implications for Intra-Species Taxonomy. Front. Microbiol. 2018, 8, 2613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawabe, T.; Ogura, Y.; Matsumura, Y.; Feng, G.; Amin, A.R.; Mino, S.; Nakagawa, S.; Sawabe, T.; Kumar, R.; Fukui, Y.; et al. Updating the Vibrio clades defined by multilocus sequence phylogeny: Proposal of eight new clades, and the description of Vibrio tritonius sp. nov. Front. Microbiol. 2013, 4, 414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirk, S.; Huhulescu, S.; Allerberger, F.; Lepuschitz, S.; Rehak, S.; Weil, S.; Gschwandtner, E.; Hermann, M.; Neuhold, S.; Zoufaly, A.; et al. Necrotizing fasciitis due to Vibrio cholerae non-O1/non-O139 after exposure to Austrian bathing sites. Wien. Klin. Wochenschr. 2016, 128, 141–145. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, G.; Joshi, S.; Bhattacharya, S.; Sekar, U.; Birajdar, B.; Bhattacharyya, A.; Shinoda, S.; Ramamurthy, T. Extraintestinal infections caused by non-toxigenic Vibrio cholerae non-O1/non-O139. Front. Microbiol. 2016, 7, 144. [Google Scholar] [CrossRef] [Green Version]
- Petsaris, O.; Nousbaum, J.B.; Quilici, M.L.; Le Coadou, G.; Payan, C.; Abalain, M.L. Non-O1, non-O139 Vibrio cholerae bacteraemia in a cirrhotic patient. J. Med. Microbiol. 2010, 59, 1260–1262. [Google Scholar] [CrossRef] [Green Version]
- Tobin-D’Angelo, M.; Smith, A.R.; Bulens, S.N.; Thomas, S.; Hodel, M.; Izumiya, H.; Arakawa, E.; Morita, M.; Watanabe, H.; Marin, C.; et al. Severe Diarrhea Caused by Cholera Toxin–Producing Vibrio cholerae Serogroup O75 Infections Acquired in the Southeastern United States. Clin. Infect. Dis. 2008, 47, 1035–1040. [Google Scholar] [CrossRef] [Green Version]
- Valáriková, J.; Korcová, J.; Ziburová, J.; Rosinský, J.; Čížová, A.; Bieliková, S.; Sojka, M.; Farkaš, P. Potential pathogenicity and antibiotic resistance of aquatic Vibrio isolates from freshwater in Slovakia. Folia Microbiol. 2020, 65, 545–555. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio spp. infections. Nat. Rev. Dis. Primers 2018, 4, 1–19. [Google Scholar] [CrossRef]
- Heng, S.-P.; Letchumanan, V.; Deng, C.-Y.; Ab Mutalib, N.-S.; Khan, T.M.; Chuah, L.-H.; Chan, K.-G.; Goh, B.-H.; Pusparajah, P.; Lee, L.-H. Vibrio vulnificus: An Environmental and Clinical Burden. Front. Microbiol. 2017, 8, 997. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.-J.; Hsu, P.-H.; Lin, H.-T.V. A Novel Cooperative Metallo-β-Lactamase Fold Metallohydrolase from Pathogen Vibrio vulnificus Exhibits β-Lactam Antibiotic-Degrading Activities. Antimicrob. Agents Chemother. 2021, 65, AAC0032621. [Google Scholar] [CrossRef] [PubMed]
- Briet, A.; Helsens, N.; Delannoy, S.; Debuiche, S.; Brisabois, A.; Midelet, G.; Granier, S.A. NDM-1-producing Vibrio parahaemolyticus isolated from imported seafood. J. Antimicrob. Chemother. 2018, 73, 2578–2579. [Google Scholar] [CrossRef] [PubMed]
- Sadat, A.; El-Sherbiny, H.; Zakaria, A.; Ramadan, H.; Awad, A. Prevalence, antibiogram and virulence characterization of Vibrio isolates from fish and shellfish in Egypt: A possible zoonotic hazard to humans. J. Appl. Microbiol. 2021, 131, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Ramamurthy, T.; Chowdhury, G.; Pazhani, G.P.; Shinoda, S. Vibrio fluvialis: An emerging human pathogen. Front. Microbiol. 2014, 5, 91. [Google Scholar] [CrossRef] [Green Version]
- Kokashvili, T.; Whitehouse, C.A.; Tskhvediani, A.; Grim, C.J.; Elbakidze, T.; Mitaishvili, N.; Janelidze, N.; Jaiani, E.; Haley, B.J.; Lashkhi, N.; et al. Occurrence and Diversity of Clinically Important Vibrio Species in the Aquatic Environment of Georgia. Front. Public Health 2015, 3, 232. [Google Scholar] [CrossRef]
- Chowdhury, G.; Pazhani, G.P.; Dutta, D.; Guin, S.; Dutta, S.; Ghosh, S.; Izumiya, H.; Asakura, M.; Yamasaki, S.; Takeda, Y.; et al. Vibrio fluvialis in patients with diarrhea, Kolkata, India. Emerg. Infect. Dis. 2012, 18, 1868. [Google Scholar] [CrossRef] [Green Version]
- Ina-Salwany, M.Y.; Al-Saari, N.; Mohamad, A.; Mursidi, F.A.; Mohd-Aris, A.; Amal, M.N.A.; Kasai, H.; Mino, S.; Sawabe, T.; Zamri-Saad, M. Vibriosis in Fish: A Review on Disease Development and Prevention. J. Aquat. Anim. Health 2019, 31, 3–22. [Google Scholar] [CrossRef]
- Liu, L.; Xiao, J.; Zhang, M.; Zhu, W.; Xia, X.; Dai, X.; Pan, Y.; Yan, S.; Wang, Y. A Vibrio owensii strain as the causative agent of AHPND in cultured shrimp, Litopenaeus vannamei. J. Invertebr. Pathol. 2018, 153, 156–164. [Google Scholar] [CrossRef]
- Costa, R.A.; Colares, L.P.; Lima, R.A.; Fernandes Vieira, R.H.S.D.; de Sousa, O.V. Effect of Seawater on the Activity of Antibiotics AgainstVibriosIsolated from the Hemolymph of Cultured Pacific White Shrimp. J. World Aquac. Soc. 2012, 43, 727–732. [Google Scholar] [CrossRef]
- Dahanayake, P.; Hossain, S.; Wickramanayake, M.; Heo, G.J. Prevalence of virulence and extended-spectrum β-lactamase (ESBL) genes harbouring Vibrio spp. isolated from cockles (Tegillarca granosa) marketed in Korea. Lett. Appl. Microbiol. 2020, 71, 61–69. [Google Scholar] [CrossRef]
- Håkonsholm, F.; Lunestad, B.T.; Aguirre Sánchez, J.R.; Martinez-Urtaza, J.; Marathe, N.P.; Svanevik, C.S. Vibrios from the Norwegian marine environment: Characterization of associated antibiotic resistance and virulence genes. MicrobiologyOpen 2020, 9, e1093. [Google Scholar] [CrossRef]
- Ng, C.; Goh, S.G.; Saeidi, N.; Gerhard, W.A.; Gunsch, C.K.; Gin, K.Y.H. Occurrence of Vibrio species, beta-lactam resistant Vibrio species, and indicator bacteria in ballast and port waters of a tropical harbor. Sci. Total Environ. 2018, 610, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Zanetti, S.; Spanu, T.; Deriu, A.; Romano, L.; Sechi, L.A.; Fadda, G. In vitro susceptibility of Vibrio spp. isolated from the environment. Int. J. Antimicrob. Agents 2001, 17, 407–409. [Google Scholar] [CrossRef]
- Garrity, G.M.; Bell, J.A.; Lilburn, T.G. Taxonomic outline of the prokaryotes. In Bergey’s Manual of Systematic Bacteriology; Springer: New York, NY, USA, 2004. [Google Scholar]
- Turner, J.W.; Good, B.; Cole, D.; Lipp, E.K. Plankton composition and environmental factors contribute to Vibrio seasonality. ISME J. 2009, 3, 1082–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker-Austin, C.; Trinanes, J.A.; Salmenlinna, S.; Löfdahl, M.; Siitonen, A.; Taylor, N.G.; Martinez-Urtaza, J. Heat wave–associated vibriosis, Sweden and Finland, 2014. Emerg. Infect. Dis. 2016, 22, 1216. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, D.; Hasan, N.A.; Huq, A.; Colwell, R.R. Distribution and dynamics of epidemic and pandemic Vibrio parahaemolyticus virulence factors. Front. Cell. Infect. Microbiol. 2013, 3, 97. [Google Scholar] [CrossRef] [Green Version]
- Mansergh, S.; Zehr, J. Vibrio diversity and dynamics in the Monterey Bay upwelling region. Front. Microbiol. 2014, 5, 48. [Google Scholar] [CrossRef]
- Escudero, J.A.; Mazel, D. Genomic Plasticity of Vibrio cholerae. Int. Microbiol. 2017, 20, 138–148. [Google Scholar]
- Das, B.; Verma, J.; Kumar, P.; Ghosh, A.; Ramamurthy, T. Antibiotic resistance in Vibrio cholerae: Understanding the ecology of resistance genes and mechanisms. Vaccine 2020, 38, A83–A92. [Google Scholar] [CrossRef]
- Li, Y.; Wood, T.K.; Zhang, W.; Li, C. Vibrio splendidus persister cells induced by host coelomic fluids show a similar phenotype to antibiotic-induced counterparts. Environ. Microbiol. 2021, 23, 5605–5620. [Google Scholar] [CrossRef]
- Kirn, T.J.; Jude, B.A.; Taylor, R.K. A colonization factor links Vibrio cholerae environmental survival and human infection. Nature 2005, 438, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Meibom, K.L.; Blokesch, M.; Dolganov, N.A.; Wu, C.-Y.; Schoolnik, G.K. Chitin Induces Natural Competence in Vibrio cholerae. Science 2005, 310, 1824–1827. [Google Scholar] [CrossRef] [PubMed]
- Cooper, V.S.; Vohr, S.H.; Wrocklage, S.C.; Hatcher, P.J. Why Genes Evolve Faster on Secondary Chromosomes in Bacteria. PLOS Comput. Biol. 2010, 6, e1000732. [Google Scholar] [CrossRef] [PubMed]
- Dryselius, R.; Kurokawa, K.; Iida, T. Vibrionaceae, a versatile bacterial family with evolutionarily conserved variability. Res. Microbiol. 2007, 158, 479–486. [Google Scholar] [CrossRef] [PubMed]
- San Millan, A.; Escudero, J.A.; Gifford, D.R.; Mazel, D.; MacLean, R.C. Multicopy plasmids potentiate the evolution of antibiotic resistance in bacteria. Nat. Ecol. Evol. 2016, 1, 10. [Google Scholar] [CrossRef] [PubMed]
- Lutz, C.; Erken, M.; Noorian, P.; Sun, S.; McDougald, D. Environmental reservoirs and mechanisms of persistence of Vibrio cholerae. Front. Microbiol. 2013, 4, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.S.; Zaman, M.; Islam, M.S.; Ahmed, N.; Clemens, J. Environmental reservoirs of Vibrio cholerae. Vaccine 2020, 38, A52–A62. [Google Scholar] [CrossRef]
- Abd, H.; Saeed, A.; Weintraub, A.; Nair, G.B.; Sandström, G. Vibrio cholerae O1 strains are facultative intracellular bacteria, able to survive and multiply symbiotically inside the aquatic free-living amoeba Acanthamoeba castellanii. FEMS Microbiol. Ecol. 2007, 60, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Bankapalli, L.K.; Mishra, R.C.; Raychaudhuri, S. VopE, a Vibrio cholerae Type III Effector, Attenuates the Activation of CWI-MAPK Pathway in Yeast Model System. Front. Cell. Infect. Microbiol. 2017, 7, 82. [Google Scholar] [CrossRef] [Green Version]
- Blow, N.S.; Salomon, R.N.; Garrity, K.; Reveillaud, I.; Kopin, A.; Jackson, F.R.; Watnick, P.I. Vibrio cholerae Infection of Drosophila melanogaster Mimics the Human Disease Cholera. PLOS Pathog. 2005, 1, e8. [Google Scholar] [CrossRef] [Green Version]
- Hartwick, M.A.; Berenson, A.; Whistler, C.A.; Naumova, E.N.; Jones, S.H. The Seasonal Microbial Ecology of Plankton and Plankton-Associated Vibrio parahaemolyticus in the Northeast United States. Appl. Environ. Microbiol. 2021, 87, e02973-20. [Google Scholar] [CrossRef] [PubMed]
- SreeKalpana, M.; Suresh, P.; Kumar, G.R.; Kulkarni, S. Concurrent outbreak of Cholera and Shigella in Ronihal village, Bijapur, Karnataka, India, 2016. Int. J. Infect. Dis. 2020, 101, 270. [Google Scholar] [CrossRef]
- D’Souza, C.; Kumar, B.K.; Kapinakadu, S.; Shetty, R.; Karunasagar, I.; Karunasagar, I. PCR-based evidence showing the presence of Vibrio vulnificus in wound infection cases in Mangaluru, India. Int. J. Infect. Dis. 2018, 68, 74–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.-T.V.; Massam-Wu, T.; Lin, C.-P.; Wang, Y.-J.A.; Shen, Y.-C.; Lu, W.-J.; Hsu, P.-H.; Chen, Y.-H.; Borges-Walmsley, M.I.; Walmsley, A.R. The Vibrio cholerae var regulon encodes a metallo-β-lactamase and an antibiotic efflux pump, which are regulated by VarR, a LysR-type transcription factor. PLoS ONE 2017, 12, e0184255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letchumanan, V.; Chan, K.G.; Lee, L.-H. Vibrio parahaemolyticus: A review on the pathogenesis, prevalence, and advance molecular identification techniques. Front. Microbiol. 2014, 5, 705. [Google Scholar] [CrossRef] [Green Version]
- Choopun, N.; Louis, V.; Huq, A.; Colwell, R.R. Simple Procedure for Rapid Identification of Vibrio cholerae from the Aquatic Environment. Appl. Environ. Microbiol. 2002, 68, 995–998. [Google Scholar] [CrossRef] [Green Version]
- Kumar, B.K.; Raghunath, P.; Devegowda, D.; Deekshit, V.K.; Venugopal, M.N.; Karunasagar, I.; Karunasagar, I. Development of monoclonal antibody based sandwich ELISA for the rapid detection of pathogenic Vibrio parahaemolyticus in seafood. Int. J. Food Microbiol. 2011, 145, 244–249. [Google Scholar] [CrossRef]
- Nandi, B.; Nandy, R.K.; Mukhopadhyay, S.; Nair, G.B.; Shimada, T.; Ghose, A.C. Rapid Method for Species-Specific Identification of Vibrio cholerae Using Primers Targeted to the Gene of Outer Membrane Protein OmpW. J. Clin. Microbiol. 2000, 38, 4145–4151. [Google Scholar] [CrossRef] [Green Version]
- Federici, S.; Serrazanetti, D.I.; Guerzoni, M.E.; Campana, R.; Ciandrini, E.; Baffone, W.; Gianotti, A. Development of a rapid PCR protocol to detect Vibrio parahaemolyticus in clams. J. Food Sci. Technol. 2017, 55, 749–759. [Google Scholar] [CrossRef]
- Dieckmann, R.; Strauch, E.; Alter, T. Rapid identification and characterization of Vibrio species using whole-cell MALDI-TOF mass spectrometry. J. Appl. Microbiol. 2010, 109, 199–211. [Google Scholar] [CrossRef]
- Loo, K.-Y.; Law, J.W.-F.; Tan, L.T.-H.; Pusparajah, P.; Letchumanan, V.; Lee, L.-H. Diagnostic techniques for rapid detection of Vibrio species. Aquaculture 2022, 561, 738628. [Google Scholar] [CrossRef]
- Lepuschitz, S.; Baron, S.; Larvor, E.; Granier, S.A.; Pretzer, C.; Mach, R.L.; Farnleitner, A.H.; Ruppitsch, W.; Pleininger, S.; Indra, A.; et al. Phenotypic and genotypic antimicrobial resistance traits of Vibrio cholerae non-O1/non-O139 isolated from a large Austrian lake frequently associated with cases of human infection. Front. Microbiol. 2019, 10, 2600. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Kaushik, A.; Kumar, D.; Bag, S. Foodborne Pathogenic Vibrios: Antimicrobial Resistance. Front. Microbiol. 2021, 12, 638331. [Google Scholar] [CrossRef] [PubMed]
- Elmahdi, S.; DaSilva, L.V.; Parveen, S. Antibiotic resistance of Vibrio parahaemolyticus and Vibrio vulnificus in various countries: A review. Food Microbiol. 2016, 57, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.; Pires, S.M.; Ellis-Iversen, J.; Tang, Z.; Zhang, X.; Liu, J.; Li, W.; Cui, Q.; Zou, J.; Fu, P.; et al. Prevalence of Antimicrobial Resistant of Vibrio parahaemolyticus Isolated from Diarrheal Patients—Six PLADs, China, 2016−2020. China CDC Wkly. 2021, 3, 615. [Google Scholar] [CrossRef] [PubMed]
- Sneha, K.; Anas, A.; Jayalakshmy, K.; Jasmin, C.; Das, P.V.; Pai, S.S.; Pappu, S.; Nair, M.; Muraleedharan, K.; Sudheesh, K.; et al. Distribution of multiple antibiotic resistant Vibrio spp. across Palk Bay. Reg. Stud. Mar. Sci. 2016, 3, 242–250. [Google Scholar] [CrossRef]
- Tran, H.D.; Alam, M.; Trung, N.V.; Van Kinh, N.; Nguyen, H.H.; Pham, V.C.; Ansaruzzaman, M.; Rashed, S.M.; Bhuiyan, N.A.; Dao, T.T.; et al. Multi-drug resistant Vibrio cholerae O1 variant El Tor isolated in northern Vietnam between 2007 and 2010. J. Med. Microbiol. 2012, 61, 431–437. [Google Scholar] [CrossRef]
- Udoekong, N.S.; Bassey, B.E.; Asuquo, A.E.; Akan, O.D.; Ifeanyi, C.I.C. Prevalence and Antimicrobial Resistance of Gram-Negative Bacteria Isolates in Shellfish Samples from Two River Estuaries in South-South Nigeria. Adv. Microbiol. 2021, 11, 428–443. [Google Scholar] [CrossRef]
- Sperling, L.; Alter, T.; Huehn, S. Prevalence and Antimicrobial Resistance of Vibrio spp. in Retail and Farm Shrimps in Ecuador. J. Food Prot. 2015, 78, 2089–2092. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.W.; Rukayadi, Y.; Hasan, H.; Thung, T.Y.; Lee, E.; Rollon, W.D.; Hara, H.; Kayali, A.Y.; Nishibuchi, M.; Radu, S. Prevalence and antibiotic resistance patterns of Vibrio parahaemolyticus isolated from different types of seafood in Selangor, Malaysia. Saudi J. Biol. Sci. 2020, 27, 1602–1608. [Google Scholar] [CrossRef]
- Loo, K.Y.; Letchumanan, V.; Law, J.W.F.; Pusparajah, P.; Goh, B.H.; Ab Mutalib, N.S.; He, Y.W.; Lee, L.H. Incidence of antibiotic resistance in Vibrio spp. Rev. Aquac. 2020, 12, 2590–2608. [Google Scholar] [CrossRef]
- Karthikeyan, K.; Thirunarayan, M.A.; Krishnan, P. Coexistence of blaOXA-23 with blaNDM-1 and armA in clinical isolates of Acinetobacter baumannii from India. J. Antimicrob. Chemother. 2010, 65, 2253–2254. [Google Scholar] [CrossRef]
- Bonnin, R.; Poirel, L.; Naas, T.; Pirs, M.; Seme, K.; Schrenzel, J.; Nordmann, P. Dissemination of New Delhi metallo-β-lactamase-1-producing Acinetobacter baumannii in Europe. Clin. Microbiol. Infect. 2012, 18, E362–E365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamal, W.Y.; Albert, M.J.; Rotimi, V.O. High Prevalence of New Delhi Metallo-β-Lactamase-1 (NDM-1) Producers among Carbapenem-Resistant Enterobacteriaceae in Kuwait. PLoS ONE 2016, 11, e0152638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guh, A.Y.; Limbago, B.M.; Kallen, A.J. Epidemiology and prevention of carbapenem-resistant Enterobacteriaceae in the United States. Expert Rev. Anti Infect. Ther. 2014, 12, 565–580. [Google Scholar] [CrossRef]
- Munoz-Price, L.S.; Poirel, L.; Bonomo, R.A.; Schwaber, M.J.; Daikos, G.L.; Cormican, M.; Cornaglia, G.; Garau, J.; Gniadkowski, M.; Hayden, M.K.; et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect. Dis. 2013, 13, 785–796. [Google Scholar] [CrossRef] [Green Version]
- Hammerl, J.A.; Jäckel, C.; Bortolaia, V.; Schwartz, K.; Bier, N.; Hendriksen, R.S.; Guerra, B.; Strauch, E. Carbapenemase VCC-1–producing Vibrio cholerae in coastal waters of Germany. Emerg. Infect. Dis. 2017, 23, 1735. [Google Scholar] [CrossRef]
- Gxalo, O.; Digban, T.O.; Igere, B.E.; Olapade, O.A.; Okoh, A.I.; Nwodo, U.U. Virulence and Antibiotic Resistance Characteristics of Vibrio Isolates from Rustic Environmental Freshwaters. Front. Cell. Infect. Microbiol. 2021, 11, 765. [Google Scholar] [CrossRef]
- Igbinosa, E.O.; Beshiru, A.; Igbinosa, I.H.; Ogofure, A.G.; Uwhuba, K.E. Prevalence and Characterization of Food-Borne Vibrio parahaemolyticus from African Salad in Southern Nigeria. Front. Microbiol. 2021, 12, 632266. [Google Scholar] [CrossRef]
- Hossain, S.; Wickramanayake, M.V.K.S.; Dahanayake, P.S.; Heo, G.-J. Occurrence of virulence and extended-spectrum β-lactamase determinants in Vibrio spp. isolated from marketed hard-shelled mussel (Mytilus coruscus). Microb. Drug Resist. 2020, 26, 391–401. [Google Scholar] [CrossRef]
- Jun, J.W.; Kim, J.H.; Choresca, C.H., Jr.; Shin, S.P.; Han, J.E.; Han, S.Y.; Chai, J.Y.; Park, S.C. Isolation, molecular characterization, and antibiotic susceptibility of Vibrio parahaemolyticus in Korean seafood. Foodborne Pathog. Dis. 2012, 9, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Zheng, Z.; Ye, L.; Chen, S. Identification of a Novel Metallo-β-Lactamase, VAM-1, in a Foodborne Vibrio alginolyticus Isolate from China. Antimicrob. Agents Chemother. 2021, 65, e01129-21. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.A.; Westman, E.L.; Wright, G.D. The antibiotic resistome: What’s new? Curr. Opin. Microbiol. 2014, 21, 45–50. [Google Scholar] [CrossRef]
- Martínez, J.L.; Coque, T.M.; Baquero, F. What is a resistance gene? Ranking risk in resistomes. Nat. Rev. Genet. 2014, 13, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, D.; Alam, M.; Huq, A.; Colwell, R.R. Reduced Susceptibility to Extended-Spectrum β-Lactams in Vibrio cholerae Isolated in Bangladesh. Front. Public Health 2016, 4, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yahyai, S. Optimization of a method for testing ballast water for enterococci and an investigation on the occurrence of antibiotic resistance in Vibrio cholerae. Master’s Thesis, University of Maryland, College Park, MD, USA, 2013. [Google Scholar]
- Gu, W.; Yin, J.; Yang, J.; Li, C.; Chen, Y.; Yin, J.; Xu, W.; Zhao, S.; Liang, J.; Jing, H.; et al. Characterization of Vibrio cholerae from 1986 to 2012 in Yunnan Province, southwest China bordering Myanmar. Infect. Genet. Evol. 2013, 21, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yong-Hong, X.; Jin, W.; Yan, Z.; Lei, G. 2008 Mohnarin report: Antibiotic resistance analysis of Neisseria gonorrhoeae, Gardnerella vaginalis, Aeromonas and Vibrio parahaemolyticus. Chin. J. Antibiot. 2010, 35, S5–S8. [Google Scholar]
- Chen, Y.; Chen, X.; Yu, F.; Wu, M.; Wang, R.; Zheng, S.; Han, D.; Yang, Q.; Kong, H.; Zhou, F.; et al. Serological, Virulence, Antimicrobial Susceptibility and Molecular Characteristics of Clinical Vibrio parahaemolyticus Strains Circulating in Southeastern China: 2009 to 2013. Clin. Microbiol. Infect. 2016, 22, 258.e9–258.e16. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Yu, T.; Yang, Y.; Yu, S.; Wu, J.; Lin, R.; Li, Y.; Fang, J.; Zhu, C. Co-occurrence of Antibiotic and Heavy Metal Resistance and Sequence Type Diversity of Vibrio parahaemolyticus Isolated from Penaeus vannamei at Freshwater Farms, Seawater Farms, and Markets in Zhejiang Province, China. Front. Microbiol. 2020, 11, 1294. [Google Scholar] [CrossRef]
- Zheng, Z.; Li, R.; Ye, L.; Chan, E.W.-C.; Chen, S. Identification and characterization of IncA/C conjugative, bla NDM-1-bearing plasmid in Vibrio alginolyticus of food origin. Antimicrob. Agents Chemother. 2018, 62, e01897-18. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Z.; Ye, L.; Chan, E.W.-C.; Chen, S. Identification and characterization of a conjugative blaVIM-1-bearing plasmid in Vibrio alginolyticus of food origin. J. Antimicrob. Chemother. 2019, 74, 1842–1847. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Luo, X.; Zhang, J.; Zhu, J. Characteristics of and germicidal efficacy at different temperatures against Vibrio parahaemolyticus. Chin. J. Microbiol. Immunol. 2019, 12, 527–531. [Google Scholar]
- Zheng, Z.; Cheng, Q.; Chan, E.W.C.; Chen, S. Genetic and biochemical characterization of VMB-1, a novel metallo-β-lactamase encoded by a conjugative, broad-host range IncC plasmid from Vibrio spp. Adv. Biosyst. 2020, 4, 1900221. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, W.; Peng, K.; Wang, Z.; Li, R. Identification of a Novel Plasmid-Mediated Carbapenemase-Encoding Gene, bla VMB-2, in Vibrio diabolicus. Antimicrob. Agents Chemother. 2021, 65, AAC0020621. [Google Scholar] [CrossRef]
- Walsh, T.R.; Weeks, J.; Livermore, D.M.; Toleman, M.A. Dissemination of NDM-1 positive bacteria in the New Delhi environment and its implications for human health: An environmental point prevalence study. Lancet Infect. Dis. 2011, 11, 355–362. [Google Scholar] [CrossRef]
- Mandal, J.; Sangeetha, V.; Ganesan, V.; Parveen, M.; Preethi, V.; Harish, B.N.; Srinivasan, S.; Parija, S.C. Third-Generation Cephalosporin-Resistant Vibrio cholerae, India. Emerg. Infect. Dis. 2012, 18, 1326–1328. [Google Scholar] [CrossRef]
- Silvester, R.; Pires, J.; Van Boeckel, T.P.; Madhavan, A.; Balakrishnan Meenakshikutti, A.; Hatha, M. Occurrence of β-lactam resistance genes and plasmid-mediated resistance among Vibrios isolated from Southwest Coast of India. Microb. Drug Resist. 2019, 25, 1306–1315. [Google Scholar] [CrossRef]
- Di, D.Y.; Shin, H.; Han, D.; Unno, T.; Hur, H.-G. High genetic diversity of Vibrio parahaemolyticus isolated from tidal water and mud of southern coast of South Korea. FEMS Microbiol. Ecol. 2019, 95, fiz022. [Google Scholar] [CrossRef]
- Kim, D.-H.; Rajapaksha, L.; Gunasekara, C.; Wimalasena, S.; Pathirana, H.; Kim, S.-R.; Seo, B.-J.; Heo, G.-J.; Shin, G.-W. Phylogenetic relationships and antibiotic resistance of Vibrio parahaemolyticus isolates related to acute hepatopancreatic necrosis disease in Korea. Aquaculture 2021, 545, 737253. [Google Scholar] [CrossRef]
- Noorlis, A.; Ghazali, F.; Cheah, Y.; Tuan Zainazor, T.; Wong, W.; Tunung, R.; Pui, C.; Nishibuchi, M.; Nakaguchi, Y.; Son, R. Antibiotic resistance and biosafety of Vibrio cholerae and Vibrio parahaemolyticus from freshwater ish at retail level. Int. Food Res. J. 2011, 18, 1523. [Google Scholar]
- Diep, T.T.; Nguyen, N.T.N.; Nguyen, T.N.C.; An, H.K.; Nguyen, T.Q.; Nguyen, V.H.; Nguyen, T.V.; Nguyen, T.N.A.; Izumiya, H.; Ohnishi, M.; et al. Isolation of New Delhi metallo-β-lactamase 1-producing Vibrio cholerae non-O1, non-O139 strain carrying ctxA, st and hly genes in southern Vietnam. Microbiol. Immunol. 2015, 59, 262–267. [Google Scholar] [CrossRef]
- McAuliffe, G.N.; Hennessy, J.; Baird, R.W. Relative frequency, characteristics, and antimicrobial susceptibility patterns of Vibrio spp., Aeromonas spp., Chromobacterium violaceum, and Shewanella spp. in the northern territory of Australia, 2000–2013. Am. J. Trop. Med. Hyg. 2015, 92, 605. [Google Scholar] [CrossRef] [PubMed]
- Igere, B.E.; Okoh, A.I.; Nwodo, U.U. Antibiotic Susceptibility Testing (AST) Reports: A Basis for Environmental/Epidemiological Surveillance and Infection Control Amongst Environmental Vibrio cholerae. Int. J. Environ. Res. Public Health 2020, 17, 5685. [Google Scholar] [CrossRef] [PubMed]
- Okoh, A.I.; Igbinosa, E.O. Antibiotic susceptibility profiles of some Vibrio strains isolated from wastewater final effluents in a rural community of the Eastern Cape Province of South Africa. BMC Microbiol. 2010, 10, 143. [Google Scholar] [CrossRef] [Green Version]
- Onohuean, H.; Okoh, A.I.; Nwodo, U.U. Antibiogram signatures of Vibrio species recovered from surface waters in South Western districts of Uganda: Implications for environmental pollution and infection control. Sci. Total Environ. 2022, 807, 150706. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, D.; Medici, L.; Talevi, G.; Napoleoni, M.; Serratore, P.; Zavatta, E.; Bignami, G.; Masini, L.; Chierichetti, S.; Fisichella, S.; et al. Molecular characterization and drug susceptibility of non-O1/O139 V. ácholerae strains of seafood, environmental and clinical origin, Italy. Food Microbiol. 2018, 72, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Zago, V.; Veschetti, L.; Patuzzo, C.; Malerba, G.; Lleo, M.M. Shewanella algae and Vibrio spp. strains isolated in Italian aquaculture farms are reservoirs of antibiotic resistant genes that might constitute a risk for human health. Mar. Pollut. Bull. 2020, 154, 111057. [Google Scholar] [CrossRef] [PubMed]
- Serratore, P.; Zavatta, E.; Fiocchi, E.; Serafini, E.; Serraino, A.; Giacometti, F.; Bignami, G. Preliminary study on the antimicrobial susceptibility pattern related to the genotype of Vibrio vulnificus strains isolated in the north-western Adriatic Sea coastal area. Ital. J. Food Saf. 2017, 6, 6843. [Google Scholar] [CrossRef] [Green Version]
- Aberkane, S.; Compain, F.; Barraud, O.; Ouédraogo, A.-S.; Bouzinbi, N.; Vittecoq, M.; Jean-Pierre, H.; Decré, D.; Godreuil, S. Non-O1/non-O139 Vibrio cholerae avian isolate from France cocarrying the bla VIM-1 and bla VIM-4 genes. Antimicrob. Agents Chemother. 2015, 59, 6594–6596. [Google Scholar] [CrossRef] [Green Version]
- Pariente Martín, M.; Escribano Garaizábal, E.; Liria Sánchez, P.J.; Crespo Sánchez, M.D. Vibrio metschnikovii from a human infected leg ulcer. Rev. Inst. Med. Trop. São Paulo 2008, 50, 311–312. [Google Scholar] [CrossRef] [Green Version]
- Darley, E.; Weeks, J.; Jones, L.; Daniels, V.; Wootton, M.; MacGowan, A.; Walsh, T. NDM-1 polymicrobial infections including Vibrio cholerae. Lancet 2012, 380, 1358. [Google Scholar] [CrossRef]
- Shaw, K.S.; Rosenberg Goldstein, R.E.; He, X.; Jacobs, J.M.; Crump, B.C.; Sapkota, A.R. Antimicrobial Susceptibility of Vibrio vulnificus and Vibrio parahaemolyticus Recovered from Recreational and Commercial Areas of Chesapeake Bay and Maryland Coastal Bays. PLoS ONE 2014, 9, e89616. [Google Scholar] [CrossRef] [PubMed]
- Cherak, Z.; Loucif, L.; Moussi, A.; Rolain, J.-M. Carbapenemase-producing Gram-negative bacteria in aquatic environments: A review. J. Glob. Antimicrob. Resist. 2021, 25, 287–309. [Google Scholar] [CrossRef] [PubMed]
- De, R. Mobile Genetic Elements of Vibrio cholerae and the Evolution of Its Antimicrobial Resistance. Front. Trop. Dis. 2021, 2, 7. [Google Scholar] [CrossRef]
- Cross, T.; Ransegnola, B.; Shin, J.-H.; Weaver, A.; Fauntleroy, K.; VanNieuwenhze, M.S.; Westblade, L.F.; Dörr, T. Spheroplast-Mediated Carbapenem Tolerance in Gram-Negative Pathogens. Antimicrob. Agents Chemother. 2019, 63, e00756-19. [Google Scholar] [CrossRef]
- Morita, Y.; Tomida, J.; Kawamura, Y. MexXY multidrug efflux system of Pseudomonas aeruginosa. Front. Microbiol. 2012, 3, 408. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, K.; Gotoh, N.; Nishino, T. Alterations of susceptibility of Pseudomonas aeruginosa by overproduction of multidrug efflux systems, MexAB-OprM, MexCD-OprJ, and MexXY/OprM to carbapenems: Substrate specificities of the efflux systems. J. Infect. Chemother. 2002, 8, 371–373. [Google Scholar] [CrossRef]
- Chetri, S.; Bhowmik, D.; Paul, D.; Pandey, P.; Chanda, D.D.; Chakravarty, A.; Bora, D.; Bhattacharjee, A. AcrAB-TolC efflux pump system plays a role in carbapenem non-susceptibility in Escherichia coli. BMC Microbiol. 2019, 19, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Atzori, A.; Malloci, G.; Cardamone, F.; Bosin, A.; Vargiu, A.V.; Ruggerone, P. Molecular Interactions of Carbapenem Antibiotics with the Multidrug Efflux Transporter AcrB of Escherichia coli. Int. J. Mol. Sci. 2020, 21, 860. [Google Scholar] [CrossRef] [Green Version]
- Zago, V.; Veschetti, L.; Patuzzo, C.; Malerba, G.; Lleo, M.M. Resistome, Mobilome and Virulome Analysis of Shewanella algae and Vibrio spp. Strains Isolated in Italian Aquaculture Centers. Microorganisms 2020, 8, 572. [Google Scholar] [CrossRef] [Green Version]
- Bonomo, R.A.; Burd, E.M.; Conly, J.; Limbago, B.M.; Poirel, L.; Segre, J.A.; Westblade, L.F. Carbapenemase-Producing Organisms: A Global Scourge. Clin. Infect. Dis. 2018, 66, 1290–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dortet, L.; Poirel, L.; Nordmann, P. Worldwide Dissemination of the NDM-Type Carbapenemases in Gram-Negative Bacteria. BioMed Res. Int. 2014, 2014, 249856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamy, B.; Laurent, F.; Kodjo, A.; Roger, F.; Jumas-Bilak, E.; Marchandin, H. Which antibiotics and breakpoints should be used for Aeromonas susceptibility testing? Considerations from a comparison of agar dilution and disk diffusion methods using Enterobacteriaceae breakpoints. Eur. J. Clin. Microbiol. 2012, 31, 2369–2377. [Google Scholar] [CrossRef] [PubMed]
- Jean, S.-S.; Lee, W.-S.; Lam, C.; Hsu, C.-W.; Chen, R.-J.; Hsueh, P.-R. Carbapenemase-producing Gram-negative bacteria: Current epidemics, antimicrobial susceptibility and treatment options. Futur. Microbiol. 2015, 10, 407–425. [Google Scholar] [CrossRef]
- Diene, S.M.; Rolain, J.M. Carbapenemase genes and genetic platforms in Gram-negative bacilli: Enterobacteriaceae, Pseudomonas and Acinetobacter species. Clin. Microbiol. Infect. 2014, 20, 831–838. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A. Updated Functional Classification of β-Lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Jones, L.S. Mobile genetic elements associated with blaNDM-1 in Acinetobacter spp. and Vibrio cholerae. Ph.D. Thesis, Cardiff University, Wales, UK, 2015. [Google Scholar]
- Tamma, P.D.; Simner, P.J. Phenotypic Detection of Carbapenemase-Producing Organisms from Clinical Isolates. J. Clin. Microbiol. 2018, 56, e01140-18. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Ganta, S.R.; Perumal, S.; Pagadala, S.R.R.; Samuelsen, Ø.; Spencer, J.; Pratt, R.; Buynak, J.D. Approaches to the simultaneous inactivation of metallo- and serine-β-lactamases. Bioorganic Med. Chem. Lett. 2009, 19, 1618–1622. [Google Scholar] [CrossRef] [Green Version]
- Bebrone, C. Metallo-β-lactamases (classification, activity, genetic organization, structure, zinc coordination) and their superfamily. Biochem. Pharmacol. 2007, 74, 1686–1701. [Google Scholar] [CrossRef]
- Dewi, D.A.P.R.; Thomas, T.; Ahmad Mokhtar, A.M.; Mat Nanyan, N.S.; Zulfigar, S.B.; Salikin, N.H. Carbapenem Resistance among Marine Bacteria—An Emerging Threat to the Global Health Sector. Microorganisms 2021, 9, 2147. [Google Scholar] [CrossRef] [PubMed]
- Garg, A.; Garg, J.; Kumar, S.; Bhattacharya, A.; Agarwal, S.; Upadhyay, G. Molecular epidemiology & therapeutic options of carbapenem-resistant Gram-negative bacteria. Indian J. Med Res. 2019, 149, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-β-lactamase gene, bla NDM-1, and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, A.; Tripathi, A. An in silico approach for understanding the molecular evolution of clinically important metallo-beta-lactamases. Infect. Genet. Evol. 2013, 20, 39–47. [Google Scholar] [CrossRef]
- Blokesch, M. A quorum sensing-mediated switch contributes to natural transformation of Vibrio cholerae. Mob. Genet. Elem. 2012, 2, 224–227. [Google Scholar] [CrossRef]
- Neiman, J.; Guo, Y.; Rowe-Magnus, D.A. Chitin-Induced Carbotype Conversion in Vibrio vulnificus. Infect. Immun. 2011, 79, 3195–3203. [Google Scholar] [CrossRef] [Green Version]
- Rajpara, N.; Nair, M.; Bhardwaj, A.K. A Highly Promiscuous Integron, Plasmids, Extended Spectrum Beta Lactamases and Efflux Pumps as Factors Governing Multidrug Resistance in a Highly Drug Resistant Vibrio fluvialis Isolate BD146 from Kolkata, India. Indian J. Microbiol. 2018, 58, 60–67. [Google Scholar] [CrossRef]
- MacLean, R.C.; San Millan, A. Microbial Evolution: Towards Resolving the Plasmid Paradox. Curr. Biol. 2015, 25, R764–R767. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A. Plasmids and the spread of resistance. Int. J. Med. Microbiol. 2013, 303, 298–304. [Google Scholar] [CrossRef]
- Letchumanan, V.; Chan, K.-G.; Lee, L.-H. An insight of traditional plasmid curing in Vibrio species. Front. Microbiol. 2015, 6, 735. [Google Scholar] [CrossRef]
- Johnson, A.P.; Woodford, N. Global spread of antibiotic resistance: The example of New Delhi metallo-β-lactamase (NDM)-mediated carbapenem resistance. J. Med. Microbiol. 2013, 62, 499–513. [Google Scholar] [CrossRef]
- Wang, R.; Yu, D.; Zhu, L.; Li, J.; Yue, J.; Kan, B. IncA/C plasmids harboured in serious multidrug-resistant Vibrio cholerae serogroup O139 strains in China. Int. J. Antimicrob. Agents 2015, 45, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.; Huguet, K.T.; Grenier, F.; Burrus, V. IncC conjugative plasmids and SXT/R391 elements repair double-strand breaks caused by CRISPR–Cas during conjugation. Nucleic Acids Res. 2020, 48, 8815–8827. [Google Scholar] [CrossRef] [PubMed]
- Magagnin, C.M.; Campos, J.C.; da Rocha, D.A.; Sampaio, S.C.; Rozáles, F.P.; Barth, A.L.; Zavascki, A.P.; Sampaio, J.L. Dissemination of blaOXA-370 is mediated by IncX plasmids and the Tn6435 transposon. Eur. J. Clin. Microbiol. 2018, 37, 2165–2169. [Google Scholar] [CrossRef] [PubMed]
- Cambray, G.; Guerout, A.-M.; Mazel, D. Integrons. Annu. Rev. Genet. 2010, 44, 141–166. [Google Scholar] [CrossRef]
- Hall, R.M. Mobile Gene Cassettes and Integrons: Moving Antibiotic Resistance Genes in Gram-Negative Bacteria. In Antibiotic Resistance: Origins, Evolution, Selection and Spread, Proceedings of the Ciba Foundation Symposium 207, Chichester, UK, 2007; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2007; pp. 192–205. [Google Scholar]
- Hall, R.M.; Collis, C.M. Mobile gene cassettes and integrons: Capture and spread of genes by site-specific recombination. Mol. Microbiol. 1995, 15, 593–600. [Google Scholar] [CrossRef]
- Gillings, M.; Boucher, Y.; Labbate, M.; Holmes, A.; Krishnan, S.; Holley, M.; Stokes, H.W. The Evolution of Class 1 Integrons and the Rise of Antibiotic Resistance. J. Bacteriol. 2008, 190, 5095–5100. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Fujihara, K.; Sato, T.; Ito, H.; Garg, P.; Chakrabarty, R.; Ramamurthy, T.; Nair, G.B.; Takeda, Y.; Yamasaki, S. Distribution and characterization of integrons in various serogroups of Vibrio cholerae strains isolated from diarrhoeal patients between 1992 and 2000 in Kolkata, India. J. Med. Microbiol. 2006, 55, 575–583. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L.; Walsh, T.R.; Livermore, D.M. The emerging NDM carbapenemases. Trends Microbiol. 2011, 19, 588–595. [Google Scholar] [CrossRef]
- Ceccarelli, D.; Salvia, A.M.; Sami, J.; Cappuccinelli, P.; Colombo, M.M. New Cluster of Plasmid-Located Class 1 Integrons in Vibrio cholerae O1 and a dfrA15 Cassette-Containing Integron in Vibrio parahaemolyticus Isolated in Angola. Antimicrob. Agents Chemother. 2006, 50, 2493–2499. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, É.L.; dos Santos Freitas, F.; Vieira, V.V.; Vicente, A.C. New qnr gene cassettes associated with superintegron repeats in Vibrio cholerae O1. Emerg. Infect. Dis. 2008, 14, 1129. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.K.; Knabel, S.J.; Kwan, B.W. Bacterial Persister Cell Formation and Dormancy. Appl. Environ. Microbiol. 2013, 79, 7116–7121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, K. Persister cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Silva-Valenzuela, C.A.; Lazinski, D.W.; Kahne, S.C.; Nguyen, Y.; Molina-Quiroz, R.C.; Camilli, A. Growth arrest and a persister state enable resistance to osmotic shock and facilitate dissemination of Vibrio cholerae. ISME J. 2017, 11, 2718–2728. [Google Scholar] [CrossRef] [Green Version]
- Ayrapetyan, M.; Williams, T.C.; Baxter, R.; Oliver, J.D. Viable but Nonculturable and Persister Cells Coexist Stochastically and Are Induced by Human Serum. Infect. Immun. 2015, 83, 4194–4203. [Google Scholar] [CrossRef]
- Singh, S.; Singh, S.K.; Chowdhury, I.; Singh, R. Understanding the mechanism of bacterial biofilms resistance to antimicrobial agents. Open Microbiol. J. 2017, 11, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abisado, R.G.; Benomar, S.; Klaus, J.R.; Dandekar, A.A.; Chandler, J.R. Bacterial quorum sensing and microbial community interactions. MBio 2018, 9, e02331-17. [Google Scholar] [CrossRef] [Green Version]
- Penesyan, A.; Gillings, M.; Paulsen, I.T. Antibiotic Discovery: Combatting Bacterial Resistance in Cells and in Biofilm Communities. Molecules 2015, 20, 5286–5298. [Google Scholar] [CrossRef] [Green Version]
- Penesyan, A.; Paulsen, I.T.; Gillings, M.R.; Kjelleberg, S.; Manefield, M.J. Secondary Effects of Antibiotics on Microbial Biofilms. Front. Microbiol. 2020, 11, 2109. [Google Scholar] [CrossRef]
- Murtha, A.N.; Kazi, M.I.; Schargel, R.D.; Cross, T.; Fihn, C.; Carlson, E.E.; Boll, J.M.; Dörr, T. The PhoPQ two-component system promotes carbapenem tolerance in Enterobacterales. bioRxiv 2021. [Google Scholar] [CrossRef]
- Cheng, A.T.; Ottemann, K.M.; Yildiz, F.H. Vibrio cholerae Response Regulator VxrB Controls Colonization and Regulates the Type VI Secretion System. PLOS Pathog. 2015, 11, e1004933. [Google Scholar] [CrossRef] [PubMed]
- Dörr, T.; Alvarez, L.; Delgado, F.; Davis, B.M.; Cava, F.; Waldor, M.K. A cell wall damage response mediated by a sensor kinase/response regulator pair enables beta-lactam tolerance. Proc. Natl. Acad. Sci. USA 2016, 113, 404–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, A.I.; Murphy, S.G.; Umans, B.D.; Tallavajhala, S.; Onyekwere, I.; Wittels, S.; Shin, J.-H.; VanNieuwenhze, M.; Waldor, M.K.; Dörr, T. Genetic Determinants of Penicillin Tolerance in Vibrio cholerae. Antimicrob. Agents Chemother. 2018, 62, e01326-18. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, E.; Daniel, S.; Hernández, S.B.; Goudin, A.; Cava, F.; Barre, F.-X.; Galli, E. l -Arabinose Induces the Formation of Viable Nonproliferating Spheroplasts in Vibrio cholerae. Appl. Environ. Microbiol. 2021, 87, e02305–e02320. [Google Scholar] [CrossRef]
- Monahan, L.G.; Turnbull, L.; Osvath, S.R.; Birch, D.; Charles, I.G.; Whitchurch, C.B. Rapid Conversion of Pseudomonas aeruginosa to a Spherical Cell Morphotype Facilitates Tolerance to Carbapenems and Penicillins but Increases Susceptibility to Antimicrobial Peptides. Antimicrob. Agents Chemother. 2014, 58, 1956–1962. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.; Higgs, E.; Rutman, A.; Cole, P. Isolation of spheroplastic forms of Haemophilus influenzae from sputum in conventionally treated chronic bronchial sepsis using selective medium supplemented with N-acetyl-D-glucosamine: Possible reservoir for re-emergence of infection. BMJ 1984, 289, 1409–1412. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, H.P.; Bugrysheva, J.; Sue, D. Optical microscopy reveals the dynamic nature of B. pseudomallei morphology during β-lactam antimicrobial susceptibility testing. BMC Microbiol. 2020, 20, 209. [Google Scholar] [CrossRef]
- Zou, J.; Kou, S.-H.; Xie, R.; VanNieuwenhze, M.S.; Qu, J.; Peng, B.; Zheng, J. Non-walled spherical Acinetobacter baumannii is an important type of persister upon β-lactam antibiotic treatment. Emerg. Microbes Infect. 2020, 9, 1149–1159. [Google Scholar] [CrossRef]
- Kopterides, P.; Koletsi, P.K.; Michalopoulos, A.; Falagas, M.E. Exposure to quinolones is associated with carbapenem resistance among colistin-susceptible Acinetobacter baumannii blood isolates. Int. J. Antimicrob. Agents 2007, 30, 409–414. [Google Scholar] [CrossRef]
- Kumar, P.; Bag, S.; Ghosh, T.S.; Dey, P.; Dayal, M.; Saha, B.; Verma, J.; Pant, A.; Saxena, S.; Desigamani, A.; et al. Molecular Insights into Antimicrobial Resistance Traits of Multidrug Resistant Enteric Pathogens isolated from India. Sci. Rep. 2017, 7, 14468. [Google Scholar] [CrossRef] [Green Version]
- Gelband, H.; Miller, P.; Molly; Pant, S.; Gandra, S.; Levinson, J.; Barter, D.; White, A.; Laxminarayan, R. The state of the world’s antibiotics 2015. Wound Health S. Afr. 2015, 8, 30–34. [Google Scholar]
- Kardos, N. CRE (Carbapenem Resistant Enterobacteriaceae) and the Globalization of Antimicrobial Resistance: Problems and Solutions. SunText Rev. Biotechnol. 2020, 1, 103. [Google Scholar]
- Lee, Y.-T.; Fung, C.-P.; Wang, F.-D.; Chen, C.-P.; Chen, T.-L.; Cho, W.-L. Outbreak of imipenem-resistant Acinetobacter calcoaceticus–Acinetobacter baumannii complex harboring different carbapenemase gene-associated genetic structures in an intensive care unit. J. Microbiol. Immunol. Infect. 2012, 45, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Al Salah, D.M.M.; Ngweme, G.N.; Laffite, A.; Otamonga, J.-P.; Mulaji, C.; Poté, J. Hospital wastewaters: A reservoir and source of clinically relevant bacteria and antibiotic resistant genes dissemination in urban river under tropical conditions. Ecotoxicol. Environ. Saf. 2020, 200, 110767. [Google Scholar] [CrossRef] [PubMed]
- Lamba, M.; Graham, D.W.; Ahammad, S.Z. Hospital Wastewater Releases of Carbapenem-Resistance Pathogens and Genes in Urban India. Environ. Sci. Technol. 2017, 51, 13906–13912. [Google Scholar] [CrossRef] [Green Version]
- Weingarten, R.A.; Johnson, R.C.; Conlan, S.; Ramsburg, A.M.; Dekker, J.P.; Lau, A.F.; Khil, P.; Odom, R.T.; Deming, C.; Park, M.; et al. Genomic Analysis of Hospital Plumbing Reveals Diverse Reservoir of Bacterial Plasmids Conferring Carbapenem Resistance. mBio 2018, 9, e02011–e02017. [Google Scholar] [CrossRef] [Green Version]
- Larsson, D.; Flach, C.-F. Antibiotic resistance in the environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef]
- Ranjan, R.; Thatikonda, S. β-Lactam Resistance Gene NDM-1 in the Aquatic Environment: A Review. Curr. Microbiol. 2021, 78, 3634–3643. [Google Scholar] [CrossRef]
- Theethakaew, C.; Nakamura, S.; Motooka, D.; Matsuda, S.; Kodama, T.; Chonsin, K.; Suthienkul, O.; Iida, T. Plasmid dynamics in Vibrio parahaemolyticus strains related to shrimp Acute Hepatopancreatic Necrosis Syndrome (AHPNS). Infect. Genet. Evol. 2017, 51, 211–218. [Google Scholar] [CrossRef]
- Kumar, R.; Ng, T.H.; Wang, H.-C. Acute hepatopancreatic necrosis disease in penaeid shrimp. Rev. Aquac. 2020, 12, 1867–1880. [Google Scholar] [CrossRef] [Green Version]
- Dolejska, M.; Papagiannitsis, C.C. Plasmid-mediated resistance is going wild. Plasmid 2018, 99, 99–111. [Google Scholar] [CrossRef]
- Jung, D.; Morrison, B.J.; Rubin, J.E. A review of antimicrobial resistance in imported foods. Can. J. Microbiol. 2022, 68, 1–15. [Google Scholar] [CrossRef]
- Randa, M.A.; Polz, M.F.; Lim, E. Effects of Temperature and Salinity on Vibrio vulnificus Population Dynamics as Assessed by Quantitative PCR. Appl. Environ. Microbiol. 2004, 70, 5469–5476. [Google Scholar] [CrossRef] [Green Version]
- Vezzulli, L.; Colwell, R.R.; Pruzzo, C. Ocean Warming and Spread of Pathogenic Vibrios in the Aquatic Environment. Microb. Ecol. 2013, 65, 817–825. [Google Scholar] [CrossRef]
- Lupo, A.; Coyne, S.; Berendonk, T.U. Origin and Evolution of Antibiotic Resistance: The Common Mechanisms of Emergence and Spread in Water Bodies. Front. Microbiol. 2012, 3, 18. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.R.; Stedtfeld, R.D.; Guo, X.; Hashsham, S.A. Antimicrobial Resistance in the Environment. Water Environ. Res. 2016, 88, 1951–1967. [Google Scholar] [CrossRef] [Green Version]
- Mills, M.C.; Lee, J. The threat of carbapenem-resistant bacteria in the environment: Evidence of widespread contamination of reservoirs at a global scale. Environ. Pollut. 2019, 255, 113143. [Google Scholar] [CrossRef]
- Grant, S.L.; Tamason, C.C.; Hoque, B.A.; Jensen, P.K.M. Drinking cholera: Salinity levels and palatability of drinking water in coastal Bangladesh. Trop. Med. Int. Health 2015, 20, 455–461. [Google Scholar] [CrossRef]
- Sedas, V.T.P. Influence of environmental factors on the presence of Vibrio cholerae in the marine environment: A climate link. J. Infect. Dev. Ctries. 2007, 1, 224–241. [Google Scholar] [CrossRef]
- Vezzulli, L.; Baker-Austin, C.; Kirschner, A.; Pruzzo, C.; Martinez-Urtaza, J. Global emergence of environmental non-O1/O139 Vibrio cholerae infections linked with climate change: A neglected research field? Environ. Microbiol. 2020, 22, 4342–4355. [Google Scholar] [CrossRef]
- Vezzulli, L.; Brettar, I.; Pezzati, E.; Reid, P.C.; Colwell, R.R.; Höfle, M.G.; Pruzzo, C. Long-term effects of ocean warming on the prokaryotic community: Evidence from the vibrios. ISME J. 2012, 6, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vezzulli, L.; Pezzati, E.; Brettar, I.; Höfle, M.; Pruzzo, C. Effects of Global Warming on Vibrio Ecology. Microbiol. Spectr. 2015, 3, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paz, S.; Bisharat, N.; Paz, E.; Kidar, O.; Cohen, D. Climate change and the emergence of Vibrio vulnificus disease in Israel. Environ. Res. 2007, 103, 390–396. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Trinanes, J.A.; Taylor, N.G.H.; Hartnell, R.; Siitonen, A.; Martinez-Urtaza, J. Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim. Chang. 2012, 3, 73–77. [Google Scholar] [CrossRef]
- Brehm, T.T.; Berneking, L.; Martins, M.S.; Dupke, S.; Jacob, D.; Drechsel, O.; Bohnert, J.; Becker, K.; Kramer, A.; Christner, M.; et al. Heatwave-associated Vibrio infections in Germany, 2018 and 2019. Eurosurveillance 2021, 26, 2002041. [Google Scholar] [CrossRef]
- Vezzulli, L.; Grande, C.; Reid, P.C.; Hélaouët, P.; Edwards, M.; Höfle, M.G.; Brettar, I.; Colwell, R.R.; Pruzzo, C. Climate influence on Vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc. Natl. Acad. Sci. USA 2016, 113, E5062–E5071. [Google Scholar] [CrossRef] [Green Version]
- Binsztein, N.; Costagliola, M.C.; Pichel, M.; Jurquiza, V.; Ramiírez, F.C.; Akselman, R.; Vacchino, M.; Huq, A.; Colwell, R. Viable but Nonculturable Vibrio cholerae O1 in the Aquatic Environment of Argentina. Appl. Environ. Microbiol. 2004, 70, 7481–7486. [Google Scholar] [CrossRef]
- Dörr, T.; Davis, B.M.; Waldor, M.K. Endopeptidase-Mediated Beta Lactam Tolerance. PLOS Pathog. 2015, 11, e1004850. [Google Scholar] [CrossRef] [Green Version]
- Halpern, M.; Senderovich, Y.; Izhaki, I. Waterfowl—the missing link in epidemic and pandemic cholera dissemination? PLoS Pathog. 2008, 4, e1000173. [Google Scholar] [CrossRef] [Green Version]
- Senderovich, Y.; Izhaki, I.; Halpern, M. Fish as Reservoirs and Vectors of Vibrio cholerae. PLoS ONE 2010, 5, e8607. [Google Scholar] [CrossRef] [Green Version]
- Kiiyukia, C.; Nakajima, A.; Nakai, T.; Muroga, K.; Kawakami, H.; Hashimoto, H. Vibrio cholerae non-O1 isolated from ayu fish (Plecoglossus altivelis) in Japan. Appl. Environ. Microbiol. 1992, 58, 3078–3082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, B.; Rehnstam-Holm, A.-S.; Lindmark, B.; Pal, A.; Wai, S.N.; Hernroth, B. The Origin of Vibrio cholerae Influences Uptake and Persistence in the Blue MusselMytilus edulis. J. Shellfish Res. 2012, 31, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Deriu, A.; Sechi, L.A.; Molicotti, P.; Spanu, M.; Zanetti, S. Virulence genes in halophilic Vibrio spp. isolated in common mussels. Microbiol. Q. J. Microbiol. Sci. 2002, 25, 93–96. [Google Scholar]
- Halpern, M.; Broza, Y.B.; Mittler, S.; Arakawa, E.; Broza, M. Chironomid Egg Masses as a Natural Reservoir of Vibrio cholerae Non-O1 and Non-O139 in Freshwater Habitats. Microb. Ecol. 2003, 47, 341–349. [Google Scholar] [CrossRef]
- Kesy, K.; Labrenz, M.; Scales, B.S.; Kreikemeyer, B.; Oberbeckmann, S. Vibrio Colonization Is Highly Dynamic in Early Microplastic-Associated Biofilms as Well as on Field-Collected Microplastics. Microorganisms 2021, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Bowley, J.; Baker-Austin, C.; Porter, A.; Hartnell, R.; Lewis, C. Oceanic Hitchhikers—Assessing Pathogen Risks from Marine Microplastic. Trends Microbiol. 2021, 29, 107–116. [Google Scholar] [CrossRef]
- Shikuma, N.J.; Hadfield, M.G. Marine biofilms on submerged surfaces are a reservoir for Escherichia coli and Vibrio cholerae. Biofouling 2010, 26, 39–46. [Google Scholar] [CrossRef]
- Alam, M.; Sultana, M.; Nair, G.B.; Siddique, A.K.; Hasan, N.A.; Sack, R.B.; Sack, D.A.; Ahmed, K.U.; Sadique, A.; Watanabe, H.; et al. Viable but nonculturable Vibrio cholerae O1 in biofilms in the aquatic environment and their role in cholera transmission. Proc. Natl. Acad. Sci. USA 2007, 104, 17801–17806. [Google Scholar] [CrossRef]
- Kumarasamy, K.K.; Toleman, M.A.; Walsh, T.R.; Bagaria, J.; Butt, F.; Balakrishnan, R.; Chaudhary, U.; Doumith, M.; Giske, C.G.; Irfan, S.; et al. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: A molecular, biological, and epidemiological study. Lancet Infect. Dis. 2010, 10, 597–602. [Google Scholar] [CrossRef]
- Castanheira, M.; Deshpande, L.M.; Mathai, D.; Bell, J.M.; Jones, R.N.; Mendes, R.E. Early Dissemination of NDM-1- and OXA-181-Producing Enterobacteriaceae in Indian Hospitals: Report from the SENTRY Antimicrobial Surveillance Program, 2006–2007. Antimicrob. Agents Chemother. 2011, 55, 1274–1278. [Google Scholar] [CrossRef] [Green Version]
- Collignon, P.; Beggs, J.J.; Walsh, T.R.; Gandra, S.; Laxminarayan, R. Anthropological and socioeconomic factors contributing to global antimicrobial resistance: A univariate and multivariable analysis. Lancet Planet. Health 2018, 2, e398–e405. [Google Scholar] [CrossRef]
- Anjali, R.; Shanthakumar, S. Insights on the current status of occurrence and removal of antibiotics in wastewater by advanced oxidation processes. J. Environ. Manag. 2019, 246, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhuan, R. Degradation of antibiotics by advanced oxidation processes: An overview. Sci. Total Environ. 2020, 701, 135023. [Google Scholar] [CrossRef]
- McKinney, C.W.; Pruden, A. Ultraviolet Disinfection of Antibiotic Resistant Bacteria and Their Antibiotic Resistance Genes in Water and Wastewater. Environ. Sci. Technol. 2012, 46, 13393–13400. [Google Scholar] [CrossRef]
- Lüddeke, F.; Heß, S.; Gallert, C.; Winter, J.; Güde, H.; Löffler, H. Removal of total and antibiotic resistant bacteria in advanced wastewater treatment by ozonation in combination with different filtering techniques. Water Res. 2015, 69, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Petterson, S.R.; Ashbolt, N.J. QMRA and water safety management: Review of application in drinking water systems. J. Water Health 2016, 14, 571–589. [Google Scholar] [CrossRef] [PubMed]
- Noman, E.; Al-Gheethi, A.; Mohamed, R.M.S.R.; Talip, B.; Al-Sahari, M.; Al-Shaibani, M. Quantitative Microbiological Risk Assessment of Complex Microbial Community in Prawn Farm Wastewater and Applicability of Nanoparticles and Probiotics for Eliminating of Antibiotic-Resistant Bacteria. J. Hazard. Mater. 2021, 419, 126418. [Google Scholar] [CrossRef]
- Li, M.-F.; Liu, Y.-G.; Zeng, G.-M.; Liu, N.; Liu, S.-B. Graphene and graphene-based nanocomposites used for antibiotics removal in water treatment: A review. Chemosphere 2019, 226, 360–380. [Google Scholar] [CrossRef]
- Noman, E.; Al-Gheethi, A.; Talip, B.A.; Mohamed, R.; Kassim, A.H. Inactivating pathogenic bacteria in greywater by biosynthesized Cu/Zn nanoparticles from secondary metabolite of Aspergillus iizukae; optimization, mechanism and techno economic analysis. PLoS ONE 2019, 14, e0221522. [Google Scholar] [CrossRef] [Green Version]
- Gopinath, P.M.; Dhanasekaran, D.; Ranjani, A.; Thajuddin, N.; Akbarsha, M.A.; Velmurugan, M.; Panneerselvam, A. Optimization of sporicidal activity and environmental Bacillus endospores decontamination by biogenic silver nanoparticle. Futur. Microbiol. 2015, 10, 725–741. [Google Scholar] [CrossRef]
- Collivignarelli, M.C.; Abbà, A.; Benigna, I.; Sorlini, S.; Torretta, V. Overview of the Main Disinfection Processes for Wastewater and Drinking Water Treatment Plants. Sustainability 2018, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Azman, A.S.; Lauer, S.A.; Bhuiyan, T.R.; Luquero, F.J.; Leung, D.T.; Hegde, S.T.; Harris, J.B.; Paul, K.K.; Khaton, F.; Ferdous, J.; et al. Vibrio cholerae O1 transmission in Bangladesh: Insights from a nationally representative serosurvey. Lancet Microbe 2020, 1, e336–e343. [Google Scholar] [CrossRef]
- Singer, R.S.; Ward, M.P.; Maldonado, G. Erratum: Can landscape ecology untangle the complexity of antibiotic resistance? Nat. Rev. Microbiol. 2007, 5, 82. [Google Scholar] [CrossRef]
- Saha, G.K.; Ganguly, N.K. Spread and Endemicity of Cholera in India: Factors Beyond the Numbers. J. Infect. Dis. 2021, 224 (Suppl. S7), S710–S716. [Google Scholar] [CrossRef]
- Janecko, N.; Martz, S.-L.; Avery, B.P.; Daignault, D.; Desruisseau, A.; Boyd, D.; Irwin, R.J.; Mulvey, M.R.; Reid-Smith, R.J. Carbapenem-Resistant Enterobacter spp. in Retail Seafood Imported from Southeast Asia to Canada. Emerg. Infect. Dis. 2016, 22, 1675–1677. [Google Scholar] [CrossRef]
- Newton, A.; Kendall, M.; Vugia, D.J.; Henao, O.L.; Mahon, B.E. Increasing rates of vibriosis in the United States, 1996–2010: Review of surveillance data from 2 systems. Clin. Infect. Dis. 2012, 54, S391–S395. [Google Scholar] [CrossRef]
- World Health Organization. National Antimicrobial Resistance Surveillance Systems and Participation in the Global Antimicrobial Resistance Surveillance System (GLASS): A Guide to Planning, Implementation, and Monitoring and Evaluation; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Gentilini, F.; Turba, M.E.; Pasquali, F.; Mion, D.; Romagnoli, N.; Zambon, E.; Terni, D.; Peirano, G.; Pitout, J.D.D.; Parisi, A.; et al. Hospitalized Pets as a Source of Carbapenem-Resistance. Front. Microbiol. 2018, 9, 2872. [Google Scholar] [CrossRef]
- Levy, S. ECDC Vibrio Map Viewer: Tracking the Whereabouts of Pathogenic Species. Environ. Health Perspect. 2018, 126, 034003. [Google Scholar] [CrossRef]
- Racault, M.-F.; Abdulaziz, A.; George, G.; Menon, N.; Punathil, M.; McConville, K.; Loveday, B.; Platt, T.; Sathyendranath, S.; Vijayan, V. Environmental Reservoirs of Vibrio cholerae: Challenges and Opportunities for Ocean-Color Remote Sensing. Remote Sens. 2019, 11, 2763. [Google Scholar] [CrossRef] [Green Version]
- Yadav, R.; Landersdorfer, C.B.; Nation, R.L.; Boyce, J.D.; Bulitta, J.B. Novel Approach to Optimize Synergistic Carbapenem-Aminoglycoside Combinations against Carbapenem-Resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 2015, 59, 2286–2298. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Jang, A.; Yoon, Y.K.; Kim, Y. Development of Novel Peptides for the Antimicrobial Combination Therapy against Carbapenem-Resistant Acinetobacter baumannii Infection. Pharmaceutics 2021, 13, 1800. [Google Scholar] [CrossRef] [PubMed]
- Zhanel, G.G.; Lawrence, C.K.; Adam, H.; Schweizer, F.; Zelenitsky, S.; Zhanel, M.; Lagacé-Wiens, P.R.; Walkty, A.; Denisuik, A.; Golden, A.; et al. Imipenem–relebactam and meropenem–vaborbactam: Two novel carbapenem-β-lactamase inhibitor combinations. Drugs 2018, 78, 65–98. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.C.; Zmarlicka, M.T.; Shaeer, K.M.; Pardo, J. Meropenem/vaborbactam, the first carbapenem/β-lactamase inhibitor combination. Ann. Pharmacother. 2018, 52, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Everett, M.; Sprynski, N.; Coelho, A.; Castandet, J.; Bayet, M.; Bougnon, J.; Lozano, C.; Davies, D.T.; Leiris, S.; Zalacain, M.; et al. Discovery of a Novel Metallo-β-Lactamase Inhibitor That Potentiates Meropenem Activity against Carbapenem-Resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2018, 62, e00074-18. [Google Scholar] [CrossRef] [Green Version]
- Bhowmick, T.; Weinstein, M.P. Microbiology of Meropenem-Vaborbactam: A Novel Carbapenem Beta-Lactamase Inhibitor Combination for Carbapenem-Resistant Enterobacterales Infections. Infect. Dis. Ther. 2020, 9, 757–767. [Google Scholar] [CrossRef]
- El Hafi, B.; Rasheed, S.S.; Abou Fayad, A.; Araj, G.F.; Matar, G.M. Evaluating the Efficacies of Carbapenem/β-Lactamase Inhibitors Against Carbapenem-Resistant Gram-Negative Bacteria in vitro and in vivo. Front. Microbiol. 2019, 10, 933. [Google Scholar] [CrossRef]
- Tan, L.T.-H.; Lee, L.-H.; Goh, B.-H. The Bioprospecting of Anti-Vibrio Streptomyces species: Prevalence and Applications. Prog. Microbes Mol. Biol. 2019, 2, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Tan, L.T.-H.; Chan, K.-G.; Lee, L.-H.; Goh, B.-H. Streptomyces bacteria as potential probiotics in aquaculture. Front. Microbiol. 2016, 7, 79. [Google Scholar] [CrossRef]
- Newman, A.M.; Arshad, M. The Role of Probiotics, Prebiotics and Synbiotics in Combating Multidrug-Resistant Organisms. Clin. Ther. 2020, 42, 1637–1648. [Google Scholar] [CrossRef]
- Piatek, J.; Bernatek, M.; Ciechelska-Rybarczyk, A.; Oleskow, B.; Sommermeyer, H. Inhibition of carbapenem-resistant ndm-1 Klebsiella pneumoniae isolated from a Hospital outbreak patient by a synbiotic: A nonantibiotic treatment option. Int. J. Med. Res. Health Sci. 2019, 8, 12–20. [Google Scholar]
- Tan, L.T.-H.; Lee, L.-H.; Goh, B.-H. Critical review of fermentation and extraction of anti-Vibrio compounds from Streptomyces. Prog. Microbes Mol. Biol. 2020, 3, 1–14. [Google Scholar] [CrossRef]
- Goh, J.X.; Tan, L.T.; Law, J.W.; Ser, H.L.; Khaw, K.Y.; Letchumanan, V.; Lee, L.H.; Goh, B.H. Harnessing the potentialities of probiotics, prebiotics, synbiotics, paraprobiotics, and postbiotics for shrimp farming. Rev. Aquac. 2022, 14, 1478–1557. [Google Scholar] [CrossRef]
- Letchumanan, V.; Chan, K.-G.; Pusparajah, P.; Saokaew, S.; Duangjai, A.; Goh, B.-H.; Ab Mutalib, N.-S.; Lee, L.-H. Insights into Bacteriophage Application in Controlling Vibrio Species. Front. Microbiol. 2016, 7, 1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Zhang, Y.; Lan, W.; Sun, X. Characterization of vB_VpaP_MGD2, a newly isolated bacteriophage with biocontrol potential against multidrug-resistant Vibrio parahaemolyticus. Arch. Virol. 2021, 166, 413–426. [Google Scholar] [CrossRef]
- Tan, T.; Chan, K.; Lee, L. Application of bacteriophage in biocontrol of major foodborne bacterial pathogens. J. Mol. Biol. Mol. Imaging 2014, 1, 1–9. [Google Scholar]
- Zhong, S.; He, S. Quorum Sensing Inhibition or Quenching in Acinetobacter baumannii: The Novel Therapeutic Strategies for New Drug Development. Front. Microbiol. 2021, 12, 558003. [Google Scholar] [CrossRef]
- Thakur, P.; Chawla, R.; Tanwar, A.; Chakotiya, A.S.; Narula, A.; Goel, R.; Arora, R.; Sharma, R.K. Attenuation of adhesion, quorum sensing and biofilm mediated virulence of carbapenem resistant Escherichia coli by selected natural plant products. Microb. Pathog. 2016, 92, 76–85. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Continent | Country | Sample Type | Vibrio Species (Number of Strain) | Susceptibility to Carbapenem Antibiotics | Carbapenem Resistance Number of Strain (%) | MIC | Carbapenemase Gene Identified (Location) | Year of Isolation | Reference | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| R | I | S | |||||||||
| Asia | Bangladesh Chhatak (north-eastern Bangladesh) | Clinical | V. cholerae O1 (141) | IMI | 0 | 0 | 141 (100%) | - | Absent | 2009–2014 | [138] |
| Environmental | V. cholerae O1 (21) | IMI | 0 | 0 | 21 (100%) | - | Absent | ||||
| Bangladesh Mathbaria (south-western Bangladesh) | Clinical | V. cholerae O1 (178) | IMI | 0 | 0 | 178 (100%) | - | Absent | |||
| Environmental | V. cholerae O1 (120) | IMI | 0 | 0 | 120 (100%) | - | Absent | ||||
| Bangladesh Chhatak (north-eastern Bangladesh) | Clinical | V. cholerae O1 (68) | IMI | 0 | 1 (1%) | 67 (99%) | - | - | 2013 | [139] | |
| Environmental | V. cholerae O1 (6) | IMI | 1 (17%) | 0 | 5 (83%) | - | - | ||||
| Bangladesh Mathbaria (south-western Bangladesh) | Clinical [diarrheal sample] | V. cholerae O1 (77) | IMI | 0 | 0 | 77 (100%) | - | - | |||
| Environmental [ponds] | V. cholerae O1 (57) | IMI | 0 | 0 | 57 (100%) | - | - | ||||
| China [16 counties] | Clinical (429) Environment [water (77), soil/surface (62)] | V. cholerae El Tor (568) | IMI | 568 (100%) | 0 | 0 | - | - | 1986–2012 | [140] | |
| MER | 57 (10%) | 0 | 0 | - | |||||||
| China (Mohnarin hospitals in different areas) | Clinical | V. parahaemolyticus (51) | Carbapenem | - | - | (>90%) | - | - | 2010 | [141] | |
| China (south-eastern China) | Clinical [diarrheal sample] | V. parahaemolyticus (563) | Carbapenem | - | - | (>95%) | - | - | 2015 | [142] | |
| China [6 counties] | Clinical [diarrheal sample] | V. parahaemolyticus (2871) | IMI | (<1%) | - | - | Interpretive standard of resistance: MIC ≥ 4 µg/mL | - | 2016–2020 | [118] | |
| China Zhejiang | Freshwater farm, Seawater farm, Market | V. parahaemolyticus (360) | IMI (10 µg) | - | - | 360 (100%) | - | - | 2017–2019 | [143] | |
| China | Seafood [shrimp] | V. alginolyticus Vb1394 | IMI | 0 | 1 | 0 | 2 mg/L | - | 2018 | [144] | |
| MER | 0 | 1 | 0 | 2 mg/L | - | ||||||
| China Shenzhen | Seafood [shrimp] | V. alginolyticus Vb1978 | IMI | - | - | - | 0.5 mg/L | NDM-1, VIM-1 (plasmid;50 kb) | 2019 | [145] | |
| MER | - | - | - | 0.03 mg/L | |||||||
| China Bohai Bay, Tianjin | Environmental [Coastal seawater)] | Vibrio sp. (carrying blaOXA58) | IMI | - | - | - | 2 µg/mL | OXA-58 (plasmid) | 2019 | [23] | |
| MER | - | - | - | 8 µg/mL | |||||||
| China | Clinical [diarrheal sample] Seafood [from markets] | V. parahaemolyticus (107) | Carbapenem | - | - | (90–100%) | - | - | 2019 | [146] | |
| China | Seafood [shrimp] | V. alginolyticus Vb1796 | IMI | - | - | - | 0.12 mg/L | VMB-1 | 2020 | [147] | |
| MER | - | - | - | 1 mg/L | |||||||
| China | Seafood [shrimp] | V. alginolyticusVb1579 | IMI | 0 | 0 | 1 | Standard agar dilution method: (0.06 mg/L) | VAM-1 | 2021 | [135] | |
| MER | 0 | 0 | 1 | (0.12 mg/L) | |||||||
| China | Seafood [shrimp] | V. diabolicus SLV18 | IMI | 0 | 0 | 1 | - | VMB-2 - | - | [148] | |
| MER | - | - | - | 4 µg/mL (Highly resistant) | |||||||
| China | collection center | V. vulnificus | - | - | - | - | - | Vmh (chromosome) | - | [73] | |
| India Kolkata (east India) Delhi (north India) | Clinical [diarrheal sample] | V. cholerae (443) | IMI | ~ <20 (~ <5%) | - | - | - | NDM-1 (chromosome) | 2008–2015 | [59] | |
| India | Clinical [diarrheal sample] | V. fluvialis (115) | IMI | 27 (24%) | - | - | 4-32 µg/mL | 27 (24%) NDM-1 (Class 1 integron) 27 (24%) OXA-1, OXA-7, OXA-9 | 2009–2013 | [57] | |
| India Central New Delhi | Environmental [seepage, tap water, sewage effluent] | V. cholerae 116-14 | IMI | - | - | - | 8 mg/L | NDM-1 (chromosome and plasmid; 400 kb) | 2010 | [149] | |
| MER | - | - | - | 8 mg/L | |||||||
| V. cholerae 116-17 | IMI | - | - | - | 16 mg/L | NDM-1 (plasmid Inc A/C;170kb) | |||||
| MER | - | - | - | 1 mg/L | |||||||
| India Puducherry | Clinical [fecal sample of a child] | V. cholerae O1 El Tor Ogawa | - | - | - | - | - | NDM-1 (plasmid) | 2012 | [150] | |
| India (Southwest coast) | Environmental [water, sediment sample from estuary] | Vibrio sp. (180) | - | - | - | - | - | (13%) NDM-1 | 2012 | [151] | |
| Environmental [water, sediment sample from shrimp farms] | Vibrio sp. (70) | - | - | - | - | - | (14%) NDM-1 | ||||
| Seafood [retail] | Vibrio sp. (30) | - | - | - | - | - | (7%) NDM-1 | ||||
| India Kerala | Environmental [sediment, water sample from aquaculture farm] Seafood [shrimp] | V. parahaemolyticus (27) | MER (10 µg) | 1 | 8 | 18 | - | (4%) produces carbapenemase | 2015–2016 | [34] | |
| Korea | Seafood [10 types] | V. parahaemolyticus | IMI | (70%) | (13%) | (17%) | - | - | 2009 | [134] | |
| V. parahaemolyticus | MER | (50%) | (29%) | (221%) | - | - | |||||
| Korea Southern coast of South Korea | Environment [mud, tidal water] | V. parahaemolyticus (1720) | MER (10 µg) | 4 (<1%) | - | - | 64 µg/mL 128 µg/mL 128 µg/mL 1024 µg/mL | - | 2013–2014 | [152] | |
| Korea Cheongju | Seafood [shrimp from retail outlet] | V. parahaemolyticus (27) | IMI | - | - | - | MIC50 = ≤1 mg/L; MIC90 = ≤1 mg/L Imipenem (range = 1–8 mg/L) | - | 2016 | [153] | |
| Korea | Seafood [Cockles] | V. parahaemolyticus (4) | IMI (10 µg) | 0 | 3 (75%) | 1 (25%) | - | - | - | [82] | |
| V. alginolyticus (11) | IMI (10 µg) | 0 | 3 (27%) | 8 (73%) | - | ||||||
| V. diabolicus (14) | IMI (10 µg) | 0 | 5 (36%) | 9 (64%) | - | ||||||
| V. harveyi (3) | IMI (10 µg) | 0 | 2 (67%) | 1 (33%) | - | ||||||
| Korea | Seafood [hard shell mussel] | Total Vibrio sp. (32) V. parahaemolyticus (2) V. harveyi (1) V. alginolyticus (13) V. diabolicus (16) | IMI (10 µg) | (3%) | (3%) | (94%) | - | 1(3%) OXA [V. diabolicus] | - | [133] | |
| Malaysia Selangor | Seafood [freshwater fish] | V. parahaemolyticus (49) V. cholerae (8) | IMI | 0 | 0 | 57 (100%) | - | - | - | [154] | |
| Malaysia Selangor | Seafood [shellfish] | V. parahaemolyticus (200) | IMI (10 µg) | 1 (<1%) | 18 (9%) | 181 (90%) | - | - | 2014 | [29] | |
| Malaysia | Seafood [shrimp] | V. parahaemolyticus (185) | IMI (10 µg) | 4 (2%) | - | - | - | - | 2014 | [30] | |
| Seafood [shrimp and shellfish] | V. parahaemolyticus (385) | IMI (10 µg) | 5 (1%) | 18 (5%) | 362 (94%) | - | - | - | [28] | ||
| Malaysia Selangor | Seafood [marine and freshwater fish] | V. parahaemolyticus (165) | IMI (10 µg) | 19 (12%) | 6 (4%) | 140 (85%) | - | - | 2016 | [27] | |
| Malaysia Selangor | Seafood [shrimps, clams, squid] | V. parahaemolyticus (120) | IMI (10 µg) | 0 | 2 (2%) | 118 (98%) | - | - | 2018 | [123] | |
| MER (10 µg) | 0 | 2 (2%) | 118 (98%) | - | |||||||
| Malaysia Selangor | Seafood [shellfish] | V. parahaemolyticus (43) | IMI (10 µg) | 10 (23%) | 0 | 33 (77%) | - | - | - | [32] | |
| Singapore | Environment [harbor, ballast water from ships] | V. alginolyticus | MER | 3 | - | - | - | - | 2016 | [84] | |
| V. parahaemolyticus | MER | 2 | - | - | - | - | |||||
| V. vulnificus | MER | 6 | - | - | - | - | |||||
| V. brasiliensis | MER | 1 | - | - | - | - | |||||
| V. campbellii | MER | 18 | - | - | - | - | |||||
| V. rotiferianus | MER | 1 | - | - | - | - | |||||
| V. tubiashii | MER | 1 | - | - | - | - | |||||
| Vietnam (Southern Vietnam) | Environmental | V. cholerae (100) non-O1, non-O139 | - | - | - | - | - | 3 (3%) NDM-1 | 2010–2013 | [155] | |
| Oceania | Australia Northern territory | Clinical [infection site] | Vibrio sp. (44) | MER | - | - | (93%) | - | - | 2000–2013 | [156] |
| Africa | Egypt Mansoura | Seafood [fish and shellfish] | V. parahaemolyticus (50) | IMI (10 µg) | 12 (24%) | 4 (8%) | 34 (68%) | - | - | - | [75] |
| V. alginolyticus (42) | IMI (10 µg) | 8 (19%) | 10 (24%) | 24 (57%) | - | - | |||||
| Nigeria (South-south Nigeria) | Seafood [shellfish] | Vibrio sp. (6) | IMI (10 µg) | 6 (100%) | 0 | 0 | - | - | 2015–2017 | [121] | |
| Nigeria (Eight states in Southern Nigeria) | African salad | V. parahaemolyticus (63) | IMI (10 µg) | 2 (3%) | 7 (11%) | 54 (86%) | - | - | 2018–2019 | [132] | |
| Nigeria Lagos | Environment [sea water, wet and dry sand] | V. parahaemolyticus (26) | IMI | (4%) | - | - | - | 1 (4%) NDM-1 | - | [60] | |
| MER | (4%) | - | - | - | - | ||||||
| ETP | (4%) | - | - | - | - | ||||||
| V. vulnificus (14) | IMI | (57%) | - | - | - | 5 (36%) NDM-1 | |||||
| MER | (57%) | - | - | - | - | ||||||
| ETP | (57%) | - | - | - | - | ||||||
| South Africa [Chris Hani, Amahlathi, Lukhanji] | Environment [Final effluents from dams, earth canals, rivers, receiving water bodies, tap water and wastewater treatment units] | V. cholerae (61) | IMI (10 µg) | 0 | 2 (3%) | 59 (97%) | - | 19 (31%) NDM-1, GES, IMP, VIM 14 (23%) produce carbapenemase | 2018 | [157] | |
| MER (10 µg) | 0 | 0 | 61 (100%) | - | |||||||
| ETP (10 µg) | 5 (8%) | 3 (5%) | 53 (87%) | - | |||||||
| DOR (10 µg) | 1 (2%) | 2 (3%) | 59 (97%) | - | |||||||
| South Africa [Eastern Cape] | Environment [wastewater effluent] | Total Vibrio sp. (52) V. parhaemolyticus (12) V. vulnificus (18) V. fluvialis (19) V. metschnikovii (3) | IMI (10 µg) MER (10 µg) | 0 | 0 | 52 (100%) | - | - | 2010 | [158] | |
| South Africa [Eastern Cape] | Environment [river water] | Total Vibrio sp. (118) | IMI (10 µg) | 39 (33%) | 40 (34%) | 39 (33%) | - | 35(30%) VIM, 13 (11%) OXA-48, 18 (15%) IMP, 20 (17%) GES, 7 (6%) KPC | 2018 | [131] | |
| MER (10 µg) | 32 (27%) | 21 (18%) | 65 (55%) | - | - | ||||||
| V. mimicus (40) | IMI (10 µg) | 2 (5%) | 17 (43%) | 21 (53%) | - | - | |||||
| MER (10 µg) | 7 (18%) | 8 (20%) | 25 (63%) | - | - | ||||||
| V. vulnificus (37) | IMI (10 µg) | 26 (70%) | 10 (27%) | 1 (3%) | - | - | |||||
| MER (10 µg) | 17 (46%) | 7 (19%) | 13 (35%) | - | - | ||||||
| V. fluvialis (41) | IMI (10 µg) | 11 (27%) | 13 (32%) | 17 (41%) | - | - | |||||
| MER (10 µg) | 8 (20%) | 6 (15%) | 27 (66%) | - | - | ||||||
| Uganda (South-western district) | Environmental [surface water] | Vibrio sp. (392) | MER (10 µg) | 180 (46%) | 38 (10%) | 174 (44%) | - | - | 2019 | [159] | |
| Europe | Italy [Northern Sardinia] | Environmental [water sample from coast and gulf] | V. alginolyticus (40) | IMI (10 µg) | - | - | - | MIC50 = 0.06 mg/L; MIC90 = 0.12 mg/L Imipenem (range = 0.06–2 mg/L) | - | - | [85] |
| MER (10 µg) | - | - | - | MIC50 = 0.06 mg/L; MIC90 = 0.06 mg/L Meropenem (range = 0.03–0.25 mg/L) | - | ||||||
| V. parahaemolyticus (8) | IMI (10 µg) | - | - | - | MIC50 = 0.125 mg/L; MIC90 = 1 mg/L Imipenem (range = 0.06–1 mg/L) | - | |||||
| MER (10 µg) | - | - | - | MIC50 = 0.006 mg/L; MIC90 = 0.12mg/L Meropenem (range = 0.004–0.12 mg/L) | - | ||||||
| V. vulnificus (6) | IMI (10 µg) | - | - | - | MIC50 = 0.12 mg/L; MIC90 = 0.12 mg/L Imipenem (range = 0.06–2 mg/L) | - | |||||
| MER (10 µg) | - | - | - | MIC50 = 0.006 mg/L; MIC90 = 0.12mg/L Meropenem (range = 0.004–0.25 mg/L) | - | ||||||
| Italy | Environmental [Seawater] | V. cholerae (12) | MER (10 µg) | 0 | 6 (50%) | 6 (50%) | - | - | 2003–2014 | [160] | |
| Environmental [Freshwater] | V. cholerae (5) | MER (10 µg) | 0 | 1 (20%) | 4 (80%) | - | - | ||||
| Seafood | V. cholerae (25) | MER (10 µg) | 1 (4%) | 4 (16%) | 20 (80%) | - | - | ||||
| Italy (north-western Adriatic Sea coasts) | Seafood [European Seabass] | V. anguillarum 28AD | IMI (10 µg) | 0 | 0 | 1 | 4 µg/mL | - | 2007–2011 | [161] | |
| Environmental [Water from Celeri Lagoon] | V. parahaemolyticus VPE116 | IMI (10 µg) | 0 | 0 | 1 | 0.125 µg/mL | - | ||||
| Italy (north-western Adriatic Sea coasts) | Seafood [shellfish] Wildlife [turtle blood] Environmental [beach, brackish water] | V. vulnificus (40) | IMI (10 µg) | 0 | 0 | 40 (100%) | - | - | - | [162] | |
| MER (10 µg) | 0 | 0 | 40 (100%) | - | |||||||
| France Port-Saint-Louis | Wildlife [gull] | V. cholerae non-O1/non-O139 | IMI | - | - | - | 3 mg/L | VIM-1, VIM-4 (plasmid) | 2013 | [163] | |
| MER | - | - | - | 0.5 mg/L | |||||||
| ETP | - | - | - | 0.19 mg/L | |||||||
| DOR | - | - | - | 0.75 mg/L | |||||||
| France (imported) | Seafood [shrimp] | V. parahaemolyticus | IMI | 0 | 0 | (100%) | - | NDM-1 Produces carbapenemase | 2016 | [74] | |
| Germany [Baltic Sea, the North Sea, Ems and Weser River estuaries] | Total (184) | V. cholerae V. vulnificus | IMI (10 µg) | (2%) | (1%) | (97%) | - | - | 2004–2014 | [21] | |
| MER (10 µg) | (<1%) | (2%) | (98%) | ||||||||
| Retail (35) | V. cholerae V. vulnificus | IMI (10 µg) | 0 | 0 | (100%) | - | - | ||||
| MER (10 µg) | 0 | 0 | (100%) | ||||||||
| Clinical (18) | V. cholerae V. vulnificus | IMI (10 µg) | 0 | 0 | (100%) | - | - | ||||
| MER (10 µg) | 0 | 0 | (100%) | ||||||||
| Environmental (131) | V. cholerae V. vulnificus | IMI (10 µg) | (3%) | (2%) | (95%) | - | - | ||||
| MER (10 µg) | (1%) | (2%) | (97%) | ||||||||
| North Sea (52) | V. cholerae V. vulnificus | IMI (10 µg) | (6%) | 0 | (94%) | - | - | ||||
| MER (10 µg) | (2%) | (4%) | (94%) | ||||||||
| Baltic Sea (79) | V. cholerae V. vulnificus | IMI (10 µg) | (1%) | (3%) | (96%) | - | - | ||||
| MER (10 µg) | 0 | (1%) | (99%) | ||||||||
| Germany [Baltic Sea, the North Sea] | Environmental [Coastal water] | V. cholerae (4) | IMI MER | 4 (100%) | 0 | 0 | - | 3 VCC-1 | - | [130] | |
| Norway [temperate and Polar Oceanic area] | Environmental [Seawater, sea creatures] | V. alginolyticus (53) | IMI (10 µg) | 0 | 2 (4%) | 51 (96%) | 2–8 µg/mL | - | 2018 | [83] | |
| MER (10 µg) | 0 | 0 | 53 (100%) | - | |||||||
| V. anguillarum (21) | IMI (10 µg) | 21 (100%) | 0 | 0 | - | - | |||||
| MER (10 µg) | 0 | 0 | 21 (100%) | - | |||||||
| V. antiquaries (2) | IMI (10 µg) MER (10 µg) | 0 | 0 | 2 (100%) | - | - | |||||
| V. fujianensis (2) | IMI (10 µg) | 0 | 1 (50%) | 1 (50%) | - | - | |||||
| MER (10 µg) | 0 | 0 | 2 (100%) | - | |||||||
| V. metschnovikovii (38) | IMI (10 µg) MER (10 µg) | 0 | 0 | 38(100%) | - | - | |||||
| Slovakia (Eastern and southern) | Environmental [freshwater] | V. cholerae (21) | IMI (10 µg) | 0 | 8 (38%) | 13 (62%) | - | - | 2016 | [70] | |
| Spain | Clinical [leg ulcer] | V. metschnikovii | IMI | 0 | 0 | 1 | <1 µg/mL | - | 2008 | [164] | |
| UK | Clinical [blood sample of a burn patient] | V. cholerae | MER | 1 | 0 | 0 | - | NDM-1 | 2011 | [165] | |
| South America | Brazil | Seafood [shrimp] | Vibrio (26) | IMI (10 µg) [in seawater] | 1 (4%) [V. navarrensis] | 0 | 25 (96%) | - | - | - | [81] |
| IMI (10 µg) [in distilled water] | 0 | 0 | 26 (100%) | - | |||||||
| Brazil Guanabara Bay | Environment [water sample from polluted estuary] | V. parahaemolyticus (150) | IMI | 1 (<1%) | 2 (1%) | 147 (99%) | - | - | 2018–2019 | [20] | |
| Environment [water sample from polluted estuary] | V. alginolyticus (1) | IMI | 1 | 0 | 0 | - | - | ||||
| Ecuador Cuenca | Seafood [shrimp] | V. parahaemolyticus (154) | IMI (10 µg) | (<1%) | 0 | (99%) | - | - | 2012 | [122] | |
| MER | 0 | 0 | (100%) | - | - | ||||||
| North America | Maryland Coastal Bays, Chesapeake Bay | Environmental [Surface water] | V. parahaemolyticus (77) | IMI | 0 | 0 | 77 (100%) | (IMI; 2–16 µg/mL) | - | 2009 | [166] |
| MER | 0 | 0 | 77 (100%) | (MERO; 2–16 µg/mL) | |||||||
| V. vulnificus (120) | IMI (10 µg) | 2(2%) | 0 | 118 (98%) | - | - | |||||
| MER (10 µg) | 0 | 0 | 120 (100%) | - | |||||||
| Canada (imported) | Seafood [shrimp; imported from India] | V. cholerae | DOR, ETP, IMI, MER | 1 | 0 | 0 | >32 µg/mL | VCC-1 (plasmid) | 2014 | [31] | |
| - | - | Culture collection center | V. cholerae (20) | IMI (10 µg) | - | - | - | MIC50 = 0.25 µg/mL; MIC90 = 0.25 µg/mL Imipenem (range = 0.0–0.5 µg/mL) | - | - | [51] |
| MER (10 µg) | - | - | - | MIC50 = 2 mg/L; MIC90 = 2 mg/L Meropenem (range = 1–2 mg/L) | |||||||
| V. parahaemolyticus (20) | IMI (10 µg) | - | - | - | MIC50 = 0.06 µg/mL; MIC90 = 0.06 µg/mL Imipenem (range = 0.06 µg/mL) | - | |||||
| MER (10 µg) | - | - | - | MIC50 = 0.06 mg/L MIC90 = 0.5mg/L Meropenem (range = 0.06–0.5 mg/L) | |||||||
| V. vulnificus (20) | IMI (10 µg) | - | - | - | MIC50 = 0.06 µg/mL Imipenem (range = 0.06 µg/mL) | - | |||||
| MER (10 µg) | - | - | - | MIC50 = 1.12 mg/L; MIC90 = 0.25 mg/L Meropenem (range = 0.06–0.25 mg/L) | |||||||
| Ambler Class | Active Site | Representative Carbapenemase Type | References |
|---|---|---|---|
| A | Serine | K. pneumoniae carbapenemase (KPC) * VCC * Imipenemase MBL (IMP) * Guiana extended-spectrum (GES) * Serratia fonticola carbapenemase (SFC) Serratia marcescens enzyme (SME) Non-metallo-carbapenemase-A (NMC-A) | [4,41,47,152] |
| B | Zinc (cofactor) | New Delhi MBL (NDM)* Imipenemase; active-on-imipenem (IMP) * Verona integron-encoded MBL (VIM) * German imipenemase (GIM) Seoul imipenemase (SIM) Australian imipenemase (AIM-1) Dutch imipenemase (DIM-1) Florence imipenemase (FIM-1) S. marcescens MBL (SMB-1) Sao Paulo MBL(SPM) Kyorin Health Science MBL (KHM-1) Tripoli MBL (TMB-1) Pseudomonas Fluorescens MBL (PFM) Vibrio alginolyticus MBL (VAM) * Vibrio MBL (VMB) * MBL fold metallohydrolase (Vmh) * | [47,73,147,183] |
| C | Serine | Not considered as carbapenemase | [47] |
| D | Serine | Oxacillinases (OXA) * OXA-23 OXA-24/-40 OXA-25 OXA-26 OXA-27 OXA-40 OXA−48 * OXA-49 OXA−51 OXA−58 * OXA-72 OXA-143 | [42,149] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goh, J.X.H.; Tan, L.T.-H.; Law, J.W.-F.; Khaw, K.-Y.; Ab Mutalib, N.-S.; He, Y.-W.; Goh, B.-H.; Chan, K.-G.; Lee, L.-H.; Letchumanan, V. Insights into Carbapenem Resistance in Vibrio Species: Current Status and Future Perspectives. Int. J. Mol. Sci. 2022, 23, 12486. https://doi.org/10.3390/ijms232012486
Goh JXH, Tan LT-H, Law JW-F, Khaw K-Y, Ab Mutalib N-S, He Y-W, Goh B-H, Chan K-G, Lee L-H, Letchumanan V. Insights into Carbapenem Resistance in Vibrio Species: Current Status and Future Perspectives. International Journal of Molecular Sciences. 2022; 23(20):12486. https://doi.org/10.3390/ijms232012486
Chicago/Turabian StyleGoh, Joanna Xuan Hui, Loh Teng-Hern Tan, Jodi Woan-Fei Law, Kooi-Yeong Khaw, Nurul-Syakima Ab Mutalib, Ya-Wen He, Bey-Hing Goh, Kok-Gan Chan, Learn-Han Lee, and Vengadesh Letchumanan. 2022. "Insights into Carbapenem Resistance in Vibrio Species: Current Status and Future Perspectives" International Journal of Molecular Sciences 23, no. 20: 12486. https://doi.org/10.3390/ijms232012486
APA StyleGoh, J. X. H., Tan, L. T. -H., Law, J. W. -F., Khaw, K. -Y., Ab Mutalib, N. -S., He, Y. -W., Goh, B. -H., Chan, K. -G., Lee, L. -H., & Letchumanan, V. (2022). Insights into Carbapenem Resistance in Vibrio Species: Current Status and Future Perspectives. International Journal of Molecular Sciences, 23(20), 12486. https://doi.org/10.3390/ijms232012486