

Macrophage Phenotypes and Gene Expression Patterns Are Unique in Naturally Occurring Metabolically Healthy Obesity
 , , and
, , and
Abstract
:1. Introduction
2. Results
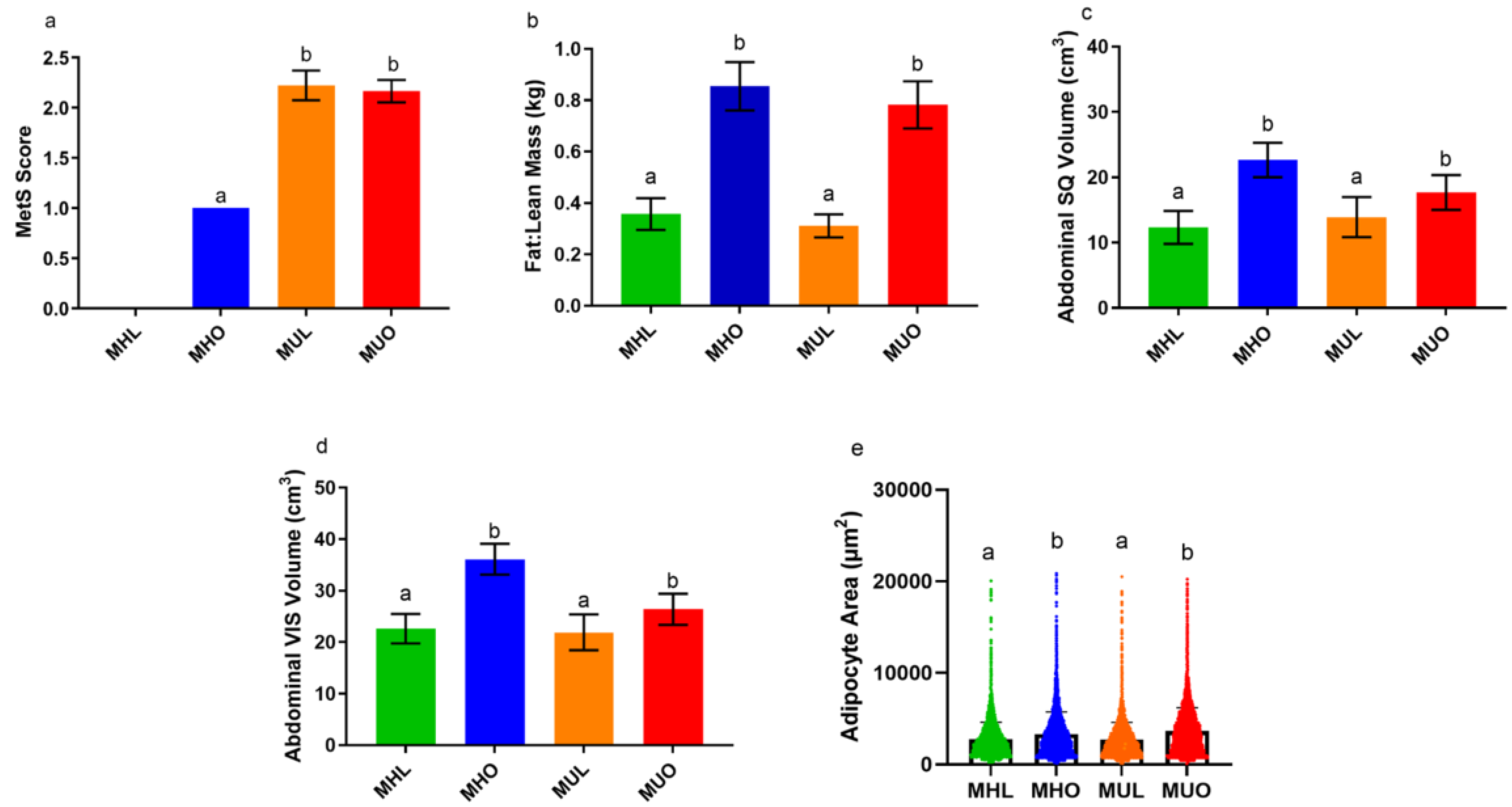
2.1. Abdominal Adipose Tissue Distribution by Depot, Adipocyte Size, and Circulating Insulin Do Not Explain Poor Metabolic Health
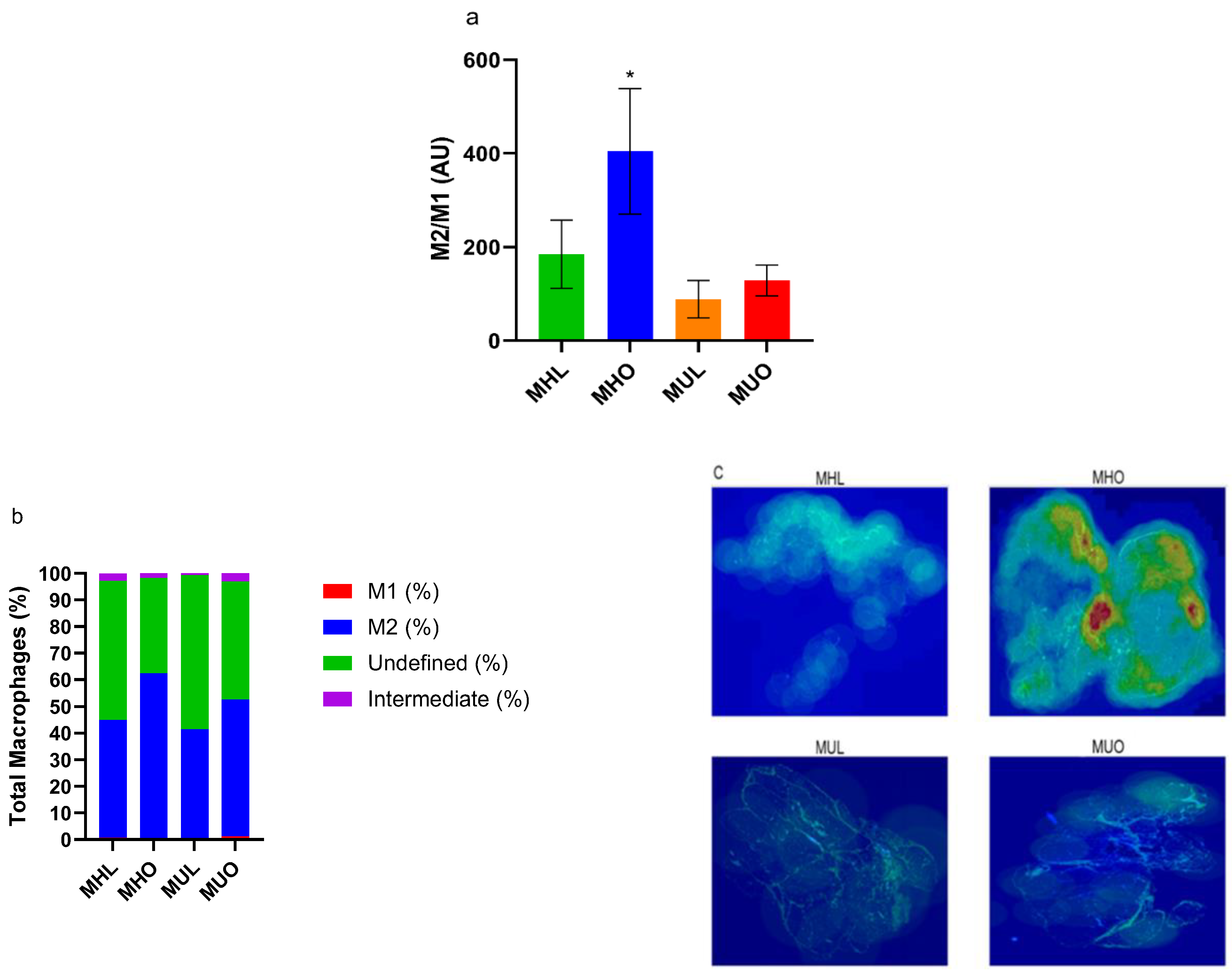
2.2. Metabolically Healthy Obese Subcutaneous Adipose Demonstrates Increased Anti-Inflammatory M2 Macrophage Accumulation
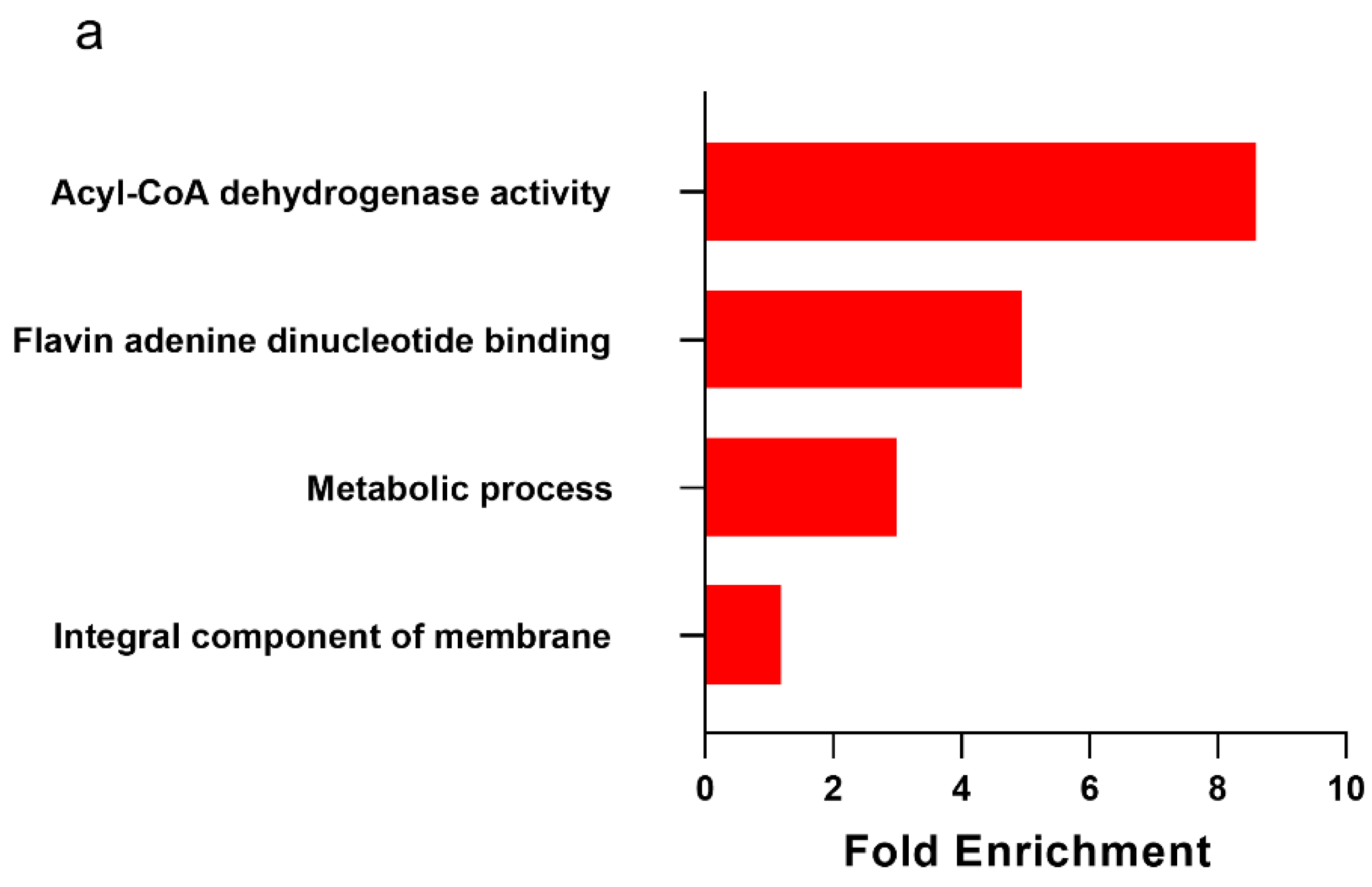
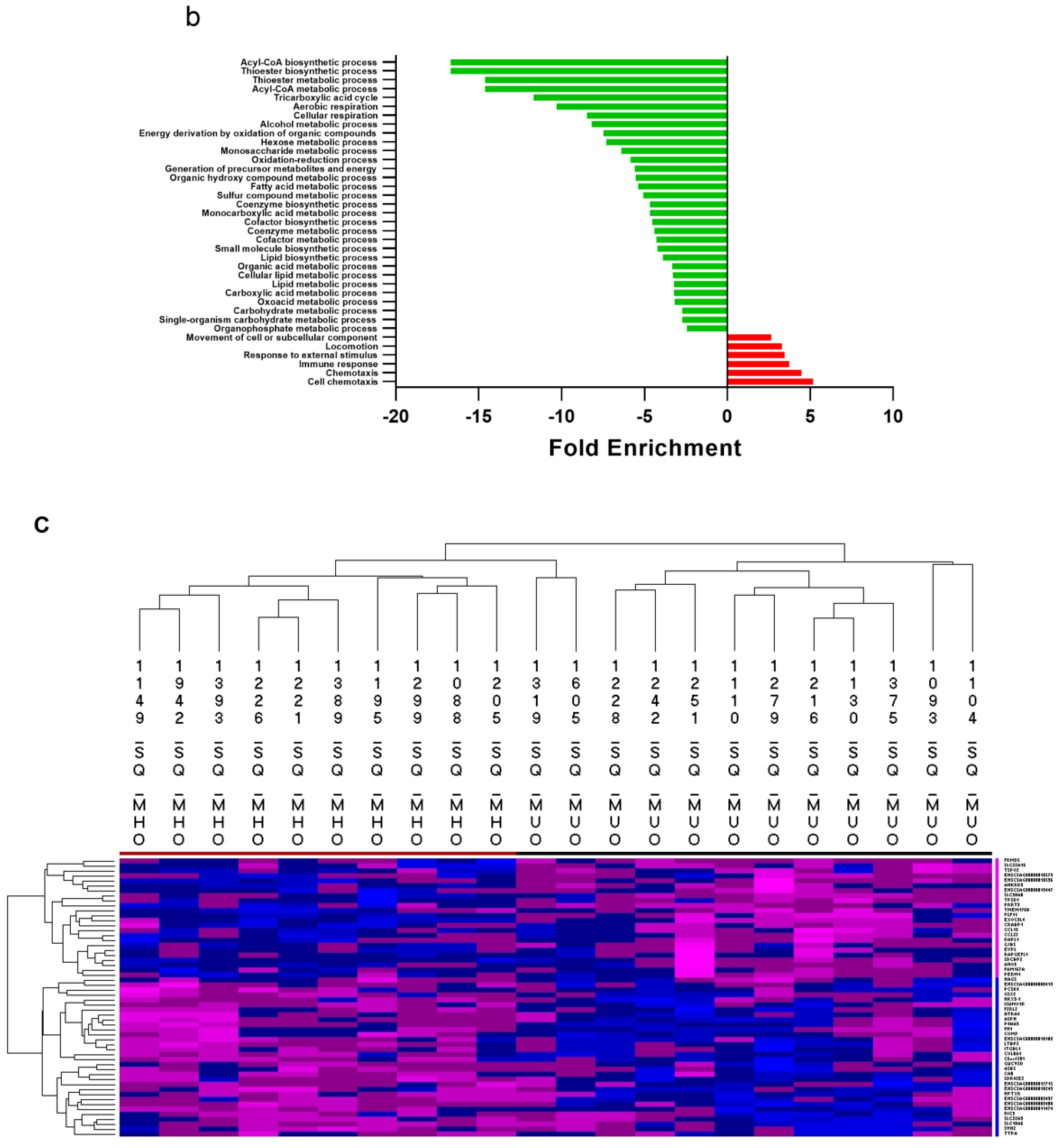
2.3. Metabolically Healthy Obese Subcutaneous Adipose Demonstrates Upregulation of Lipid Handling-Related Transcripts without Immune Activation
2.4. Tissue Cytokine Burdens Match Tissue Macrophage Profiles
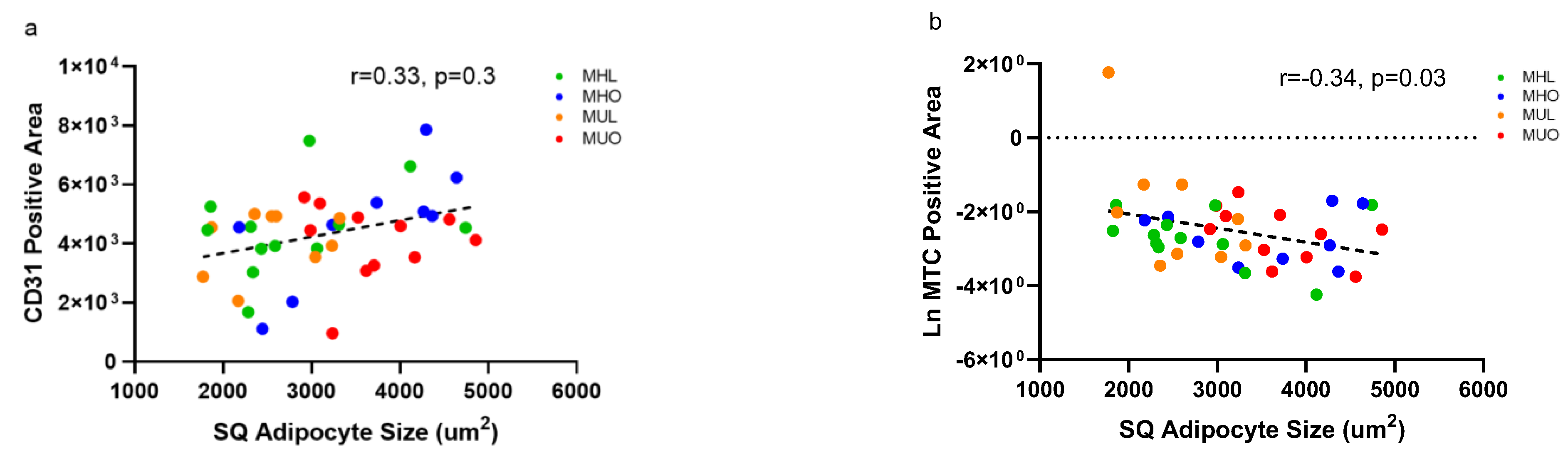
2.5. Changes in Tissue Vasculature or Fibrosis Do Not Explain Poor Health but Relate to Adipocyte Hypertrophy
3. Discussion
4. Methods and Materials
4.1. Study Design and Cohort Selection
4.2. Blood and Metabolic Measures
4.3. Body Composition
4.4. Tissue Collections
4.5. Cytokine Evaluations
4.6. Histology and Immunohistochemistry
4.7. Transcriptomic Analyses
4.8. Statistical Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 18 October 2022).
- Aung, K.; Lorenzo, C.; Hinojosa, M.A.; Haffner, S.M. Risk of developing diabetes and cardiovascular disease in metabolically unhealthy normal-weight and metabolically healthy obese individuals. J. Clin. Endocrinol. Metab. 2014, 99, 462–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahba, I.M.; Mak, R.H. Obesity and obesity-initiated metabolic syndrome: Mechanistic links to chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2007, 2, 550–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Russo, L.; Lumeng, C.N. Properties and functions of adipose tissue macrophages in obesity. Immunology 2018, 155, 407–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chylikova, J.; Dvorackova, J.; Tauber, Z.; Kamarad, V. M1/M2 macrophage polarization in human obese adipose tissue. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech. Repub. 2018, 162, 79–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Smith, W.; Hao, D.; He, B.; Kong, L. M1 and M2 macrophage polarization and potentially therapeutic naturally occurring compounds. Int. Immunopharmacol. 2019, 70, 459–466. [Google Scholar] [CrossRef]
- Wentworth, J.M.; Naselli, G.; Brown, W.A.; Doyle, L.; Phipson, B.; Smyth, G.K.; Wabitsch, M.; O’Brien, P.E.; Harrison, L.C. Pro-inflammatory CD11c+ CD206+ adipose tissue macrophages are associated with insulin resistance in human obesity. Diabetes 2010, 59, 1648–1656. [Google Scholar] [CrossRef] [Green Version]
- Wynn, T.A.; Vannella, K.M. Macrophages in tissue repair, regeneration, and fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef] [Green Version]
- Brestoff, J.R.; Wilen, C.B.; Moley, J.R.; Li, Y.; Zou, W.; Malvin, N.P.; Rowen, M.N.; Saunders, B.T.; Ma, H.; Mack, M.R. Intercellular mitochondria transfer to macrophages regulates white adipose tissue homeostasis and is impaired in obesity. Cell Metab. 2021, 33, 270–282.e278. [Google Scholar] [CrossRef]
- Nguyen, K.D.; Qiu, Y.; Cui, X.; Goh, Y.; Mwangi, J.; David, T.; Mukundan, L.; Brombacher, F.; Locksley, R.M.; Chawla, A. Alternatively activated macrophages produce catecholamines to sustain adaptive thermogenesis. Nature 2011, 480, 104–108. [Google Scholar] [CrossRef]
- Gao, J.; Qiao, Z.; Liu, S.; Xu, J.; Wang, S.; Yang, X.; Wang, X.; Tang, R. A small molecule nanodrug consisting of pH-sensitive ortho ester–dasatinib conjugate for cancer therapy. Eur. J. Pharm. Biopharm. 2021, 163, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Wellen, K.E.; Hotamisligil, G.S. Obesity-induced inflammatory changes in adipose tissue. J. Clin. Investig. 2003, 112, 1785–1788. [Google Scholar] [CrossRef] [PubMed]
- Paczkowska-Abdulsalam, M.; Kretowski, A. Obesity, metabolic health and omics: Current status and future directions. World J. Diabetes 2021, 12, 420. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, K.; Davis, A.T.; Peters, D.E.; LeGrand, A.C.; Bharadwaj, M.S.; Molina, A.J. Regulators of mitochondrial quality control differ in subcutaneous fat of metabolically healthy and unhealthy obese monkeys. Obesity 2017, 25, 689–696. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Härtlova, A.; Gierliński, M.; Prescott, A.; Castellvi, J.; Losa, J.H.; Petersen, S.K.; Wenzel, U.A.; Dill, B.D.; Emmerich, C.H. Triggering MSR1 promotes JNK-mediated inflammation in IL-4-activated macrophages. EMBO J. 2019, 38, e100299. [Google Scholar] [CrossRef]
- De Oliveira, C.; Khatua, B.; Noel, P.; Kostenko, S.; Bag, A.; Balakrishnan, B.; Patel, K.S.; Guerra, A.A.; Martinez, M.N.; Trivedi, S. Pancreatic triglyceride lipase mediates lipotoxic systemic inflammation. J. Clin. Investig. 2020, 130, 1931–1947. [Google Scholar] [CrossRef] [Green Version]
- Mehta, J.L.; Chen, J.; Hermonat, P.L.; Romeo, F.; Novelli, G. Lectin-like, oxidized low-density lipoprotein receptor-1 (LOX-1): A critical player in the development of atherosclerosis and related disorders. Cardiovasc. Res. 2006, 69, 36–45. [Google Scholar] [CrossRef]
- Das, S.K.; Ma, L.; Sharma, N.K. Adipose tissue gene expression and metabolic health of obese adults. Int. J. Obes. 2015, 39, 869–873. [Google Scholar] [CrossRef] [Green Version]
- Crewe, C.; An, Y.A.; Scherer, P.E. The ominous triad of adipose tissue dysfunction: Inflammation, fibrosis, and impaired angiogenesis. J. Clin. Investig. 2017, 127, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Naukkarinen, J.; Heinonen, S.; Hakkarainen, A.; Lundbom, J.; Vuolteenaho, K.; Saarinen, L.; Hautaniemi, S.; Rodriguez, A.; Frühbeck, G.; Pajunen, P. Characterising metabolically healthy obesity in weight-discordant monozygotic twins. Diabetologia 2014, 57, 167–176. [Google Scholar] [CrossRef]
- Schlauch, K.A.; Kulick, D.; Subramanian, K.; De Meirleir, K.L.; Palotás, A.; Lombardi, V.C. Single-nucleotide polymorphisms in a cohort of significantly obese women without cardiometabolic diseases. Int. J. Obes. 2019, 43, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Gharooi Ahangar, O.; Javanrouh, N.; Daneshpour, M.S.; Barzin, M.; Valizadeh, M.; Azizi, F.; Hosseinpanah, F. Genetic markers and continuity of healthy metabolic status: Tehran cardio-metabolic genetic study (TCGS). Sci. Rep. 2020, 10, 1–8. [Google Scholar]
- Fujisaka, S. The role of adipose tissue M1/M2 macrophages in type 2 diabetes mellitus. Diabetol. Int. 2021, 12, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Miyachi, Y.; Tsuchiya, K.; Shiba, K.; Mori, K.; Komiya, C.; Ogasawara, N.; Ogawa, Y. A reduced M1-like/M2-like ratio of macrophages in healthy adipose tissue expansion during SGLT2 inhibition. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bäckdahl, J.; Franzén, L.; Massier, L.; Li, Q.; Jalkanen, J.; Gao, H.; Andersson, A.; Bhalla, N.; Thorell, A.; Rydén, M. Spatial mapping reveals human adipocyte subpopulations with distinct sensitivities to insulin. Cell Metab. 2021, 33, 1869–1882.e1866. [Google Scholar] [CrossRef]
- Kavanagh, K.; Jones, K.L.; Sawyer, J.; Kelley, K.; Carr, J.J.; Wagner, J.D.; Rudel, L.L. Trans fat diet induces abdominal obesity and changes in insulin sensitivity in monkeys. Obesity 2007, 15, 1675–1684. [Google Scholar] [CrossRef]
- Kavanagh, K.; Wylie, A.T.; Tucker, K.L.; Hamp, T.J.; Gharaibeh, R.Z.; Fodor, A.A.; Cullen, J.M. Dietary fructose induces endotoxemia and hepatic injury in calorically controlled primates. Am. J. Clin. Nutr. 2013, 98, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Nappo, F.; Esposito, K.; Cioffi, M.; Giugliano, G.; Molinari, A.M.; Paolisso, G.; Marfella, R.; Giugliano, D. Postprandial endothelial activation in healthy subjects and in type 2 diabetic patients: Role of fat and carbohydrate meals. J. Am. Coll. Cardiol. 2002, 39, 1145–1150. [Google Scholar] [CrossRef] [Green Version]
- Spencer, M.; Finlin, B.S.; Unal, R.; Zhu, B.; Morris, A.J.; Shipp, L.R.; Lee, J.; Walton, R.G.; Adu, A.; Erfani, R. Omega-3 fatty acids reduce adipose tissue macrophages in human subjects with insulin resistance. Diabetes 2013, 62, 1709–1717. [Google Scholar] [CrossRef] [Green Version]
- Brykczynska, U.; Geigges, M.; Wiedemann, S.J.; Dror, E.; Böni-Schnetzler, M.; Hess, C.; Donath, M.Y.; Paro, R. Distinct transcriptional responses across tissue-resident macrophages to short-term and long-term metabolic challenge. Cell Rep. 2020, 30, 1627–1643.e1627. [Google Scholar] [CrossRef]
- Fox, C.S.; Massaro, J.M.; Hoffmann, U.; Pou, K.M.; Maurovich-Horvat, P.; Liu, C.-Y.; Vasan, R.S.; Murabito, J.M.; Meigs, J.B.; Cupples, L.A. Abdominal visceral and subcutaneous adipose tissue compartments: Association with metabolic risk factors in the Framingham Heart Study. Circulation 2007, 116, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.A.; Hamer, M.; Batty, G.D.; Singh-Manoux, A.; Sabia, S.; Kivimäki, M. Incidence of metabolic risk factors among healthy obese adults: 20-year follow-up. J. Am. Coll. Cardiol. 2015, 66, 871–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolahdouzi, S.; Talebi-Garakani, E.; Hamidian, G.; Safarzade, A. Exercise training prevents high-fat diet-induced adipose tissue remodeling by promoting capillary density and macrophage polarization. Life Sci. 2019, 220, 32–43. [Google Scholar] [CrossRef]
- Heo, J.S.; Choi, Y.; Kim, H.O. Adipose-derived mesenchymal stem cells promote M2 macrophage phenotype through exosomes. Stem Cells Int. 2019, 2019, 7921760. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Shang, Q.; Pan, Z.; Bai, Y.; Li, Z.; Zhang, H.; Zhang, Q.; Guo, C.; Zhang, L.; Wang, Q. Exosomes from adipose-derived stem cells attenuate adipose inflammation and obesity through polarizing M2 macrophages and beiging in white adipose tissue. Diabetes 2018, 67, 235–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrero, A.; Alegret, M.; Sánchez, R.M.; Adzet, T.; Laguna, J.C.; Vázquez, M. Bezafibrate reduces mRNA levels of adipocyte markers and increases fatty acid oxidation in primary culture of adipocytes. Diabetes 2001, 50, 1883–1890. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, B.; Long, L.; Luo, P.; Xiang, W.; Li, X.; Wang, H.; Jiang, Q.; Tan, X.; Luo, S. Improvement of obesity-associated disorders by a small-molecule drug targeting mitochondria of adipose tissue macrophages. Nat. Commun. 2021, 12, 1–16. [Google Scholar] [CrossRef]
- Camhi, S.M.; Must, A.; Gona, P.N.; Hankinson, A.; Odegaard, A.; Reis, J.; Gunderson, E.P.; Jacobs, D.R.; Carnethon, M.R. Duration and stability of metabolically healthy obesity over 30 years. Int. J. Obes. 2019, 43, 1803–1810. [Google Scholar] [CrossRef] [PubMed]
- Comuzzie, A.G.; Cole, S.A.; Martin, L.; Carey, K.D.; Mahaney, M.C.; Blangero, J.; VandeBerg, J.L. The baboon as a nonhuman primate model for the study of the genetics of obesity. Obes. Res. 2003, 11, 75–80. [Google Scholar] [CrossRef]
- Laparra, A.; Tricot, S.; Le Van, M.; Damouche, A.; Gorwood, J.; Vaslin, B.; Favier, B.; Benoist, S.; Ho Tsong Fang, R.; Bosquet, N. The frequencies of immunosuppressive cells in adipose tissue differ in human, non-human primate, and mouse models. Front. Immunol. 2019, 10, 117. [Google Scholar] [CrossRef]
- Bousquet-Mélou, A.; Galitzky, J.; Lafontan, M.; Berlan, M. Control of lipolysis in intra-abdominal fat cells of nonhuman primates: Comparison with humans. J. Lipid Res. 1995, 36, 451–461. [Google Scholar] [CrossRef]
- Ziaei, S.; Mohseni, H. Correlation between hormonal statuses and metabolic syndrome in postmenopausal women. J. Fam. Reprod. Health 2013, 7, 63. [Google Scholar]
- Misumi, I.; Starmer, J.; Uchimura, T.; Beck, M.A.; Magnuson, T.; Whitmire, J.K. Obesity expands a distinct population of T cells in adipose tissue and increases vulnerability to infection. Cell Rep. 2019, 27, 514–524.e515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Parekh, V.V.; Hsiao, J.; Kitamura, D.; Van Kaer, L. Spleen supports a pool of innate-like B cells in white adipose tissue that protects against obesity-associated insulin resistance. Proc. Natl. Acad. Sci. USA 2014, 111, E4638–E4647. [Google Scholar] [CrossRef] [Green Version]
- Morgan-Bathke, M.; Harteneck, D.; Jaeger, P.; Sondergaard, E.; Karwoski, R.; Espinosa De Ycaza, A.; Carranza-Leon, B.G.; Faubion, W.A., Jr.; Oliveira, A.M.; Jensen, M.D. Comparison of methods for analyzing human adipose tissue macrophage content. Obesity 2017, 25, 2100–2107. [Google Scholar] [CrossRef] [PubMed]
- Barros, M.H.M.; Hauck, F.; Dreyer, J.H.; Kempkes, B.; Niedobitek, G. Macrophage polarisation: An immunohistochemical approach for identifying M1 and M2 macrophages. PLoS ONE 2013, 8, e80908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leopold Wager, C.M.; Hole, C.R.; Wozniak, K.L.; Olszewski, M.A.; Mueller, M.; Wormley, F.L., Jr. STAT1 signaling within macrophages is required for antifungal activity against Cryptococcus neoformans. Infect. Immun. 2015, 83, 4513–4527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Tong, Z.; Ding, C.; Luo, F.; Wu, S.; Wu, C.; Albeituni, S.; He, L.; Hu, X.; Tieri, D. Transcription factor c-Maf is a checkpoint that programs macrophages in lung cancer. J. Clin. Investig. 2020, 130, 2081–2096. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, J.; Li, S.; Yu, Z.; Liu, B.; Song, B.; Su, Y. The clinical dynamic changes of macrophage phenotype and function in different stages of human wound healing and hypertrophic scar formation. Int. Wound J. 2019, 16, 360–369. [Google Scholar] [CrossRef]
- Xu, Z.-J.; Gu, Y.; Wang, C.-Z.; Jin, Y.; Wen, X.-M.; Ma, J.-C.; Tang, L.-J.; Mao, Z.-W.; Qian, J.; Lin, J. The M2 macrophage marker CD206: A novel prognostic indicator for acute myeloid leukemia. Oncoimmunology 2020, 9, 1683347. [Google Scholar] [CrossRef] [Green Version]
- Everaert, C.; Luypaert, M.; Maag, J.L.; Cheng, Q.X.; Dinger, M.E.; Hellemans, J.; Mestdagh, P. Benchmarking of RNA-sequencing analysis workflows using whole-transcriptome RT-qPCR expression data. Sci. Rep. 2017, 7, 1–11. [Google Scholar]
- Ruiz-Ojeda, F.J.; Méndez-Gutiérrez, A.; Aguilera, C.M.; Plaza-Díaz, J. Extracellular matrix remodeling of adipose tissue in obesity and metabolic diseases. Int. J. Mol. Sci. 2019, 20, 4888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model. Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef]
- Kavanagh, K.; Fairbanks, L.A.; Bailey, J.N.; Jorgensen, M.J.; Wilson, M.; Zhang, L.; Rudel, L.L.; Wagner, J.D. Characterization and heritability of obesity and associated risk factors in vervet monkeys. Obesity 2007, 15, 1666–1674. [Google Scholar] [CrossRef]
- Araújo, J.; Cai, J.; Stevens, J. Prevalence of optimal metabolic health in American adults: National Health and Nutrition Examination Survey 2009–2016. Metab. Syndr. Relat. Disord. 2019, 17, 46–52. [Google Scholar] [CrossRef]
- Moore, J.X.; Chaudhary, N.; Akinyemiju, T. Peer reviewed: Metabolic syndrome prevalence by race/ethnicity and sex in the United States, National Health and Nutrition Examination Survey, 1988–2012. Prev. Chronic Dis. 2017, 14, E24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Cholesterol Education Program (US); Expert Panel on Detection, Treatment of High Blood Cholesterol in Adults. Third Report of the National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III); The Program: Bethesda, MD, USA, 2002. [Google Scholar]
- Lean, M.; Han, T.; Morrison, C. Waist circumference as a measure for indicating need for weight management. BMJ 1995, 311, 158–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palaniappan, L.; Carnethon, M.R.; Wang, Y.; Hanley, A.J.; Fortmann, S.P.; Haffner, S.M.; Wagenknecht, L. Predictors of the incident metabolic syndrome in adults: The Insulin Resistance Atherosclerosis Study. Diabetes Care 2004, 27, 788–793. [Google Scholar] [CrossRef] [Green Version]
- Vemuri, R.; Ruggiero, A.; Whitfield, J.B.; Dugan, G.O.; Cline, J.M.; Block, M.R.; Guo, H.; Kavanagh, K. Hypertension drives microbial translocation and shifts in the fecal microbiome of non-human primates. bioRxiv 2021. [Google Scholar] [CrossRef]
- Bacarella, N.; Ruggiero, A.; Davis, A.T.; Uberseder, B.; Davis, M.A.; Bracy, D.P.; Wasserman, D.H.; Cline, J.M.; Sherrill, C.; Kavanagh, K. Whole body irradiation induces diabetes and adipose insulin resistance in nonhuman primates. Int. J. Radiat. Oncol. Biol. Phys. 2020, 106, 878–886. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement | MHL | MHO | MUL | MUO | ANOVA p-Value | Health p-Value | Obesity p-Value |
|---|---|---|---|---|---|---|---|
| Age (yrs) | 15.15 (1.30) | 15.14 (1.55) | 14.32 (1.99) | 15.87 (0.92) | 0.74 | 0.69 | 0.26 |
| Body Weight (kg) | 5.25 (0.26) | 6.47 (0.24) | 4.95 (0.24) | 6.91 (0.26) | <0.001 | 0.19 | 0.01 |
| Waist Circumference (cm) | 34.29 (0.97) | 42.86 (1.07) | 34.80 (0.78) | 43.23 (0.85) | <0.001 | 0.02 | <0.001 |
| Fasting Glucose (mg/dL) | 73.5 (3.20) | 67.2 (3.59) | 108.67 (19.15) | 104.32 (12.37) | 0.01 | 0.36 | 0.86 |
| Glycosylated Hemoglobin A1c (%) | 4.07 (0.05) | 4.07 (0.07) | 4.83 (0.58) | 5.38 (0.57) | 0.009 | 0.012 | 0.31 |
| Systolic Blood Pressure (mmHg) | 114.44 (3.79) | 110.10 (5.43) | 144.78 (4.95) | 111.62 (5.06) | <0.001 | 0.48 | 0.31 |
| Diastolic Blood Pressure (mmHg) | 67.53 (3.38) | 61.57 (3.80) | 89.22 (4.70) | 66.79 (2.46) | <0.001 | 0.25 | 0.42 |
| High Density Lipoprotein Cholesterol (mg/dL) | 68.08 (3.37) | 56.10 (5.76) | 73.44 (10.38) | 62.50 (4.39) | 0.24 | 0.33 | 0.06 |
| Triglycerides (mg/dL) | 56.34 (3.46) | 55.90 (5.89) | 79.10 (15.67) | 90.52 (10.80) | 0.03 | 0.24 | 0.76 |
| Liver Density (HU) | 60.37 (2.26) | 55.71 (4.03) | 59.54 (2.84) | 57.59 (1.94) | 0.86 | 0.69 | 0.83 |
| % Fat Mass | 18.35 (2.69) | 36.59 (2.81) | 16.70 (2.07) | 35.98 (3.05) | 0.01 | 0.94 | 0.06 |
| % Lean Mass | 54.13 (1.56) | 47.29 (2.89) | 55.03 (1.52) | 45.63 (1.60) | 0.02 | 0.91 | 0.07 |
| Visceral:Subcutaneous Adipose Area (cm2) | 3.44 (0.46) | 1.78 (0.19) | 3.86 (0.83) | 1.73 (0.30) | 0.31 | 0.88 | 0.19 |
| Plasma Insulin (mU/L) | 16.80 (4.15) | 19.98 (3.87) | 8.84 (2.08) | 51.12 (11.18) | 0.0055 | 0.71 | 0.0019 |
| Transcript | Log Fold Change | Adjusted p-Value |
|---|---|---|
| Fatty Acid Binding Protein 7 (FABP7) | 1.51 | 0.010 |
| Prostaglandin D2 Synthase (PTGDS) | 1.53 | 0.015 |
| Aldehyde Dehydrogenase 1 Family Member B1 (ALDH1B1) | 1.56 | 0.044 |
| Transferrin (TF) | 1.66 | 0.024 |
| Thyroid Stimulating hormone receptor (TSHR) | 1.70 | 0.0036 |
| Lactate Dehydrogenase D (LDHD) | 1.74 | 0.00016 |
| Phosphoenolpyruvate Carboxykinase 2 (PCK2) | 1.77 | 0.0063 |
| ATPase Phospholipid Transporting 8B3 (ATP8B3) | 1.79 | 0.0063 |
| Adrenoceptor Beta 3 (ADRB3) | 1.79 | 0.010 |
| Trefoil Factor 3 (TFF3) | 1.88 | 0.027 |
| ATP Binding Cassette Subfamily C Member 6 (ABCC6) | 1.89 | 0.0051 |
| Phosphoenolpyruvate Carboxykinase 1 (PCK1) | 2.05 | 0.0075 |
| Monoacylglycerol O-Acyltransferase 1 (MOGAT1) | 2.12 | 0.00071 |
| Glycerol-3-Phosphate Acyltransferase 3 (GPAT3) | 2.16 | 0.015 |
| Protein Phosphatase 1 Regulatory Inhibitor Subunit 1B (PPP1R1B) | 2.38 | 0.034 |
| ADAM-Like Decysin 1 (ADAMDEC1) | −4.11 | 0.00011 |
| Cartilage Oligomeric Matrix Protein (COMP) | −3.13 | 0.00022 |
| Sterile Alpha Motif Domain Containing 5 (SAMD5) | −2.98 | 0.000056 |
| Dendrocyte Expressed Seven Transmembrane Protein (DCSTAMP) | −2.93 | 0.014 |
| R-Spondin 1 (RSPO1) | −2.76 | 0.0034 |
| Thymic Stromal Lymphopoietin (TSLP) | −2.52 | 0.00067 |
| Osteoclast Stimulatory Transmembrane Protein (OCSTAMP) | −2.46 | 0.045 |
| CUB and Sushi Multiple Domains 2 (CSMD2) | −2.31 | 0.011 |
| ETS Homologous Factor (EHF) | −2.28 | 0.027 |
| Oxidized Low Density Lipoprotein Receptor 1 (OLR1) | −2.23 | 0.0063 |
| Carboxypeptidase Z (CPZ) | −2.20 | 0.029 |
| C-C Motif Chemokine Receptor 1 (CCR1) | −2.17 | 0.00070 |
| Serpin Family A Member 5 (SERPINA5) | −2.11 | 0.0020 |
| Pancreatic Lipase Related Protein 1 (PNLIPRP1) | −2.10 | 0.020 |
| Macrophage Scavenger Receptor 1 (MSR1) | −2.08 | 0.014 |
| Transcript | Log Fold Change | Adjusted p-Value |
|---|---|---|
| Glycogen Synthase 2 (GYS2) | 3.72 | 0.0059 |
| Collagen Type IV Alpha 3 Chain (COL4A3) | 2.69 | 0.0084 |
| Family with Sequence Similarity 131 Member C (FAM131C) | 2.62 | 0.0067 |
| Serum Amyloid A4, Constitutive (SAA4) | 2.45 | 0.028 |
| SH3 Domain Binding Kinase Family Member 2 (SBK2) | 2.41 | 0.0011 |
| Glycine-N-Acyltransferase (GLYAT) | 2.40 | 0.00033 |
| Plasminogen (PLG) | 2.33 | 0.000012 |
| Adrenoceptor Beta 3 (ADRB3) | 2.28 | 0.0019 |
| Stathmin 4 (STMN4) | 2.23 | 0.0456 |
| C-type Lectin Domain Family 4 Member G (CLEC4G) | 2.22 | 0.0060 |
| Thyroid Stimulating Hormone Receptor (TSHR) | 2.21 | 0.000049 |
| Monoacylglycerol O-Acyltransferase 1 (MOGAT1) | 2.19 | 0.00013 |
| Polypeptide N-Acetylgalactosaminyltransferase 9 (GALNT9) | 2.17 | 0.0016 |
| V-Set Furthermore, Transmembrane Domain Containing 2 Like (VSTM2L) | 2.17 | 0.0043 |
| Cat Eye Syndrome Chromosome Region, Candidate 2(CECR2) | 2.11 | 0.0017 |
| Glycoprotein Hormones, Alpha Polypeptide (CGA) | −2.62 | 0.0054 |
| Osteoclast Stimulatory Transmembrane Protein (OCSTAMP) | −2.63 | 0.013 |
| WAP Four-Disulfide Core Domain 2 (WFDC2) | −2.67 | 0.00025 |
| Solute Carrier Family 5 Member 9 (SLC5A9) | −2.68 | 0.00038 |
| Glutamate Ionotropic Receptor AMPA Type Subunit 1 (GRIA1) | −2.91 | 0.000011 |
| Cadherin 1 (CDH1) | −2.98 | 0.0047 |
| Sterile Alpha Motif Domain Containing 5 (SAMD5) | −2.99 | 0.000010 |
| ETS Homologous Factor (EHF) | −3.00 | 0.00013 |
| Dendrocyte Expressed Seven Transmembrane Protein (DCSTAMP) | −3.02 | 0.0040 |
| WD Repeat Domain 64 (WDR64) | −3.12 | 0.0000085 |
| Matrix Metallopeptidase 7 (MMP7) | −3.29 | 0.035 |
| Solute Carrier Family 24 Member 2 (SLC24A2) | −3.32 | 0.00021 |
| S100 Calcium Binding Protein A7A (S100A7A) | −3.32 | 0.00040 |
| ADAM Like Decysin 1 (ADAMDEC1) | −4.06 | 0.000028 |
| Death Associated Protein Like 1 (DAPL1) | −4.27 | 0.000056 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruggiero, A.D.; Vemuri, R.; Block, M.; DeStephanis, D.; Davis, M.; Chou, J.; Williams, A.; Brock, A.; Das, S.K.; Kavanagh, K. Macrophage Phenotypes and Gene Expression Patterns Are Unique in Naturally Occurring Metabolically Healthy Obesity. Int. J. Mol. Sci. 2022, 23, 12680. https://doi.org/10.3390/ijms232012680
Ruggiero AD, Vemuri R, Block M, DeStephanis D, Davis M, Chou J, Williams A, Brock A, Das SK, Kavanagh K. Macrophage Phenotypes and Gene Expression Patterns Are Unique in Naturally Occurring Metabolically Healthy Obesity. International Journal of Molecular Sciences. 2022; 23(20):12680. https://doi.org/10.3390/ijms232012680
Chicago/Turabian StyleRuggiero, Alistaire D., Ravichandra Vemuri, Masha Block, Darla DeStephanis, Matthew Davis, Jeff Chou, Abigail Williams, Ashlynn Brock, Swapan Kumar Das, and Kylie Kavanagh. 2022. "Macrophage Phenotypes and Gene Expression Patterns Are Unique in Naturally Occurring Metabolically Healthy Obesity" International Journal of Molecular Sciences 23, no. 20: 12680. https://doi.org/10.3390/ijms232012680
APA StyleRuggiero, A. D., Vemuri, R., Block, M., DeStephanis, D., Davis, M., Chou, J., Williams, A., Brock, A., Das, S. K., & Kavanagh, K. (2022). Macrophage Phenotypes and Gene Expression Patterns Are Unique in Naturally Occurring Metabolically Healthy Obesity. International Journal of Molecular Sciences, 23(20), 12680. https://doi.org/10.3390/ijms232012680