

Isomelezitose Overproduction by Alginate-Entrapped Recombinant E. coli Cells and In Vitro Evaluation of Its Potential Prebiotic Effect

 ,
,  ,
, 
 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
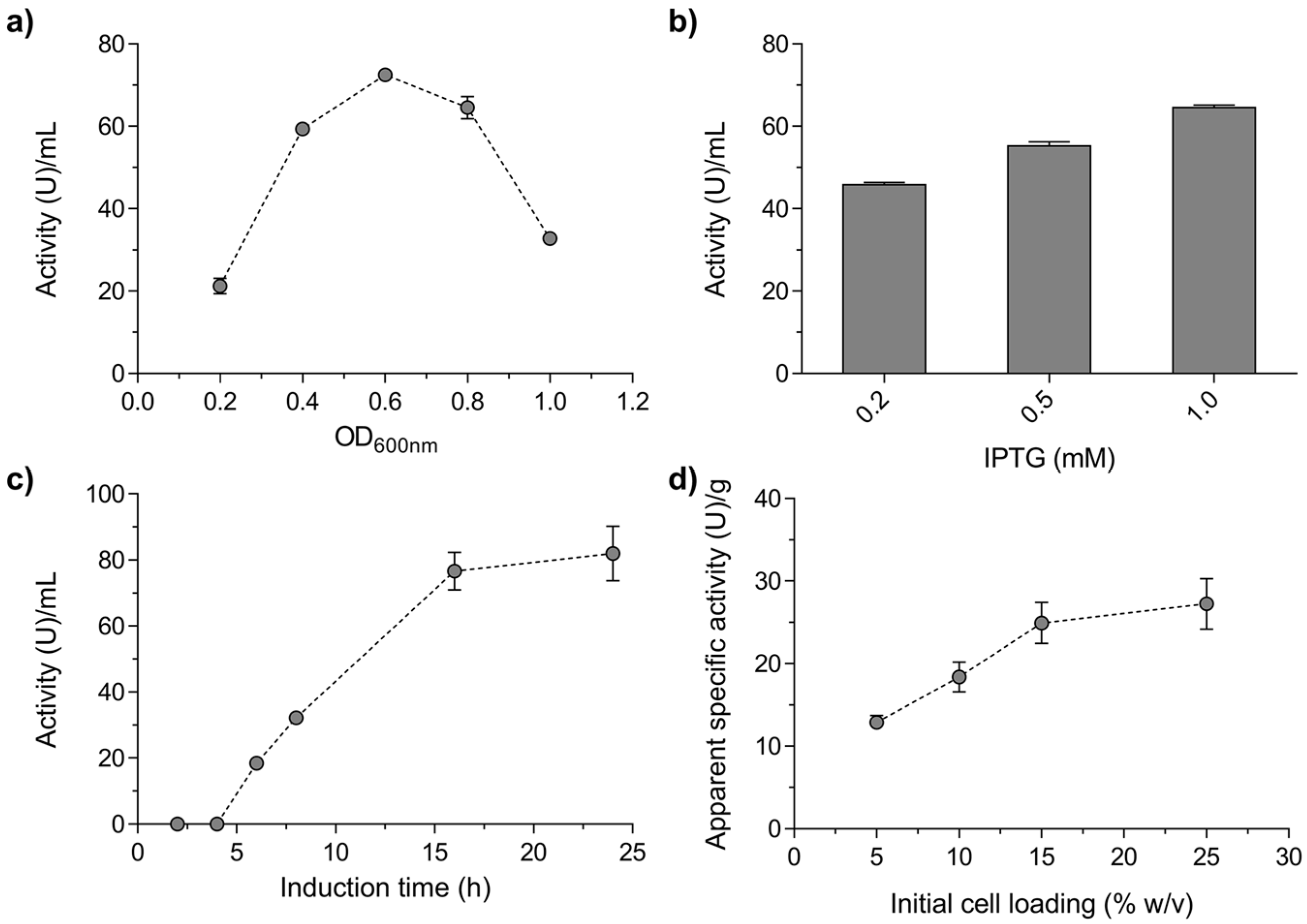
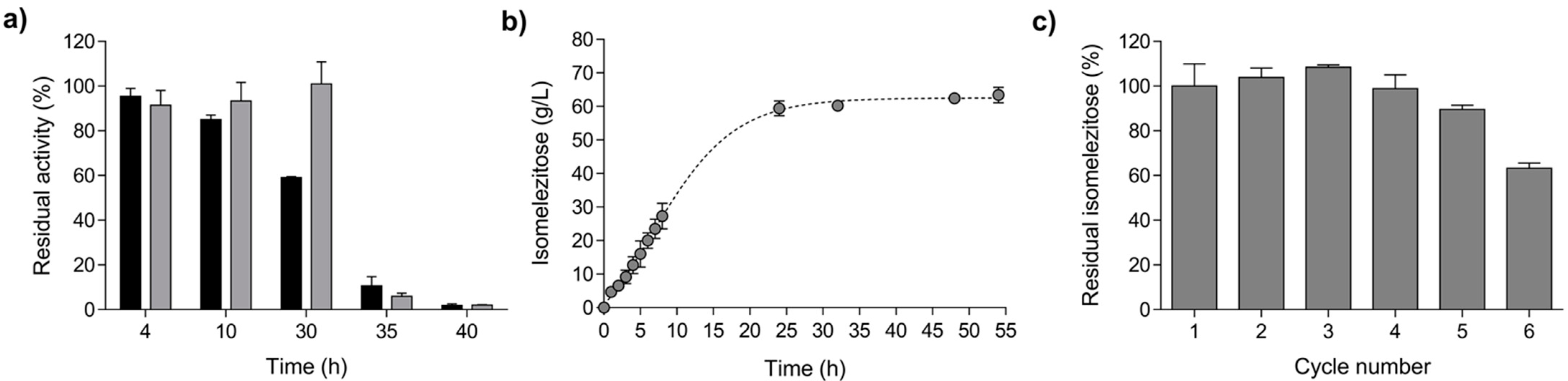
2.1. Production and Characterisation of a Biocatalyst Based on Alginate-Entrapped E. coli Cells
2.2. Isomelezitose Overproduction Employing the Immobilised E. coli Cells in Alginate Beads
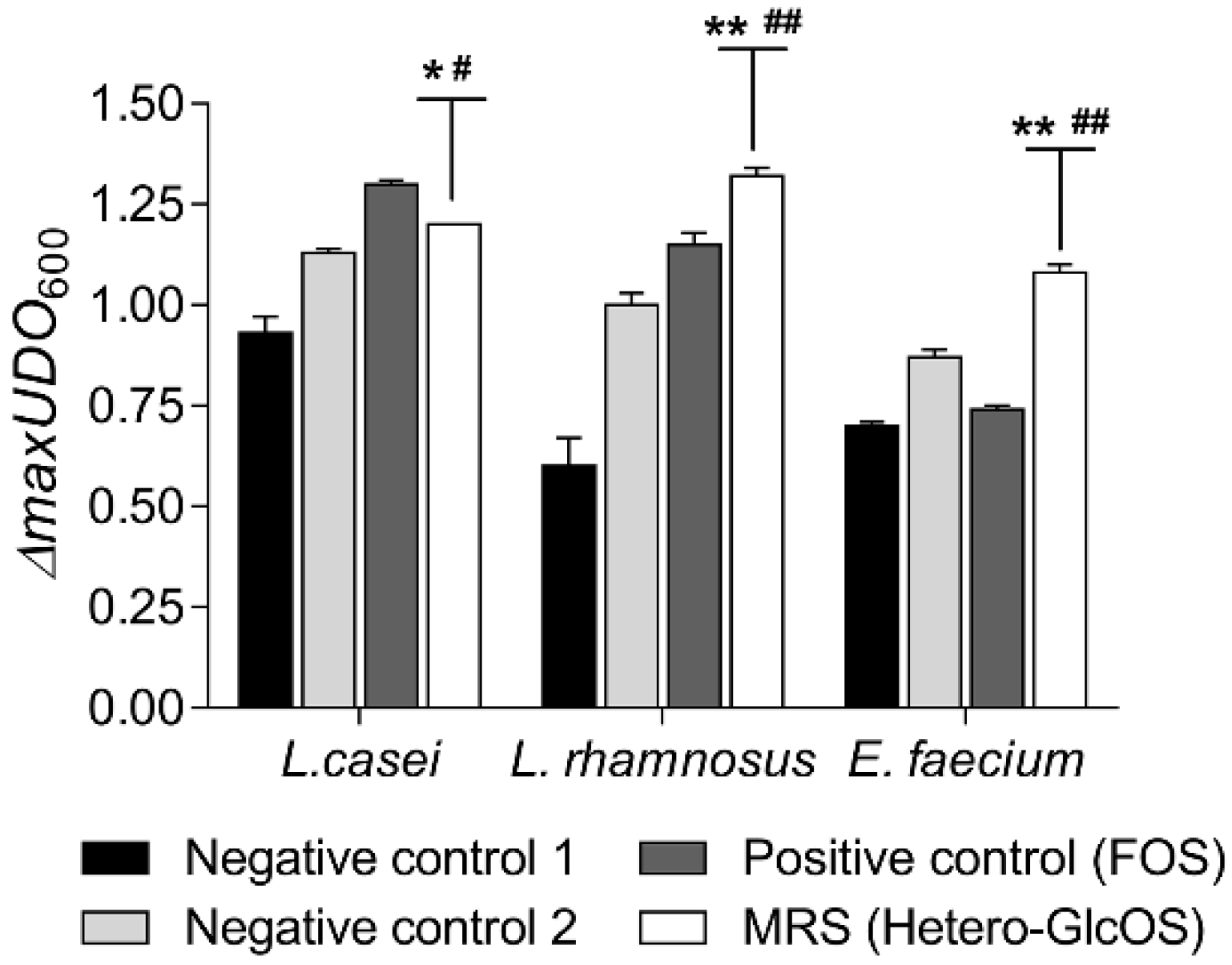
2.3. Potential Prebiotic Effect of the Hetero-GlcOS Generated by the Alginate-Entrapped E. coli Cells Expressing Mr-αGlu
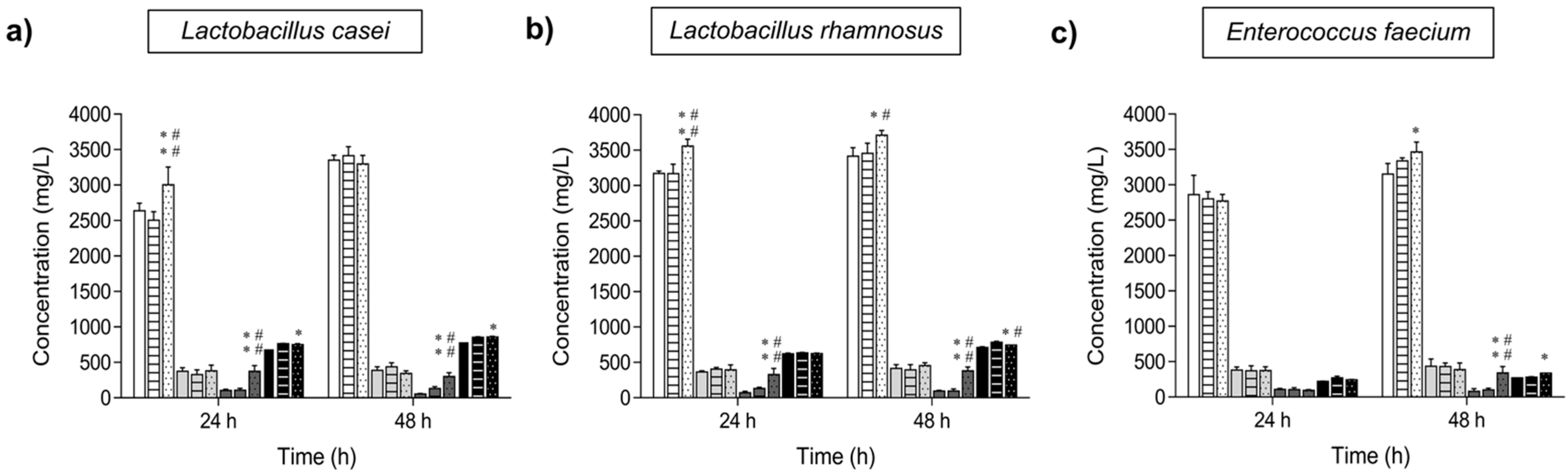
2.4. Analysis of the Potential Prebiotic Activity of Isomelezitose
3. Materials and Methods
3.1. Materials and Reagents
3.2. Microorganisms and Culture Conditions
3.3. Optimal Conditions for Mr-αGlu Expression in E. coli
3.4. Standard Hydrolytic Activity Assay
3.5. Entrapment of E. coli Cells Harbouring the Mr-αGlu Activity into Alginate Beads
3.6. Thermal and pH Stability of the Alginate-Entrapped E. coli Cells
3.7. Production of Honey Hetero-GlcOS and Operational Stability of Immobilised Biocatalysts in Transglucosylation Reactions
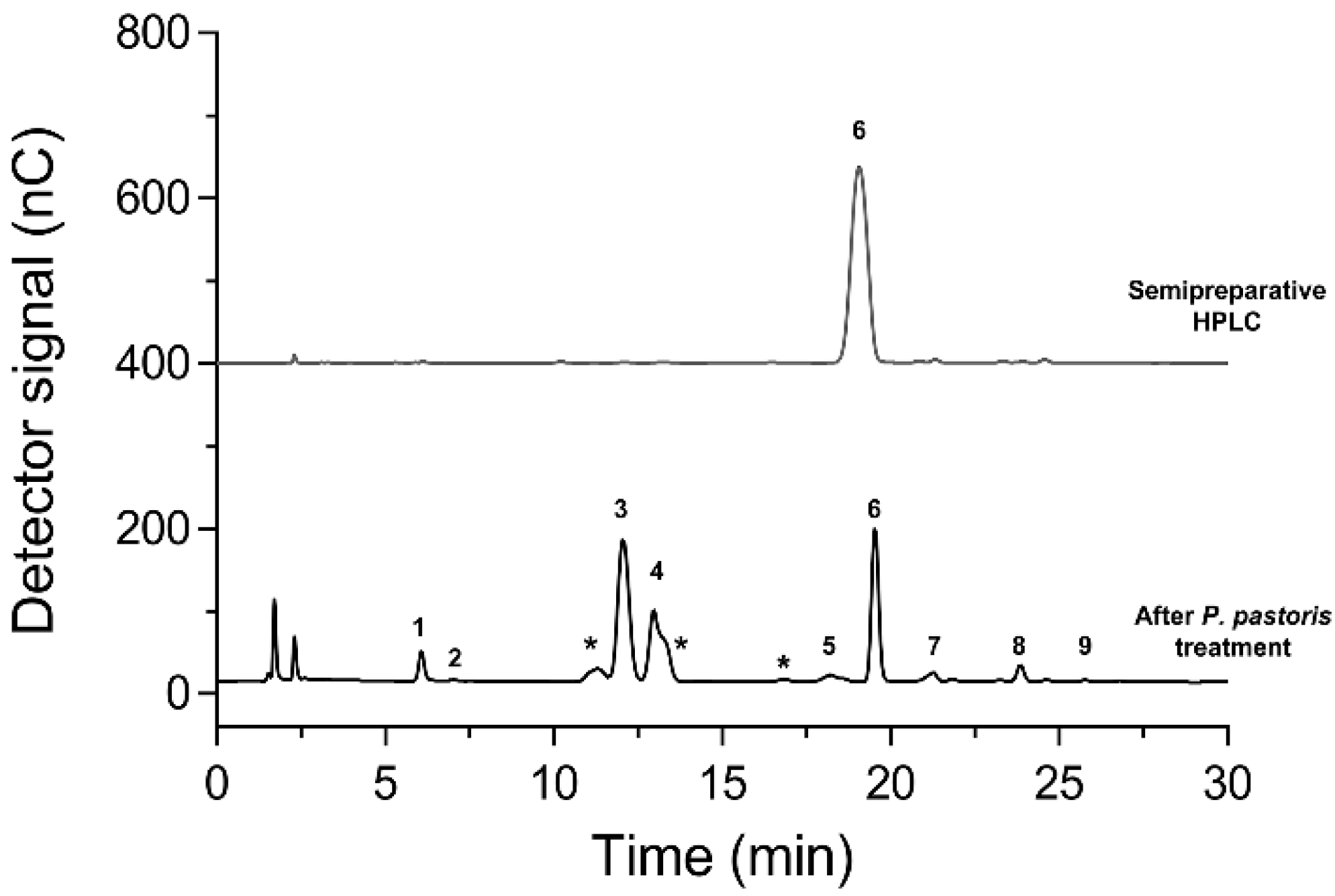
3.8. Partial Purification of Hetero-GlcOS Mixture by Alginate-Entrapped K. phaffii and Isomelezitose Purification
3.9. Effect of Isomelezitose and Commercial Prebiotic Sugars on the Grow and Metabolic Activity of Probiotic Bacteria
3.10. Analytical Analysis
3.11. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salamone, D.; Rivellese, A.A.; Vetrani, C. The Relationship between Gut Microbiota, Short-Chain Fatty Acids and Type 2 Diabetes Mellitus: The Possible Role of Dietary Fibre. Acta Diabetol. 2021, 58, 1131–1138. [Google Scholar] [CrossRef]
- Seo, K.; Seo, J.; Yeun, J.; Choi, H.; Kim, Y.I.; Chang, S.Y. The Role of Mucosal Barriers in Human Gut Health. Arch. Pharm. Res. 2021, 44, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Whon, T.W.; Lim, M.Y.; Kim, Y.B.; Kim, N.; Kwon, M.; Kim, J.; Lee, S.H.; Choi, H.; Nam, I.; et al. The Human Gut Archaeome: Identification of Diverse Haloarchaea in Korean Subjects. Microbiome 2020, 8, 114. [Google Scholar] [CrossRef] [PubMed]
- Date, Y.; Nakanishi, Y.; Fukuda, S.; Nuijima, Y.; Kato, T.; Umehara, M.; Ohno, H.; Kikuchi, J. In Vitro Evaluation Method for Screening of Candidate Prebiotic Foods. Food Chem. 2014, 152, 251–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinto, E.J.; Jiménez, P.; Caro, I.; Tejero, J.; Mateo, J.; Girbés, T. Probiotic Lactic Acid Bacteria: A Review. Food Nutr. Sci. 2014, 5, 1765–1775. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Liu, J.; Ling, Z. Short-Chain Fatty Acids-Producing Probiotics: A Novel Source of Psychobiotics. Crit. Rev. Food Sci. Nutr. 2021, 62, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Zalán, Z.; Hudáček, J.; Štětina, J.; Chumchalová, J.; Halász, A. Production of Organic Acids by Lactobacillus Strains in Three Different Media. Eur. Food Res. Technol. 2010, 230, 395–404. [Google Scholar] [CrossRef]
- Singh, S.P.; Jadaun, J.S.; Narnoliya, L.K.; Pandey, A. Prebiotic Oligosaccharides: Special Focus on Fructooligosaccharides, Its Biosynthesis and Bioactivity. Appl. Biochem. Biotechnol. 2017, 183, 613–635. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohanty, D.; Misra, S.; Mohapatra, S.; Sahu, P.S. Prebiotics and Symbiotics: Recent Concepts in Nutrition. Food Biosci. 2018, 26, 152–160. [Google Scholar] [CrossRef]
- Lee, S.; Park, J.; Jang, J.K.; Lee, B.H.; Park, Y.S. Structural Analysis of Gluco-Oligosaccharides Produced by Leuconostoc lactis and Their Prebiotic Effect. Molecules 2019, 24, 3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Gonzalez, M.; Minguet-Lobato, M.; Plou, F.J.; Fernandez-Lobato, M. Molecular Characterization and Heterologous Expression of Two α-Glucosidases from Metschnikowia Spp., Both Producers of Honey Sugars. Microb. Cell Fact. 2020, 19, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Suarez, J.; Giampieri, F.; Battino, M. Honey as a Source of Dietary Antioxidants: Structures, Bioavailability and Evidence of Protective Effects Against Human Chronic Diseases. Curr. Med. Chem. 2013, 20, 621–638. [Google Scholar] [CrossRef]
- Pascual-Maté, A.; Osés, S.M.; Marcazzan, G.L.; Gardini, S.; Fernández Muiño, M.A.; Teresa Sancho, M. Sugar Composition and Sugar-Related Parameters of Honeys from the Northern Iberian Plateau. J. Food Compos. Anal. 2018, 74, 34–43. [Google Scholar] [CrossRef]
- Mohan, A.; Quek, S.-Y.; Gutierrez-Maddox, N.; Gao, Y.; Shu, Q. Effect of Honey in Improving the Gut Microbial Balance. Food Qual. Saf. 2017, 1, 107–115. [Google Scholar] [CrossRef]
- Siddiqui, A.J.; Musharraf, S.G.; Choudhary, M.I.; Rahman, A. Application of Analytical Methods in Authentication and Adulteration of Honey. Food Chem. 2017, 217, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.L.; Polemis, N.; Morales, V.; Corzo, N.; Drakoularakou, A.; Gibson, G.R.; Rastall, R.A. In Vitro Investigation into the Potential Prebiotic Activity of Honey Oligosaccharides. J. Agric. Food Chem. 2005, 53, 2914–2921. [Google Scholar] [CrossRef]
- Gietl, E.; Mengerink, W.; de Slegte, J.; Gibson, G.; Rastall, R.; van den Heuvel, E. Factors Involved in the In Vitro Fermentability of Short Carbohydrates in Static Faecal Batch Cultures. Int. J. Carbohydr. Chem. 2012, 2012, 140. [Google Scholar] [CrossRef] [Green Version]
- Lane, J.A.; Calonne, J.; Slattery, H.; Hickey, R.M. Oligosaccharides Isolated from MGO™ Manuka Honey Inhibit the Adhesion of Pseudomonas aeruginosa, Escherichia coli O157: H7 and Staphylococcus Aureus to Human HT-29 Cells. Foods 2019, 8, 446. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, F.V.; Neifar, S.; Merdzo, Z.; Viña-Gonzalez, J.; Fernandez-Arrojo, L.; Ballesteros, A.O.; Fernandez-Lobato, M.; Bejar, S.; Plou, F.J. A Three-Step Process for the Bioconversion of Whey Permeate into a Glucose D-Free Tagatose Syrup. Catalysts 2020, 10, 647. [Google Scholar] [CrossRef]
- Cervantes, F.V.; Fernandez-polo, D.; Merdzo, Z.; Miguez, N.; Garcia-Gonzalez, M.; Ballesteros, A.O.; Fernandez-lobato, M.; Plou, F.J. Reuse of Immobilized Komagataella Phaffii Cells for the Elimination Of D-glucose in Syrups of Bioactive Carbohydrates. ACS Food Sci. Technol. 2022, 2, 682–690. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, M.; Plou, F.J.; Cervantes, F.V.; Remacha, M.; Poveda, A.; Jiménez-Barbero, J.; Fernandez-Lobato, M. Efficient Production of Isomelezitose by a Glucosyltransferase Activity in Metschnikowia reukaufii Cell Extracts. Microb. Biotechnol. 2019, 12, 1274–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Görl, J.; Timm, M.; Seibel, J. Mechanism-Oriented Redesign of an Isomaltulose Synthase to an Isomelezitose Synthase by Site-Directed Mutagenesis. ChemBioChem 2012, 13, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Munir, M.; Vogel, M. Method for Process for Producing Isomelezitose and Isomelezitose-Containing Sweeteners. German Patent DE19747642B4, 6 May 1999. [Google Scholar]
- Chiba, S.; Murata, M.; Matsusaka, K.; Shimomura, T. A New Trisaccharide, 6F-α-D-Glucosyl-Sucrose, Synthesized by Transglucosylation Reaction of Brewer’s Yeast α-Glucosidase. Agric. Biol. Chem. 1979, 43, 775–779. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Nakayama, T.; Noguchi, A.; Yano, M.; Hemmi, H.; Nishino, T.; Ueda, T. Site-Specific Mutagenesis at Positions 272 and 273 of the Bacillus Sp. SAM1606 α-Glucosidase to Screen Mutants with Altered Specificity for Oligosaccharide Production by Transglucosylation. J. Mol. Catal. B Enzym. 2002, 16, 265–274. [Google Scholar] [CrossRef]
- Ernits, K.; Kjeldsen, C.; Persson, K.; Grigor, E.; Alamäe, T.; Visnapuu, T. Structural Insight into a Yeast Maltase—The BaAG2 from Blastobotrys adeninivorans with Transglycosylating Activity. J. Fungi 2021, 7, 816. [Google Scholar] [CrossRef]
- Slininger, P.J.; Côté, G.L.; Shea-Andersh, M.A.; Dien, B.S.; Skory, C.D. Application of Isomelezitose as an Osmoprotectant for Biological Control Agent Preservation during Drying and Storage. Biocontrol Sci. Technol. 2021, 31, 132–152. [Google Scholar] [CrossRef]
- Míguez, N.; Gimeno-Pérez, M.; Fernández-Polo, D.; Cervantes, F.V.; Ballesteros, A.O.; Fernández-Lobato, M.; Ribeiro, M.H.; Plou, F.J. Immobilization of the β-Fructofuranosidase from Xanthophyllomyces dendrorhous by Entrapment in Polyvinyl Alcohol and Its Application to Neo-Fructooligosaccharides Production. Catalysts 2018, 8, 201. [Google Scholar] [CrossRef] [Green Version]
- Sakai, T.; Tsuji, H.; Shibata, S.; Hayakawa, K.; Matsumoto, K. Repeated-Batch Production of Galactooligosaccharides from Lactose at High Concentration by Using Alginate-Immobilized Cells of Sporobolomyces singularis YIT 10047. J. Gen. Appl. Microbiol. 2008, 54, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.T.; Park, K.M.; Kang, K.H.; Oh, D.J.; Lee, S.W.; Baig, S.Y.; Lee, J.H. Novel Method for Cell Immobilization and Its Application for Production of Oligosaccharides from Sucrose. Lett. Appl. Microbiol. 2004, 38, 176–179. [Google Scholar] [CrossRef]
- Rodrigo-Frutos, D.; Piedrabuena, D.; Sanz-Aparicio, J.; Fernández-Lobato, M. Yeast Cultures Expressing the Ffase from Schwanniomyces occidentalis, a Simple System to Produce the Potential Prebiotic Sugar 6-Kestose. Appl. Microbiol. Biotechnol. 2019, 103, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Rosendale, D.I.; Maddox, I.S.; Miles, M.C.; Rodier, M.; Skinner, M.; Sutherland, J. High-Throughput Microbial Bioassays to Screen Potential New Zealand Functional Food Ingredients Intended to Manage the Growth of Probiotic and Pathogenic Gut Bacteria. Int. J. Food Sci. Technol. 2008, 43, 2257–2267. [Google Scholar] [CrossRef]
- Goulas, A.K.; Grandison, A.S.; Rastall, R.A. Fractionation of Oligosaccharides by Nanofiltration. J. Sci. Food Agric. 2003, 83, 675–680. [Google Scholar] [CrossRef]
- Feng, Y.M.; Chang, X.L.; Wang, W.H.; Ma, R.Y. Separation of Galacto-Oligosaccharides Mixture by Nanofiltration. J. Taiwan Inst. Chem. Eng. 2009, 40, 326–332. [Google Scholar] [CrossRef]
- Lian, Z.; Wang, Y.; Luo, J.; Lai, C.; Yong, Q.; Yu, S. An Integrated Process to Produce Prebiotic Xylooligosaccharides by Autohydrolysis, Nanofiltration and Endo-xylanase from Alkali-Extracted Xylan. Bioresour. Technol. 2020, 314, 123685. [Google Scholar] [CrossRef]
- Yang, Z.D.; Guo, Y.S.; Huang, J.S.; Gao, Y.F.; Peng, F.; Xu, R.Y.; Su, H.H.; Zhang, P.J. Isomaltulose Exhibits Prebiotic Activity, and Modulates Gut Microbiota, the Production of Short Chain Fatty Acids, and Secondary Bile Acids in Rats. Molecules 2021, 26, 2464. [Google Scholar] [CrossRef]
- Rad, A.H.; Aghebati-Maleki, L.; Kafil, H.S.; Gilani, N.; Abbasi, A.; Khani, N. Postbiotics, as Dynamic Biomolecules, and Their Promising Role in Promoting Food Safety. Biointerface Res. Appl. Chem. 2021, 11, 14529–14544. [Google Scholar] [CrossRef]
- Aho, V.T.E.; Houser, M.C.; Pereira, P.A.B.; Chang, J.; Rudi, K.; Paulin, L.; Hertzberg, V.; Auvinen, P.; Tansey, M.G.; Scheperjans, F. Relationships of Gut Microbiota, Short-Chain Fatty Acids, Inflammation, and the Gut Barrier in Parkinson’s Disease. Mol. Neurodegener. 2021, 16, 6. [Google Scholar] [CrossRef]
- Campos-Perez, W.; Martinez-Lopez, E. Effects of Short Chain Fatty Acids on Metabolic and Inflammatory Processes in Human Health. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 1866, 158900. [Google Scholar] [CrossRef]
- Gill, P.A.; van Zelm, M.C.; Muir, J.G.; Gibson, P.R. Review Article: Short Chain Fatty Acids as Potential Therapeutic Agents in Human Gastrointestinal and Inflammatory Disorders. Aliment. Pharmacol. Ther. 2018, 48, 15–34. [Google Scholar] [CrossRef]
- Fernandez-Arrojo, L.; Santos-Moriano, P.; Rodriguez-Colinas, B.; Ballesteros, A.O.; Plou, F.J. Micro-Scale Procedure for Enzyme Immobilization Screening and Operational Stability Assays. Biotechnol. Lett. 2015, 37, 1593–1600. [Google Scholar] [CrossRef] [PubMed]
- Nobre, C.; Sousa, S.C.; Silva, S.P.; Pinheiro, A.C.; Coelho, E.; Vicente, A.A.; Gomes, A.M.P.; Coimbra, M.A.; Teixeira, J.A.; Rodrigues, L.R. In Vitro Digestibility and Fermentability of Fructo-Oligosaccharides Produced by Aspergillus ibericus. J. Funct. Foods 2018, 46, 278–287. [Google Scholar] [CrossRef] [Green Version]
- De la Rubia, M.A.; Villamil, J.; Rodriguez, J.; Borja, R.; Mohedano, A. Mesophilic anaerobic co-digestion of the organic fraction of municipal solid waste with the liquid fraction from hydrothermal carbonization of sewage sludge. Waste Manag. 2018, 76, 315–322. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glc + Fru | Suc | Tre | Isom | Rest of Hetero-GlcOS | Total Hetero-GlcOS | |
|---|---|---|---|---|---|---|
| Entrapped E. coli cells | 217 ± 8.9 | 65 ± 1.6 | 95 ± 3.8 | 75 ± 1.1 | 48 ± 1 | 218 ± 6 |
| Soluble Mr-αGlu | 97 ± 2 | 146 ± 16 | 61 ± 2 | 173 ± 14 | 23 ± 1.2 | 256 ± 17 |
| Bacteria | Grow Parameters on MRS Media | |||||
|---|---|---|---|---|---|---|
| ΔmaxOD600a | g (h) b | |||||
| NCS | FOS | Isom | NCS | FOS | Isom | |
| L. casei | 0.63 ± 0.03 | 0.71 ± 0.03 | 0.76 ± 0.01 * | 1.75 ± 0.01 | 1.34 ± 0.09 | 1.41 ± 0.08 * |
| L. rhamnosus | 0.94 ± 0.01 | 1.09 ± 0.02 | 1.13 ± 0.02 * | 1.30 ± 0.07 | 0.87 ± 0.04 | 0.80 ± 0.02 * |
| E. faecium | 0.58 ± 0.03 | 0.79 ± 0.03 | 0.89 ± 0.05 *,# | 1.70 ± 0.20 | 0.71 ± 0.14 | 0.87 ± 0.06 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Gonzalez, M.; Cervantes, F.V.; Ipiales, R.P.; de la Rubia, A.; Plou, F.J.; Fernández-Lobato, M. Isomelezitose Overproduction by Alginate-Entrapped Recombinant E. coli Cells and In Vitro Evaluation of Its Potential Prebiotic Effect. Int. J. Mol. Sci. 2022, 23, 12682. https://doi.org/10.3390/ijms232012682
Garcia-Gonzalez M, Cervantes FV, Ipiales RP, de la Rubia A, Plou FJ, Fernández-Lobato M. Isomelezitose Overproduction by Alginate-Entrapped Recombinant E. coli Cells and In Vitro Evaluation of Its Potential Prebiotic Effect. International Journal of Molecular Sciences. 2022; 23(20):12682. https://doi.org/10.3390/ijms232012682
Chicago/Turabian StyleGarcia-Gonzalez, Martin, Fadia V. Cervantes, Ricardo P. Ipiales, Angeles de la Rubia, Francisco J. Plou, and María Fernández-Lobato. 2022. "Isomelezitose Overproduction by Alginate-Entrapped Recombinant E. coli Cells and In Vitro Evaluation of Its Potential Prebiotic Effect" International Journal of Molecular Sciences 23, no. 20: 12682. https://doi.org/10.3390/ijms232012682
APA StyleGarcia-Gonzalez, M., Cervantes, F. V., Ipiales, R. P., de la Rubia, A., Plou, F. J., & Fernández-Lobato, M. (2022). Isomelezitose Overproduction by Alginate-Entrapped Recombinant E. coli Cells and In Vitro Evaluation of Its Potential Prebiotic Effect. International Journal of Molecular Sciences, 23(20), 12682. https://doi.org/10.3390/ijms232012682