
Gene Expression Detects the Factors Influencing the Reproductive Success and the Survival Rates of Paracentrotus lividus Offspring
 ,
, 

 and
and
Abstract
:1. Introduction
1.1. Sea Urchins and Aquaculture
1.2. Factors Influencing Larval Development
2. Results
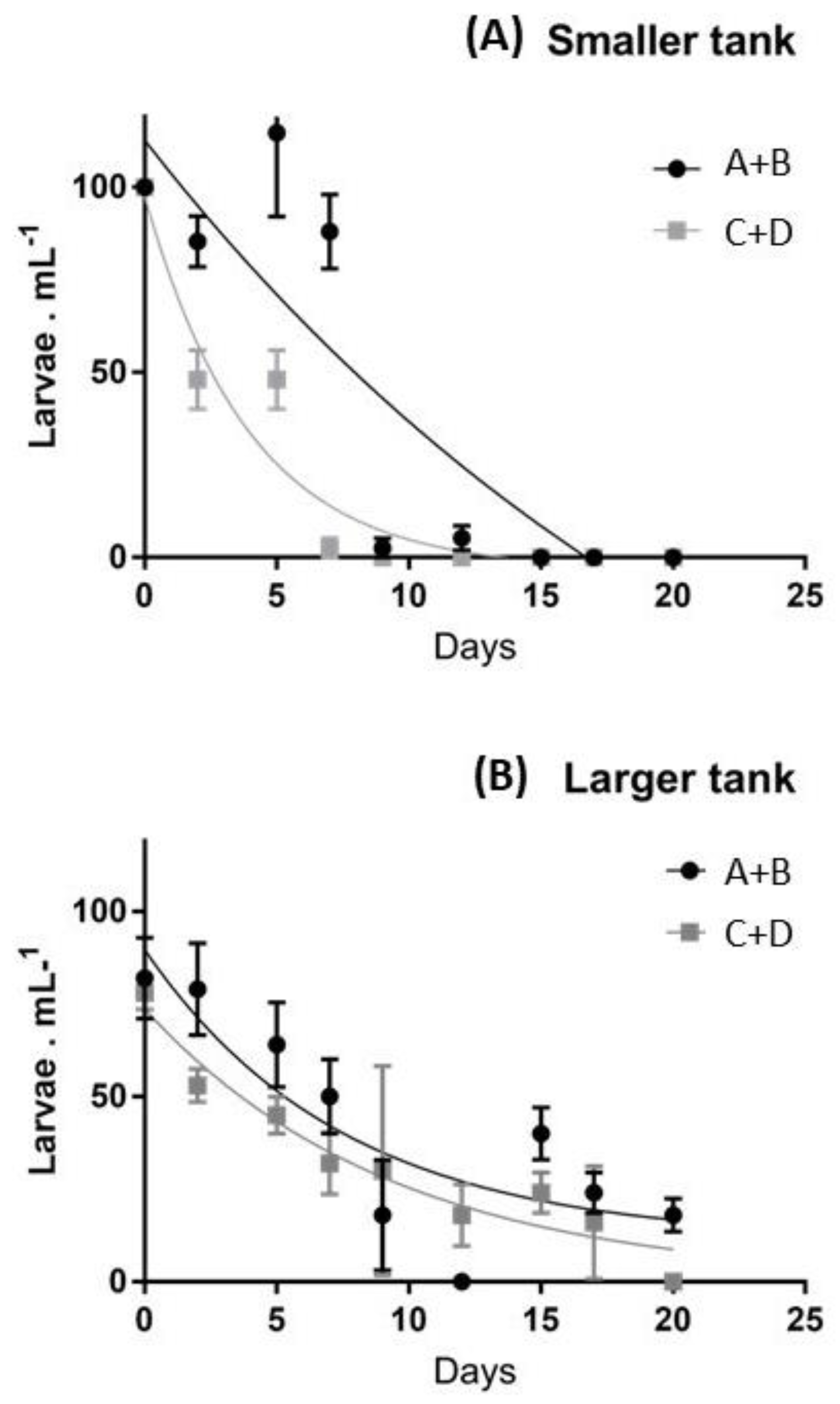
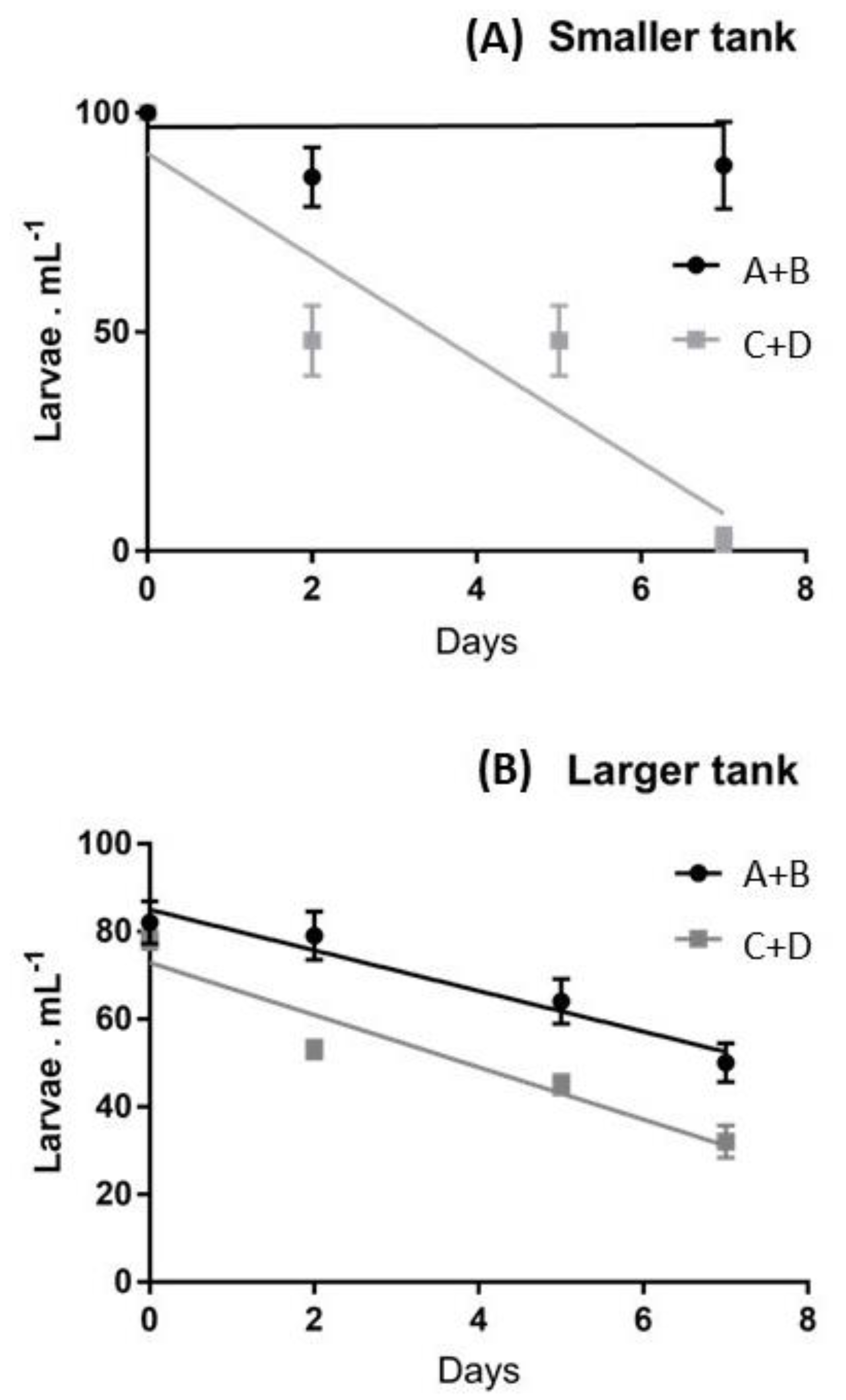
2.1. Survival Rates
2.2. Molecular Analyses of Embryos Deriving from Various Females
2.2.1. Genes Involved in Stress Response
2.2.2. Skeletogenic Genes
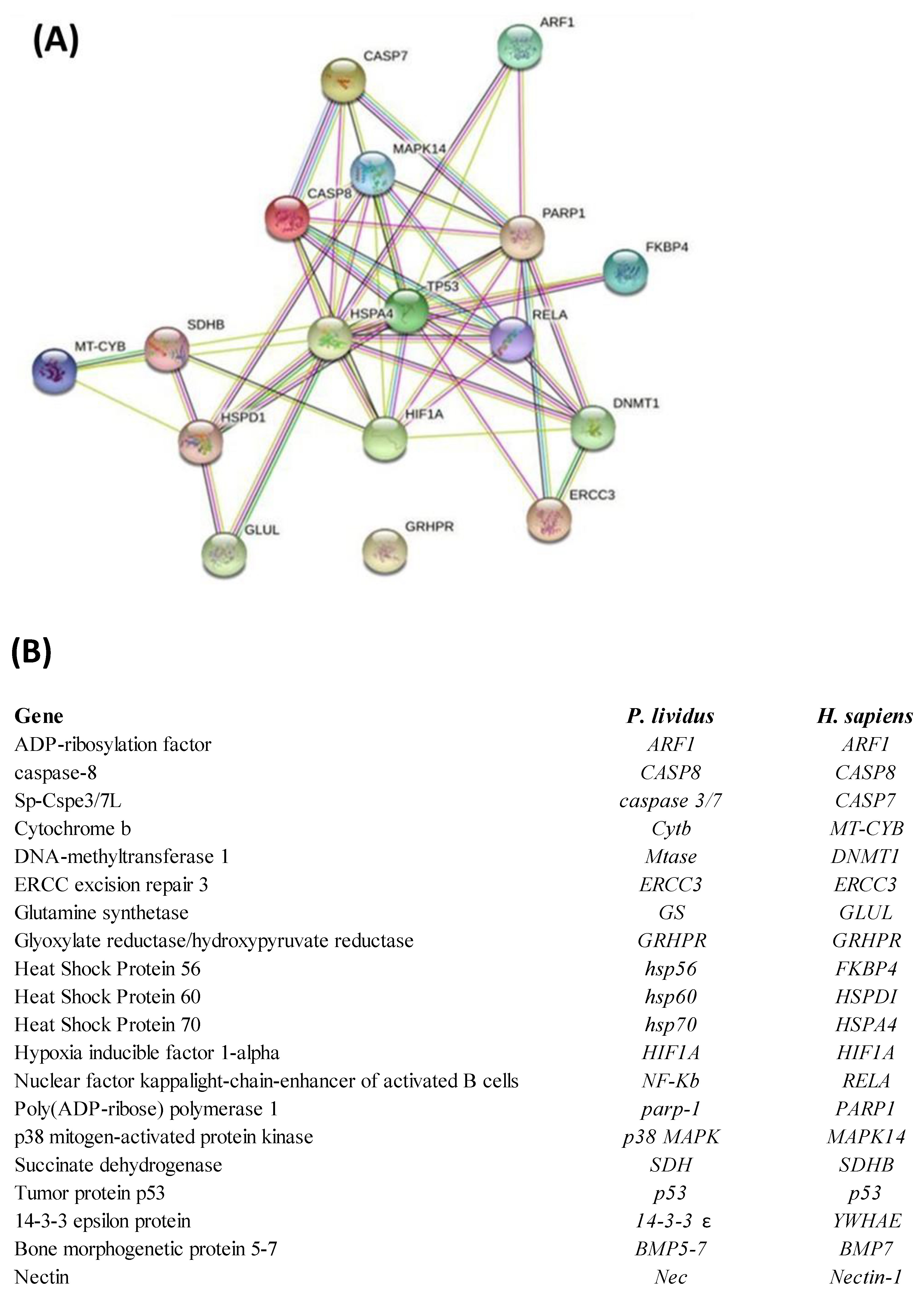
2.3. Network Analysis
3. Discussion
4. Materials and Method
4.1. Gamete Collection and Embryo Culture
4.2. Statistical Analyses
4.3. RNA Extraction and cDNA Synthesis
4.4. Real Time qPCR Experiments
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Böttger, S.A.; Devin, M.G.; Walker, C.W. Suspension of Annual Gametogenesis in North American Green Sea Urchins (Strongylocentrotus Droebachiensis) Experiencing Invariant Photoperiod—Applications for Land-Based Aquaculture. Aquaculture 2006, 261, 1422–1431. [Google Scholar] [CrossRef]
- Adonin, L.; Drozdov, A.; Barlev, N.A. Sea Urchin as a Universal Model for Studies of Gene Networks. Front. Genet. 2021, 11, 627259. [Google Scholar] [CrossRef] [PubMed]
- Marrone, V.; Piscopo, M.; Romano, G.; Ianora, A.; Palumbo, A.; Costantini, M. Defensome against Toxic Diatom Aldehydes in the Sea Urchin Paracentrotus Lividus. PLoS ONE 2012, 7, e31750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruocco, N.; Costantini, M.; Santella, L. New Insights into Negative Effects of Lithium on Sea Urchin Paracentrotus Lividus Embryos. Sci. Rep. 2016, 6, 32157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, G.; Costantini, M.; Buttino, I.; Ianora, A.; Palumbo, A. Nitric Oxide Mediates the Stress Response Induced by Diatom Aldehydes in the Sea Urchin Paracentrotus Lividus. PLoS ONE 2011, 6, e25980. [Google Scholar] [CrossRef] [Green Version]
- Luis, O.; Delgado, F.; Gago, J. Year-Round Captive Spawning Performance of the Sea Urchin Paracentrotus Lividus: Relevance for the Use of Its Larvae as Live Feed. Aquat. Living Resour. 2005, 18, 45–54. [Google Scholar] [CrossRef]
- Hammer, H.; Hammer, B.; Watts, S.; Lawrence, A.; Lawrence, J. The Effect of Dietary Protein and Carbohydrate Concentration on the Biochemical Composition and Gametogenic Condition of the Sea Urchin Lytechinus Variegatus. J. Exp. Mar. Biol. Ecol. 2006, 334, 109–121. [Google Scholar] [CrossRef]
- Carboni, S.; Vignier, J.; Chiantore, M.; Tocher, D.R.; Migaud, H. Effects of Dietary Microalgae on Growth, Survival and Fatty Acid Composition of Sea Urchin Paracentrotus Lividus throughout Larval Development. Aquaculture 2012, 324, 250–258. [Google Scholar] [CrossRef]
- Cárcamo, P.F.; Candia, A.I.; Chaparro, O.R. Larval Development and Metamorphosis in the Sea Urchin Loxechinus Albus (Echinodermata: Echinoidea): Effects of Diet Type and Feeding Frequency. Aquaculture 2005, 249, 375–386. [Google Scholar] [CrossRef]
- Castilla-Gavilán, M.; Buzin, F.; Cognie, B.; Dumay, J.; Turpin, V.; Decottignies, P. Optimising Microalgae Diets in Sea Urchin Paracentrotus Lividus Larviculture to Promote Aquaculture Diversification. Aquaculture 2018, 490, 251–259. [Google Scholar] [CrossRef]
- Kelly, M.S.; Hunter, A.J.; Scholfield, C.L.; McKenzie, J.D. Morphology and Survivorship of Larval Psammechinus Miliaris (Gmelin) (Echinodermata: Echinoidea) in Response to Varying Food Quantity and Quality. Aquaculture 2000, 183, 223–240. [Google Scholar] [CrossRef]
- Liu, H.; Kelly, M.S.; Cook, E.J.; Black, K.; Orr, H.; Zhu, J.X.; Dong, S.L. The Effect of Diet Type on Growth and Fatty Acid Composition of the Sea Urchin Larvae, II. Psammechinus Miliaris (Gmelin). Aquaculture 2007, 264, 263–278. [Google Scholar] [CrossRef]
- Guedes, A.C.; Malcata, F.X. Nutritional Value and Uses of Microalgae in Aquaculture. Aquaculture 2012, 10, 59–78. [Google Scholar]
- Brown, M.R.; Jeffrey, S.W.; Volkman, J.K.; Dunstan, G.A. Nutritional Properties of Microalgae for Mariculture. Aquaculture 1997, 151, 315–331. [Google Scholar] [CrossRef]
- Zupo, V.; Glaviano, F.; Caramiello, D.; Mutalipassi, M. Effect of Five Benthic Diatoms on the Survival and Development of Paracentrotus Lividus Post-Larvae in the Laboratory. Aquaculture 2018, 495, 13–20. [Google Scholar] [CrossRef]
- Ruocco, N.; Costantini, S.; Zupo, V.; Lauritano, C.; Caramiello, D.; Ianora, A.; Budillon, A.; Romano, G.; Nuzzo, G.; D’Ippolito, G. Toxigenic Effects of Two Benthic Diatoms upon Grazing Activity of the Sea Urchin: Morphological, Metabolomic and de Novo Transcriptomic Analysis. Sci. Rep. 2018, 8, 5622. [Google Scholar] [CrossRef] [Green Version]
- Kipryushina, Y.O.; Yakovlev, K.V. Maternal Control of Early Patterning in Sea Urchin Embryos. Differentiation 2020, 113, 28–37. [Google Scholar] [CrossRef]
- Tu, Q.; Brown, C.T.; Davidson, E.H.; Oliveri, P. Sea Urchin Forkhead Gene Family: Phylogeny and Embryonic Expression. Dev. Biol. 2006, 300, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Yaguchi, S.; Yaguchi, J.; Angerer, R.C.; Angerer, L.M. A Wnt-FoxQ2-Nodal Pathway Links Primary and Secondary Axis Specification in Sea Urchin Embryos. Dev. Cell 2008, 14, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Burton, T.; McKelvey, S.; Stewart, D.C.; Armstrong, J.D.; Metcalfe, N.B. Early Maternal Experience Shapes Offspring Performance in the Wild. Ecology 2013, 94, 618–626. [Google Scholar] [CrossRef] [Green Version]
- Venturelli, P.A.; Murphy, C.A.; Shuter, B.J.; Johnston, T.A.; van Coeverden de Groot, P.J.; Boag, P.T.; Casselman, J.M.; Montgomerie, R.; Wiegand, M.D.; Leggett, W.C. Maternal Influences on Population Dynamics: Evidence from an Exploited Freshwater Fish. Ecology 2010, 91, 2003–2012. [Google Scholar] [CrossRef] [PubMed]
- Green, B.S. Maternal Effects in Fish Populations. Adv. Mar. Biol. 2008, 54, 1–105. [Google Scholar] [PubMed]
- Marshall, D.J. Transgenerational Plasticity in the Sea: Context-dependent Maternal Effects across the Life History. Ecology 2008, 89, 418–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uller, T. Developmental Plasticity and the Evolution of Parental Effects. Trends Ecol. Evol. 2008, 23, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Benton, T.G.; St. Clair, J.J.H.; Plaistow, S.J. Maternal Effects Mediated by Maternal Age: From Life Histories to Population Dynamics. J. Anim. Ecol. 2008, 77, 1038–1046. [Google Scholar] [CrossRef]
- Leber, K.M.; Lorenzen, K.; Main, K.L.; Moe, M., Jr.; Vaughan, D.; Capo, T.; Bardales, A.; Gillette, P. Developing Restoration Methods to Aid in Recovery of a Key Herbivore, Diadema Antillarum, on Florida Coral Reefs-2008/2009 Final Report (April 1, 2008 to March 31, 2009); Mote Marine Laboratory: Sarasota, FL, USA; University of Miami: Coral Gables, FL, USA, 2009. [Google Scholar]
- Grosjean, P. Growth Model of the Reared Sea Urchin Paracentrotus Lividus (Lamarck, 1816). Ph.D. Thesis, Université de Mons, Mons, Belgium, 2001. [Google Scholar]
- Grosjean, P.; Spirlet, C.; Jangoux, M. Experimental Study of Growth in the Echinoid Paracentrotus Lividus (Lamarck, 1816) (Echinodermata). J. Exp. Mar. Biol. Ecol. 1996, 201, 173–184. [Google Scholar] [CrossRef]
- Varrella, S.; Romano, G.; Ianora, A.; Bentley, M.G.; Ruocco, N.; Costantini, M. Molecular Response to Toxic Diatom-Derived Aldehydes in the Sea Urchin Paracentrotus Lividus. Mar. Drugs 2014, 12, 2089–2113. [Google Scholar] [CrossRef] [Green Version]
- Varrella, S.; Romano, G.; Ruocco, N.; Ianora, A.; Bentley, M.G.; Costantini, M. First Morphological and Molecular Evidence of the Negative Impact of Diatom-Derived Hydroxyacids on the Sea Urchin Paracentrotus Lividus. Toxicol. Sci. 2016, 151, 419–433. [Google Scholar] [CrossRef] [Green Version]
- Ruocco, N.; Costantini, S.; Zupo, V.; Romano, G.; Ianora, A.; Fontana, A.; Costantini, M. High-Quality RNA Extraction from the Sea Urchin Paracentrotus Lividus Embryos. PLoS ONE 2017, 12, e0172171. [Google Scholar] [CrossRef]
- Morroni, L.; Sartori, D.; Costantini, M.; Genovesi, L.; Magliocco, T.; Ruocco, N.; Buttino, I. First Molecular Evidence of the Toxicogenetic Effects of Copper on Sea Urchin Paracentrotus Lividus Embryo Development. Water Res. 2019, 160, 415–423. [Google Scholar] [CrossRef]
- Albarano, L.; Zupo, V.; Guida, M.; Libralato, G.; Caramiello, D.; Ruocco, N.; Costantini, M. PAHs and PCBs Affect Functionally Intercorrelated Genes in the Sea Urchin Paracentrotus Lividus Embryos. Int. J. Mol. Sci. 2021, 22, 12498. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Pinsino, A.; Costa, C.; Bonaventura, R.; Matranga, V.; Zito, F. The Newly Characterized Pl-jun Is Specifically Expressed in Skeletogenic Cells of the Paracentrotus Lividus Sea Urchin Embryo. FEBS J. 2014, 281, 3828–3843. [Google Scholar] [CrossRef] [PubMed]
- Varrella, S.; Romano, G.; Costantini, S.; Ruocco, N.; Ianora, A.; Bentley, M.G.; Costantini, M. Toxic Diatom Aldehydes Affect Defence Gene Networks in Sea Urchins. PLoS ONE 2016, 11, e0149734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruocco, N.; Varrella, S.; Romano, G.; Ianora, A.; Bentley, M.G.; Somma, D.; Leonardi, A.; Mellone, S.; Zuppa, A.; Costantini, M. Diatom-Derived Oxylipins Induce Cell Death in Sea Urchin Embryos Activating Caspase-8 and Caspase 3/7. Aquat. Toxicol. 2016, 176, 128–140. [Google Scholar] [CrossRef]
- Ruocco, N.; Annunziata, C.; Ianora, A.; Libralato, G.; Manfra, L.; Costantini, S.; Costantini, M. Toxicity of Diatom-Derived Polyunsaturated Aldehyde Mixtures on Sea Urchin Paracentrotus Lividus Development. Sci. Rep. 2019, 9, 517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, R.; Ruocco, N.; Albarano, L.; Ianora, A.; Manfra, L.; Libralato, G.; Costantini, M. Combined Effects of Diatom-Derived Oxylipins on the Sea Urchin Paracentrotus Lividus. Int. J. Mol. Sci. 2020, 21, 719. [Google Scholar] [CrossRef] [Green Version]
- Ross, P.M.; Parker, L.; Byrne, M. Transgenerational Responses of Molluscs and Echinoderms to Changing Ocean Conditions. ICES J. Mar. Sci. 2016, 73, 537–549. [Google Scholar] [CrossRef] [Green Version]
- Byrne, M.; Prowse, T.A.A.; Sewell, M.A.; Dworjanyn, S.; Williamson, J.E.; Vaïtilingon, D. Maternal Provisioning for Larvae and Larval Provisioning for Juveniles in the Toxopneustid Sea Urchin Tripneustes Gratilla. Mar. Biol. 2008, 155, 473–482. [Google Scholar] [CrossRef]
- Kovalchuk, I. Transgenerational Epigenetic Inheritance in Animals. Front. Genet. 2012, 3, 76. [Google Scholar] [CrossRef] [Green Version]
- Munday, P.L.; Warner, R.R.; Monro, K.; Pandolfi, J.M.; Marshall, D.J. Predicting Evolutionary Responses to Climate Change in the Sea. Ecol. Lett. 2013, 16, 1488–1500. [Google Scholar] [CrossRef]
- Byrne, M.; Selvakumaraswamy, P.; Ho, M.A.; Woolsey, E.; Nguyen, H.D. Sea Urchin Development in a Global Change Hotspot, Potential for Southerly Migration of Thermotolerant Propagules. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 712–719. [Google Scholar]
- Shama, L.N.S.; Strobel, A.; Mark, F.C.; Wegner, K.M. Transgenerational Plasticity in Marine Sticklebacks: Maternal Effects Mediate Impacts of a Warming Ocean. Funct. Ecol. 2014, 28, 1482–1493. [Google Scholar] [CrossRef]
- Hamdoun, A.; Epel, D. Embryo Stability and Vulnerability in an Always Changing World. Proc. Natl. Acad. Sci. USA 2007, 104, 1745–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suckling, C.C.; Clark, M.S.; Richard, J.; Morley, S.A.; Thorne, M.A.S.; Harper, E.M.; Peck, L.S. Adult Acclimation to Combined Temperature and pH Stressors Significantly Enhances Reproductive Outcomes Compared to Short-term Exposures. J. Anim. Ecol. 2015, 84, 773–784. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, L.; Shi, D.; Ding, J.; Yin, D.; Sun, J.; Zhang, B.; Zhang, L.; Chang, Y. Transgenerational Effects of Ocean Warming on the Sea Urchin Strongylocentrotus Intermedius. Ecotoxicol. Environ. Saf. 2018, 151, 212–219. [Google Scholar] [CrossRef]
- Schlegel, P.; Havenhand, J.N.; Gillings, M.R.; Williamson, J.E. Individual Variability in Reproductive Success Determines Winners and Losers under Ocean Acidification: A Case Study with Sea Urchins. PLoS ONE 2012, 7, e53118. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.N.; Andersson, A.J.; Day, J.M.D.; Taylor, J.R.A.; Frank, M.B.; Jung, J.-Y.; McKittrick, J.; Levin, L.A. Response of Sea Urchin Fitness Traits to Environmental Gradients across the Southern California Oxygen Minimum Zone. Front. Mar. Sci. 2018, 5, 258. [Google Scholar] [CrossRef]
- Vafidis, D.; Antoniadou, C.; Kyriakouli, K. Reproductive Cycle of the Edible Sea Urchin Paracentrotus Lividus (Echinodermata: Echinoidae) in the Aegean Sea. Water 2019, 11, 1029. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Chang, Y.; Luo, S.; Zhou, H.; Tian, X.; Ding, J.; Chen, X. Effects of Biofilms as the Main and as a Supplementary Food on the Survival, Somatic Growth and Gonad Enhancement of Sea Urchin Strongylocentrotus Intermedius. Aquac. Int. 2014, 22, 925–936. [Google Scholar] [CrossRef]
- Rinde, E.; Christie, H.; Fagerli, C.W.; Bekkby, T.; Gundersen, H.; Norderhaug, K.M.; Hjermann, D.Ø. The Influence of Physical Factors on Kelp and Sea Urchin Distribution in Previously and Still Grazed Areas in the NE Atlantic. PLoS ONE 2014, 9, e100222. [Google Scholar] [CrossRef] [Green Version]
- López, S.; Turon, X.; Montero, E.; Palacín, C.; Duarte, C.M.; Tarjuelo, I. Larval Abundance, Recruitment and Early Mortality in Paracentrotus Lividus (Echinoidea). Interannual Variability and Plankton-Benthos Coupling. Mar. Ecol. Prog. Ser. 1998, 172, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Ding, J.; Ding, S.; Chang, Y. Integrated Metabolomic and Transcriptomic Analyses Identify Critical Genes in Eicosapentaenoic Acid Biosynthesis and Metabolism in the Sea Urchin Strongylocentrotus Intermedius. Sci. Rep. 2020, 10, 1697. [Google Scholar] [CrossRef] [PubMed]
- Picard, V.; Mulner-Lorillon, O.; Bourdon, J.; Morales, J.; Cormier, P.; Siegel, A.; Bellé, R. Model of the Delayed Translation of Cyclin B Maternal MRNA after Sea Urchin Fertilization. Mol. Reprod. Dev. 2016, 83, 1070–1082. [Google Scholar] [CrossRef] [Green Version]
- Shi, D.; Zhao, C.; Chen, Y.; Ding, J.; Zhang, L.; Chang, Y. Transcriptomes Shed Light on Transgenerational and Developmental Effects of Ocean Warming on Embryos of the Sea Urchin Strongylocentrotus Intermedius. Sci. Rep. 2020, 10, 7931. [Google Scholar] [CrossRef] [PubMed]
- Klosin, A.; Lehner, B. Mechanisms, Timescales and Principles of Trans-Generational Epigenetic Inheritance in Animals. Curr. Opin. Genet. Dev. 2016, 36, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Bonduriansky, R.; Crean, A.J.; Day, T. The Implications of Nongenetic Inheritance for Evolution in Changing Environments. Evol. Appl. 2012, 5, 192–201. [Google Scholar] [CrossRef]
- Bertoldo, M.J.; Locatelli, Y.; O’Neill, C.; Mermillod, P.; Bertoldo, M.J.; Locatelli, Y.; O’Neill, C.; Mermillod, P. Impacts of and Interactions between Environmental Stress and Epigenetic Programming during Early Embryo Development. Reprod. Fertil. Dev. 2015, 27, 1125–1136. [Google Scholar] [CrossRef]
- Masullo, T.; Biondo, G.; Natale, M.D.; Tagliavia, M.; Bennici, C.D.; Musco, M.; Ragusa, M.A.; Costa, S.; Cuttitta, A.; Nicosia, A. Gene Expression Changes after Parental Exposure to Metals in the Sea Urchin Affect Timing of Genetic Programme of Embryo Development. Biology 2021, 10, 103. [Google Scholar] [CrossRef]
- Goldstone, J.V.; Hamdoun, A.; Cole, B.J.; Howard-Ashby, M.; Nebert, D.W.; Scally, M.; Dean, M.; Epel, D.; Hahn, M.E.; Stegeman, J.J. The Chemical Defensome: Environmental Sensing and Response Genes in the Strongylocentrotus Purpuratus Genome. Dev. Biol. 2006, 300, 366–384. [Google Scholar]
- Harris, L.G.; Eddy, S.D. Sea Urchin Ecology and Biology. In Echinoderm Aquaculture; Wiley: Hoboken, NJ, USA, 2015; Part I, Chapter 1; pp. 1–24. [Google Scholar]
- McBride, S.C. Sea Urchin Aquaculture. In American Fisheries Society Symposium; American Fisheries Society: Bethesda, MD, USA, 2005; Volume 46, p. 179. [Google Scholar]
- Ragusa, M.A.; Costa, S.; Gianguzza, M.; Roccheri, M.C.; Gianguzza, F. Effects of Cadmium Exposure on Sea Urchin Development Assessed by SSH and RT-QPCR: Metallothionein Genes and Their Differential Induction. Mol. Biol. Rep. 2013, 40, 2157–2167. [Google Scholar]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative Expression Software Tool (REST©) for Group-Wise Comparison and Statistical Analysis of Relative Expression Results in Real-Time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P. STRING V11: Protein–Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Paired t Test | Smaller | Larger |
|---|---|---|
| pvalue | 0.0830 | 0.1169 |
| pvalue summary | n.s. | n.s. |
| t | 1.980 | 1.758 |
| df | 8 | 8 |
| Mean of differences (B-A) | −21.93 | −8.778 |
| SD of differences | 33.22 | 14.98 |
| SEM of differences | 11.07 | 4.994 |
| 95% confidence interval | −47.46 to 3.610 | −20.29 to 2.738 |
| R squared (partial eta squared) | 0.3289 | 0.2786 |
| Correlation coefficient (r) | 0.7608 | 0.8595 |
| p value (one tailed) | 0.0086 | 0.0015 |
| p value summary | ** | ** |
| Significantly effective pairing | Yes | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Federico, S.; Glaviano, F.; Esposito, R.; Pinto, B.; Gharbi, M.; Di Cosmo, A.; Costantini, M.; Zupo, V. Gene Expression Detects the Factors Influencing the Reproductive Success and the Survival Rates of Paracentrotus lividus Offspring. Int. J. Mol. Sci. 2022, 23, 12790. https://doi.org/10.3390/ijms232112790
Federico S, Glaviano F, Esposito R, Pinto B, Gharbi M, Di Cosmo A, Costantini M, Zupo V. Gene Expression Detects the Factors Influencing the Reproductive Success and the Survival Rates of Paracentrotus lividus Offspring. International Journal of Molecular Sciences. 2022; 23(21):12790. https://doi.org/10.3390/ijms232112790
Chicago/Turabian StyleFederico, Serena, Francesca Glaviano, Roberta Esposito, Bruno Pinto, Maissa Gharbi, Anna Di Cosmo, Maria Costantini, and Valerio Zupo. 2022. "Gene Expression Detects the Factors Influencing the Reproductive Success and the Survival Rates of Paracentrotus lividus Offspring" International Journal of Molecular Sciences 23, no. 21: 12790. https://doi.org/10.3390/ijms232112790
APA StyleFederico, S., Glaviano, F., Esposito, R., Pinto, B., Gharbi, M., Di Cosmo, A., Costantini, M., & Zupo, V. (2022). Gene Expression Detects the Factors Influencing the Reproductive Success and the Survival Rates of Paracentrotus lividus Offspring. International Journal of Molecular Sciences, 23(21), 12790. https://doi.org/10.3390/ijms232112790