
The N-Linked Glycosylation Site N191 Is Necessary for PKA Signal Transduction in Eel Follicle-Stimulating Hormone Receptor


 ,
, 
Abstract
:1. Introduction
2. Results
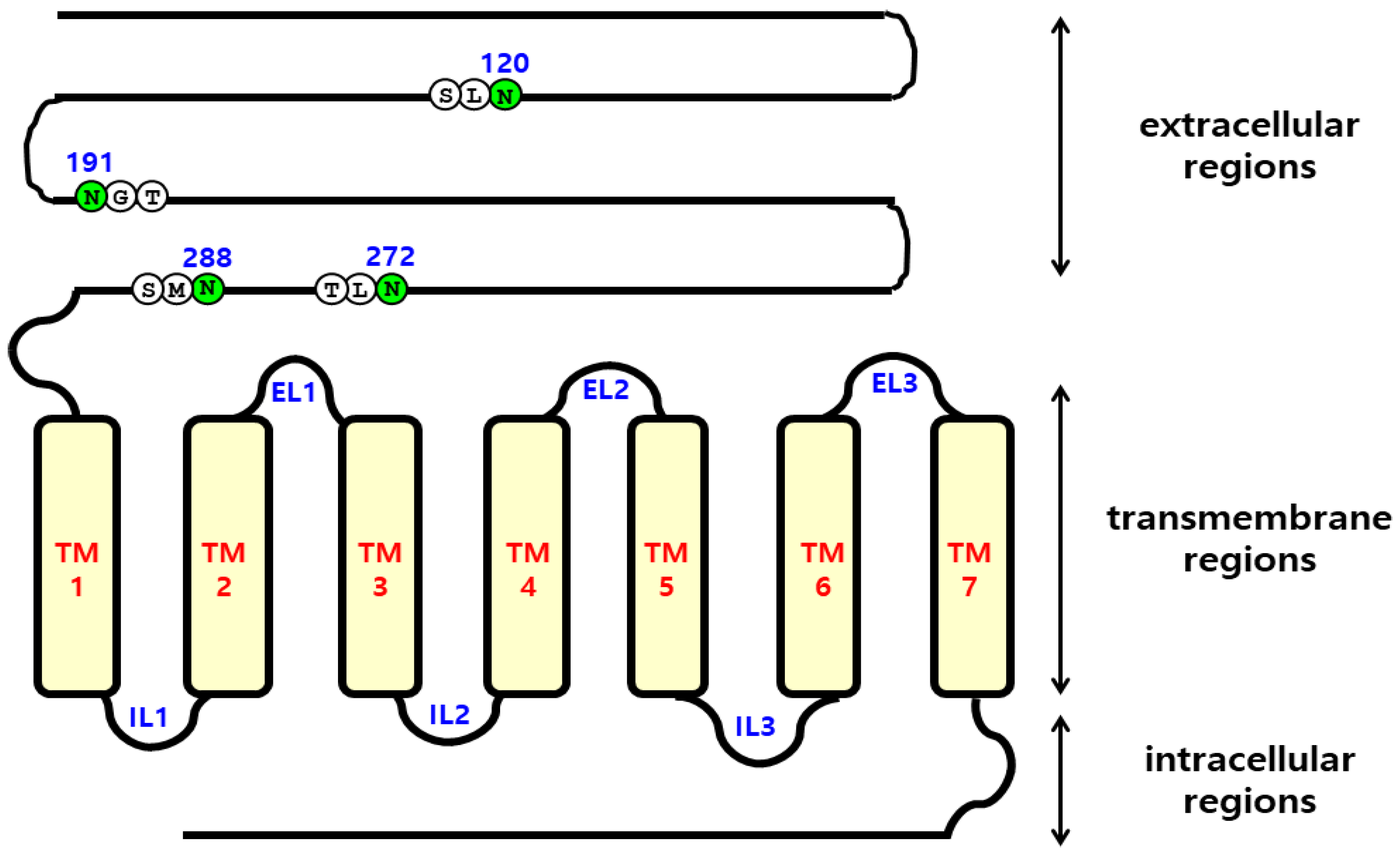
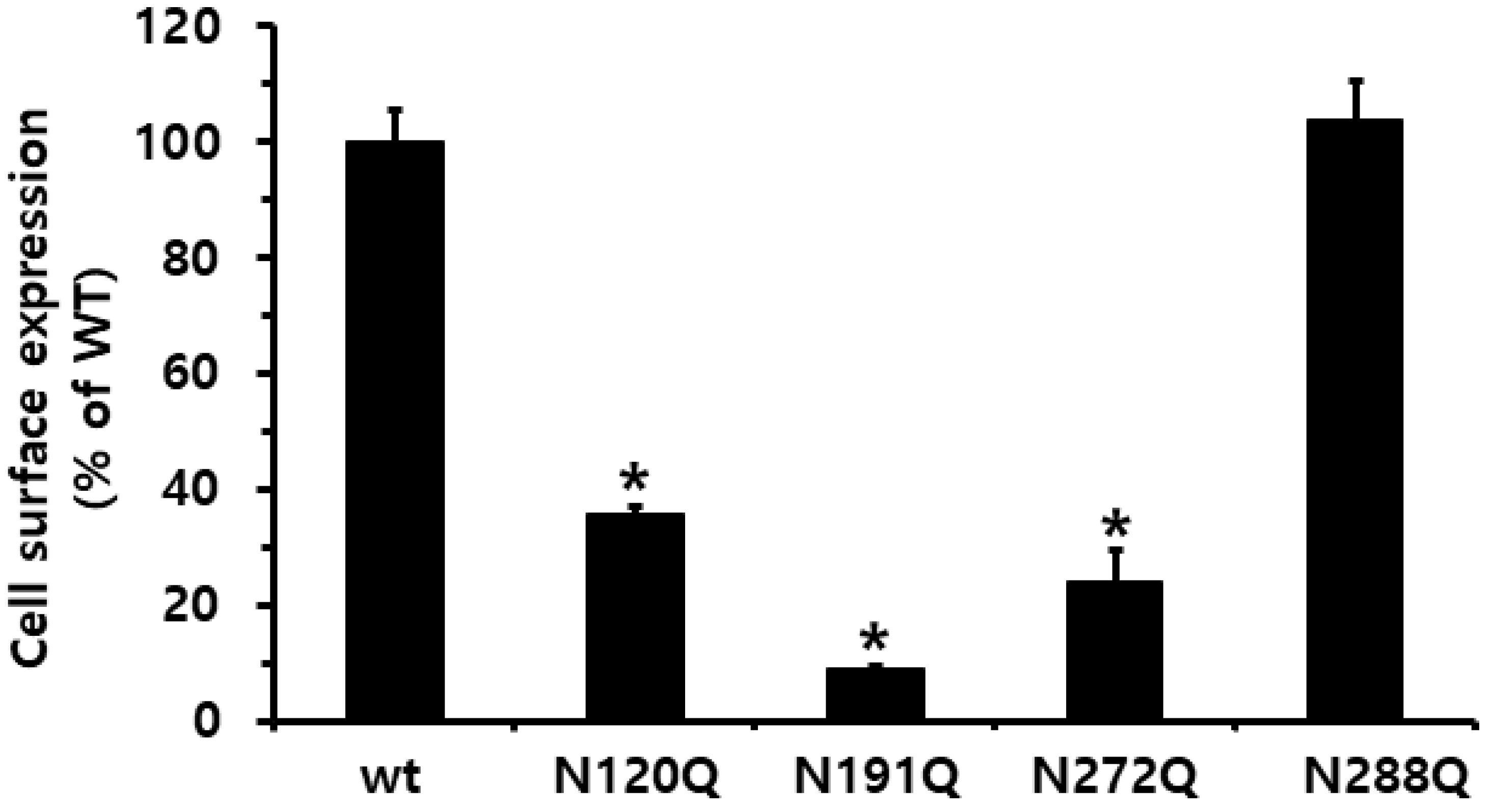
2.1. Preparation and Cell-Surface Expression of Eel FSHR Mutants and Expression Vectors
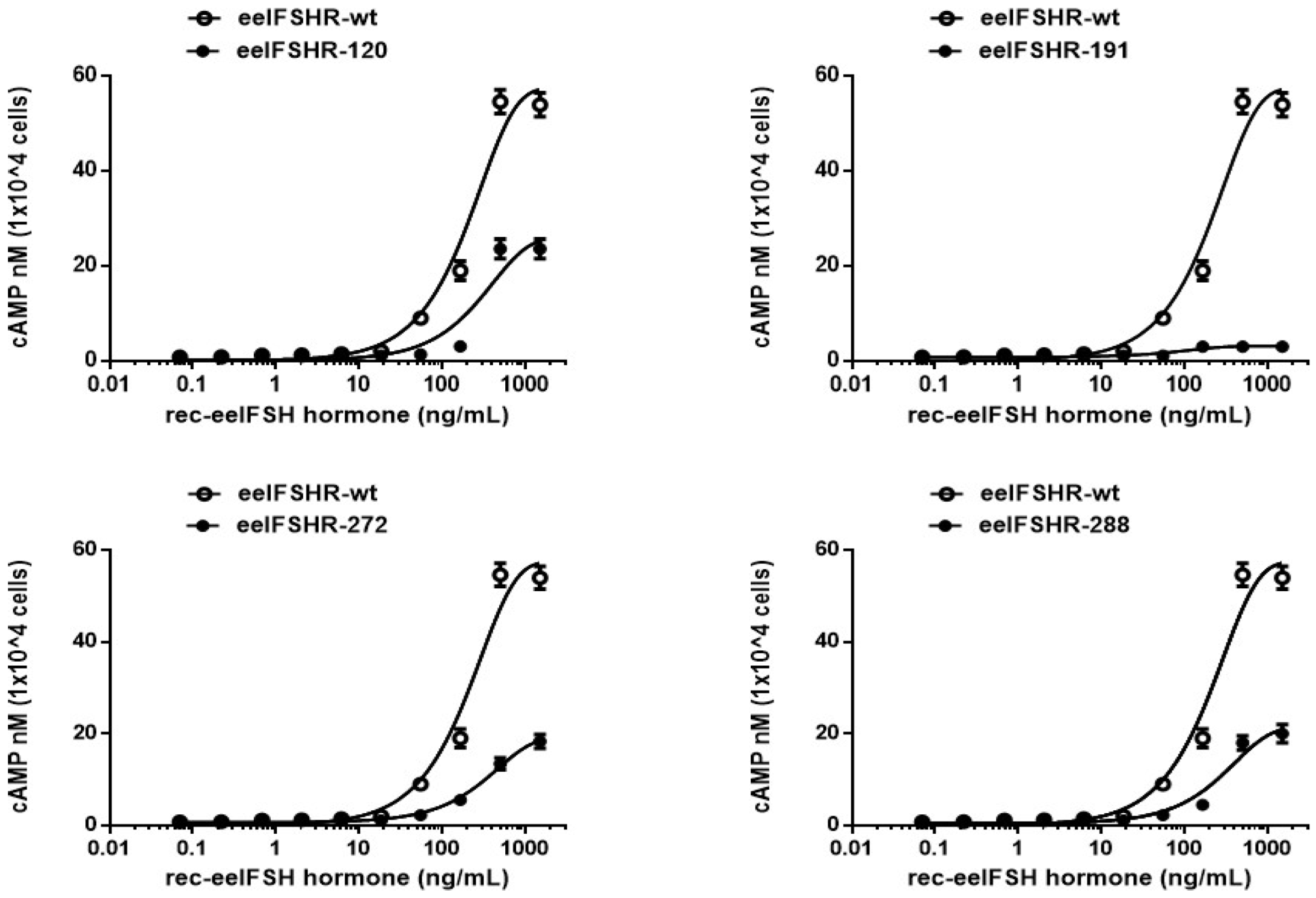
2.2. cAMP Responsiveness Induced by Agonist in N-Linked Glycosylated Mutants
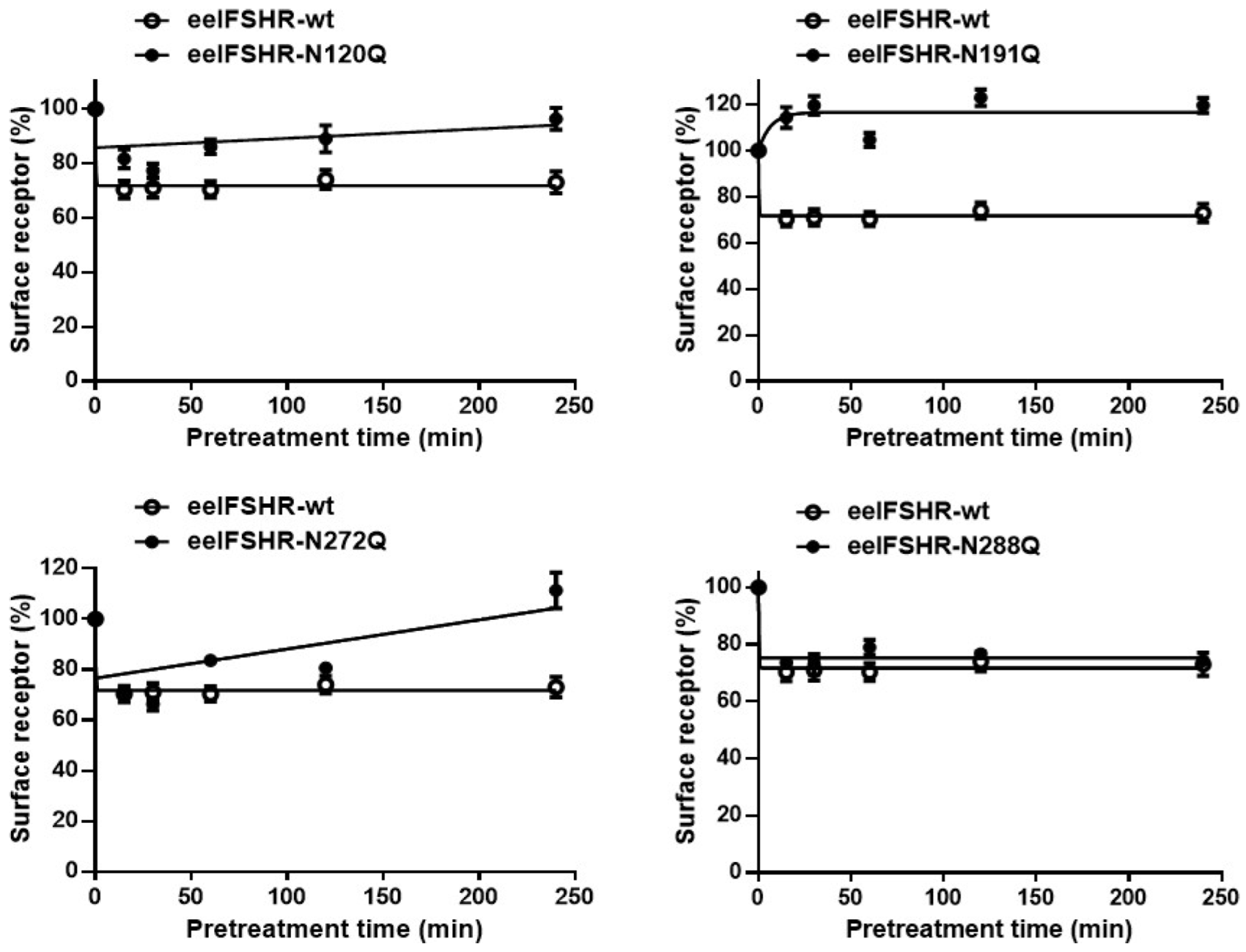
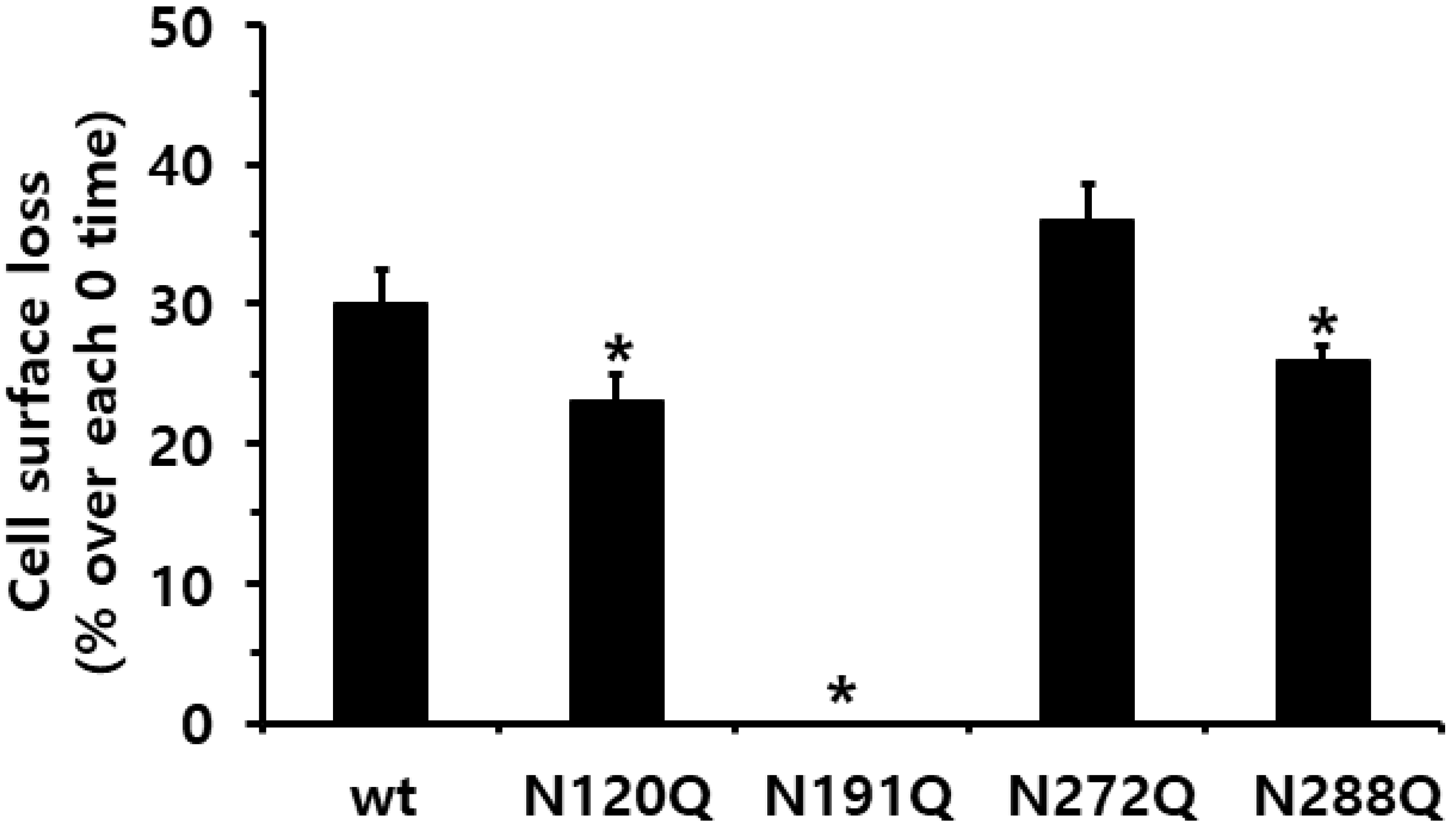
2.3. Cell-Surface Receptor Loss Induced by Treatment with the Eel FSH Agonist
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Site-Directed Mutagenesis
4.3. Vector Construction for Transfection in Mammalian Cells
4.4. Production and ELISA Analysis of Recombinant-eel FSH (rec-eel FSH)
4.5. Transient Transfection of Eel FSHR Mutant Genes
4.6. cAMP Analysis by Homogeneous Time-Resolved Fluorescence (HTRF)
4.7. Agonist-Induced Loss of Cell Surface Receptor
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nakamura, K.; Krupnic, J.G.; Benovic, J.L.; Ascoli, M. Signaling and phosphorylation-impaired mutants of the rat follitropin receptor reveal an activation- and phosphorylation-independent but arrestin-dependent pathway for internalization. J. Biol. Chem. 1998, 273, 24336–24354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hipkin, R.W.; Liu, X.; Ascoli, M. Truncation of the C-terminal tail of the follitropin receptor does not impair the agonist- or phorbol ester-induced receptor phosphorylation and uncoupling. J. Biol. Chem. 1995, 270, 26682–26689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byambaragchaa, M.; Kim, J.S.; Park, H.K.; Kim, D.J.; Hong, S.M.; Kang, M.H.; Min, K.S. Constitutive activation and inactivation of mutations inducing cell surface loss of receptor and impairing of signal transduction of agonist-stimulated eel follicle-stimulating hormone receptor. Int. J. Mol. Sci. 2020, 21, 7075. [Google Scholar] [CrossRef] [PubMed]
- Byambaragchaa, M.; Seong, H.K.; Choi, S.H.; Kim, D.J.; Kang, M.H.; Min, K.S. Constitutively activating mutants of equine LH/CGR constitutively induce signal transduction and inactivating mutations impair biological activity and cell-surface receptor loss in vitro. Int. J. Mol. Sci. 2021, 22, 10723. [Google Scholar] [CrossRef] [PubMed]
- Byambaragchaa, M.; Ahn, T.Y.; Choi, S.H.; Kang, M.H.; Min, K.S. Functional characterization of naturally-occurring constitutively activating/inactivating mutations in equine follicle-stimulating hormone receptor. Anim. Biosci. 2022, 35, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Eggert, U.S. Non-traditional roles of G protein-coupled receptors in basic cell biology. Mol. Biosyst. 2013, 9, 586–595. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, S.G.; DeVree, B.T.; Zou, Y.; Kruse, A.C.; Chung, K.Y.; Kobilka, T.S.; Thian, F.S.; Chae, P.S.; Pardon, E.; Calinski, D.; et al. Crystal structure of the β2 adrenergic receptor-Gs protein complex. Nature 2011, 477, 549–555. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.J.; Park, C.W.; Byambaragchaa, M.; Kim, S.K.; Lee, B.I.; Hwang, H.K.; Myeong, J.I.; Hong, S.M.; Kang, M.H.; Min, K.S. Data on the characterization of follicle-stimulating hormone monoclonal antibodies and localization in Japanese eel pituitary. Data Brief 2016, 8, 404–410. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.M.; Munkhuu, O.; Byambaragchaa, M.; Lee, B.I.; Kim, S.K.; Kang, M.H.; Kim, D.J.; Min, K.S. Site-specific roles of N-linked oligosaccharides in recombinant eel follicle-stimulating hormone for secretion and signal transduction. Gen. Comp. Endocrinol. 2019, 276, 37–44. [Google Scholar] [CrossRef]
- Pfleger, J.; Gresham, K.; Koch, W.J. G protein-coupled receptor kinases as therapeutic targets in the heart. Nat. Rev. Cardiol. 2019, 16, 612–622. [Google Scholar] [CrossRef]
- Kobayashi, M.; Morita, T.; Ikeguchi, K.; Yshizaki, G.; Suzuki, T.; Watabe, S. In vivo biological activity of recombinant goldfish gonadotropins produced by baculovirus in silkworm larvae. Aquaculture 2006, 256, 433–442. [Google Scholar] [CrossRef]
- Ko, H.Y.; Park, W.D.; Kim, D.J.; Kobayashi, M.; Sohn, Y.C. Biological activities of recombinant Manchurian trout FSH and LH: Their receptor specificity, steroidogenic and vitellogenic potencies. J. Mol. Endocrinol. 2007, 38, 99–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogerd, J.; Blomenrohr, M.; Andersson, E.; van der Putten, H.H.; Tensen, C.P.; Visher, H.F.; Granneman, J.C.; Janssen-Dommerholt, C.; Goos, H.J.; Schulz, R.W. Discrepancy between molecular structure and ligand selectivity of a testicular follicle-stimulating hormone receptor of the African catfish (Clarias gariepinus). Biol. Reprod. 2001, 64, 1633–1643. [Google Scholar] [CrossRef] [PubMed]
- Vischer, H.F.; Bogerd, J. Cloning and functional characterization of a gonadal luteinizing hormone receptor complementary DNA for the African catfish (Clarias gariepinus). Biol. Reprod. 2003, 68, 262–271. [Google Scholar] [CrossRef] [Green Version]
- Kwok, H.F.; So, W.K.; Wang, Y.; Ge, W. Zebrafish gonadotropins and their receptors: I. Cloning and characterization of zebrafish follicle-stimulating hormone and luteinizing hormone receptors—Evidence for their distinct functions in follicle development. Biol. Reprod. 2005, 72, 1370–1381. [Google Scholar] [CrossRef]
- So, W.K.; Kwok, H.F.; Ge, W. Zebrafish gonadotropins and their receptors: II. Cloning and characterization of zebrafish follicle-stimulating hormone and luteinizing hormone receptors—Their spatial-temporal expression patterns and receptor specificity. Biol. Reprod. 2005, 72, 1382–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byambaragchaa, M.; Choi, S.H.; Kim, D.W.; Min, K.S. Constitutive activating eel luteinizing hormone receptors induce constitutively signal transduction and inactivating mutants impair biological activity. Dev. Reprod. 2021, 25, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Byambaragchaa, M.; Choi, S.H.; Kim, D.W.; Min, K.S. Cell-surface loss of constitutive activating and inactivating mutants of eel luteinizing hormone receptors. Dev. Reprod. 2021, 25, 225–234. [Google Scholar] [CrossRef]
- Byambaragchaa, M.; Joo, H.E.; Kim, S.K.; Kim, Y.J.; Park, G.E.; Min, K.S. Signal transduction of C-terminal phosphorylation regions for equine luteinizing hormone/chorionic gonadotropin receptor (eLH/CGR). Dev. Reprod. 2022, 26, 1–12. [Google Scholar] [CrossRef]
- Byambaragchaa, M.; Kim, D.J.; Kang, M.H.; Min, K.S. Site specificity of eel luteinizing hormone N-linked oligosaccharides in signal transduction. Gen. Comp. Endocrinol. 2018, 268, 50–56. [Google Scholar] [CrossRef]
- Min, K.S.; Hiyama, T.; Seong, H.W.; Hattori, N.; Tanaka, S.; Shiota, K. Biological activities of tethered equine chorionic gonadotropin (eCG) and its deglycosylated mutants. J. Reprod. Dev. 2004, 50, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Min, K.S.; Park, J.J.; Byambaragchaa, M.; Kang, M.H. Characterization of tethered equine chorionic gonadotropin and its deglycosylated mutants by ovulation stimulation in mice. BMC Biotechnol. 2019, 19, 60. [Google Scholar] [CrossRef] [Green Version]
- Min, K.S.; Park, J.J.; Lee, S.Y.; Byambaragchaa, M.; Kang, M.H. Comparative gene expression profiling of mouse ovaries upon stimulation with natural equine chorionic gonadotropin (N-eCG) and tethered recombinant-eCG (R-eCG). BMC Biotechnol. 2020, 20, 59. [Google Scholar] [CrossRef]
- Lee, S.Y.; Byambaragchaa, M.; Choi, S.H.; Kang, H.J.; Kang, M.H.; Min, K.S. Roles of N-linked and O-linked glycosylation sites in the activity of equine chorionic gonadotropin in cells expressing rat luteinizing hormone/chorionic gonadotropin receptor and follicle-stimulating hormone receptor. BMC Biotechnol. 2021, 21, 52. [Google Scholar] [CrossRef]
- Byambaragchaa, M.; Park, A.; Gil, S.J.; Lee, H.W.; Ko, Y.J.; Choi, S.H.; Kang, M.H.; Min, K.S. Luteinizing hormone-like and follicle-stimulating hormone-like activities of equine chorionic gonadotropin β-subunit mutants in cells expressing rat luteinizing hormone/chorionic gonadotropin receptor and rat follicle-stimulating hormone receptor. Anim. Cells Syst. 2021, 25, 171–181. [Google Scholar] [CrossRef]
- Ji, I.; Slaughter, R.G.; Ji, T.H. N-linked oligosaccharides are not required for hormone binding of the lutropin receptor in a Leydig tumor cell line and rat granulosa cells. Endocrinology 1990, 127, 494–496. [Google Scholar] [CrossRef]
- Liu, X.; Davis, D.; Segaloff, D.L. Distribution of potential sites for N-linked glycosylation does not impair hormone binding to the lutropin/choriogonadotropin receptor if Asn-173 is left intact. J. Biol. Chem. 1993, 268, 1513–1516. [Google Scholar] [CrossRef]
- Davis, D.P.; Liu, X.; Segaloff, D.L. Identification of the sites of N-linked glycosylation on the follicle-stimulating hormone (FSH) receptor and assessment of their role in the FSH receptor function. Mol. Endocrinol. 1995, 9, 159–170. [Google Scholar]
- Davis, D.P.; Rozell, T.G.; Liu, X.; Segaloff, D.L. The six-N-linked carbohydrates of the lutropin/choriogonadotropin receptor are not absolutely required for correct folding, cell surface expression, hormone binding, or signal transduction. Mol. Endocrinol. 1997, 11, 550–562. [Google Scholar] [CrossRef]
- Davis, D.P.; Segaloff, D.L. N-linked carbohydrates on G protein-coupled receptors: Mapping sites of attachment and determining functional roles. Methods Enzymol. 2002, 343, 200–212. [Google Scholar]
- Russo, D.; Chazenbalk, G.D.; Nagayama, Y.; Wadsworth, H.L.; Rapoport, B. Site-directed mutagenesis of the human thyrotropin receptor: Role of asparagine-linked oligosaccharides in the expression of a functional receptor. Mol. Endocrinol. 1991, 5, 29–33. [Google Scholar] [CrossRef] [Green Version]
- Kakinuma, A.; Chazenbalk, G.D.; Tanaka, K.; Nagayama, Y.; Mclachlan, S.M.; Rapoport, B. An N-linked glycosylation motif from the noncleaving luteinizing hormone receptor substituted for the homologous region (Gly369 to Glu369) of the thyrotropin receptor prevents cleavage at its second, downstream site. J. Biol. Chem. 1997, 272, 28296–282300. [Google Scholar] [CrossRef]
- Keinanen, K.P. Effect of deglycosylation on the structure and hormone-binding activity of the lutropin receptor. Biochem. J. 1988, 256, 719–724. [Google Scholar] [CrossRef] [Green Version]
- Rajaniemi, H.J.; Petaja-Repo, U.E.; Pietila, E.M. Structure and functional significance of the carbohydrates of the LH/CG receptor. Mol. Cell Endocrinol. 1996, 125, 101–105. [Google Scholar] [CrossRef]
- Wang, T.; Nakagawa, S.; Miyake, T.; Setsu, G.; Kunisue, S.; Goto, K.; Hirawawa, A.; Okamura, H.; Yamaguchi, Y.; Doi, M. Identification and functional characterization of N-linked glycosylation of the orphan G protein-coupled receptor Gpr176. Sci. Rep. 2020, 10, 4429. [Google Scholar] [CrossRef] [Green Version]
- Goth, C.K.; Petaja-Repo, U.E.; Rosenkilde, M.M. G protein-coupled receptors in the sweet spot: Glycosylation and other post-translational modifications. ACS Pharmacol. Transl. Sci. 2020, 3, 237–245. [Google Scholar] [CrossRef]
- Rowland, R.; Brandariz-Nunez, A. Analysis of the role of N-linked glycosylation in cell surface expression, function, and binding properties of SARS-CoV-2 receptor ACE2. Microbiol. Spectr. 2021, 9, e0119921. [Google Scholar] [CrossRef]
- Pang, R.T.; Ng, S.S.; Cheng, C.H.; Holtmann, M.H.; Miller, L.J.; Chow, B.K. Role of N-linked glycosylation on the function and expression of the human secretin receptor. Endocrinology 1999, 140, 5102–5111. [Google Scholar] [CrossRef]
- Norskov-lauritsen, L.; Jorgensen, S.; Brauner-Osborne, H. N-glycosylation and disulfide bonding affects GPRC6A receptor expression, function, and dimerization. FEBS Lett. 2015, 589, 588–597. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.A.; Dupakuntla, M.; Pathak, B.R.; Mahale, S.D. FSH receptor-specific residue L501 and I505 in extracellular loop 2 are essential for its function. J. Mol. Endocrinol. 2015, 54, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.A.; Mahale, S.D. Extracellular loop 3 substitutions k589N and A590S in FSH receptor increase FSH-induced receptor internalization and along with S588T substitution exhibit impaired ERK1/2 phosphorylation. Arch. Biochem. Biophys. 2018, 659, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Cottom, J.; Salvador, L.M.; Maizels, E.T.; Reierstad, S.; Park, Y.; Carr, D.W.; Davare, M.A.; Hell, J.W.; Palmer, S.S.; Dent, P.; et al. Follicle-stimulating hormone activates extracellular signal-regulated kinase but not extracellular signal-regulated kinase kinase through a 100-kDa phosphotyrosine phosphatase. J. Biol. Chem. 2003, 278, 7167–7179. [Google Scholar] [CrossRef] [PubMed]
- Kara, E.; Crepieux, P.; Gauthier, C.; Martinat, N.; Piketty, V.; Guillou, F.; Reiter, E. A phosphorylation cluster of five serine and threonine residues in the C-terminus of the follicle-stimulating hormone receptor is important for desensitization but not for β-arrestin-mediated ERK activation. Mol. Endocrinol. 2006, 20, 3014–3026. [Google Scholar] [CrossRef] [PubMed]
- Luttrell, L.M.; Lefkowitz, R.J. The role of beta-arrestins in the termination and transduction of G-protein-coupled receptor signals. J. Cell Sci. 2002, 115, 455–465. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| cAMP Responses | |||
|---|---|---|---|
| eel FSH Receptors | Basal a (nM/104 Cells) | EC50 (ng/mL) | Rmax b (nM/104 Cells) |
| eel FSHR-wt | 0.2 ± 0.1 | 201.1 (1.0-fold) (165.2 to 256.8) c | 57.5 ± 2.0 (100%) |
| eel FSHR- N120Q | 0.1 ± 0.1 | 283.3 (0.70-fold) (199.2 to 490.4) | 25.9 ± 1.9 (45.0%) |
| eel FSHR- N191Q | 0.2 ± 0.1 | - d | 3.1 ± 0.1 (0.05%) |
| eel FSHR- N272Q | 0.3 ± 0.1 | 326.8 (0.61-fold) (284.2 to 384.6) | 19.2 ± 0.4 (33.4%) |
| eel FSHR- N288Q | 0.4 ± 0.2 | 281.0 (0.71-fold) (218.4 to 394.0) | 21.4 ± 0.3 (37.2%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byambaragchaa, M.; Park, H.-K.; Kim, D.-J.; Lee, J.-H.; Kang, M.-H.; Min, K.-S. The N-Linked Glycosylation Site N191 Is Necessary for PKA Signal Transduction in Eel Follicle-Stimulating Hormone Receptor. Int. J. Mol. Sci. 2022, 23, 12792. https://doi.org/10.3390/ijms232112792
Byambaragchaa M, Park H-K, Kim D-J, Lee J-H, Kang M-H, Min K-S. The N-Linked Glycosylation Site N191 Is Necessary for PKA Signal Transduction in Eel Follicle-Stimulating Hormone Receptor. International Journal of Molecular Sciences. 2022; 23(21):12792. https://doi.org/10.3390/ijms232112792
Chicago/Turabian StyleByambaragchaa, Munkhzaya, Hong-Kyu Park, Dae-Jung Kim, Jong-Hyuk Lee, Myung-Hwa Kang, and Kwan-Sik Min. 2022. "The N-Linked Glycosylation Site N191 Is Necessary for PKA Signal Transduction in Eel Follicle-Stimulating Hormone Receptor" International Journal of Molecular Sciences 23, no. 21: 12792. https://doi.org/10.3390/ijms232112792
APA StyleByambaragchaa, M., Park, H.-K., Kim, D.-J., Lee, J.-H., Kang, M.-H., & Min, K.-S. (2022). The N-Linked Glycosylation Site N191 Is Necessary for PKA Signal Transduction in Eel Follicle-Stimulating Hormone Receptor. International Journal of Molecular Sciences, 23(21), 12792. https://doi.org/10.3390/ijms232112792