
Cluster Analysis of Short Sensory Profile Data Reveals Sensory-Based Subgroups in Autism Spectrum Disorder


Abstract
:1. Introduction
2. Results
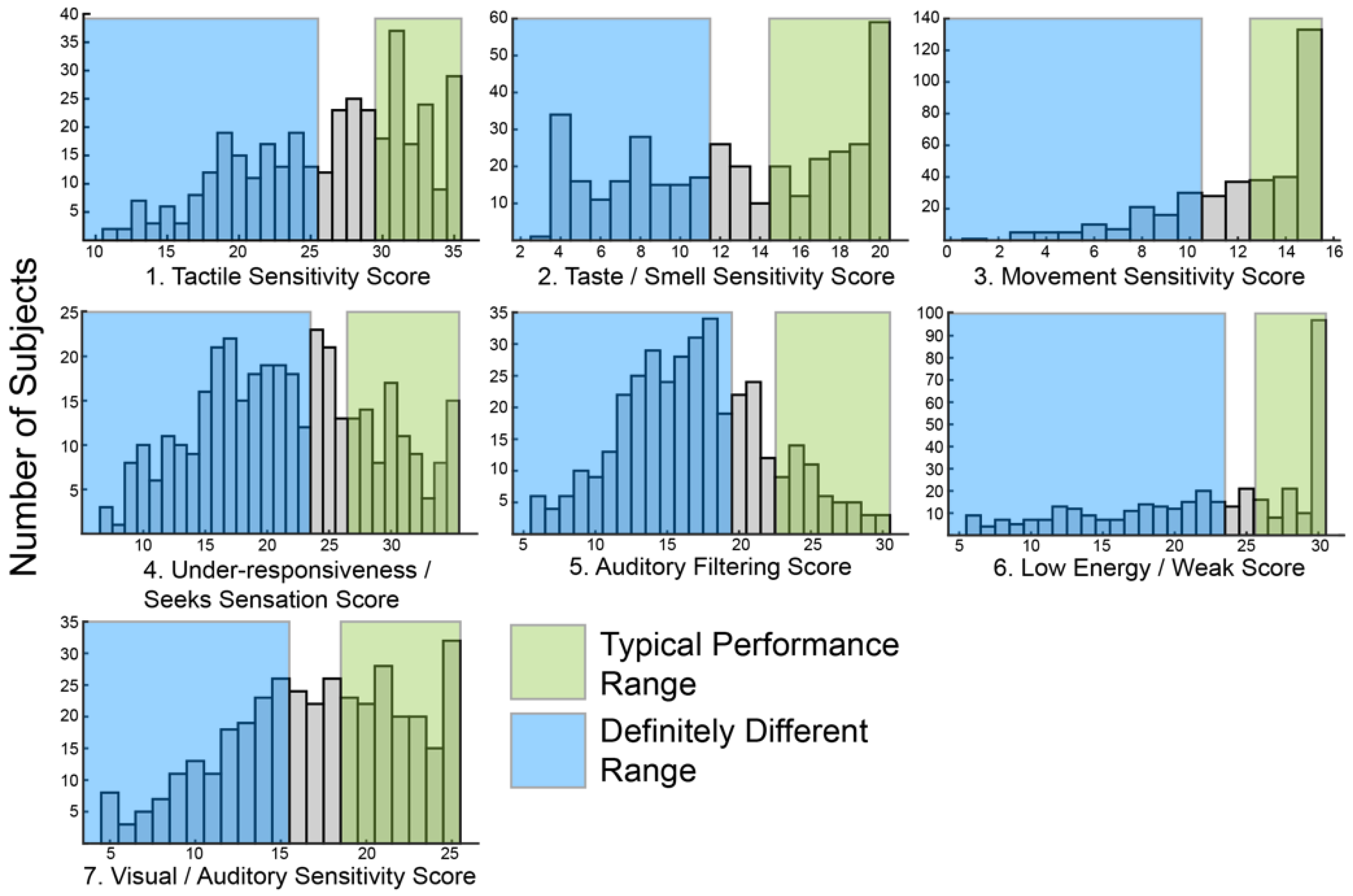
2.1. Short Sensory Profile Responses in ASD Are Heterogeneous
2.2. Cluster Analysis Optimizes at Six Sensory Based Subgroups
2.3. Subgroups Are Characterized by Unique Patterns of Genetic Variants
3. Discussion
3.1. Sensory Features Can Be Used to Identify Unique Subgroups in ASD
3.2. Genes Enriched within Each Sensory Subgroup Are Relevant to ASD
3.3. Limitations
4. Materials and Methods
4.1. Procedures
4.2. Data Analysis
4.3. Variant Extraction and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-5, 5th ed.; American Psychiatric Publishing: Arlington, VA, USA, 2013. [Google Scholar]
- Lyall, K.; Croen, L.; Daniels, J.; Fallin, M.D.; Ladd-Acosta, C.; Lee, B.K.; Park, B.Y.; Snyder, N.W.; Schendel, D.; Volk, H.; et al. The Changing Epidemiology of Autism Spectrum Disorders. Annu. Rev. Public Health 2017, 38, 81–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, A.J.; Brugha, T.S.; Erskine, H.E.; Scheurer, R.W.; Vos, T.; Scott, J.G. The epidemiology and global burden of autism spectrum disorders. Psychol. Med. 2015, 45, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Kogan, M.D.; Strickland, B.B.; Blumberg, S.J.; Singh, G.K.; Perrin, J.M.; van Dyck, P.C. A national profile of the health care experiences and family impact of autism spectrum disorder among children in the United States 2005–2006. Pediatrics 2008, 122, e1149–e1158. [Google Scholar] [CrossRef] [PubMed]
- Pandina, G.; Ring, R.H.; Bangerter, A.; Ness, S. Current Approaches to the Pharmacologic Treatment of Core Symptoms Across the Lifespan of Autism Spectrum Disorder. Psychiatr. Clin. N. Am. 2020, 43, 629–645. [Google Scholar] [CrossRef] [PubMed]
- Betancur, C. Etiological heterogeneity in autism spectrum disorders: More than 100 genetic and genomic disorders and still counting. Brain Res. 2011, 1380, 42–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeste, S.S.; Geschwind, D.H. Disentangling the heterogeneity of autism spectrum disorder through genetic findings. Nat. Rev. Neurol. 2014, 10, 74–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingram, D.G.; Takahashi, T.N.; Miles, J.H. Defining autism subgroups: A taxometric solution. J. Autism Dev. Disord. 2008, 38, 950–960. [Google Scholar] [CrossRef]
- Jacob, S.; Wolff, J.J.; Steinbach, M.S.; Doyle, C.B.; Kumar, V.; Elison, J.T. Neurodevelopmental heterogeneity and computational approaches for understanding autism. Transl. Psychiatry 2019, 9, 63. [Google Scholar] [CrossRef] [Green Version]
- Nordahl, C.W.; Andrews, D.S.; Dwyer, P.; Waizbard-Bartov, E.; Restrepo, B.; Lee, J.K.; Heath, B.; Saron, C.; Rivera, S.M.; Solomon, M.; et al. The Autism Phenome Project: Toward Identifying Clinically Meaningful Subgroups of Autism. Front. Neurosci. 2022, 15, 786220. [Google Scholar] [CrossRef]
- Wolfers, T.; Floris, D.L.; Dinga, R.; van Rooij, D.; Isakoglou, C.; Kia, S.M.; Zabihi, M.; Llera, A.; Chowdanayaka, R.; Kumar, V.J.; et al. From pattern classification to stratification: Towards conceptualizing the heterogeneity of Autism Spectrum Disorder. Neurosci. Biobehav. Rev. 2019, 104, 240–254. [Google Scholar] [CrossRef] [PubMed]
- Posar, A.; Visconti, P. Sensory abnormalities in children with autism spectrum disorder. J. Pediatr. (Rio J.) 2018, 94, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.E.; Molloy, C.A.; Bishop, S.L. Classification of children with autism spectrum disorder by sensory subtype: A case for sensory-based phenotypes. Autism Res. 2014, 7, 322–333. [Google Scholar] [CrossRef] [PubMed]
- Heald, M.; Adams, D.; Oliver, C. Profiles of atypical sensory processing in Angelman, Cornelia de Lange and Fragile X syndromes). J. Intellect. Disabil. Res. 2020, 64, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Mieses, A.M.; Tavassoli, T.; Li, E.; Soorya, L.; Lurie, S.; Wang, A.T.; Siper, P.M.; Kolevzon, A. Brief Report: Sensory Reactivity in Children with Phelan-McDermid Syndrome. J. Autism Dev. Disord. 2016, 46, 2508–2513. [Google Scholar] [CrossRef] [PubMed]
- Tavassoli, T.; Layton, C.; Levy, T.; Rowe, M.; George-Jones, J.; Zweifach, J.; Lurie, S.; Buxbaum, J.D.; Kolevzon, A.; Siper, P.M. Sensory Reactivity Phenotype in Phelan-McDermid Syndrome Is Distinct from Idiopathic ASD. Genes 2021, 12, 977. [Google Scholar] [CrossRef] [PubMed]
- Lyons-Warren, A.M.; McCormack, M.C.; Holder, J.L., Jr. Sensory Processing Phenotypes in Phelan-McDermid Syndrome and SYNGAP1-Related Intellectual Disability. Brain Sci. 2022, 12, 137. [Google Scholar] [CrossRef] [PubMed]
- Kirby, A.V.; Bilder, D.A.; Wiggins, L.D.; Hughes, M.M.; Davis, J.; Hall-Lande, J.A.; Lee, L.C.; McMahon, W.M.; Bakian, A.V. Sensory features in autism: Findings from a large population-based surveillance system. Autism Res. 2022, 15, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Klintwall, L.; Holm, A.; Eriksson, M.; Carlsson, L.H.; Olsson, M.B.; Hedvall, A.; Gillberg, C.; Fernell, E. Sensory abnormalities in autism. A brief report. Res. Dev. Disabil. 2011, 32, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Simpson, K.; Adams, D.; Alston-Knox, C.; Heussler, H.S.; Keen, D. Exploring the Sensory Profiles of Children on the Autism Spectrum Using the Short Sensory Profile-2 (SSP-2). J. Autism Dev. Disord. 2019, 49, 2069–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheerer, N.E.; Curcin, K.; Stojanoski, B.; Anagnostou, E.; Nicolson, R.; Kelley, E.; Georgiades, S.; Liu, X.; Stevenson, R.A. Exploring Sensory Phenotypes in Autism Spectrum Disorder. Mol. Autism. 2021, 12, 67. [Google Scholar] [CrossRef]
- Yuen, R.K.C.; Merico, D.; Bookman, M.; LHowe, J.; Thiruvahindrapuram, B.; Patel, R.V.; Whitney, J.; Deflaux, N.; Bingham, J.; Wang, Z.; et al. Whole genome sequencing resource identifies 18 new candidate genes for autism spectrum disorder. Nat. Neurosci. 2017, 20, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, J.; Ziviani, J.; Rodger, S. Sensory processing and classroom emotional, behavioral, and educational outcomes in children with autism spectrum disorder. Am. J. Occup. 2008, 62, 564–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.H.; Rodgers, J.; McConachie, H. Restricted and repetitive behaviours, sensory processing and cognitive style in children with autism spectrum disorders. J. Autism Dev. Disord. 2009, 39, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Rogers, S.J.; Hepburn, S.; Wehner, E. Parent reports of sensory symptoms in toddlers with autism and those with other developmental disorders. J. Autism Dev. Disord. 2003, 33, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Tomchek, S.D.; Dunn, W. Sensory processing in children with and without autism: A comparative study using the short sensory profile. Am. J. Occup. 2007, 61, 190–200. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, D.N.; Miller, L.J.; Shyu, V. Development and validation of the Short Sensory Profile. In Sensory Profile Manual; Dunn, W., Ed.; Psychological Corporation: San Antonio, TX, USA, 1999; pp. 59–73. [Google Scholar]
- Robak, L.A.; Jansen, I.E.; van Rooij, J.; Uitterlinden, A.G.; Kraaij, R.; Jankovic, J.; International Parkinson’s Disease Genomics Consortium (IPDGC); Heutink, P.; Shulman, J.M. Excessive burden of lysosomal storage disorder gene variants in Parkinson’s disease. Brain 2017, 140, 3191–3203. [Google Scholar] [CrossRef] [Green Version]
- Uljarević, M.; Lane, A.; Kelly, A.; Leekam, S. Sensory subtypes and anxiety in older children and adolescents with autism spectrum disorder. Autism Res. 2016, 9, 1073–1078. [Google Scholar] [CrossRef]
- Ausderau, K.K.; Furlong, M.; Sideris, J.; Bulluck, J.; Little, L.M.; Watson, L.R.; Boyd, B.A.; Belger, A.; Dickie, V.A.; Baranek, G.T. Sensory subtypes in children with autism spectrum disorder: Latent profile transition analysis using a national survey of sensory features. J. Child Psychol. Psychiatry 2014, 55, 935–944. [Google Scholar] [CrossRef] [Green Version]
- Ben-Sasson, A.; Cermak, S.A.; Orsmond, G.I.; Tager-Flusberg, H.; Kadlec, M.B.; Carter, A.S. Sensory clusters of toddlers with autism spectrum disorders: Differences in affective symptoms. J. Child Psychol. Psychiatry 2008, 49, 817–825. [Google Scholar] [CrossRef]
- Lane, A.E.; Young, R.L.; Baker, A.E.; Angley, M.T. Sensory processing subtypes in autism: Association with adaptive behavior. J. Autism Dev. Disord. 2010, 40, 112–122. [Google Scholar] [CrossRef]
- Lane, A.; Dennis, S.J.; Geraghty, M.E. Brief report: Further evidence of sensory subtypes in autism. J. Autism Dev. Disord. 2011, 41, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Liss, M.; Saulnier, C.; Fein, D.; Kinsbourne, M. Sensory and attention abnormalities in autistic spectrum disorders. Autism 2006, 10, 155–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, L.M.; Dean, E.; Tomchek, S.D.; Dunn, W. Classifying sensory profiles of children in the general population. Child Care Health Dev. 2017, 43, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Tomchek, S.D.; Little, L.M.; Myers, J.; Dunn, W. Sensory Subtypes in Preschool Aged Children with Autism Spectrum Disorder. J. Autism Dev. Disord. 2018, 48, 2139–2147. [Google Scholar] [CrossRef]
- Genovese, A.; Butler, M.G. Clinical Assessment, Genetics, and Treatment Approaches in Autism Spectrum Disorder (ASD). Int. J. Mol. Sci. 2020, 21, 4726. [Google Scholar] [CrossRef]
- Eaves, L.C.; Ho, H.H.; Eaves, D.M. Subtypes of autism by cluster analysis. J. Autism Dev. Disord. 1994, 24, 3–22. [Google Scholar] [CrossRef]
- Qi, S.; Morris, R.; Turner, J.A.; Fu, Z.; Jiang, R.; Deramus, T.P.; Zhi, D.; Calhoun, V.D.; Sui, J. Common and unique multimodal covarying patterns in autism spectrum disorder subtypes. Mol. Autism 2020, 11, 90. [Google Scholar] [CrossRef]
- Hu, V.W.; Bi, C. Phenotypic Subtyping and Re-analyses of Existing Transcriptomic Data from Autistic Probands in Simplex Families Reveal Differentially Expressed and ASD Trait-Associated Genes. Front. Neurol. 2020, 11, 578972. [Google Scholar] [CrossRef]
- Amadesi, S.; Grant, A.D.; Cottrell, G.S.; Vaksman, N.; Poole, D.P.; Rozengurt, E.; Bunnett, N.W. Protein kinase D isoforms are expressed in rat and mouse primary sensory neurons and are activated by agonists of protease-activated receptor 2. J. Comp. Neurol. 2009, 516, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Haeberle, H.; Bryan, L.A.; Vadakkan, T.J.; Dickinson, M.E.; Lumpkin, E.A. Swelling-activated Ca2+ channels trigger Ca2+ signals in Merkel cells. PLoS ONE 2008, 3, e1750. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Ehlers, M.D. Modeling autism by SHANK gene mutations in mice. Neuron 2013, 78, 8–27. [Google Scholar] [CrossRef] [Green Version]
- Sungur, A.Ö.; Schwarting, R.K.W.; Wöhr, M. Behavioral phenotypes and neurobiological mechanisms in the Shank1 mouse model for autism spectrum disorder: A translational perspective. Behav. Brain Res. 2018, 352, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Silverman, J.L.; Turner, S.M.; Barkan, C.L.; Tolu, S.S.; Saxena, R.; Hung, A.Y.; Sheng, M.; Crawley, J.N. Sociability and motor functions in Shank1 mutant mice. Brain Res. 2011, 1380, 120–137. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Chen, W.; Myers, S.J.; Yuan, H.; Traynelis, S.F. Human GRIN2B variants in neurodevelopmental disorders. J. Pharm. Sci. 2016, 132, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwyer, P.; Saron, C.D.; Rivera, S.M. Identification of Longitudinal Sensory Subtypes in Typical Development and Autism Spectrum Development Using Growth Mixture Modelling. Res. Autism Spectr. Disord. 2020, 78, 101645. [Google Scholar] [CrossRef] [PubMed]
- Dunn, W. The impact of sensory processing abilities on the daily lives of young children and their families: A conceptual model. Infants Young Child. 1997, 9, 23–35. [Google Scholar] [CrossRef]
- Dunn, W. Sensory Profile-2; Pearson Publishing: San Antonio, TX, USA, 2014. [Google Scholar]
- Bruni, M.; Cameron, D.; Dua, S.; Noy, S. Reported sensory processing of children with Down syndrome. Phys. Occup. Pediatr. 2010, 30, 280–293. [Google Scholar] [CrossRef] [PubMed]
- Carr, J.L.; Agnihotri, S.; Keightley, M. Sensory processing and adaptive behavior deficits of children across the fetal alcohol spectrum disorder continuum. Alcohol. Clin. Exp. Res. 2010, 34, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- John, A.E.; Mervis, C.B. Sensory modulation impairments in children with Williams syndrome. Am. J. Med. Genet C Semin. Med Genet. 2010, 154C, 266–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangeot, S.D.; Miller, L.J.; McIntosh, D.N.; McGrath-Clarke, J.; Simon, J.; Hagerman, R.J.; Goldson, E. Sensory modulation dysfunction in children with attention-deficit-hyperactivity disorder. Dev. Med. Child Neurol. 2001, 43, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Kircher, M.; Witten, D.M.; Jain, P.; O’Roak, B.J.; Cooper, G.M.; Shendure, J. A general framework for estimating the relative pathogenicity of human genetic variants. Nat. Genet. 2014, 46, 310–315. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | N |
|---|---|
| Definitely or probably different in all areas | 54 |
| Definitely or probably different in all areas EXCEPT Low Energy/Weak | 15 |
| Definitely or probably different in all areas EXCEPT Taste/Smell | 14 |
| Definitely or probably different in all areas EXCEPT Movement | 13 |
| Definitely or probably different in: Visual/Auditory, Taste/Smell, Under-responsiveness/Seeks Sensation, Tactile and Auditory Filtering | 13 |
| Definitely or probably different in: Under-responsiveness/Seeks Sensation, and Auditory Filtering | 13 |
| Definitely or probably different in: Under-responsiveness/Seeks Sensation, Low Energy/Weak, and Auditory Filtering | 11 |
| Definitely or probably different in: Under-responsiveness/Seeks Sensation, Tactile, and Auditory Filtering | 10 |
| Definitely or probably different just in Auditory Filtering | 10 |
| Typical performance in all areas | 9 |
| Sensory Domain | SSP Question Number and Text | |
|---|---|---|
| Vision Hypersensitivity | 36 | Is bothered by bright lights after others have adapted to the light |
| 38 | Covers eyes or squints to protect eyes from light | |
| Auditory Hypersensitivity | 34 | Responds negatively to unexpected or loud noises (for example, cries or hides at noise from vacuum cleaner, dog barking or hair dryer) |
| 35 | Holds hands over ears to protect ears from sound | |
| Auditory Hyposensitivity | 23 | Appears to not hear what you say (for example, does not “tune-in” to what you say, appears to ignore you) |
| 26 | Doesn’t respond when name is called but you know the child’s hearing is OK | |
| Tactile Hypersensitivity | 1 | Expresses distress during grooming (for example, fights or cries during haircutting, face washing, fingernail cutting) |
| 3 | Avoids going barefoot, especially in sand or grass | |
| 4 | Reacts aggressively or emotionally to touch | |
| 5 | Withdrawals from splashing water | |
| 7 | Rubs or scratches out a spot that has been touched | |
| Tactile Hyposensitivity | 18 | Touches people and objects |
| 19 | Doesn’t seem to notice when face or hands are messy | |
| Taste Hypersensitivity | 8 | Avoids certain tastes or food smells that are typically part of children’s diet |
| 11 | Picky eater, especially regarding food textures | |
| Subgroup | Total Number | Male N (%) | European Ancestry N (%) | Mean Age Years | Mean Adaptive Behavior Score | Mean Socialization Score | Full Scale IQ Mean (N) |
|---|---|---|---|---|---|---|---|
| 1 | 53 | 45 (84.9) | 39 (73.6) | 10.3 | 64 | 67 | 89 (33) |
| 2 | 72 | 58 (80.5) | 48 (66.7) | 10.0 | 73 | 76 | 80 (39) |
| 3 | 63 | 51 (80.9) | 46 (73.0) | 8.0 | 66 | 67 | 80 (45) |
| 4 | 62 | 55 (88.7) | 42 (67.7) | 9.8 | 66 | 70 | 73 (36) |
| 5 | 67 | 45 (67.1) | 49 (73.1) | 9.1 | 69 | 72 | 80 (44) |
| 6 | 60 | 48 (80.0) | 47 (78.3) | 12.1 | 67 | 69 | 77 (29) |
| Total | 377 | 302 (80.1) | 272 (72.0) | 9.87 | 68 | 70 | 80 (226) |
| Subgroup | Gene | Variant Frequency | Difference | Gene Card Summary | Gene Ontology | |
|---|---|---|---|---|---|---|
| Tactile and Auditory | 3 | THSD7B | 0.69 | 0.22 | Glycosylation | Protein binding |
| 3 | GRAMD1B | 0.45 | 0.21 | Cholesterol transport | Phosphatidylserine, lipid, cholesterol and phosphatidic acid binding; intermembrane cholesterol transfer activity | |
| 3 | ABCA12 | 0.45 | 0.20 | Transporter | signaling receptor binding and ATPase activity, coupled to transmembrane movement of substances | |
| 3 | ADAMTS18 | 0.42 | 0.17 | Metalloproteinase | peptidase activity and metalloendopeptidase activity | |
| 3 | ELN | 0.23 | 0.15 | Extracellular matrix | extracellular matrix structural constituent and extracellular matrix constituent conferring elasticity | |
| 3 | JPH3 | 0.29 | 0.15 | Junctional Complexes | calcium-release channel activity | |
| 3 | TMEFF2 | 0.47 | 0.15 | Oncogene | Protein binding | |
| 3 | OLA1 | 0.24 | −0.15 | GTPase protein family | GTP binding and ribosome binding | |
| 3 | PAPPA | 0.26 | −0.20 | Metalloproteinase | metalloendopeptidase activity and endopeptidase activity | |
| Tactile and Auditory Hyposensitivity | 4 | PKD1 | 0.41 | 0.16 | Glycoprotein | protein kinase binding and protein domain specific binding |
| 4 | KDM4C | 0.48 | 0.15 | Demethylase | enzyme binding and dioxygenase activity | |
| 4 | MGMT | 0.29 | −0.14 | Transfer of methyl groups | calcium ion binding and damaged DNA binding | |
| 4 | TNXB | 0.16 | −0.16 | Extracellular matrix | heparin binding and collagen binding | |
| 4 | SHANK1 | 0.17 | −0.19 | Scaffold proteins | Identical protein binding, Protein C-terminus binding | |
| Mixed | 6 | GRIN2B | 0.553571 | 0.159151 | NMDA-R subunit | calcium channel activity and ionotropic glutamate receptor activity |
| 6 | HNRNPUL2 | 0.214286 | 0.140732 | Nuclear Ribonucleoprotein | kinase activity | |
| 6 | EPHB1 | 0.303571 | −0.1433 | Receptor Tyrosine Kinase | transferase activity, transferring phosphorus-containing groups and protein tyrosine kinase activity | |
| 6 | LRPPRC | 0.142857 | −0.14576 | Mitochondrial | ubiquitin protein ligase binding | |
| 6 | SORL1 | 0.125 | −0.14501 | Vacuolar sorting protein LDL receptor | transmembrane signaling receptor activity and low-density lipoprotein particle binding | |
| 6 | MEIS1 | 0.428571 | −0.16274 | Homeobox | sequence-specific DNA binding and chromatin binding | |
| 6 | SSH2 | 0.160714 | −0.16829 | Tyrosine phosphatase | actin binding and protein tyrosine phosphatase activity | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyons-Warren, A.M.; Wangler, M.F.; Wan, Y.-W. Cluster Analysis of Short Sensory Profile Data Reveals Sensory-Based Subgroups in Autism Spectrum Disorder. Int. J. Mol. Sci. 2022, 23, 13030. https://doi.org/10.3390/ijms232113030
Lyons-Warren AM, Wangler MF, Wan Y-W. Cluster Analysis of Short Sensory Profile Data Reveals Sensory-Based Subgroups in Autism Spectrum Disorder. International Journal of Molecular Sciences. 2022; 23(21):13030. https://doi.org/10.3390/ijms232113030
Chicago/Turabian StyleLyons-Warren, Ariel M., Michael F. Wangler, and Ying-Wooi Wan. 2022. "Cluster Analysis of Short Sensory Profile Data Reveals Sensory-Based Subgroups in Autism Spectrum Disorder" International Journal of Molecular Sciences 23, no. 21: 13030. https://doi.org/10.3390/ijms232113030
APA StyleLyons-Warren, A. M., Wangler, M. F., & Wan, Y. -W. (2022). Cluster Analysis of Short Sensory Profile Data Reveals Sensory-Based Subgroups in Autism Spectrum Disorder. International Journal of Molecular Sciences, 23(21), 13030. https://doi.org/10.3390/ijms232113030