
Social Hierarchy Dictates Intestinal Radiation Injury in a Gut Microbiota-Dependent Manner

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
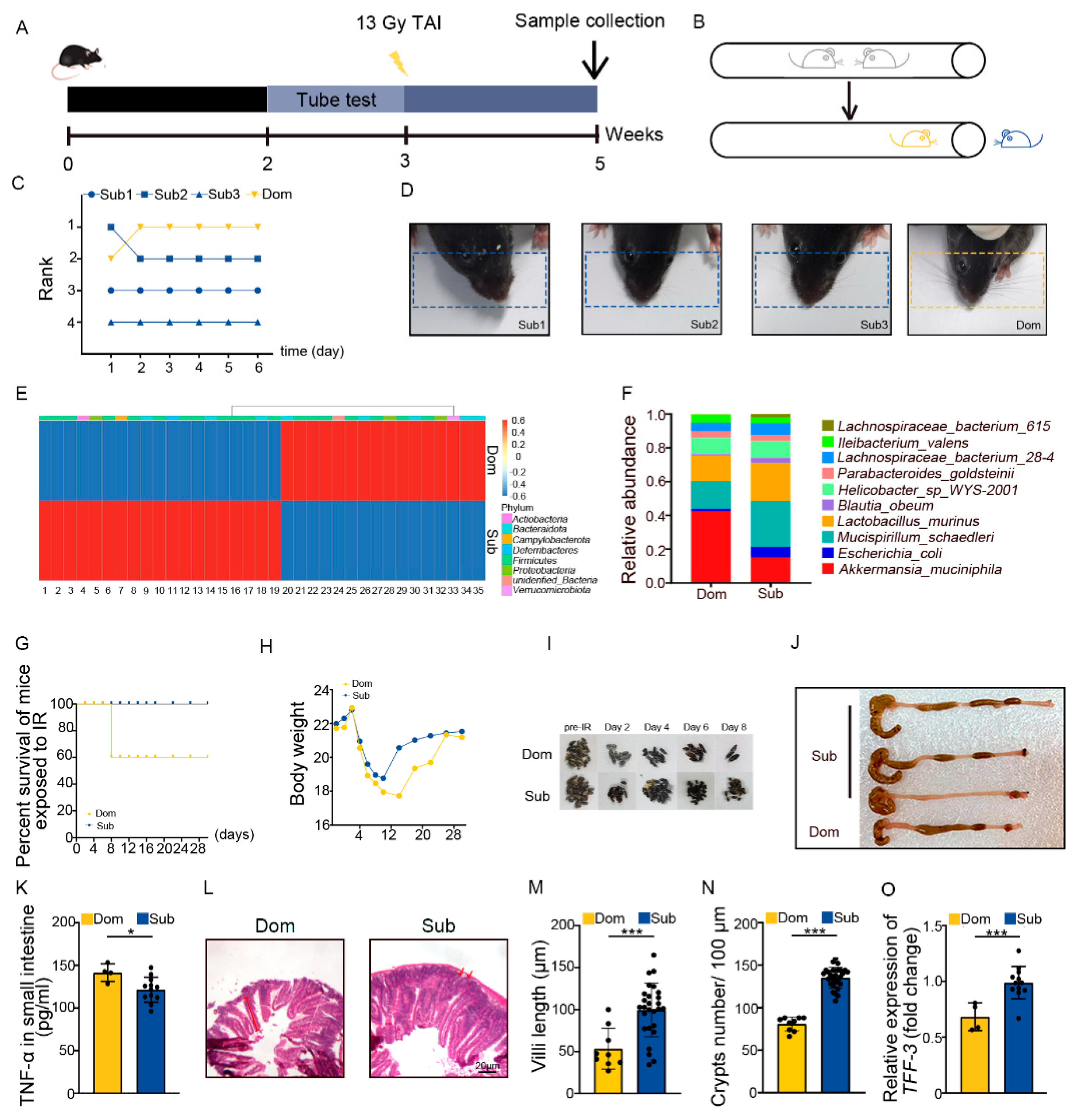
2.1. Social Hierarchy Determines the Degree of Intestinal Radiation Injury
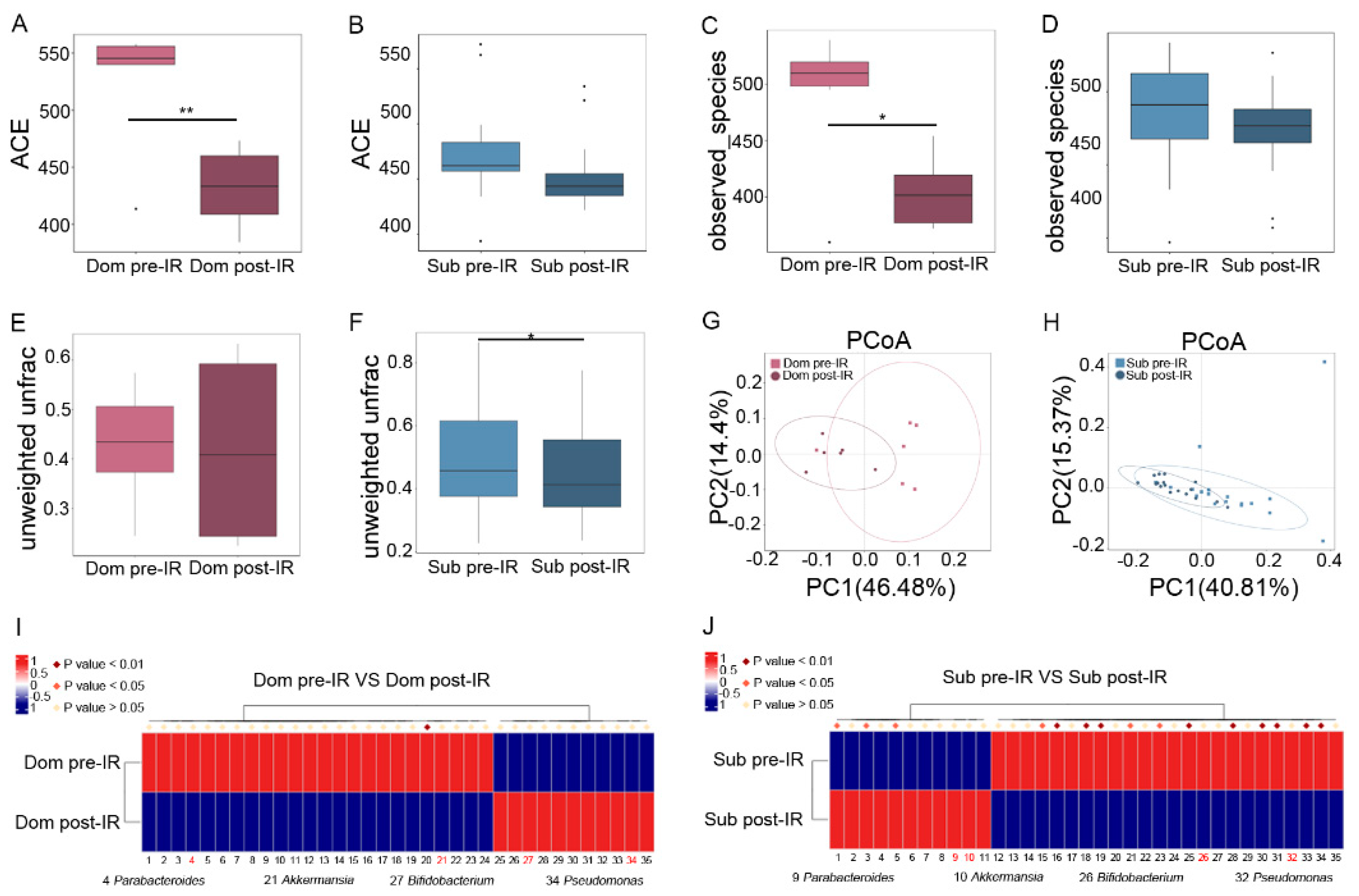
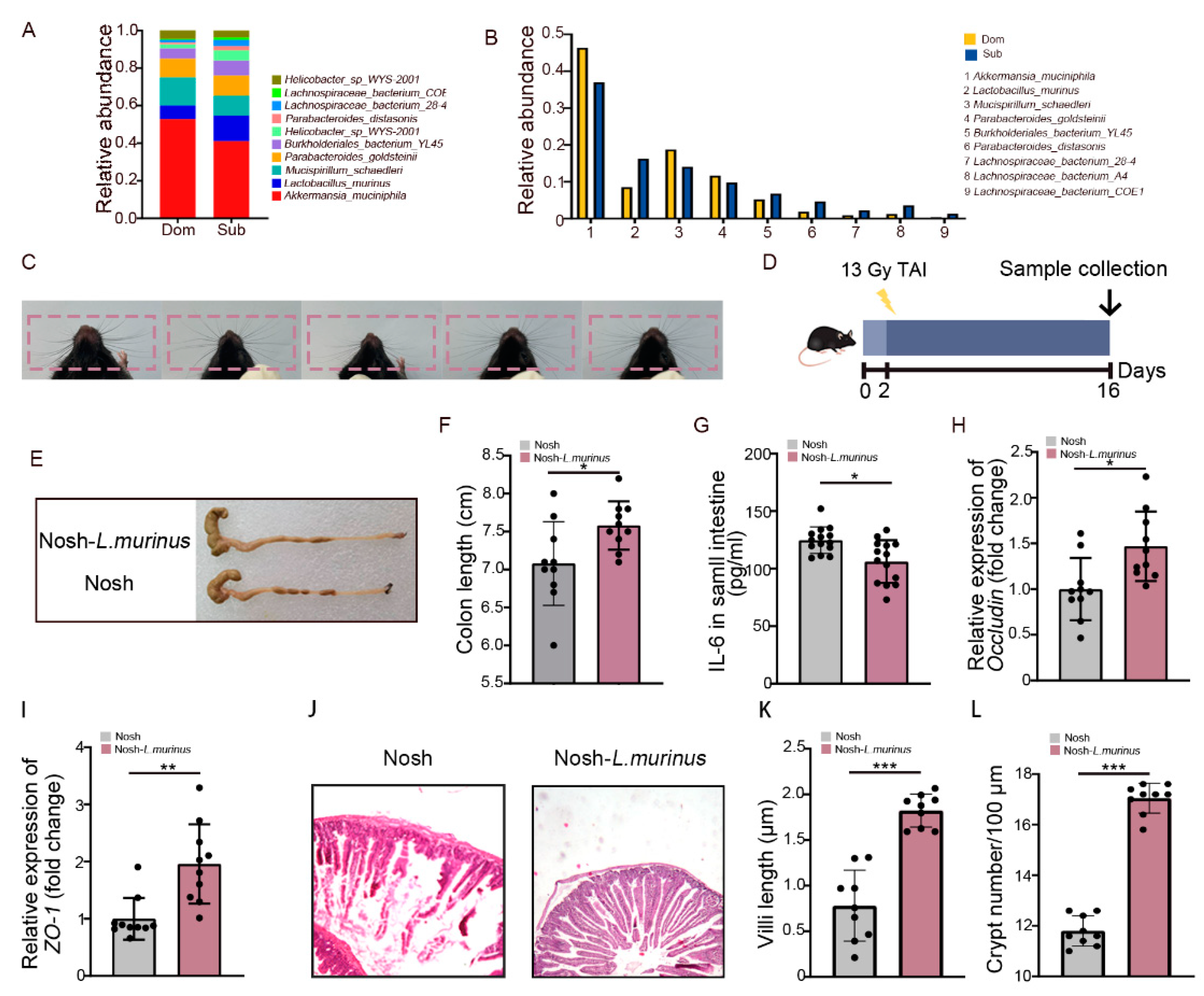
2.2. Irradiation Molds Characteristic Gut Microbiota Signature between Dom and Sub Mice
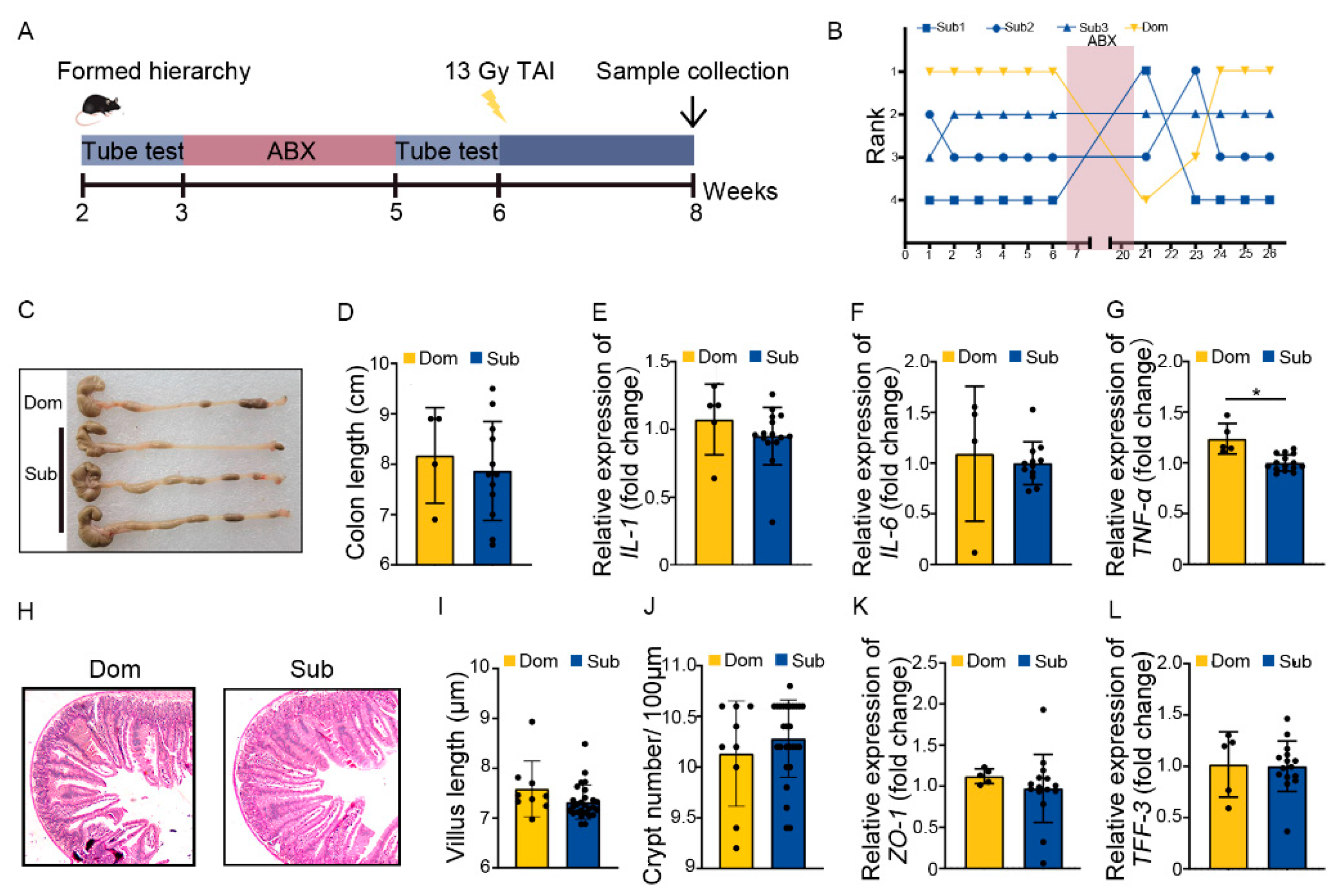
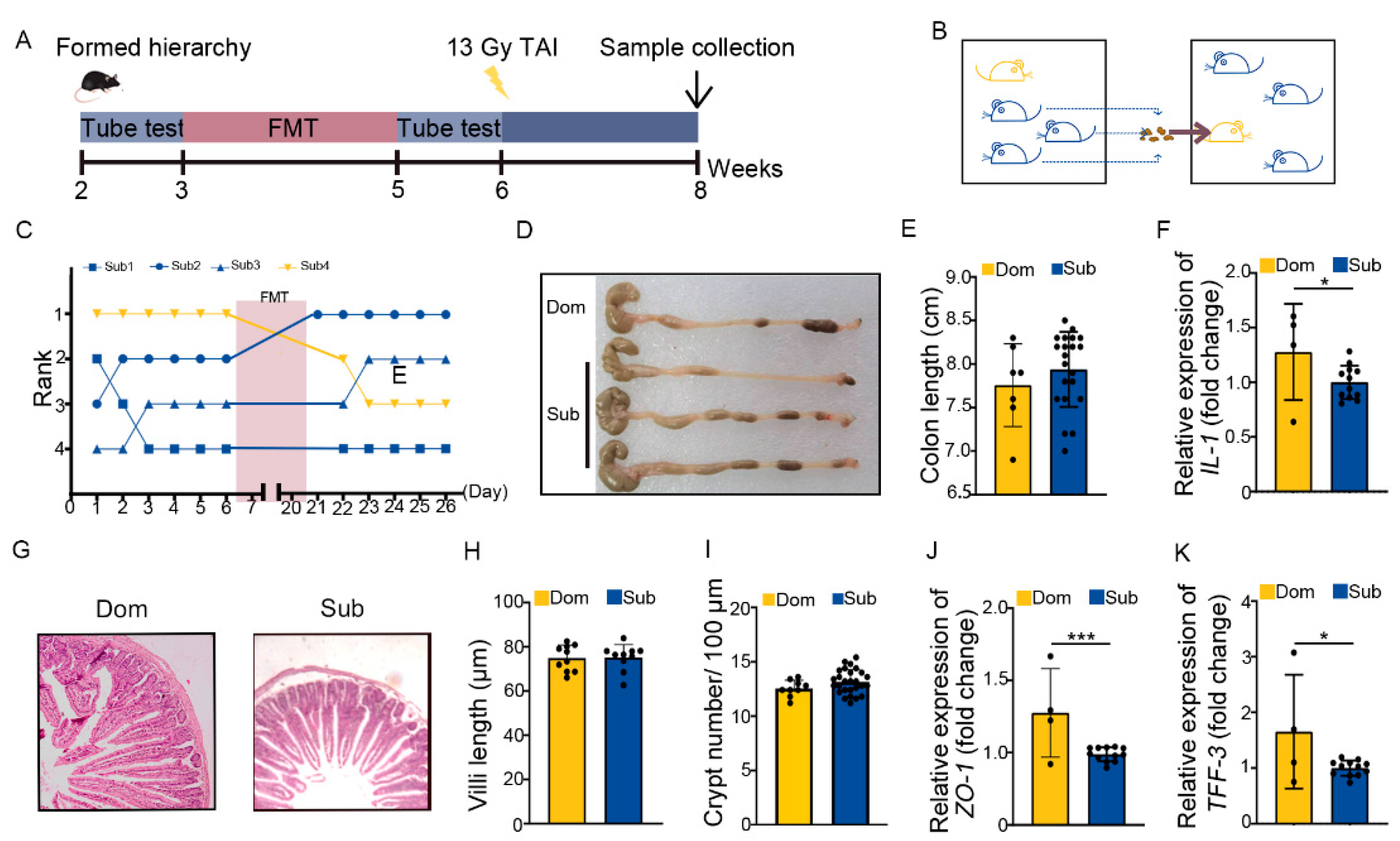
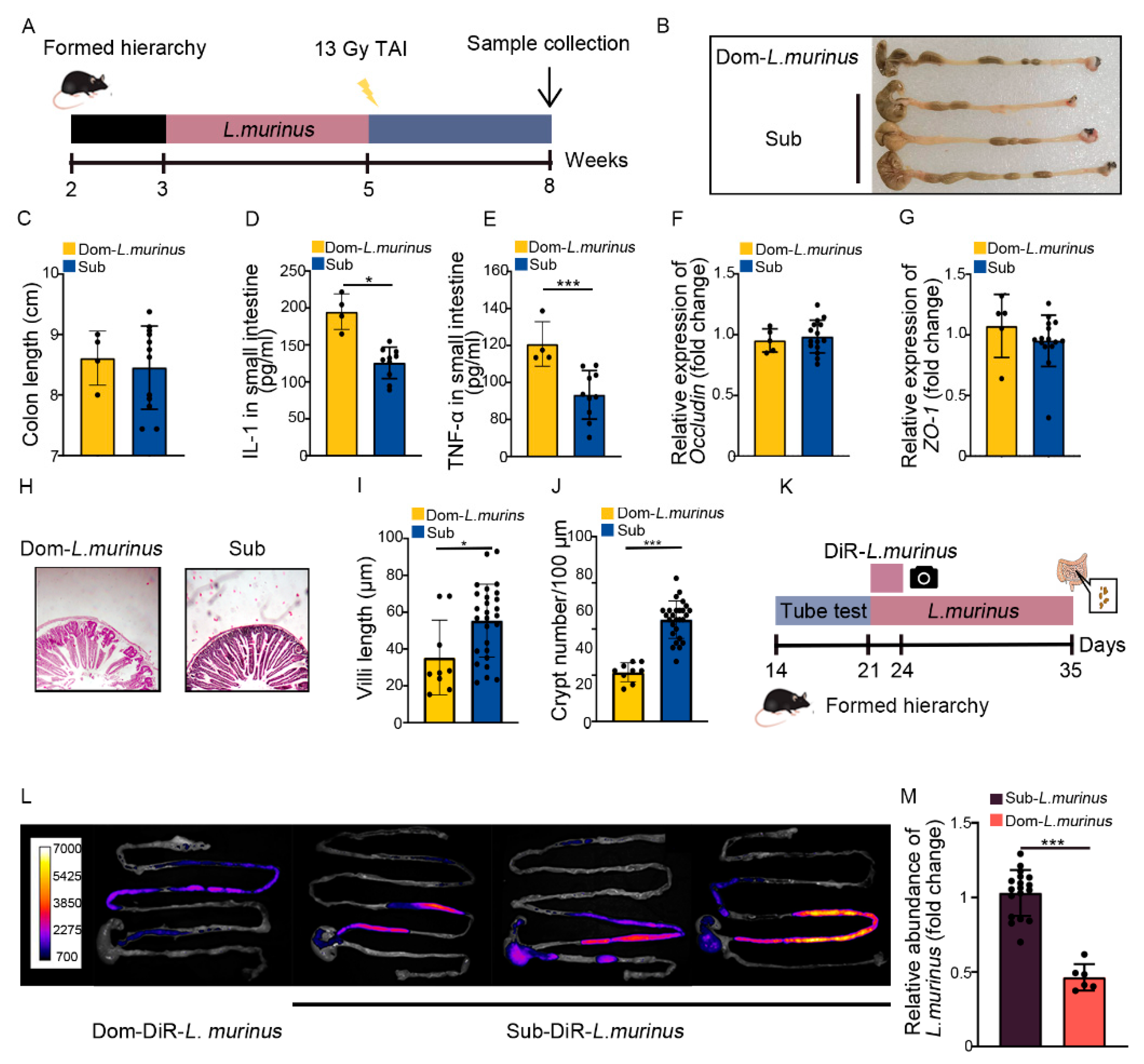
2.3. Social Hierarchy Impacts the Development of Intestinal Radiation Injuries in a Gut Microbiota-Dependent Manner
2.4. Lactobacillus Murinus Ameliorates Intestinal Radiation Injuries in Non-Social Hierarchy Mice
2.5. The Radioprotection of L. murinus Is Not Obvious for Dom Mice
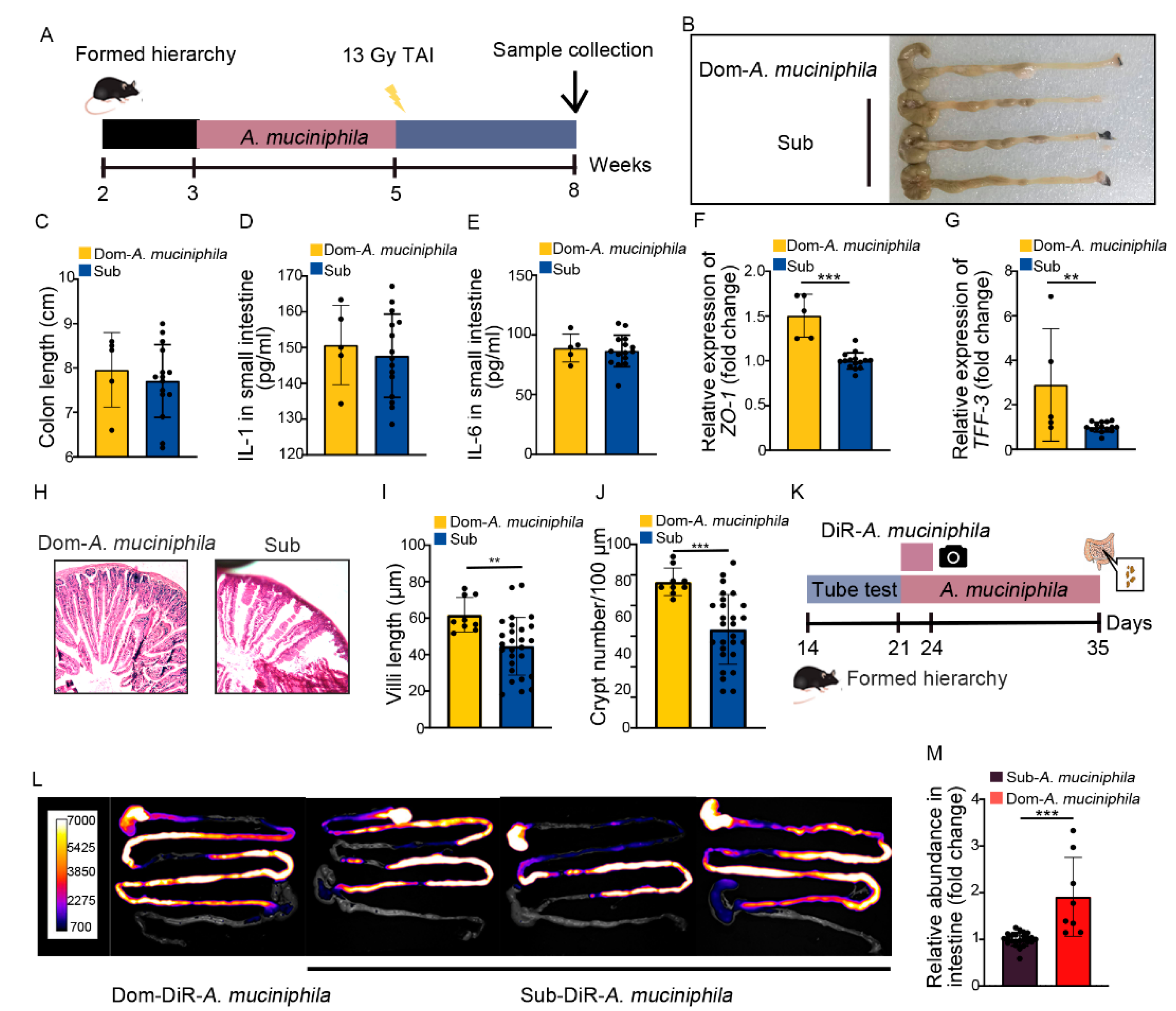
2.6. A. Muciniphila Colonizes in GI Tract Stably to Mitigate Intestinal Radiation Injury in Dom Mice
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Tube Test
4.3. Irradiation Study
4.4. Food Intake and Fecal Collection
4.5. Antibiotic Cocktail (ABX) Experiment
4.6. Fecal Microbiota Transplantation Experiment
4.7. Bacteria Strain Administration
4.8. Ex Vivo Fluorescence Imaging
4.9. Quantitative Analysis of Bacteria
4.10. RNA Isolation and Quantitative Reverse Transcription Real-Time PCR
4.11. Enzyme-Linked Immunosorbent Assay (ELISA)
4.12. Histology
4.13. Bacterial Diversity Analysis
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chou, Y.J.; Ma, Y.K.; Lu, Y.H.; King, J.T.; Tasi, W.S.; Yang, S.B.; Kuo, T.H. Potential cross-species correlations in social hierarchy and memory between mice and young children. Commun. Biol. 2022, 5, 230. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dai, G.; Gu, Z.; Liu, G.; Tang, K.; Pan, Y.H.; Chen, Y.; Lin, X.; Wu, N.; Chen, H.; et al. Accelerated evolution of an Lhx2 enhancer shapes mammalian social hierarchies. Cell Res. 2020, 30, 408–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Kessels, H.W.; Hu, H. The mouse that roared: Neural mechanisms of social hierarchy. Trends Neurosci. 2014, 37, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.R.; Toth, A.L.; Suarez, A.V.; Robinson, G.E. Genetic and genomic analyses of the division of labour in insect societies. Nat. Rev. Genet. 2008, 9, 735–748. [Google Scholar] [CrossRef]
- Evans, O.; Rodriguez-Borillo, O.; Font, L.; Currie, P.J.; Pastor, R. Alcohol Binge Drinking and Anxiety-Like Behavior in Socialized Versus Isolated C57BL/6J Mice. Alcohol. Clin. Exp. Res. 2020, 44, 244–254. [Google Scholar] [CrossRef]
- Fournier, M.A. Dimensions of human hierarchy as determinants of health and happiness. Curr. Opin. Psychol. 2020, 33, 110–114. [Google Scholar] [CrossRef]
- Gospocic, J.; Glastad, K.M.; Sheng, L.; Shields, E.J.; Berger, S.L.; Bonasio, R. Kr-h1 maintains distinct caste-specific neurotranscriptomes in response to socially regulated hormones. Cell 2021, 184, 5807–5823. [Google Scholar] [CrossRef]
- Chagas, L.A.; Batista, T.H.; Ribeiro, A.; Ferrari, M.S.; Vieira, J.S.; Rojas, V.; Kalil-Cutti, B.; Elias, L.; Giusti-Paiva, A.; Vilela, F.C. Anxiety-like behavior and neuroendocrine changes in offspring resulting from gestational post-traumatic stress disorder. Behav. Brain Res. 2021, 399, 113026. [Google Scholar] [CrossRef]
- Louch, C.D.; Higginbotham, M. The relation between social rank and plasma corticosterone levels in mice. Gen. Comp. Endocrinol. 1967, 8, 441–444. [Google Scholar] [CrossRef]
- Palle, A.; Zorzo, C.; Luskey, V.E.; McGreevy, K.R.; Fernandez, S.; Trejo, J.L. Social dominance differentially alters gene expression in the medial prefrontal cortex without affecting adult hippocampal neurogenesis or stress and anxiety-like behavior. FASEB J. 2019, 33, 6995–7008. [Google Scholar] [CrossRef]
- Stagkourakis, S.; Spigolon, G.; Williams, P.; Protzmann, J.; Fisone, G.; Broberger, C. A neural network for intermale aggression to establish social hierarchy. Nat. Neurosci. 2018, 21, 834–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, M.; Xiong, W.; Hu, F.; Deng, M.F.; Huang, X.; Chen, J.G.; Man, H.Y.; Lu, Y.; Liu, D.; Zhu, L.Q. A novel pathway regulates social hierarchy via lncRNA AtLAS and postsynaptic synapsin IIb. Cell Res. 2020, 30, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Huntingford, F.A.; Turner, A.K. The consequences of animal conflict. In Animal Conflict; Springer: Dordrecht, Germany, 1987; pp. 227–250. [Google Scholar]
- LeClair, K.B.; Chan, K.L.; Kaster, M.P.; Parise, L.F.; Burnett, C.J.; Russo, S.J. Individual history of winning and hierarchy landscape influence stress susceptibility in mice. eLife 2021, 10, e71401. [Google Scholar] [CrossRef] [PubMed]
- WARREN, S.; BOWERS, J.Z. The acute radiation syndrome in man. Ann. Intern. Med. 1950, 32, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Weisdorf, D.; Chao, N.; Waselenko, J.K.; Dainiak, N.; Armitage, J.O.; McNiece, I.; Confer, D. Acute radiation injury: Contingency planning for triage, supportive care, and transplantation. Biol. Blood Marrow Transplant. 2006, 12, 672–682. [Google Scholar] [CrossRef] [Green Version]
- Nag, D.; Bhanja, P.; Riha, R.; Sanchez-Guerrero, G.; Kimler, B.F.; Tsue, T.T.; Lominska, C.; Saha, S. Auranofin Protects Intestine against Radiation Injury by Modulating p53/p21 Pathway and Radiosensitizes Human Colon Tumor. Clin. Cancer Res. 2019, 25, 4791–4807. [Google Scholar] [CrossRef] [Green Version]
- Riehl, T.E.; Alvarado, D.; Ee, X.; Zuckerman, A.; Foster, L.; Kapoor, V.; Thotala, D.; Ciorba, M.A.; Stenson, W.F. Lactobacillus rhamnosus GG protects the intestinal epithelium from radiation injury through release of lipoteichoic acid, macrophage activation and the migration of mesenchymal stem cells. Gut 2019, 68, 1003–1013. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Z.; Jiang, Z.; Luo, P.; Liu, L.; Huang, Y.; Wang, H.; Wang, Y.; Long, L.; Tan, X.; et al. Cordycepin prevents radiation ulcer by inhibiting cell senescence via NRF2 and AMPK in rodents. Nat. Commun. 2019, 10, 2538. [Google Scholar] [CrossRef] [Green Version]
- Carmody, R.N.; Sarkar, A.; Reese, A.T. Gut microbiota through an evolutionary lens. Science 2021, 372, 462–463. [Google Scholar] [CrossRef]
- Weersma, R.K.; Zhernakova, A.; Fu, J. Interaction between drugs and the gut microbiome. Gut 2020, 69, 1510–1519. [Google Scholar] [CrossRef]
- Matenchuk, B.A.; Mandhane, P.J.; Kozyrskyj, A.L. Sleep, circadian rhythm, and gut microbiota. Sleep Med. Rev. 2020, 53, 101340. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Chang, E.B. Inflammatory Bowel Diseases (IBD) and the Microbiome-Searching the Crime Scene for Clues. Gastroenterology 2021, 160, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Tanes, C.; Bittinger, K.; Gao, Y.; Friedman, E.S.; Nessel, L.; Paladhi, U.R.; Chau, L.; Panfen, E.; Fischbach, M.A.; Braun, J.; et al. Role of dietary fiber in the recovery of the human gut microbiome and its metabolome. Cell Host Microbe 2021, 29, 394–407. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xu, J.; Xu, Y.; Xiao, J.; Bi, N.; Gu, X.; Wang, H.L. Gut microbiota shapes social dominance through modulating HDAC2 in the medial prefrontal cortex. Cell Rep. 2022, 38, 110478. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yan, C.; Liu, W.; Chen, K.; Xing, L.; Li, H.; Zhao, X. Research Note: Integrated gut microbiome and short-chain fatty acids responds to dominance hierarchy in roosters. Poult. Sci. 2022, 101, 101670. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhao, P.; Dou, M.; Mao, J.; Zhang, G.; Su, Y.; Wang, Q.; Wang, Q.; Wang, Y.; Sun, R.; et al. Exogenous microbiota-derived metabolite trimethylamine N-oxide treatment alters social behaviors: Involvement of hippocampal metabolic adaptation. Neuropharmacology 2021, 191, 108563. [Google Scholar] [CrossRef]
- Xiao, H.W.; Cui, M.; Li, Y.; Dong, J.L.; Zhang, S.Q.; Zhu, C.C.; Jiang, M.; Zhu, T.; Wang, B.; Wang, H.C.; et al. Gut microbiota-derived indole 3-propionic acid protects against radiation toxicity via retaining acyl-CoA-binding protein. Microbiome 2020, 8, 69. [Google Scholar] [CrossRef]
- Li, Y.; Dong, J.; Xiao, H.; Zhang, S.; Wang, B.; Cui, M.; Fan, S. Gut commensal derived-valeric acid protects against radiation injuries. Gut Microbes 2020, 11, 789–806. [Google Scholar] [CrossRef]
- Xiao, H.W.; Li, Y.; Luo, D.; Dong, J.L.; Zhou, L.X.; Zhao, S.Y.; Zheng, Q.S.; Wang, H.C.; Cui, M.; Fan, S.J. Hydrogen-water ameliorates radiation-induced gastrointestinal toxicity via MyD88’s effects on the gut microbiota. Exp. Mol. Med. 2018, 50, e433. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.; Xiao, H.; Li, Y.; Zhang, S.; Dong, J.; Wang, B.; Zhu, C.; Jiang, M.; Zhu, T.; He, J.; et al. Sexual Dimorphism of Gut Microbiota Dictates Therapeutics Efficacy of Radiation Injuries. Adv. Sci. 2019, 6, 1901048. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, S.Q.; Dong, J.L.; Li, Y.; Jin, Y.X.; Xiao, H.W.; Wang, H.C.; Fan, S.J.; Cui, M. Ambient temperature structures the gut microbiota of zebrafish to impact the response to radioactive pollution. Environ. Pollut. 2022, 293, 118539. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Zhu, H.; Zhou, T.; Wang, S.; Wu, Y.; Hu, H. Using the tube test to measure social hierarchy in mice. Nat. Protoc. 2019, 14, 819–831. [Google Scholar] [CrossRef]
- Hu, J.; Deng, F.; Zhao, B.; Lin, Z.; Sun, Q.; Yang, X.; Wu, M.; Qiu, S.; Chen, Y.; Yan, Z.; et al. Lactobacillus murinus alleviate intestinal ischemia/reperfusion injury through promoting the release of interleukin-10 from M2 macrophages via Toll-like receptor 2 signaling. Microbiome 2022, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Wilck, N.; Matus, M.G.; Kearney, S.M.; Olesen, S.W.; Forslund, K.; Bartolomaeus, H.; Haase, S.; Mahler, A.; Balogh, A.; Marko, L.; et al. Salt-responsive gut commensal modulates TH17 axis and disease. Nature 2017, 551, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Jin, Y.X.; Dong, J.L.; Xiao, H.W.; Zhang, S.Q.; Li, Y.; Chen, Z.Y.; Yang, X.D.; Fan, S.J.; Cui, M. Low-Intensity Exercise Modulates Gut Microbiota to Fight Against Radiation-Induced Gut Toxicity in Mouse Models. Front. Cell Dev. Biol. 2021, 9, 706755. [Google Scholar] [CrossRef]
- Duhachek-Muggy, S.; Bhat, K.; Medina, P.; Cheng, F.; He, L.; Alli, C.; Saki, M.; Muthukrishnan, S.D.; Ruffenach, G.; Eghbali, M.; et al. Radiation mitigation of the intestinal acute radiation injury in mice by 1-[(4-nitrophenyl)sulfonyl]-4-phenylpiperazine. Stem Cells Transl. Med. 2020, 9, 106–119. [Google Scholar] [CrossRef] [Green Version]
- Hobson, E.A.; Monster, D.; DeDeo, S. Aggression heuristics underlie animal dominance hierarchies and provide evidence of group-level social information. Proc. Natl. Acad. Sci. USA 2021, 118, e2022912118. [Google Scholar] [CrossRef]
- van den Berg, W.E.; Lamballais, S.; Kushner, S.A. Sex-specific mechanism of social hierarchy in mice. Neuropsychopharmacology 2015, 40, 1364–1372. [Google Scholar] [CrossRef] [Green Version]
- Russell, G.; Lightman, S. The human stress response. Nat. Rev. Endocrinol. 2019, 15, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Gerassy-Vainberg, S.; Blatt, A.; Danin-Poleg, Y.; Gershovich, K.; Sabo, E.; Nevelsky, A.; Daniel, S.; Dahan, A.; Ziv, O.; Dheer, R.; et al. Radiation induces proinflammatory dysbiosis: Transmission of inflammatory susceptibility by host cytokine induction. Gut 2018, 67, 97–107. [Google Scholar] [CrossRef]
- Caruso, R.; Ono, M.; Bunker, M.E.; Nunez, G.; Inohara, N. Dynamic and Asymmetric Changes of the Microbial Communities after Cohousing in Laboratory Mice. Cell Rep. 2019, 27, 3401–3412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virta, L.; Auvinen, A.; Helenius, H.; Huovinen, P.; Kolho, K.L. Association of repeated exposure to antibiotics with the development of pediatric Crohn’s disease--a nationwide, register-based finnish case-control study. Am. J. Epidemiol. 2012, 175, 775–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, T.; Wang, J.; Chen, L.; Shan, J.; Di, L. Lactobacillus murinus Improved the Bioavailability of Orally Administered Glycyrrhizic Acid in Rats. Front. Microbiol. 2020, 11, 597. [Google Scholar] [CrossRef]
- Qu, S.; Fan, L.; Qi, Y.; Xu, C.; Hu, Y.; Chen, S.; Liu, W.; Liu, W.; Si, J. Akkermansia muciniphila Alleviates Dextran Sulfate Sodium (DSS)-Induced Acute Colitis by NLRP3 Activation. Microbiol Spectr 2021, 9, e73021. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tang, L.; Feng, Y.; Zhao, S.; Han, M.; Zhang, C.; Yuan, G.; Zhu, J.; Cao, S.; Wu, Q.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurised bacterium blunts colitis associated tumourigenesis by modulation of CD8(+) T cells in mice. Gut 2020, 69, 1988–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, Y.J.; Lu, Y.H.; Ma, Y.K.; Su, Y.S.; Kuo, T.H. The decisive role of subordination in social hierarchy in weanling mice and young children. iScience 2021, 24, 102073. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, X.; Liu, Z.; Dong, Y.; Zhao, J.; Wang, B.; Xiao, H.; Li, Y.; Chen, Z.; Liu, X.; Liu, J.; et al. Social Hierarchy Dictates Intestinal Radiation Injury in a Gut Microbiota-Dependent Manner. Int. J. Mol. Sci. 2022, 23, 13189. https://doi.org/10.3390/ijms232113189
Zeng X, Liu Z, Dong Y, Zhao J, Wang B, Xiao H, Li Y, Chen Z, Liu X, Liu J, et al. Social Hierarchy Dictates Intestinal Radiation Injury in a Gut Microbiota-Dependent Manner. International Journal of Molecular Sciences. 2022; 23(21):13189. https://doi.org/10.3390/ijms232113189
Chicago/Turabian StyleZeng, Xiaozhou, Zhihong Liu, Yanxi Dong, Jiamin Zhao, Bin Wang, Huiwen Xiao, Yuan Li, Zhiyuan Chen, Xiaojing Liu, Jia Liu, and et al. 2022. "Social Hierarchy Dictates Intestinal Radiation Injury in a Gut Microbiota-Dependent Manner" International Journal of Molecular Sciences 23, no. 21: 13189. https://doi.org/10.3390/ijms232113189
APA StyleZeng, X., Liu, Z., Dong, Y., Zhao, J., Wang, B., Xiao, H., Li, Y., Chen, Z., Liu, X., Liu, J., Dong, J., Fan, S., & Cui, M. (2022). Social Hierarchy Dictates Intestinal Radiation Injury in a Gut Microbiota-Dependent Manner. International Journal of Molecular Sciences, 23(21), 13189. https://doi.org/10.3390/ijms232113189