
The BRCT Domain from the Homologue of the Oncogene PES1 in Leishmania major (LmjPES) Promotes Malignancy and Drug Resistance in Mammalian Cells

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
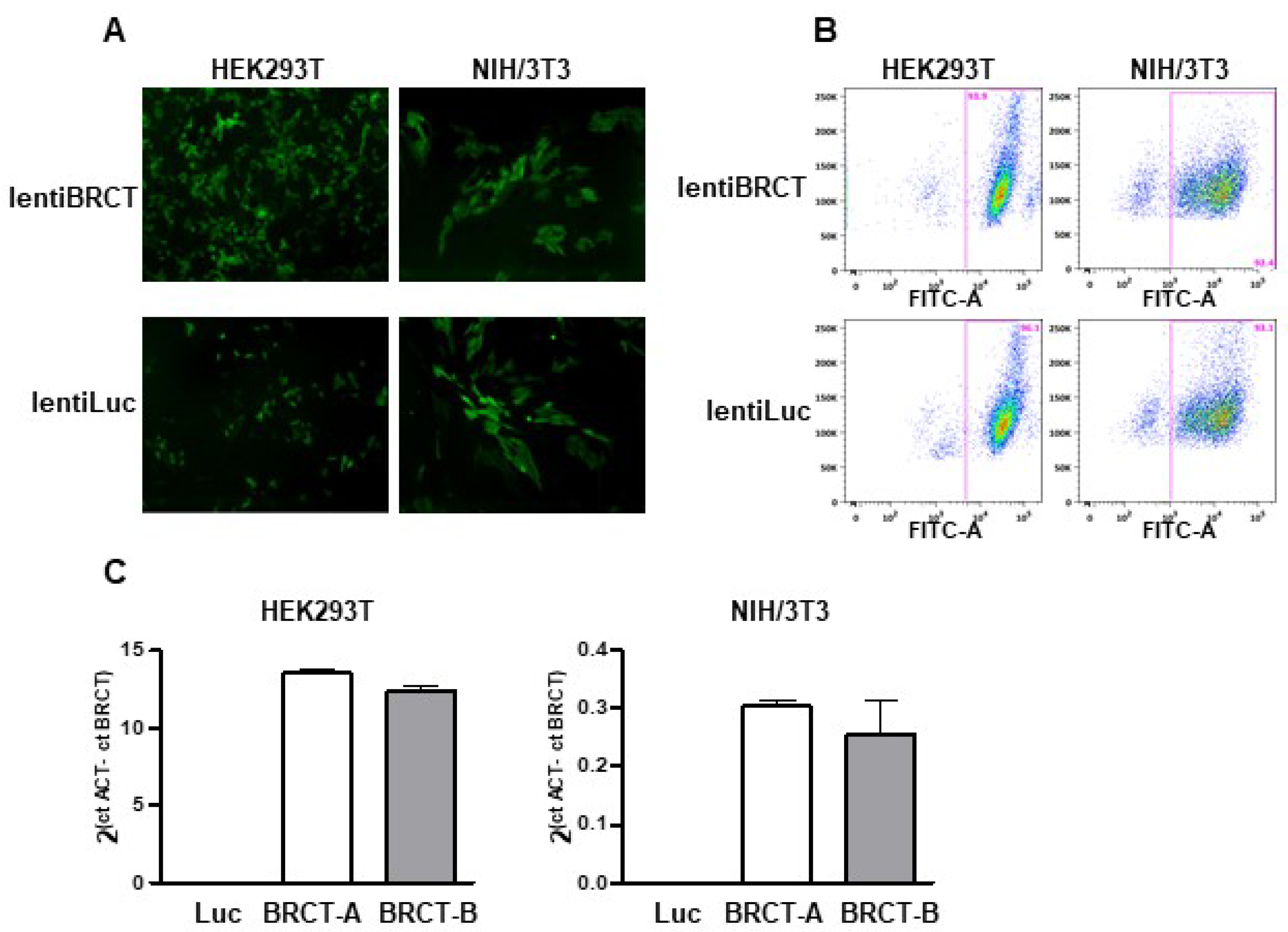
2.1. Generation of a Lentivirus Harboring the BRCT Domain Sequence from LmjPES Gene
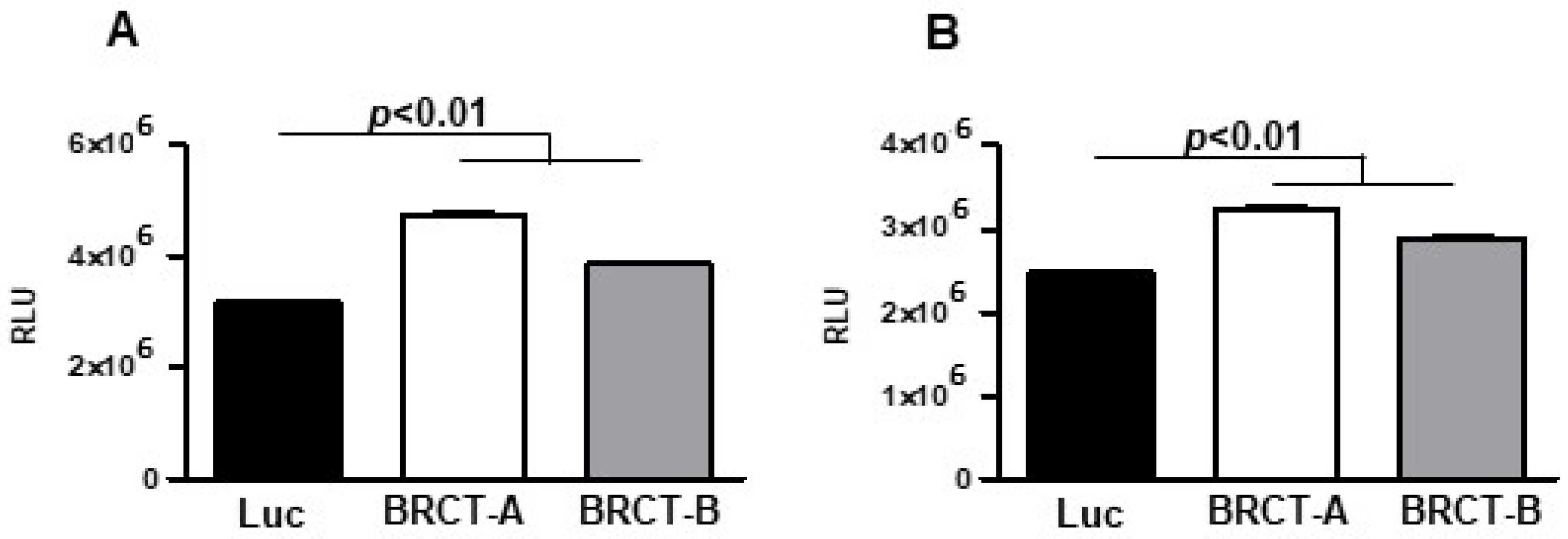
2.2. The Expression of BRCT Domain from LmjPES Dramatically Increased the Replication of Mammal Cells
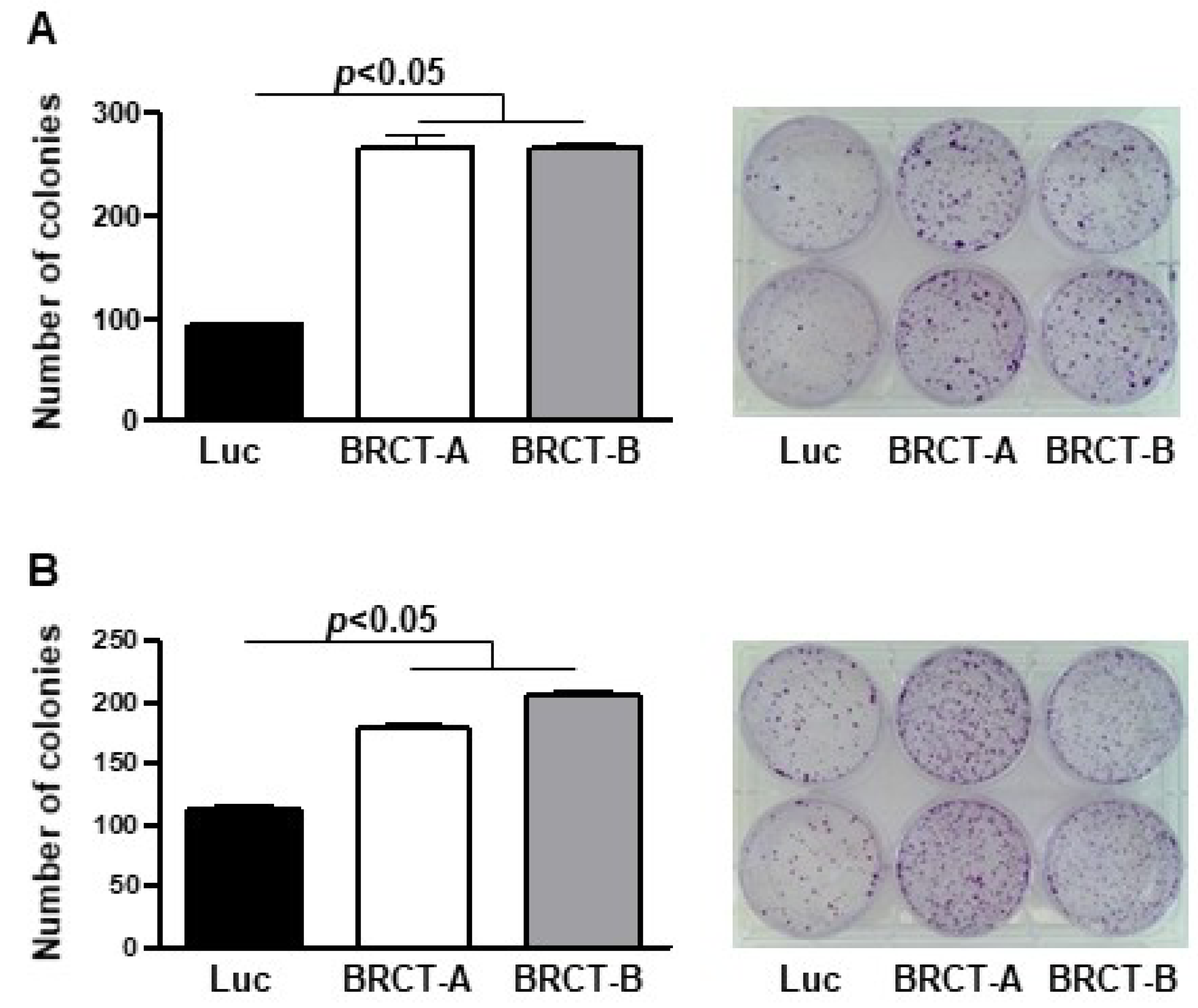
2.3. The Expression of BRCT Domain from LmjPES Protein Promoted Colony Formation in Mammalian Cells
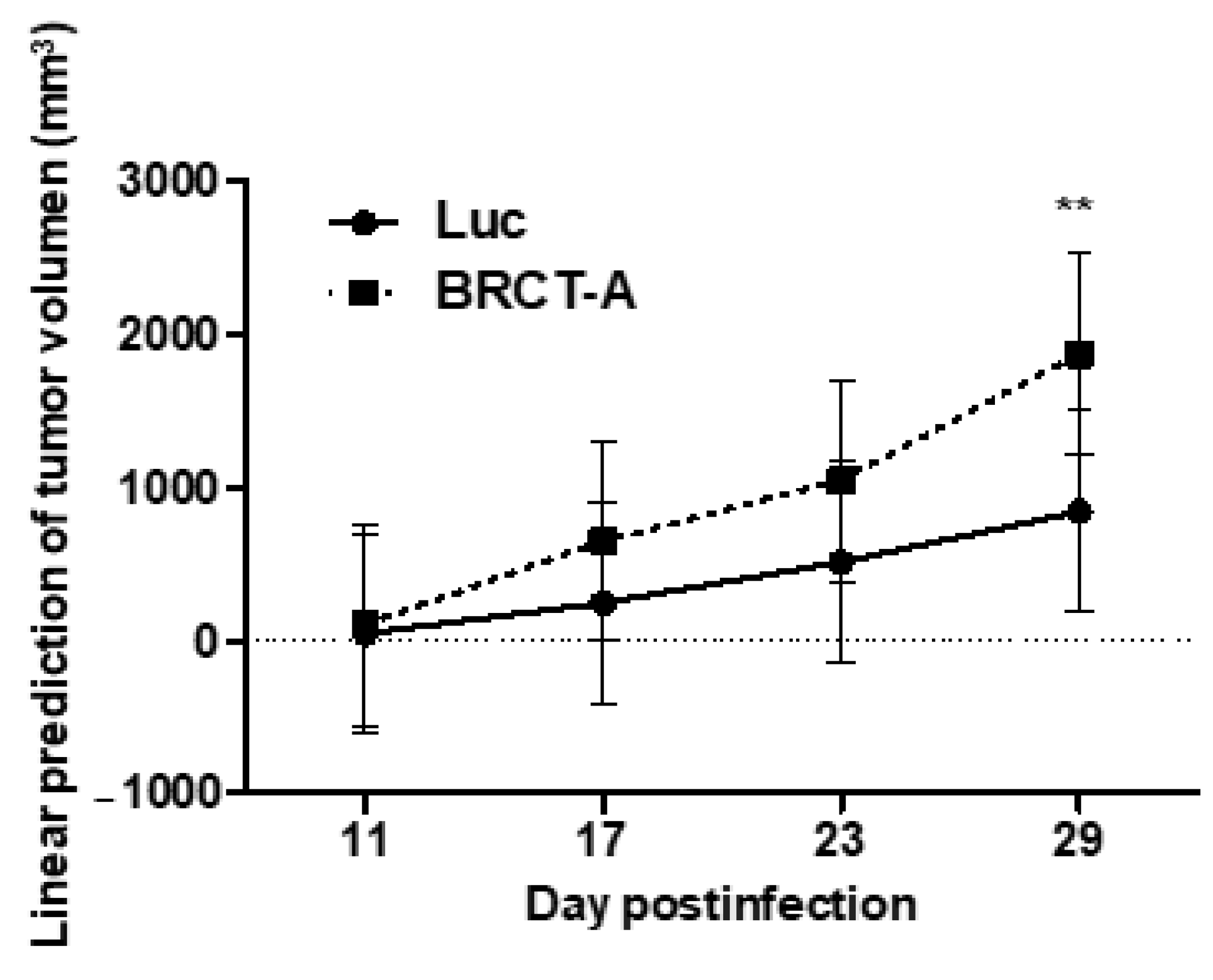
2.4. The Studied BRCT Domain Reinforced Tumorigenesis Process In Vivo
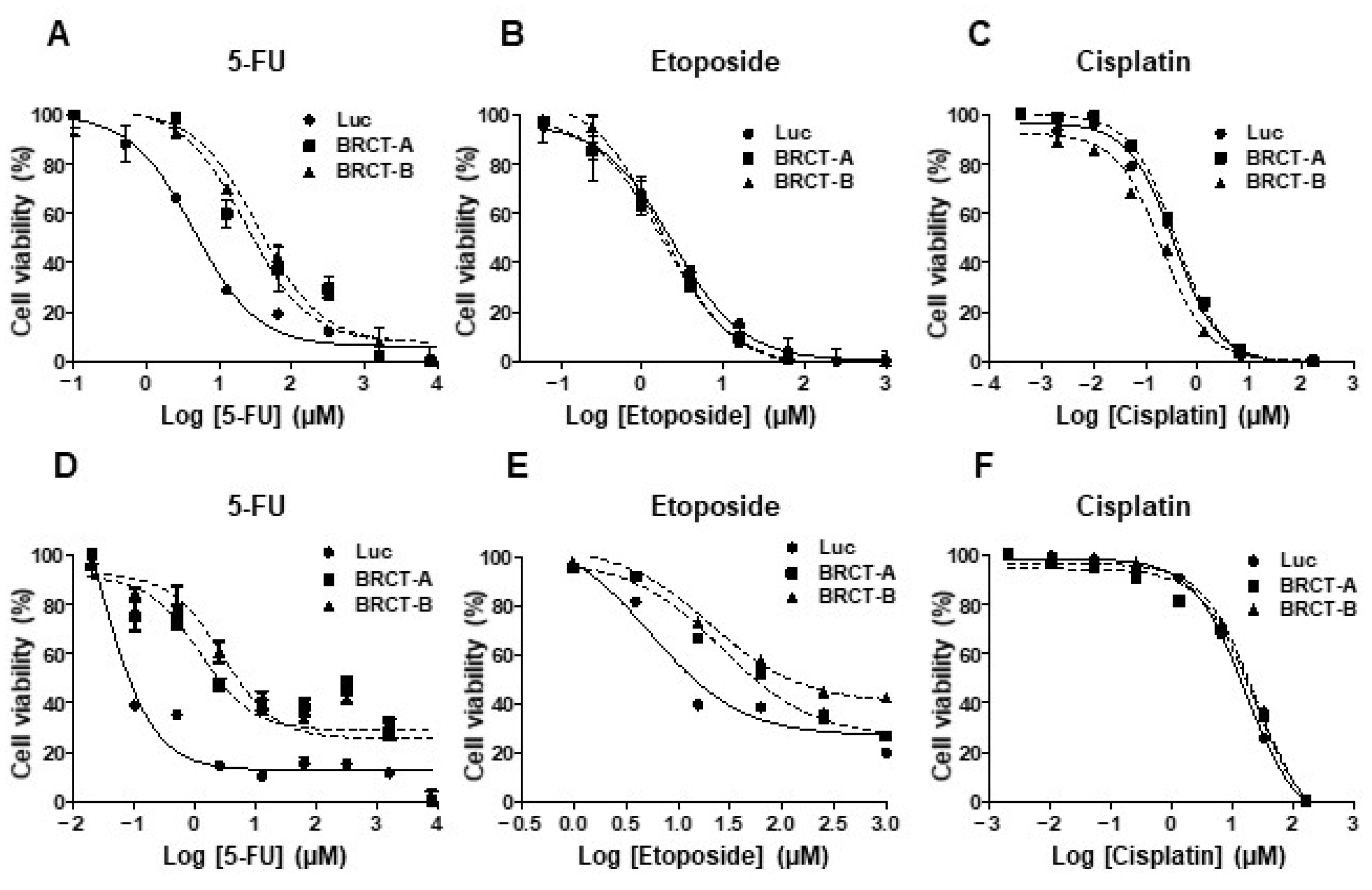
2.5. These Novel Tumorigenic Cells Were More Resistant to Death Induced by Antitumor Drugs
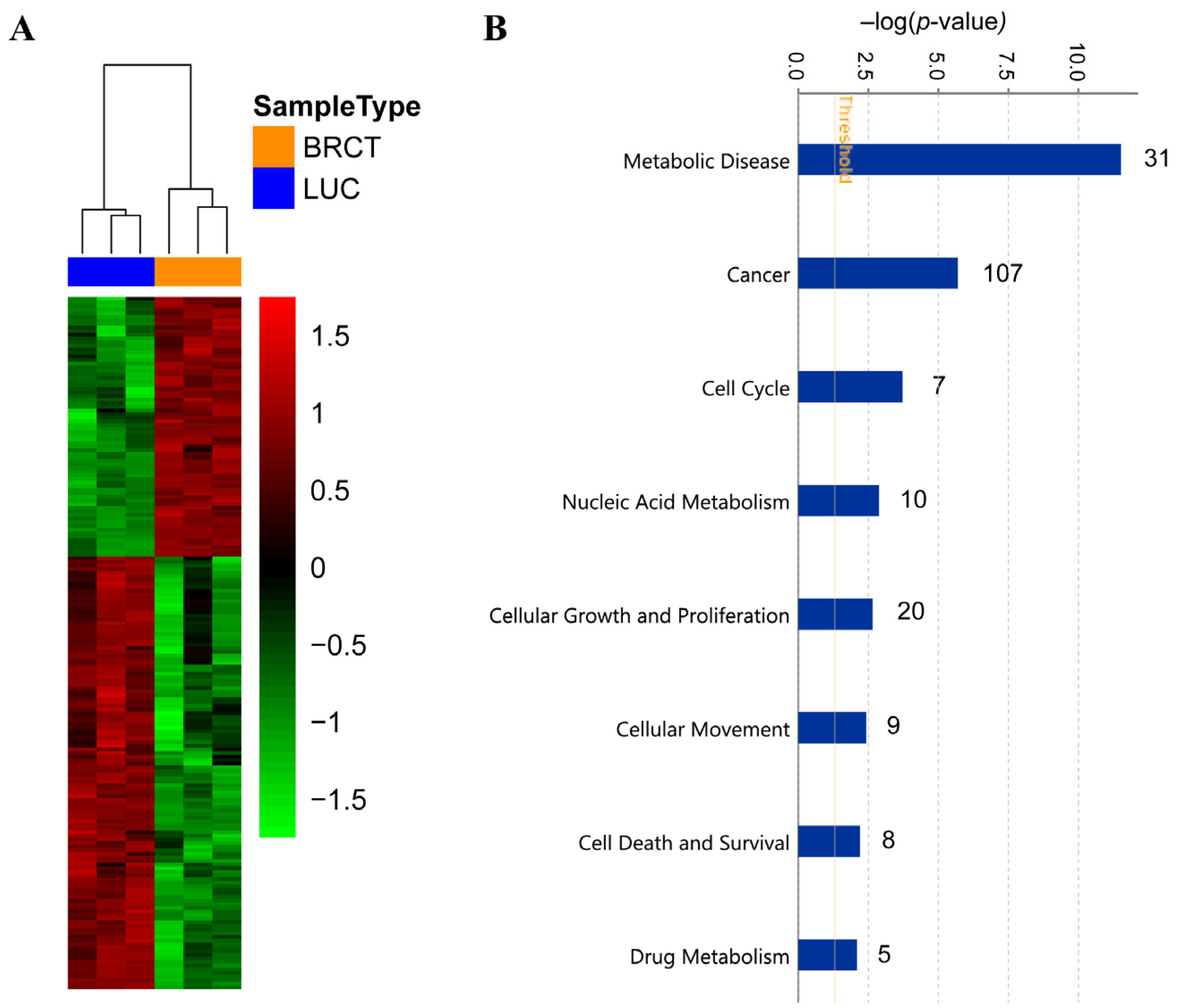
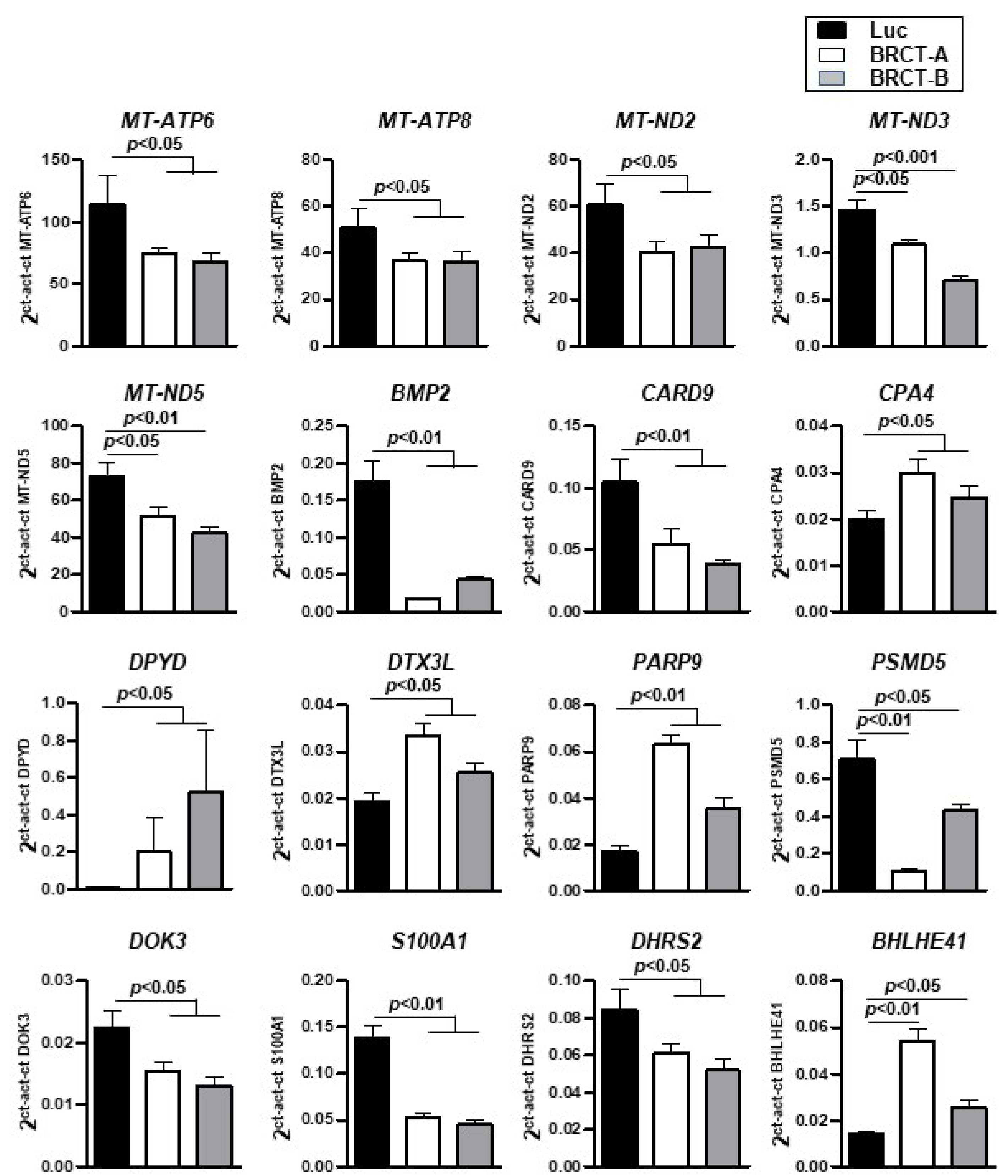
2.6. Genes Deregulated through BRCT Domain Expression Mainly Overlapped with Metabolic Disease, Cellular Proliferation, Survival and Drug Metabolism Profile
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Lentiviral Vector Assembly
4.3. Lentiviral Transduction of Mammal Cells
4.4. Cell Proliferation Assay
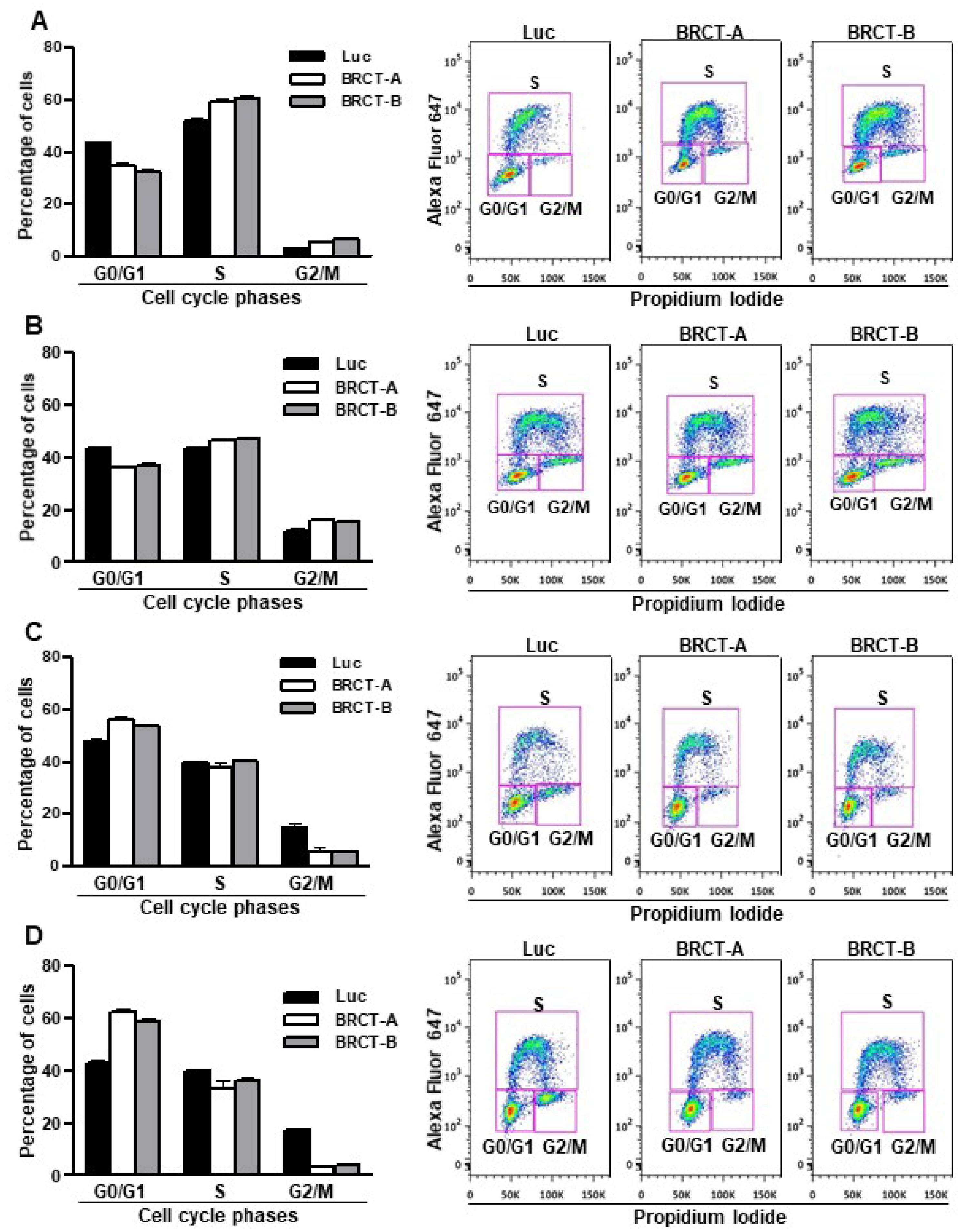
4.5. Cell Cycle Analysis
4.6. Colony Formation Assay
4.7. Drugs Resistance Study
4.8. In Vivo Tumorigenicity Assay
4.9. RNA Preparation and RNA-Sequencing (RNA-seq) Analysis
4.10. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Martel, C.; Georges, D.; Bray, F.; Ferlay, J.; Clifford, G.M. Global Burden of Cancer Attributable to Infections in 2018: A Worldwide Incidence Analysis. Lancet Glob. Health 2020, 8, e180–e190. [Google Scholar] [CrossRef] [Green Version]
- Plummer, M.; de Martel, C.; Vignat, J.; Ferlay, J.; Bray, F.; Franceschi, S. Global Burden of Cancers Attributable to Infections in 2012: A Synthetic Analysis. Lancet Glob. Health 2016, 4, e609–e616. [Google Scholar] [CrossRef] [Green Version]
- Nguewa, P.A.; Villa, T.G.; Notario, V. Microbiome Control in the Prevention and Early Management of Cancer. In New Weapons to Control Bacterial Growth; Springer International Publishing: Cham, Switzerland, 2016; pp. 219–237. ISBN 9783319283685. [Google Scholar]
- Alibek, K.; Kakpenova, A.; Baiken, Y. Role of Infectious Agents in the Carcinogenesis of Brain and Head and Neck Cancers. Infect. Agent. Cancer 2013, 8, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermida, M.D.R.; Doria, P.G.; Taguchi, A.M.P.; Mengel, J.O.; Dos-Santos, W.L.C. Leishmania Amazonensis Infection Impairs Dendritic Cell Migration from the Inflammatory Site to the Draining Lymph Node. BMC Infect. Dis. 2014, 14, 450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashidi, S.; Fernández-Rubio, C.; Manzano-Román, R.; Mansouri, R.; Shafiei, R.; Ali-Hassanzadeh, M.; Barazesh, A.; Karimazar, M.; Hatam, G.; Nguewa, P. Potential Therapeutic Targets Shared between Leishmaniasis and Cancer. Parasitology 2021, 148, 655–671. [Google Scholar] [CrossRef]
- Torres-Guerrero, E.; Quintanilla-Cedillo, M.R.; Ruiz-Esmenjaud, J.; Arenas, R. Leishmaniasis: A Review. F1000Research 2017, 6, 750. [Google Scholar] [CrossRef]
- Pace, D. Leishmaniasis. J. Infect. 2014, 69 (Suppl. S1), S10–S18. [Google Scholar] [CrossRef]
- Kopterides, P.; Mourtzoukou, E.G.; Skopelitis, E.; Tsavaris, N.; Falagas, M.E. Aspects of the Association between Leishmaniasis and Malignant Disorders. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 1181–1189. [Google Scholar] [CrossRef]
- Kopterides, P.; Halikias, S.; Tsavaris, N. Visceral Leishmaniasis Masquerading as Myelodysplasia. Am. J. Hematol. 2003, 74, 198–199. [Google Scholar] [CrossRef]
- Böer, A.; Blödorn-Schlicht, N.; Wiebels, D.; Steinkraus, V.; Falk, T.M. Unusual Histopathological Features of Cutaneous Leishmaniasis Identified by Polymerase Chain Reaction Specific for Leishmania on Paraffin-Embedded Skin Biopsies. Br. J. Dermatol. 2006, 155, 815–819. [Google Scholar] [CrossRef]
- Oetken, T.; Hiscox, B.; Orengo, I.; Rosen, T. Cutaneous Leishmaniasis Mimicking Squamous Cell Carcinoma. Dermatol. Online J. 2017, 23, 13030. [Google Scholar] [CrossRef]
- Nicodemo, A.C.; Duailibi, D.F.; Feriani, D.; Duarte, M.I.S.; Amato, V.S. Mucosal Leishmaniasis Mimicking T-Cell Lymphoma in a Patient Receiving Monoclonal Antibody against TNFα. PLoS Negl. Trop. Dis. 2017, 11, e0005807. [Google Scholar] [CrossRef] [Green Version]
- Czechowicz, R.T.; Millard, T.P.; Smith, H.R.; Ashton, R.E.; Lucas, S.B.; Hay, R.J. Reactivation of Cutaneous Leishmaniasis after Surgery. Br. J. Dermatol. 1999, 141, 1113–1116. [Google Scholar] [CrossRef]
- Dereure, J. Visceral Leishmaniasis. Persistence of Parasites in Lymph Nodes after Clinical Cure. J. Infect. 2003, 47, 77–81. [Google Scholar] [CrossRef]
- Boutros, N.; Hawkins, D.; Nelson, M.; Lampert, I.A.; Naresh, K.N. Burkitt Lymphoma and Leishmaniasis in the Same Tissue Sample in an AIDS Patient. Histopathology 2006, 48, 880–881. [Google Scholar] [CrossRef] [PubMed]
- Blum-Domínguez, S.D.C.; Martínez-Vázquez, A.; Núñez-Oreza, L.A.; Martínez-Hernández, F.; Villalobos, G.; Tamay-Segovia, P. Diffuse Cutaneous Leishmaniasis (DCL) and Visceral Leishmaniasis (VL) Concurrent with Cancer: Presentation of a Case. Gac. Med. Mex. 2017, 153, 121–124. [Google Scholar] [PubMed]
- Liao, H.; Jin, Y.; Yu, J.; Jiang, N. Concomitant T-Cell Prolymphocytic Leukemia and Visceral Leishmaniasis. Medicine 2018, 97, e12410. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.; Hanson, S.; Goldberg, L.H. Squamous Cell Carcinoma Arising in a Leishmania Scar. Dermatol. Surg. 2003, 29, 1148–1149. [Google Scholar] [CrossRef]
- Gurel, M.S.; Inal, L.; Ozardali, I.; Duzgun, S.A. Basal Cell Carcinoma in a Leishmanial Scar. Clin. Exp. Dermatol. 2005, 30, 444–445. [Google Scholar] [CrossRef]
- Osakwe, N.M.; Paulus, A.; Haggerty, P.F.; Wood, R.A.; Becker, S.J.; Weina, P.J.; Dolan, M.J.; Prakash, V. Visceral Leishmaniasis with Associated Immune Dysregulation Leading to Lymphoma. Mil. Med. 2013, 178, e386–e389. [Google Scholar] [CrossRef]
- Pintor-Tortolero, J.; Medina, J.; Durán, M.; Briceño, J. Anorectal Tumor as a Debut Form of Visceral Leishmaniasis. Surgery 2018, 164, 1390–1391. [Google Scholar] [CrossRef]
- Algarabel, M.; Fernández-Rubio, C.; Musilova, K.; Peña-Guerrero, J.; Vacas, A.; Larrea, E.; Nguewa, P.A. In Leishmania Major, the Homolog of the Oncogene PES1 May Play a Critical Role in Parasite Infectivity. Int. J. Mol. Sci. 2021, 22, 12592. [Google Scholar] [CrossRef]
- Grimm, T.; Hölzel, M.; Rohrmoser, M.; Harasim, T.; Malamoussi, A.; Gruber-Eber, A.; Kremmer, E.; Eick, D. Dominant-Negative Pes1 Mutants Inhibit Ribosomal RNA Processing and Cell Proliferation via Incorporation into the PeBoW-Complex. Nucleic Acids Res. 2006, 34, 3030–3043. [Google Scholar] [CrossRef] [Green Version]
- Tecza, A.; Bugner, V.; Kühl, M.; Kühl, S.J. Pescadillo Homologue 1 and Peter Pan Function during Xenopus Laevis Pronephros Development. Biol. cell 2011, 103, 483–498. [Google Scholar] [CrossRef]
- Fan, P.; Wang, B.; Meng, Z.; Zhao, J.; Jin, X. PES1 Is Transcriptionally Regulated by BRD4 and Promotes Cell Proliferation and Glycolysis in Hepatocellular Carcinoma. Int. J. Biochem. Cell Biol. 2018, 104, 1–8. [Google Scholar] [CrossRef]
- Spitzner, M.; Emons, G.; Kramer, F.; Gaedcke, J.; Rave-Fränk, M.; Scharf, J.-G.; Burfeind, P.; Becker, H.; Beissbarth, T.; Ghadimi, B.M.; et al. A Gene Expression Signature for Chemoradiosensitivity of Colorectal Cancer Cells. Int. J. Radiat. Oncol. Biol. Phys. 2010, 78, 1184–1192. [Google Scholar] [CrossRef]
- Maiorana, A.; Tu, X.; Cheng, G.; Baserga, R. Role of Pescadillo in the Transformation and Immortalization of Mammalian Cells. Oncogene 2004, 23, 7116–7124. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, J.; Thomas, G.; Volarević, S. Ribosome Biogenesis in Cancer: New Players and Therapeutic Avenues. Nat. Rev. Cancer 2018, 18, 51–63. [Google Scholar] [CrossRef]
- Li, J.; Yu, L.; Zhang, H.; Wu, J.; Yuan, J.; Li, X.; Li, M. Down-Regulation of Pescadillo Inhibits Proliferation and Tumorigenicity of Breast Cancer Cells. Cancer Sci. 2009, 100, 2255–2260. [Google Scholar] [CrossRef]
- Kinoshita, Y.; Jarell, A.D.; Flaman, J.M.; Foltz, G.; Schuster, J.; Sopher, B.L.; Irvin, D.K.; Kanning, K.; Kornblum, H.I.; Nelson, P.S.; et al. Pescadillo, a Novel Cell Cycle Regulatory Protein Abnormally Expressed in Malignant Cells. J. Biol. Chem. 2001, 276, 6656–6665. [Google Scholar] [CrossRef]
- Xie, W.; Qu, L.; Meng, L.; Liu, C.; Wu, J.; Shou, C. PES1 Regulates Sensitivity of Colorectal Cancer Cells to Anticancer Drugs. Biochem. Biophys. Res. Commun. 2013, 431, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Callebaut, I.; Mornon, J.-P. From BRCA1 to RAP1: A Widespread BRCT Module Closely Associated with DNA Repair. FEBS Lett. 1997, 400, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Bork, P.; Hofmann, K.; Bucher, P.; Neuwald, A.F.; Altschul, S.F.; Koonin, E.V. A Superfamily of Conserved Domains in DNA Damage-Responsive Cell Cycle Checkpoint Proteins. FASEB J. 1997, 11, 68–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, J.; Boger, S.; Li, J.; Duncan, S.A. The Murine Pes1 Gene Encodes a Nuclear Protein Containing a BRCT Domain. Genomics 2000, 70, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Glover, J.N.M. Insights into the Molecular Basis of Human Hereditary Breast Cancer from Studies of the BRCA1 BRCT Domain. Fam. Cancer 2006, 5, 89–93. [Google Scholar] [CrossRef]
- Masi, A.; Antoccia, A. NBS1 Heterozygosity and Cancer Risk. Curr. Genomics 2008, 9, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Ḧlzel, M.; Grimm, T.; Rohrmoser, M.; Malamoussi, A.; Harasim, T.; Gruber-Eber, A.; Kremmer, E.; Eick, D. The BRCT Domain of Mammalian Pes1 Is Crucial for Nucleolar Localization and RRNA Processing. Nucleic Acids Res. 2007, 35, 789–800. [Google Scholar] [CrossRef]
- Peña-Guerrero, J.; Fernández-Rubio, C.; Burguete-Mikeo, A.; El-Dirany, R.; García-Sosa, A.T.; Nguewa, P. Discovery and Validation of Lmj_04_BRCT Domain, a Novel Therapeutic Target: Identification of Candidate Drugs for Leishmaniasis. Int. J. Mol. Sci. 2021, 22, 10493. [Google Scholar] [CrossRef]
- Wijayawardena, B.K.; Minchella, D.J.; DeWoody, J.A. Hosts, Parasites, and Horizontal Gene Transfer. Trends Parasitol. 2013, 29, 329–338. [Google Scholar] [CrossRef]
- Ives, A.; Ronet, C.; Prevel, F.; Ruzzante, G.; Fuertes-Marraco, S.; Schutz, F.; Zangger, H.; Revaz-Breton, M.; Lye, L.-F.; Hickerson, S.M.; et al. Leishmania RNA Virus Controls the Severity of Mucocutaneous Leishmaniasis. Science 2011, 331, 775–778. [Google Scholar] [CrossRef]
- Bourreau, E.; Ginouves, M.; Prévot, G.; Hartley, M.-A.; Gangneux, J.-P.; Robert-Gangneux, F.; Dufour, J.; Sainte-Marie, D.; Bertolotti, A.; Pratlong, F.; et al. Presence of Leishmania RNA Virus 1 in Leishmania Guyanensis Increases the Risk of First-Line Treatment Failure and Symptomatic Relapse. J. Infect. Dis. 2016, 213, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, J.L. Role of LRV1 and RNAi in the Pathogenesis of Leishmania. Trends Parasitol. 2017, 33, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Hecht, M.M.; Nitz, N.; Araujo, P.F.; Sousa, A.O.; Rosa, A.D.C.; Gomes, D.A.; Leonardecz, E.; Teixeira, A.R.L. Inheritance of DNA Transferred from American Trypanosomes to Human Hosts. PLoS ONE 2010, 5, e9181. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.R.L.; Argañaraz, E.R.; Freitas, L.H.; Lacava, Z.G.M.; Santana, J.M.; Luna, H. Possible Integration of Trypanosoma Cruzi KDNA Minicircles into the Host Cell Genome by Infection. Mutat. Res. Mol. Mech. Mutagen. 1994, 305, 197–209. [Google Scholar] [CrossRef]
- Lerch-Gaggl, A.; Haque, J.; Li, J.; Ning, G.; Traktman, P.; Duncan, S.A. Pescadillo Is Essential for Nucleolar Assembly, Ribosome Biogenesis, and Mammalian Cell Proliferation. J. Biol. Chem. 2002, 277, 45347–45355. [Google Scholar] [CrossRef] [Green Version]
- Di Masi, A.; Gullotta, F.; Cappadonna, V.; Leboffe, L.; Ascenzi, P. Cancer Predisposing Mutations in BRCT Domains. IUBMB Life 2011, 63, 503–512. [Google Scholar] [CrossRef]
- Long, L.; He, J.-Z.; Chen, Y.; Xu, X.-E.; Liao, L.-D.; Xie, Y.-M.; Li, E.-M.; Xu, L.-Y. Riboflavin Depletion Promotes Tumorigenesis in HEK293T and NIH3T3 Cells by Sustaining Cell Proliferation and Regulating Cell Cycle–Related Gene Transcription. J. Nutr. 2018, 148, 834–843. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Heras, G.; Domínguez–Berzosa, C.; Collantes, E.; Guadalajara, H.; García-Olmo, D.; García-Olmo, D.C. NIH-3T3 Fibroblasts Cultured with Plasma from Colorectal Cancer Patients Generate Poorly Differentiated Carcinomas in Mice. Cancer Lett. 2012, 316, 85–90. [Google Scholar] [CrossRef]
- Benskey, M.J.; Manfredsson, F.P. Lentivirus Production and Purification. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2016; Volume 1382, pp. 107–114. [Google Scholar]
- Dalm, M.C.F.; Cuijten, S.M.R.; Van Grunsven, W.M.J.; Tramper, J.; Martens, D.E. Effect of Feed and Bleed Rate on Hybridoma Cells in an Acoustic Perfusion Bioreactor: Part I. Cell Density, Viability, and Cell-Cycle Distribution. Biotechnol. Bioeng. 2004, 88, 547–557. [Google Scholar] [CrossRef]
- Shen, Y.; Sun, Y.; Zhang, L.; Liu, H. Effects of DTX3L on the Cell Proliferation, Adhesion, and Drug Resistance of Multiple Myeloma Cells. Tumor Biol. 2017, 39, 101042831770394. [Google Scholar] [CrossRef]
- Fu, Y.; Su, L.; Cai, M.; Yao, B.; Xiao, S.; He, Q.; Xu, L.; Yang, L.; Zhao, C.; Wan, T.; et al. Downregulation of CPA4 Inhibits Non Small–cell Lung Cancer Growth by Suppressing the AKT/C-MYC Pathway. Mol. Carcinog. 2019, 58, 2026–2039. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Pan, J.; Ji, L.; Song, S.; Lv, H.; Yang, Z.; Guo, Y. Carboxypeptidase A4 Promotes Cell Growth via Activating STAT3 and ERK Signaling Pathways and Predicts a Poor Prognosis in Colorectal Cancer. Int. J. Biol. Macromol. 2019, 138, 125–134. [Google Scholar] [CrossRef]
- Wang, C.; Zhao, N.; Zheng, Q.; Zhang, D.; Liu, Y. BHLHE41 Promotes U87 and U251 Cell Proliferation via ERK/CyclinD1 Signaling Pathway. Cancer Manag. Res. 2019, 11, 7657–7672. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Zhu, L.; Zhang, C.; Cui, X.; Lu, J. Overexpression of BHLHE41, Correlated with DNA Hypomethylation in 3’UTR Region, Promotes the Growth of Human Clear Cell Renal Cell Carcinoma. Oncol. Rep. 2019, 41, 2137–2147. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Ayrault, O.; Zindy, F.; Kim, J.H.; Roussel, M.F. Post-Transcriptional down-Regulation of Atoh1/Math1 by Bone Morphogenic Proteins Suppresses Medulloblastoma Development. Genes Dev. 2008, 22, 722–727. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Li, N.; Zhao, X.; Liao, C.; Ye, R.; Cheng, C.; Xu, Z.; Quan, J.; Liu, J.; Cao, Y. DHRS2 Mediates Cell Growth Inhibition Induced by Trichothecin in Nasopharyngeal Carcinoma. J. Exp. Clin. Cancer Res. 2019, 38, 300. [Google Scholar] [CrossRef]
- Nettersheim, D.; Berger, D.; Jostes, S.; Skowron, M.; Schorle, H. Deciphering the Molecular Effects of Romidepsin on Germ Cell Tumours: DHRS2 Is Involved in Cell Cycle Arrest but Not Apoptosis or Induction of Romidepsin Effectors. J. Cell. Mol. Med. 2019, 23, 670–679. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, H.; Long, Y.; Hua, H.; Jiang, Y.; Jing, J. PARP9 Is Overexpressed in Human Breast Cancer and Promotes Cancer Cell Migration. Oncol. Lett. 2018, 16, 4073–4077. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, S.; Guha, M.; Kashina, A.; Avadhani, N.G. Mitochondrial Dysfunction and Mitochondrial Dynamics-The Cancer Connection. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 602–614. [Google Scholar] [CrossRef]
- Hsu, C.-C.; Tseng, L.-M.; Lee, H.-C. Role of Mitochondrial Dysfunction in Cancer Progression. Exp. Biol. Med. 2016, 241, 1281–1295. [Google Scholar] [CrossRef]
- Lund, M.; Melbye, M.; Diaz, L.J.; Duno, M.; Wohlfahrt, J.; Vissing, J. Mitochondrial Dysfunction and Risk of Cancer. Br. J. Cancer 2015, 112, 1134–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheedy, D.M.; Dimitrova, D.; Rankin, J.K.; Bass, K.L.; Lee, K.M.; Tapia-Alveal, C.; Harvey, S.H.; Murray, J.M.; O’Connell, M.J. Brc1-Mediated DNA Repair and Damage Tolerance. Genetics 2005, 171, 457–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, G.P.; Brown, J.A.R.; Glover, J.N.M.; Kobor, M.S. Rtt107 BRCT Domains Act as a Targeting Module in the DNA Damage Response. DNA Repair Amst. 2016, 37, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, K.; Wakasugi, K.; Tomonaga, R.; Utsunomiya, K.; Miyamoto, H.; Fumoto, S.; Nishida, K. Effect of Metabolic Inhibitors on the Hepatic Disposition of 5-Fluorouracil after Application to the Rat Liver Surface. Biol. Pharm. Bull. 2016, 39, 361–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.Y.; Kim, J.Y.; Jung, Y.-J.; Kang, K. Toxicological Evaluation of the Topoisomerase Inhibitor, Etoposide, in the Model Animal Caenorhabditis Elegans and 3T3-L1 Normal Murine Cells. Environ. Toxicol. 2017, 32, 1836–1843. [Google Scholar] [CrossRef]
- Dasari, S.; Bernard Tchounwou, P. Cisplatin in Cancer Therapy: Molecular Mechanisms of Action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Gajski, G.; Gerić, M.; Žegura, B.; Novak, M.; Nunić, J.; Bajrektarević, D.; Garaj-Vrhovac, V.; Filipič, M. Genotoxic Potential of Selected Cytostatic Drugs in Human and Zebrafish Cells. Environ. Sci. Pollut. Res. 2016, 23, 14739–14750. [Google Scholar] [CrossRef]
- Negrei, C.; Hudita, A.; Ginghina, O.; Galateanu, B.; Voicu, S.N.; Stan, M.; Costache, M.; Fenga, C.; Drakoulis, N.; Tsatsakis, A.M. Colon Cancer Cells Gene Expression Signature As Response to 5- Fluorouracil, Oxaliplatin, and Folinic Acid Treatment. Front. Pharmacol. 2016, 7, 172. [Google Scholar] [CrossRef] [Green Version]
- Takebe, N.; Zhao, S.-C.; Ural, A.U.; Johnson, M.R.; Banerjee, D.; Diasio, R.B.; Bertino, J.R. Retroviral Transduction of Human Dihydropyrimidine Dehydrogenase CDNA Confers Resistance to 5-Fluorouracil in Murine Hematopoietic Progenitor Cells and Human CD34+-Enriched Peripheral Blood Progenitor Cells. Cancer Gene Ther. 2001, 8, 966–973. [Google Scholar] [CrossRef] [Green Version]
- Salonga, D.; Danenberg, K.D.; Johnson, M.; Metzger, R.; Groshen, S.; Tsao-Wei, D.D.; Lenz, H.-J.; Leichman, C.G.; Leichman, L.; Diasio, R.B.; et al. Colorectal Tumors Responding to 5-Fluorouracil Have Low Gene Expression Levels of Dihydropyrimidine Dehydrogenase, Thymidylate Synthase, and Thymidine Phosphorylase. Clin. Cancer Res. 2000, 6, 1322–1327. [Google Scholar]
- Guaragnella, N.; Giannattasio, S.; Moro, L. Mitochondrial Dysfunction in Cancer Chemoresistance. Biochem. Pharmacol. 2014, 92, 62–72. [Google Scholar] [CrossRef]
- Fernández-Rubio, C.; Campbell, D.; Vacas, A.; Ibañez, E.; Moreno, E.; Espuelas, S.; Calvo, A.; Palop, J.A.; Plano, D.; Sanmartin, C.; et al. Leishmanicidal Activities of Novel Methylseleno-Imidocarbamates. Antimicrob. Agents Chemother. 2015, 59, 5705–5713. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Harrow, J.; Denoeud, F.; Frankish, A.; Reymond, A.; Chen, C.K.; Chrast, J.; Lagarde, J.; Gilbert, J.G.R.; Storey, R.; Swarbreck, D.; et al. GENCODE: Producing a Reference Annotation for ENCODE. Genome Biol. 2006, 7 (Suppl. S1), 1–9. [Google Scholar] [CrossRef] [Green Version]
- Gentleman, R.C.; Carey, V.J.; Bates, D.M.; Bolstad, B.; Dettling, M.; Dudoit, S.; Ellis, B.; Gautier, L.; Ge, Y.; Gentry, J.; et al. Bioconductor: Open Software Development for Computational Biology and Bioinformatics. Genome Biol. 2004, 5, R80. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Larrea, E.; Aldabe, R.; Gonzalez, I.; Segura, V.; Sarobe, P.; Echeverria, I.; Prieto, J. Oncostatin M Enhances the Antiviral Effects of Type I Interferon and Activates Immunostimulatory Functions in Liver Epithelial Cells. J. Virol. 2009, 83, 3298–3311. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | 5-Fluorouracil (µM) | Etoposide (µM) | Cisplatin (µM) |
|---|---|---|---|
| HEK293T-Luc HEK293T-BRCT-A HEK293T-BRCT-B | 3.75 ± 1.06 21.75 ± 2.47 * 29.65 ± 6.58 * | 2.43 ± 0.09 1.92 ± 0.11 1.82 ± 0.11 | 0.58 ± 0.32 0.54 ± 0.23 0.30 ± 0.13 |
| NIH/3T3-Luc NIH/3T3-BRCT-A NIH/3T3-BRCT-B | 0.028 ± 0.002 1.26 ± 0.05 * 3.00 ± 0.14 * | 6.37 ± 1.29 24.44 ± 2.94 * 19.94 ± 0.78 * | 16.82 ± 0.25 21.52 ± 2.15 23.26 ± 1.77 |
| Categories | Diseases or Functions Annotation | Molecules |
|---|---|---|
| Metabolic Disease | Mitochondrial cytopathy | MT-ATP6, MT-ATP8, MT-ND1, MT-ND2, MT-ND3, MT-ND4, MT-ND4L, MT-ND5 |
| Cancer | Skin cancer | ABI3BP, ACE, ANG, B3GALT1, BMP2, C1R, CARD9, CDH7, CHST8, CLSTN2, CNKSR1, COL14A1, COL21A1, CPA4, CPNE7, CSMD3, CUL9, CYP1A1, DCHS2, DOK6, DPYD, DTX3L, EOMES, FGF13, FHAD1, FLRT2, FTCD, GABRE, GALNT5, GLIS1, GNB3, IFIH1, KCNJ4, LCP1, LDHD, LRAT, LRRN2, MAGEA3/MAGEA6, MDM1, MST1R, MT-ND5, MUC6, NAP1L2, NELL1, NLRP1, NPNT, NPR2, OR51B5, PARP9, PCK1, PDE1A, PLAT, PLIN4, PLXNA4, PRKCB, RPGRIP1, SLC7A3, SLCO2A1, SNX19, SPOCK3, SYNPO, THPO, TOX, TREX2, TRPC4, WDR66, ZC3H6, ZNF114, ZNF331, ZNF525 |
| Cell cycle | Cell cycle progression | BMP2, THPO |
| Nucleic acid metabolism | Metabolism of nucleic acid component or derivative | CYP1A1, DPYD, HAAO, HAND1, MT-ATP6, MT-ATP8, NPR2, PDE1A, PRKCB, PSMD5 |
| Cellular growth and proliferation | Expansion of cells | BMP2, DHRS2, DOK3, EOMES, LCP1, PLAT, S100A1, THPO, TOX |
| Cellular movement | Invasion of cancer cell lines | BHLHE41, MST1R, PLAT |
| Cell death and survival | Cell viability of cancer cell lines | CD24, MST1R, BMP2, THPO |
| Drug metabolism | Clearance of 5-fluorouracil | DPYD |
| Target Gene | Sense Primer (5’-3’) | Antisense Primer (5’-3’) |
|---|---|---|
| MT-ATP8 | ATGGCCCACCATAATTACCC | GTTCATTTTGGTTCTCAGGG |
| PARP9 | GTTAGTTTGCAAGGGAAGCC | AATTACATCTGCCGTCTGCC |
| MT-ATP6 | TTTCCCCCTCTATTGATCCC | AGCCTATAATCACTGTGCCC |
| MT-ND5 | CATCAGTTGATGATACGCCC | GAAGGGCTATTTGTTGTGGG |
| CARD9 | CTGTACAAGAAGGTCACAGG | AGTCATCTTTGGAGCTCAGC |
| PSMD5 | GCTTAACGAGAACCATAGGG | CAATCTGGGAAAGAGTGAGG |
| S100A1 | CAAGAAGGAGCTGAAAGAGC | TATGGAGAGGGATAAGTGGG |
| DTX3L | GGTGGATACTGTTCATTGGG | TGTCACTGCGTACTCTAAGC |
| BHLHE41 | GCAAGAGAGACAGTTACTGG | CCAAGACTACAGCTTTCTCC |
| DPYD | CAACTCTGTGTTCCACTTCG | AGGCATCTCATTGCTTCTCG |
| CPA4 | TACTGAAGTTCAGCACTGGG | CATATCCATCAGGATTGGCC |
| MT-ND3 | TAATCAACACCCTCCTAGCC | ACTCATAGGCCAGACTTAGG |
| MT-ND2 | ATTTCCTCACGCAAGCAACC | TCATGTGAGAAGAAGCAGGC |
| DOK3 | TCCTCAGGATCCACAGATGC | CCTCAGTCCTCTGCACCACC |
| BMP2 | ACATGCTAGACCTGTATCGC | GTTTTCCCACTCGTTTCTGG |
| DHRS2 | ATGCTGTCAGCAGTTGCCCG | CCGATCCCACTGGTGGACCC |
| β-actin | AGCCTCGCCTTTGCCGA | CTGGTGCCTGGGGCG |
| LmjBRCT | TCTTCATATCGCGTGAGGTG | CATGCTTTTTCATCCCTGGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larrea, E.; Fernández-Rubio, C.; Peña-Guerrero, J.; Guruceaga, E.; Nguewa, P.A. The BRCT Domain from the Homologue of the Oncogene PES1 in Leishmania major (LmjPES) Promotes Malignancy and Drug Resistance in Mammalian Cells. Int. J. Mol. Sci. 2022, 23, 13203. https://doi.org/10.3390/ijms232113203
Larrea E, Fernández-Rubio C, Peña-Guerrero J, Guruceaga E, Nguewa PA. The BRCT Domain from the Homologue of the Oncogene PES1 in Leishmania major (LmjPES) Promotes Malignancy and Drug Resistance in Mammalian Cells. International Journal of Molecular Sciences. 2022; 23(21):13203. https://doi.org/10.3390/ijms232113203
Chicago/Turabian StyleLarrea, Esther, Celia Fernández-Rubio, José Peña-Guerrero, Elizabeth Guruceaga, and Paul A. Nguewa. 2022. "The BRCT Domain from the Homologue of the Oncogene PES1 in Leishmania major (LmjPES) Promotes Malignancy and Drug Resistance in Mammalian Cells" International Journal of Molecular Sciences 23, no. 21: 13203. https://doi.org/10.3390/ijms232113203
APA StyleLarrea, E., Fernández-Rubio, C., Peña-Guerrero, J., Guruceaga, E., & Nguewa, P. A. (2022). The BRCT Domain from the Homologue of the Oncogene PES1 in Leishmania major (LmjPES) Promotes Malignancy and Drug Resistance in Mammalian Cells. International Journal of Molecular Sciences, 23(21), 13203. https://doi.org/10.3390/ijms232113203