


Localization of the Neuropeptide Arginine Vasotocin and Its Receptor in the Osmoregulatory Organs of Black Porgy, Acanthopagrus schlegelii: Gills, Kidneys, and Intestines

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
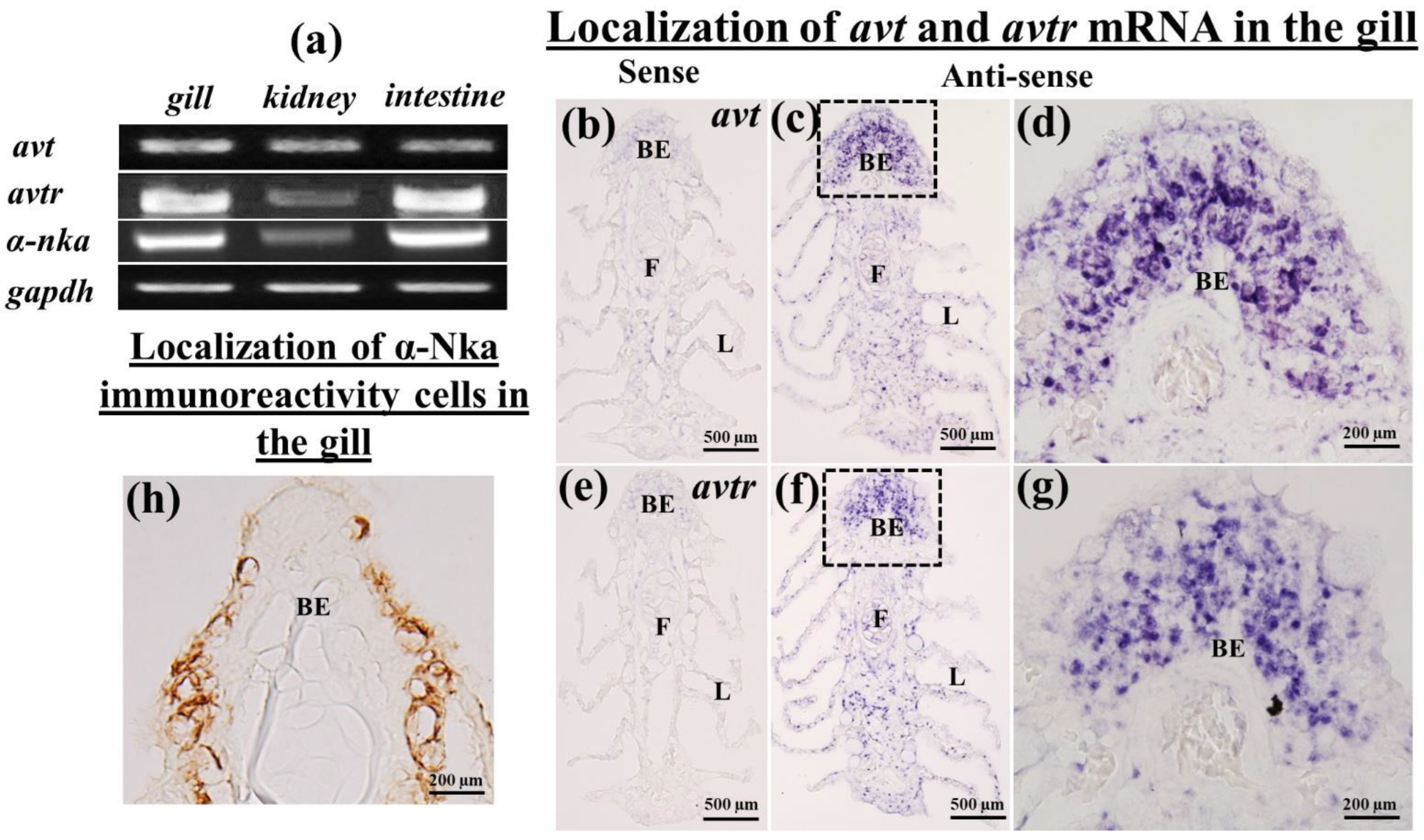
2.1. Tissue Distribution of avt, avtr, and a-nka mRNA in the Gills, Kidneys, and Intestines
2.2. Localization of avt and avtr mRNA in the Gills of the Black Porgy
2.3. Localization of avt and avtr mRNA in the Kidneys and Intestines of the Black Porgy
2.4. Immunolocalization of a-Nka-ir Cells in the Gills, Kidneys, and Intestines of the Black Porgy
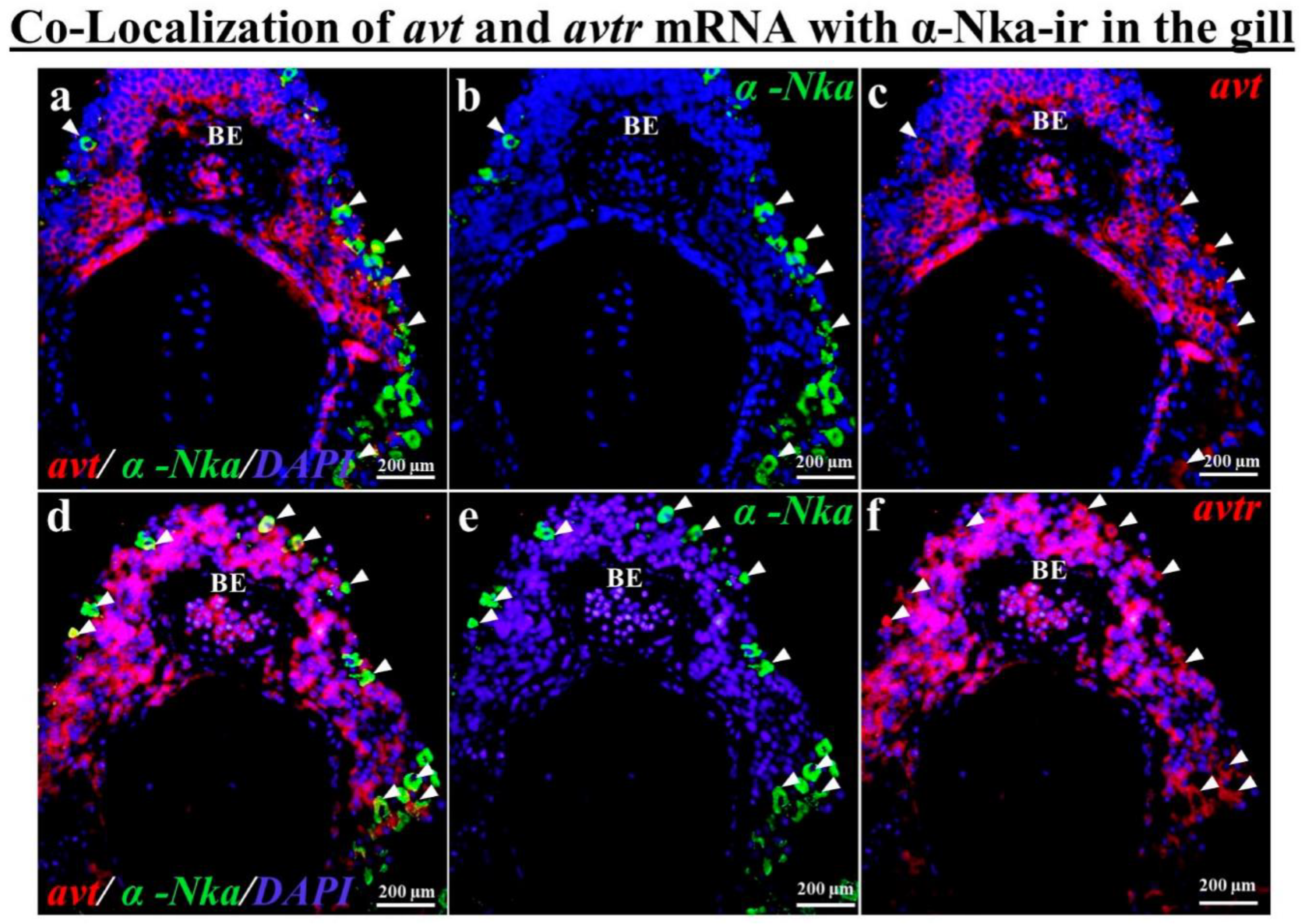
2.5. Co-Localization of avt and avtr mRNA with a-Nka-ir Cells in the Gills, Kidneys, and Intestines of the Black Porgy
3. Discussion
4. Materials and Methods
4.1. Experimental Fish
4.2. RNA Extraction and Cloning
4.3. Tissue Distribution of avt, avtr and α-nka in the Gills, Kidneys, and Intestines
4.4. In Situ Hybridization
4.5. Immunohistochemistry
4.6. Co-Localization of avt and avtr mRNA with α-Nka in the Gills, Kidneys, and Intestines
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Acher, R. Neurohypophysial peptide systems: Processing machinery, hydroosmotic regulation, adaptation and evolution. Regul. Pept. 1993, 45, 1–13. [Google Scholar] [CrossRef]
- Lema, S.C. Identification of multiple vasotocin receptor cDNAs in teleost fish: Sequences, phylogenetic analysis, sites of expression, and regulation in the hypothalamus and gill in response to hyperosmotic challenge. Mol. Cell Endocrinol. 2010, 321, 215–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lema, S.C.; Washburn, E.H.; Crowley, M.C.; Carvalho, P.G.; Egelston, J.N.; McCormick, S.D. Evidence for a role of arginine vasotocin receptors in the gill during salinity acclimation by a euryhaline teleost fish. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 316, 735–750. [Google Scholar] [CrossRef] [PubMed]
- Maetz, J.; Bourguet, J.; Lahlough, B.; Hourdry, J. Neurohypophysial Peptides and Osmoregulation in Carassius Auratus. Gen. Comp. Endocrinol. 1964, 47, 508–522. [Google Scholar] [CrossRef]
- Moon, T.W.; Mommsen, T.P. Vasoactive peptides and phenylephrine actions in isolated teleost hepatocytes. Am. J. Physiol. Endocrinol. Metabol. 1990, 259, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Masuda, H.; Amaoka, K.; Araga, C.; Uyeno, T.; Yoshino, T.; Muzik, K. The Fishes of the Japanese Archipelago; Tokai University Press: Tokyo, Japan, 1984; Volume 1, p. 437. [Google Scholar]
- Hong, W.S.; Zhang, Q.Y. Review of captive bred species and fry production of marine fish in China. Aquaculture 2003, 227, 305–318. [Google Scholar] [CrossRef]
- Shao, Q.J.; Ma, J.J.; Xu, Z.R.; Hu, W.L.; Xu, J.Z.; Xie, S.Q. Dietary phosphorus requirement of juvenile black sea bream, Sparus macrocephalus. Aquaculture 2008, 277, 92–100. [Google Scholar] [CrossRef]
- Takei, Y.; Hiroi, J.; Takahashi, H.; Sakamoto, T. Diverse mechanisms for body fluid regulation in teleost fishes. Am. J. Physiol. 2014, 307, 778–792. [Google Scholar] [CrossRef] [Green Version]
- Warren, I.A.; Ciborowski, K.L.; Casadei, E.; Hazlerigg, D.G.; Martin, S.; Jordan, W.C. Extensive local gene duplication and functional divergence among paralogs in Atlantic salmon. Genome Biol. Evol. 2014, 6, 1790–1805. [Google Scholar] [CrossRef] [Green Version]
- Takvam, M.; Wood, C.M.; Kryvi1, H.; Nilsen, T.O. Ion Transporters and Osmoregulation in the Kidney of Teleost Fishes as a Function of Salinity. Front. Physiol. 2021, 12, 664588. [Google Scholar] [CrossRef]
- Godwin, J.; Sawby, R.; Warner, R.R.; Crews, D.; Grober, M.S. Hypothalamic arginine vasotocin mRNA abundance variation across sexes and with sex change in a coral reef fish. Brain Behav. Evol. 2000, 55, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Goodson, J.L.; Bass, A.H. Forebrain peptides modulate sexually polymorphic vocal circuitry. Nature 2000, 403, 769–772. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Muro, T.; Kimura, Y.; Li, S.; Islam, Z.; Ogoshi, M. Differential expression of Na+-Cl− cotransporter and Na+-K+-Cl− cotransporter 2 in the distal nephrons of euryhaline and seawater pufferfishes. Am. J. Physiol. 2011, 300, 284–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, S.D.; Farrell, A.P.; Brauner, C.J. Euryhaline fishes. In Fish Physiology; Farrell, A.P., Brauner, C.J., Eds.; Elsevier: Waltham, MA, USA, 2013; pp. 477–533. [Google Scholar]
- Féraille, E.; Doucet, A. Sodium Potassium Adenosine triphosphatase Dependent Sodium Transport in the Kidney: Hormonal Control. Phy. Rev. 2001, 81, 345–418. [Google Scholar]
- Gonin, S.; Deschenes, G.; Roger, F.; Bens, M.; Martin, P.Y.; Carpentier, J.L.; Vandewalle, A.; Doucet, A.; Feraille, E. Cyclic AMP increases cell surface expression of functional Na-K-ATPase units in mammalian cortical collecting duct principal cells. Mol. Biol. Cell 2001, 12, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef]
- Tatsuo, S. The structure of the kidney from the freshwater teleost Carassius auratus. Anat. Embryol. 1985, 171, 31–39. [Google Scholar]
- Kazuhiro, U.; Kiyoshi, S.; Naotaka, O.; Shinji, A.; Kohei, Y. Localization of Na+, K+-ATPase in tissues of rabbit and teleosts using an antiserum directed against a partial sequence of the α-Subunit. Zoo Sci. 1996, 13, 219–227. [Google Scholar]
- Seidelin, M.; Madsen, S.S.; Blenstrup, H.; Tipsmark, C.K. Time-course changes in the expression of Na+, K+-ATPase in gills and pyloric caeca of brown trout (Salmo trutta) during acclimation to seawater. Physiol. Biochem. Zoo 2000, 73, 446–453. [Google Scholar] [CrossRef]
- Veillette, P.A.; White, R.J.; Specker, J.L.; Young, G. Osmoregulatory physiology of pyloric ceca: Regulated and adaptive changes in Chinook salmon. J. Exp. Zool. Part A Comp. Exp. Biol. 2005, 303, 608–613. [Google Scholar] [CrossRef]
- Hwang, P.P.; Lee, T.H. New insights into fish ion regulation and mitochondrion-rich cells. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 148, 479–497. [Google Scholar] [CrossRef]
- Marshall, W.S.; Grosell, M. Ion transport, osmoregulation, and acidbase balance. In The Physiology of Fishes; Evans, D.H., Claiborne, J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2006; Volume 16, pp. 177–230. [Google Scholar]
- Whittamore, J.M. Osmoregulation and epithelial water transport: Lessons from the intestine of marine teleost fish. J. Comp. Physiol. B 2012, 182, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Cádiz, L.; Padilla, J.R.; Gozdowska, M.; Kulczykowska, E.; Rodríguez, G.M.; Mancera, J.M.; Martos-Sitcha. J.A. Cortisol modulates vasotocinergic and isotocinergic pathways in the gilthead sea bream. J. Exp. Biol. 2015, 218, 316–325. [Google Scholar]
- Aruna, A.; Lin, C.-J.; Nagarajan, G.; Chang, C.-F. Neurohypophysial Hormones Associated with Osmotic Challenges in the Brain and Pituitary of the Euryhaline Black Porgy, Acanthopagrus schlegelii. Cells 2021, 10, 3086. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, G.; Aruna, A.; Chang, C.F. Neuropeptide Arginine Vasotocin Positively Interacts with Neurosteroidogenesis in the Early Brain of Orange-Spotted Grouper, Epinephelus coioides. J. Neuroendocrinol. 2015, 9, 718–736. [Google Scholar] [CrossRef]
- Balment, R.J.; Lu, W.; Weybourne, E.; Warne, J.M. Arginine vasotocin a key hormone in fish physiology and behaviour: A review with insights from mammalian models. Gen. Comp. Endocrinol. 2006, 15, 9–16. [Google Scholar] [CrossRef]
- Bennett, M.B.; Rankin, J.C. The eVect of neurohypophysial hormones on the vascular resistance of the isolated perfused gill of the European eel Anguilla anguilla. Gen. Comp. Endocrinol. 1986, 64, 60–66. [Google Scholar] [CrossRef]
- Olson, K.R. Gill circulation: Regulation of perfusion distribution and metabolism of regulatory molecules. J. Exp. Zool. 2002, 293, 320–335. [Google Scholar] [CrossRef]
- Marshall, W.S. Rapid regulation of NaCl secretion by estuarine teleost fish: Coping strategies for short-duration fresh water exposures. Biochim. Biophys. Acta (BBA)—Biomembr. 2003, 1618, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.M.; Laurent, P. Fish gill morphology: Inside out. J. Exp. Zool. 2002, 293, 192–213. [Google Scholar] [CrossRef]
- Sakamoto, T.; Uchida, K.; Yokota, S. Regulation of the ion-transporting mitochondrion-rich cell during adaptation of teleost fishes to different salinities. Zoo. Sci. 2001, 18, 1163–1174. [Google Scholar] [CrossRef] [Green Version]
- Avella, M.; Masoni, A.; Bornancin, M.; Mayer-Gostan, N. Gill morphology and sodium influx in the rainbow trout (Salmo gairdneri) acclimated to artificial freshwater environments. J. Exp. Zool. 1987, 241, 159–169. [Google Scholar] [CrossRef]
- Uchida, K.; Kaneko, T.; Yamauchi, K.; Hirano, T. Morphometrical analysis of chloride cell activity in the gill filaments and lamellae and changes in Na+,K+-ATPase activity during seawater adaptation in chum salmon fry. J. Exp. Zool. 1996, 276, 193–200. [Google Scholar] [CrossRef]
- Sasai, S.; Kaneko, T.; Hasegawa, S.; Tsukamoto, K. Morphological alteration in two types of gill chloride cells in Japanese eel (Anguilla japonica) during catadromous migration. Can. J. Zool. 1998, 76, 1480–1487. [Google Scholar] [CrossRef]
- Hirai, N.; Tagawa, M.; Kaneko, T.; Seikai, T.; Tanaka, M. Distributional changes in branchial chloride cells during freshwater adaptation in Japanese sea bass Lateolabrax japonicus. Zool. Sci. 1999, 16, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Versamos, S.; Diaz, J.P.; Charmantier, G.; Flik, G.; Blasco, C.; Connes, R. Branchial chloride cells in sea bass (Dicentrarchus labrax) adapted to fresh water, seawater, and doubly concentrated seawater. J. Exp. Zool. 2002, 293, 12–26. [Google Scholar] [CrossRef]
- Hirose, S.; Kaneko, T.; Naito, N.; Takei, Y. Molecular biology of major components of chloride cells. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 136, 593–620. [Google Scholar] [CrossRef]
- Lin, C.H.; Tsaib, R.S.; Leea, T.H. Expression and distribution of Na, K-ATPase in gill and kidney of the spotted green pufferfish, Tetraodon nigroviridis, in response to salinity challenge. Comp. Biochem. Physiol. Part A 2004, 138, 287–295. [Google Scholar] [CrossRef]
- Dressler, G.R. The cellular basis of kidney development. Annu. Rev. Cell Dev. Biol. 2006, 22, 509–529. [Google Scholar] [CrossRef]
- Carroll, R.G. Renal System and Urinary Tract. In Elsevier’s Integrated Physiology; Elsevier: Amsterdam, The Netherlands, 2007; pp. 117–137. [Google Scholar]
- Grosell, M.; Taylor, J.R. Intestinal anion exchange in teleost water balance. Comp. Biochem. Physiol. 2007, 148A, 14–22. [Google Scholar] [CrossRef]
- Crosnier, C.; Stamataki, D.; Lewis, J. Organizing cell renewal in the intestine: Stem cells, signals and combinatorial control. Nat. Rev. Genet. 2006, 7, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Leblond, C.P. Origin, differentiation and renewal of the four main epithelial cell types in the mouse small intestine I. Columnar cell. Am. J. Anat. 1974, 141, 461–479. [Google Scholar] [CrossRef]
- Phillips, A.D.; France, N.E.; Walker-Smith, J.A. The structure of the enterocyte in relation to its position on the villus in childhood: An electron microscopical study. Histopathology 1979, 3, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Madsen, S.S.; Engelund, M.B.; Cutler, C.P. Water transport and functional dynamics of aquaporins in osmoregulatory organs of fishes. Biol. Bull. 2015, 229, 70–92. [Google Scholar] [CrossRef] [PubMed]
- Sundell, K.; Sundh, H. Intestinal fluid absorption in anadromous salmonids: Importance of tight junctions and aquaporins. Front. Physiol. 2012, 3, 388. [Google Scholar] [CrossRef] [Green Version]
- Sundh, H.; Nilsen, T.O.; Lindström, J.; Hasselberg-Frank, L.; Stefansson, S.O.; McCormick, S.D. Development of intestinal ion-transporting mechanisms during smoltification and seawater acclimation in Atlantic salmon Salmo salar. J. Fish Biol. 2014, 85, 1227–1252. [Google Scholar] [CrossRef]
- Chew, S.F.; Tng, Y.Y.M.; Wee, N.L.J.; Tok, C.Y.; Wilson, J.M.; Ip, Y.K. Intestinal osmoregulatory acclimation and nitrogen metabolism in juveniles of the freshwater marble goby exposed to seawater. J. Comp. Physiol. B 2010, 180, 511–520. [Google Scholar] [CrossRef]
- Ferlazzo, A.; Carvalho, E.S.; Gregorio, S.F.; Power, D.M.; Canario, A.V.; Trischitta, F.; Fuentes, J. Prolactin regulates luminal bicarbonate secretion in the intestine of the seabream (Spaurua aurata. L.). Exp. Biol. 2012, 215, 3836–3844. [Google Scholar]
- Wallace, K.N.; Akhter, S.; Smith, E.M.; Lorent, K.; Pack, M. Intestinal growth and differentiation in zebrafish. Mech. Develop. 2005, 122, 157–173. [Google Scholar] [CrossRef]
- Marshall, W.S.; Howard, J.A.; Cozzi, R.R.; Lynch, E.M. NaCl and fluid secretionby the intestine of the teleost Fundulus heteroclitus: Involvement of CFTR. J. Exp. Biol. 2002, 205, 745–758. [Google Scholar] [CrossRef]
- Grosell, M.; Gilmour, K.M.; Perry, S.F. Intestinal carbonic anhydrase, bicarbonate, and proton carriers play a role in the acclimation of rainbow trout to seawater. Am. J. Physiol. 2007, 293, R2099–R2111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tresguerres, M.; Levin, L.R.; Buck, J.; Grosell, M. Modulation of NaCl absorption by [HCO3−] in the marine teleost intestine is mediated by soluble adenylyl cyclase. Am. J. Physiol. 2010, 299, 62–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, S.S.; Olesen, J.H.; Bedal, K.; Engelund, M.B.; Velasco-Santamaria, Y.M.; Tipsmark, C.K. Functional characterization of water transport and cellular localization of three aquaporin paralogs in the salmonid intestine. Front. Physiol. 2011, 2, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aruna, A.; Nagarajan, G.; Chang, C.F. Differential expression pattern and localization of glucocorticoid and mineralocorticoid receptors transcripts in osmoregulatory organs of freshwater and seawater acclimated tilapia. Gen. Comp. Endocrinol. 2012, 179, 465–476. [Google Scholar] [CrossRef]
- Balment, R.J. Control of water balance in mammals. In Osmoregulation and Drinking in Vertebrates; Hazon, N., Flik, G., Eds.; Bios ScientiWc Publishers Ltd.: Oxford, UK, 2002; pp. 153–168. [Google Scholar]
- Aruna, A.; Nagarajan, G.; Chang, C.F. Involvement of corticotrophin releasing hormone and corticosteroid receptor in the brain-pituitary-gill of tilapia (Oreochromis mossambicus) during the course of seawater acclimation. J. Neuroendocrinol. 2012, 24, 818–830. [Google Scholar] [CrossRef]
- Aruna, A.; Nagarajan, G.; Chang, C.F. The acute salinity changes activate the dual pathways of endocrine responses in the brain and pituitary of tilapia. Gen. Comp. Endocrinol. 2015, 211, 154–164. [Google Scholar] [CrossRef]
- Nagarajan, G.; Aruna, A.; Chang, C.F. Neurosteroidogenic enzymes and their regulation in the early brain of the protogynous grouper Epinephelus coioides: A possible role in gonadal sex differentiation. Gen. Comp. Endocrinol. 2013, 181, 271–287. [Google Scholar] [CrossRef]
- Aruna, A.; Lan, D.S.; Nagarajan, G.; Wang, T.P.; Cao, J.C.; Chen, Y.H.; Chang, C. Differential expression of hypothalamic gill-crh system with osmotic stress in the euryjaline blackporgy, Acanthopagrus Schlegelii. Front. Physiol. 2021, 12, 768122. [Google Scholar] [CrossRef]
- Nagarajan, G.; Aruna, A.; Yousef, A.A.; Roshman, T.M.; Chang, C.F. Expression and Localization of star, sf1 and dax-1 in the early brain of the orange spotted grouper Epinephelus coioides. Int. Sci. Mol. Sci. 2022, 23, 2614. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Orientation | Nucleotide Sequence (5′–3′) | Usage |
|---|---|---|---|
| avt | F | 5′-CTGTGCGTCCTGGGACTCATC-3′ | RT-PCR |
| R | 5′-GGAGCTCTGAGGCATGTTTTG-3′ | RT-PCR | |
| S | 5′- CTCCTCCGCCTGCTACATC-3′ | in situ | |
| AS | 5′- GGCTGACTCTTCAATCTTC-3′ | in situ | |
| avt-r | F | 5′-CCTGCTGGCAATGTACAACAC-3′ | RT-PCR |
| R | 5′-GGCGCCCAGCAGATAATGTAC-3′ | RT-PCR | |
| S | 5′-CAGGTCTCTGGTCTTCAG-3′ | in situ | |
| AS | 5′-GCTCTGGATATCGTTGTAAC-3′ | in situ | |
| gapdh | F | 5′-GGCCCCCCTGGCCAAAGT-3′ | RT-PCR |
| R | 5′-TGGGTGTCACCGATGAAG-3′ | RT-PCR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagarajan, G.; Adimoolam, A.; Alkhamis, Y.A.; Mathew, R.T.; Chang, C.-F. Localization of the Neuropeptide Arginine Vasotocin and Its Receptor in the Osmoregulatory Organs of Black Porgy, Acanthopagrus schlegelii: Gills, Kidneys, and Intestines. Int. J. Mol. Sci. 2022, 23, 13421. https://doi.org/10.3390/ijms232113421
Nagarajan G, Adimoolam A, Alkhamis YA, Mathew RT, Chang C-F. Localization of the Neuropeptide Arginine Vasotocin and Its Receptor in the Osmoregulatory Organs of Black Porgy, Acanthopagrus schlegelii: Gills, Kidneys, and Intestines. International Journal of Molecular Sciences. 2022; 23(21):13421. https://doi.org/10.3390/ijms232113421
Chicago/Turabian StyleNagarajan, Ganesan, Aruna Adimoolam, Yousef Ahmed Alkhamis, Roshmon Thomas Mathew, and Ching-Fong Chang. 2022. "Localization of the Neuropeptide Arginine Vasotocin and Its Receptor in the Osmoregulatory Organs of Black Porgy, Acanthopagrus schlegelii: Gills, Kidneys, and Intestines" International Journal of Molecular Sciences 23, no. 21: 13421. https://doi.org/10.3390/ijms232113421
APA StyleNagarajan, G., Adimoolam, A., Alkhamis, Y. A., Mathew, R. T., & Chang, C.-F. (2022). Localization of the Neuropeptide Arginine Vasotocin and Its Receptor in the Osmoregulatory Organs of Black Porgy, Acanthopagrus schlegelii: Gills, Kidneys, and Intestines. International Journal of Molecular Sciences, 23(21), 13421. https://doi.org/10.3390/ijms232113421