
Fungal Effectoromics: A World in Constant Evolution

 , ,
, ,
Abstract
:1. Introduction
2. Fundamentals of Effector Biology
3. Effector Identification: Past and Present
4. Effector Conservation: Effectors Shedding the “Species-Specific” Label
5. Effector Targets: Beyond the Apoplast
6. Effector Nature: The Rise of Non-Proteinaceous Effectors
6.1. RNA Effectors
6.2. Secondary Metabolite Effectors
7. Effectors: Not Just Plant-Pathogenic Molecules
8. Discussion: Coming Changes to Effectoromics
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Kamoun, S. A Catalogue of the Effector Secretome of Plant Pathogenic Oomycetes. Annu. Rev. Phytopathol. 2006, 44, 41–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamoun, S. Groovy Times: Filamentous Pathogen Effectors Revealed. Curr. Opin. Plant Biol. 2007, 10, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Van der Hoorn, R.A.L.; Terauchi, R.; Kamoun, S. Emerging Concepts in Effector Biology of Plant-Associated Organisms. Mol. Plant-Microbe Interact. 2009, 22, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraldo, M.C.; Valent, B. Filamentous Plant Pathogen Effectors in Action. Nat. Rev. Microbiol. 2013, 11, 800–814. [Google Scholar] [CrossRef] [PubMed]
- Selin, C.; de Kievit, T.R.; Belmonte, M.F.; Fernando, W.G.D. Elucidating the Role of Effectors in Plant-Fungal Interactions: Progress and Challenges. Front. Microbiol. 2016, 7, 00600. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Li, C.; Si, J.; Han, Z.; Chen, D. Action Mechanisms of Effectors in Plant-Pathogen Interaction. Int. J. Mol. Sci. 2022, 23, 6758. [Google Scholar] [CrossRef]
- Koeck, M.; Hardham, A.R.; Dodds, P.N. The Role of Effectors of Biotrophic and Hemibiotrophic Fungi in Infection. Cell. Microbiol. 2011, 13, 1849–1857. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Yuan, Q.; Tang, J.; Huang, J.; Hsiang, T.; Wei, Y.; Zheng, L. Colletotrichum higginsianum as a Model for Understanding Host–Pathogen Interactions: A Review. Int. J. Mol. Sci. 2018, 19, 2142. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Kahmann, R. Cell Wall–Associated Effectors of Plant-Colonizing Fungi. Mycologia 2021, 113, 247–260. [Google Scholar] [CrossRef]
- Kim, K.-T.; Jeon, J.; Choi, J.; Cheong, K.; Song, H.; Choi, G.; Kang, S.; Lee, Y.-H. Kingdom-Wide Analysis of Fungal Small Secreted Proteins (SSPs) Reveals Their Potential Role in Host Association. Front. Plant Sci. 2016, 7, 00186. [Google Scholar] [CrossRef]
- Shao, D.; Smith, D.L.; Kabbage, M.; Roth, M.G. Effectors of Plant Necrotrophic Fungi. Front. Plant Sci. 2021, 12, 995. [Google Scholar] [CrossRef] [PubMed]
- Samperna, S.; Masi, M.; Vurro, M.; Evidente, A.; Marra, M. Cyclopaldic Acid, the Main Phytotoxic Metabolite of Diplodia Cupressi, Induces Programmed Cell Death and Autophagy in Arabidopsis Thaliana. Toxins 2022, 14, 474. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.; Aerts, A.; Ahrén, D.; Brun, A.; Danchin, E.G.J.; Duchaussoy, F.; Gibon, J.; Kohler, A.; Lindquist, E.; Pereda, V.; et al. The Genome of Laccaria bicolor Provides Insights into Mycorrhizal Symbiosis. Nature 2008, 452, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Plett, J.M.; Kemppainen, M.; Kale, S.D.; Kohler, A.; Legué, V.; Brun, A.; Tyler, B.M.; Pardo, A.G.; Martin, F. A Secreted Effector Protein of Laccaria bicolor Is Required for Symbiosis Development. Curr. Biol. 2011, 21, 1197–1203. [Google Scholar] [CrossRef] [Green Version]
- Voß, S.; Betz, R.; Heidt, S.; Corradi, N.; Requena, N. RiCRN1, a Crinkler Effector From the Arbuscular Mycorrhizal Fungus Rhizophagus irregularis, Functions in Arbuscule Development. Front. Microbiol. 2018, 9, 2068. [Google Scholar] [CrossRef] [PubMed]
- Wawra, S.; Fesel, P.; Widmer, H.; Timm, M.; Seibel, J.; Leson, L.; Kesseler, L.; Nostadt, R.; Hilbert, M.; Langen, G.; et al. The Fungal-Specific β-Glucan-Binding Lectin FGB1 Alters Cell-Wall Composition and Suppresses Glucan-Triggered Immunity in Plants. Nat. Commun. 2016, 7, 13188. [Google Scholar] [CrossRef]
- Snelders, N.C.; Rovenich, H.; Thomma, B.P.H.J. Microbiota Manipulation through the Secretion of Effector Proteins Is Fundamental to the Wealth of Lifestyles in the Fungal Kingdom. FEMS Microbiol. Rev. 2022, 46, fuac022. [Google Scholar] [CrossRef] [PubMed]
- Redkar, A.; Sabale, M.; Zuccaro, A.; Di Pietro, A. Determinants of Endophytic and Pathogenic Lifestyle in Root Colonizing Fungi. Curr. Opin. Plant Biol. 2022, 67, 102226. [Google Scholar] [CrossRef]
- Hulin, M.T.; Armitage, A.D.; Vicente, J.G.; Holub, E.B.; Baxter, L.; Bates, H.J.; Mansfield, J.W.; Jackson, R.W.; Harrison, R.J. Comparative Genomics of Pseudomonas syringae Reveals Convergent Gene Gain and Loss Associated with Specialization onto Cherry (Prunus avium). New Phytol. 2018, 219, 672–696. [Google Scholar] [CrossRef] [Green Version]
- Frantzeskakis, L.; Di Pietro, A.; Rep, M.; Schirawski, J.; Wu, C.-H.; Panstruga, R. Rapid Evolution in Plant–Microbe Interactions –a Molecular Genomics Perspective. New Phytol. 2020, 225, 1134–1142. [Google Scholar] [CrossRef]
- Gómez Luciano, L.B.; Tsai, I.J.; Chuma, I.; Tosa, Y.; Chen, Y.-H.; Li, J.-Y.; Li, M.-Y.; Lu, M.-Y.J.; Nakayashiki, H.; Li, W.-H. Blast Fungal Genomes Show Frequent Chromosomal Changes, Gene Gains and Losses, and Effector Gene Turnover. Mol. Biol. Evol. 2019, 36, 1148–1161. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sun, M.; Zhang, Y.; Song, Z.; Zhang, S.; Zhang, Q.; Xu, J.-R.; Liu, H. Extensive Chromosomal Rearrangements and Rapid Evolution of Novel Effector Superfamilies Contribute to Host Adaptation and Speciation in the Basal Ascomycetous Fungi. Mol. Plant Pathol. 2020, 21, 330–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leelarasamee, N.; Zhang, L.; Gleason, C. The Root-Knot Nematode Effector MiPFN3 Disrupts Plant Actin Filaments and Promotes Parasitism. PLoS Pathog. 2018, 14, e1006947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kud, J.; Wang, W.; Gross, R.; Fan, Y.; Huang, L.; Yuan, Y.; Gray, A.; Duarte, A.; Kuhl, J.C.; Caplan, A.; et al. The Potato Cyst Nematode Effector RHA1B Is a Ubiquitin Ligase and Uses Two Distinct Mechanisms to Suppress Plant Immune Signaling. PLoS Pathog. 2019, 15, e1007720. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, P.A.; Escudero-Martinez, C.; Bos, J.I.B. An Aphid Effector Targets Trafficking Protein VPS52 in a Host-Specific Manner to Promote Virulence. Plant Physiol. 2017, 173, 1892–1903. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.-X.; Qian, L.-X.; Wang, X.-W.; Shao, R.-X.; Hong, Y.; Liu, S.-S.; Wang, X.-W. A Salivary Effector Enables Whitefly to Feed on Host Plants by Eliciting Salicylic Acid-Signaling Pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Plett, J.M.; Yin, H.; Mewalal, R.; Hu, R.; Li, T.; Ranjan, P.; Jawdy, S.; Paoli, H.C.D.; Butler, G.; Burch-Smith, T.M.; et al. Populus trichocarpa Encodes Small, Effector-like Secreted Proteins That Are Highly Induced during Mutualistic Symbiosis. Sci. Rep. 2017, 7, 382. [Google Scholar] [CrossRef] [Green Version]
- Su, C.; Liu, H.; Wafula, E.K.; Honaas, L.; de Pamphilis, C.W.; Timko, M.P. SHR4z, a Novel Decoy Effector from the Haustorium of the Parasitic Weed Striga gesnerioides, Suppresses Host Plant Immunity. New Phytol. 2020, 226, 891–908. [Google Scholar] [CrossRef] [Green Version]
- Alfano, J.R.; Collmer, A. Type III Secretion System Effector Proteins: Double Agents in Bacterial Disease and Plant Defense. Annu. Rev. Phytopathol. 2004, 42, 385–414. [Google Scholar] [CrossRef] [Green Version]
- Stergiopoulos, I.; de Wit, P.J.G.M. Fungal Effector Proteins. Annu. Rev. Phytopathol. 2009, 47, 233–263. [Google Scholar] [CrossRef]
- Friesen, T.L.; Faris, J.D.; Solomon, P.S.; Oliver, R.P. Host-Specific Toxins: Effectors of Necrotrophic Pathogenicity. Cell. Microbiol. 2008, 10, 1421–1428. [Google Scholar] [CrossRef] [PubMed]
- Stergiopoulos, I.; Collemare, J.; Mehrabi, R.; De Wit, P.J.G.M. Phytotoxic Secondary Metabolites and Peptides Produced by Plant Pathogenic Dothideomycete Fungi. FEMS Microbiol. Rev. 2013, 37, 67–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collemare, J.; O’Connell, R.; Lebrun, M.-H. Nonproteinaceous Effectors: The Terra Incognita of Plant–Fungal Interactions. New Phytol. 2019, 223, 590–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiberg, A.; Wang, M.; Lin, F.-M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.-D.; Jin, H. Fungal Small RNAs Suppress Plant Immunity by Hijacking Host RNA Interference Pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Weiberg, A.; Dellota, E.; Yamane, D.; Jin, H. Botrytis Small RNA Bc-SiR37 Suppresses Plant Defense Genes by Cross-Kingdom RNAi. RNA Biol. 2017, 14, 421–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Presti, L.; Lanver, D.; Schweizer, G.; Tanaka, S.; Liang, L.; Tollot, M.; Zuccaro, A.; Reissmann, S.; Kahmann, R. Fungal Effectors and Plant Susceptibility. Annu. Rev. Plant Biol. 2015, 66, 513–545. [Google Scholar] [CrossRef]
- Rocafort, M.; Fudal, I.; Mesarich, C.H. Apoplastic Effector Proteins of Plant-Associated Fungi and Oomycetes. Curr. Opin. Plant Biol. 2020, 56, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, M.; Ortiz, D.; Henningsen, E.C. Tactics of Host Manipulation by Intracellular Effectors from Plant Pathogenic Fungi. Curr. Opin. Plant Biol. 2021, 62, 102054. [Google Scholar] [CrossRef]
- Schulze-Lefert, P.; Panstruga, R. A Molecular Evolutionary Concept Connecting Nonhost Resistance, Pathogen Host Range, and Pathogen Speciation. Trends Plant Sci. 2011, 16, 117–125. [Google Scholar] [CrossRef]
- Bourras, S.; Kunz, L.; Xue, M.; Praz, C.R.; Müller, M.C.; Kälin, C.; Schläfli, M.; Ackermann, P.; Flückiger, S.; Parlange, F.; et al. The AvrPm3-Pm3 Effector-NLR Interactions Control Both Race-Specific Resistance and Host-Specificity of Cereal Mildews on Wheat. Nat. Commun. 2019, 10, 2292. [Google Scholar] [CrossRef]
- Irieda, H.; Inoue, Y.; Mori, M.; Yamada, K.; Oshikawa, Y.; Saitoh, H.; Uemura, A.; Terauchi, R.; Kitakura, S.; Kosaka, A.; et al. Conserved Fungal Effector Suppresses PAMP-Triggered Immunity by Targeting Plant Immune Kinases. Proc. Natl. Acad. Sci. USA 2019, 116, 496–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, C.V.; Bhaskar, C.K.; Ma, L.-S. A Novel Core Effector Vp1 Promotes Fungal Colonization and Virulence of Ustilago maydis. J. Fungi 2021, 7, 589. [Google Scholar] [CrossRef] [PubMed]
- Snelders, N.C.; Kettles, G.J.; Rudd, J.J.; Thomma, B.P.H.J. Plant Pathogen Effector Proteins as Manipulators of Host Microbiomes? Mol. Plant Pathol. 2018, 19, 257–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wit, P.J.G.M.; Mehrabi, R.; Van den Burg, H.A.; Stergiopoulos, I. Fungal Effector Proteins: Past, Present and Future. Mol. Plant Pathol. 2009, 10, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Tariqjaveed, M.; Mateen, A.; Wang, S.; Qiu, S.; Zheng, X.; Zhang, J.; Bhadauria, V.; Sun, W. Versatile Effectors of Phytopathogenic Fungi Target Host Immunity. J. Integr. Plant Biol. 2021, 63, 1856–1873. [Google Scholar] [CrossRef]
- He, Q.; McLellan, H.; Boevink, P.C.; Birch, P.R.J. All Roads Lead to Susceptibility: The Many Modes of Action of Fungal and Oomycete Intracellular Effectors. Plant Commun. 2020, 1, 100050. [Google Scholar] [CrossRef]
- Flor, H.H. Inheritance of Pathogenicity in Melampsora Lini. Phytopathology 1942, 32, 653–669. [Google Scholar]
- De Wit, P.J.G.M. Apoplastic Fungal Effectors in Historic Perspective; a Personal View. New Phytol. 2016, 212, 805–813. [Google Scholar] [CrossRef] [Green Version]
- Van Schie, C.C.N.; Takken, F.L.W. Susceptibility Genes 101: How to Be a Good Host. Annu. Rev. Phytopathol. 2014, 52, 551–581. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323. [Google Scholar] [CrossRef] [Green Version]
- Kapos, P.; Devendrakumar, K.T.; Li, X. Plant NLRs: From Discovery to Application. Plant Sci. 2019, 279, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Keller, H.; Boyer, L.; Abad, P. Disease Susceptibility in the Zig-Zag Model of Host–Microbe Interactions: Only a Consequence of Immune Suppression? Mol. Plant Pathol. 2016, 17, 475–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thordal-Christensen, H. A Holistic View on Plant Effector-Triggered Immunity Presented as an Iceberg Model. Cell. Mol. Life Sci. 2020, 77, 3963–3976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooney, H.C.E.; van’t Klooster, J.W.; van der Hoorn, R.A.L.; Joosten, M.H.A.J.; Jones, J.D.G.; de Wit, P.J.G.M. Cladosporium Avr2 Inhibits Tomato Rcr3 Protease Required for Cf-2-Dependent Disease Resistance. Science 2005, 308, 1783–1786. [Google Scholar] [CrossRef] [Green Version]
- Van Esse, H.P.; Bolton, M.D.; Stergiopoulos, I.; de Wit, P.J.G.M.; Thomma, B.P.H.J. The Chitin-Binding Cladosporium fulvum Effector Protein Avr4 Is a Virulence Factor. Mol. Plant. Microbe Interact. 2007, 20, 1092–1101. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, K.; Fujisaki, K.; Shimizu, M.; Takeda, T.; Saitoh, H.; Hirabuchi, A.; Hiraka, Y.; Białas, A.; Langner, T.; Kellner, R.; et al. The Blast Pathogen Effector AVR-Pik Binds and Stabilizes Rice Heavy Metal-Associated (HMA) Proteins to Co-Opt Their Function in Immunity. bioRxiv 2020. [Google Scholar] [CrossRef]
- Plissonneau, C.; Blaise, F.; Ollivier, B.; Leflon, M.; Carpezat, J.; Rouxel, T.; Balesdent, M.-H. Unusual Evolutionary Mechanisms to Escape Effector-Triggered Immunity in the Fungal Phytopathogen Leptosphaeria maculans. Mol. Ecol. 2017, 26, 2183–2198. [Google Scholar] [CrossRef]
- Dangl, J.L.; Jones, J.D. Plant Pathogens and Integrated Defence Responses to Infection. Nature 2001, 411, 826–833. [Google Scholar] [CrossRef]
- Van der Hoorn, R.A.L.; Kamoun, S. From Guard to Decoy: A New Model for Perception of Plant Pathogen Effectors. Plant Cell 2008, 20, 2009–2017. [Google Scholar] [CrossRef] [Green Version]
- Cesari, S.; Bernoux, M.; Moncuquet, P.; Kroj, T.; Dodds, P.N. A Novel Conserved Mechanism for Plant NLR Protein Pairs: The ‘Integrated Decoy’ Hypothesis. Front. Plant Sci. 2014, 5, 00606. [Google Scholar] [CrossRef] [Green Version]
- Kroj, T.; Chanclud, E.; Michel-Romiti, C.; Grand, X.; Morel, J.-B. Integration of Decoy Domains Derived from Protein Targets of Pathogen Effectors into Plant Immune Receptors Is Widespread. New Phytol. 2016, 210, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Jubic, L.M.; Saile, S.; Furzer, O.J.; El Kasmi, F.; Dangl, J.L. Help Wanted: Helper NLRs and Plant Immune Responses. Curr. Opin. Plant Biol. 2019, 50, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.-M.; Iswanto, A.B.B.; Son, G.H.; Kim, S.H. Recent Advances in Effector-Triggered Immunity in Plants: New Pieces in the Puzzle Create a Different Paradigm. Int. J. Mol. Sci. 2021, 22, 4709. [Google Scholar] [CrossRef] [PubMed]
- Win, J.; Chaparro-Garcia, A.; Belhaj, K.; Saunders, D.G.O.; Yoshida, K.; Dong, S.; Schornack, S.; Zipfel, C.; Robatzek, S.; Hogenhout, S.A.; et al. Effector Biology of Plant-Associated Organisms: Concepts and Perspectives. Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 235–247. [Google Scholar] [CrossRef]
- Madina, M.H.; Rahman, M.S.; Huang, X.; Zhang, Y.; Zheng, H.; Germain, H. A Poplar Rust Effector Protein Associates with Protein Disulfide Isomerase and Enhances Plant Susceptibility. Biology 2020, 9, 294. [Google Scholar] [CrossRef]
- Cook, D.E.; Mesarich, C.H.; Thomma, B.P.H.J. Understanding Plant Immunity as a Surveillance System to Detect Invasion. Annu. Rev. Phytopathol. 2015, 53, 541–563. [Google Scholar] [CrossRef]
- Tian, H.; Wu, Z.; Chen, S.; Ao, K.; Huang, W.; Yaghmaiean, H.; Sun, T.; Xu, F.; Zhang, Y.; Wang, S.; et al. Activation of TIR Signalling Boosts Pattern-Triggered Immunity. Nature 2021, 598, 500–503. [Google Scholar] [CrossRef]
- Ngou, B.P.M.; Ahn, H.-K.; Ding, P.; Jones, J.D.G. Mutual Potentiation of Plant Immunity by Cell-Surface and Intracellular Receptors. Nature 2021, 592, 110–115. [Google Scholar] [CrossRef]
- Carreón-Anguiano, K.G.; Islas-Flores, I.; Vega-Arreguín, J.; Sáenz-Carbonell, L.; Canto-Canché, B. EffHunter: A Tool for Prediction of Effector Protein Candidates in Fungal Proteomic Databases. Biomolecules 2020, 10, 712. [Google Scholar] [CrossRef]
- Saunders, D.G.O.; Win, J.; Cano, L.M.; Szabo, L.J.; Kamoun, S.; Raffaele, S. Using Hierarchical Clustering of Secreted Protein Families to Classify and Rank Candidate Effectors of Rust Fungi. PLoS ONE 2012, 7, e29847. [Google Scholar] [CrossRef] [PubMed]
- Raffaele, S.; Kamoun, S. Genome Evolution in Filamentous Plant Pathogens: Why Bigger Can Be Better. Nat. Rev. Microbiol. 2012, 10, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.A.B.; Rozano, L.; Debler, J.W.; Mancera, R.L.; Moolhuijzen, P.M.; Hane, J.K. An Automated and Combinative Method for the Predictive Ranking of Candidate Effector Proteins of Fungal Plant Pathogens. Sci. Rep. 2021, 11, 19731. [Google Scholar] [CrossRef] [PubMed]
- Nur, M.; Wood, K.; Michelmore, R. EffectorO: Motif-Independent Prediction of Effectors in Oomycete Genomes Using Machine Learning and Lineage Specificity. BioRxiv 2021. [Google Scholar] [CrossRef]
- Sperschneider, J.; Dodds, P.N. EffectorP 3.0: Prediction of Apoplastic and Cytoplasmic Effectors in Fungi and Oomycetes. Mol. Plant-Microbe Interact. 2021, 35, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 Predicts All Five Types of Signal Peptides Using Protein Language Models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Salvatore, M.; Emanuelsson, O.; Winther, O.; von Heijne, G.; Elofsson, A.; Nielsen, H. Detecting Sequence Signals in Targeting Peptides Using Deep Learning. Life Sci. Alliance 2019, 2, e201900429. [Google Scholar] [CrossRef] [Green Version]
- Sperschneider, J.; Catanzariti, A.-M.; DeBoer, K.; Petre, B.; Gardiner, D.M.; Singh, K.B.; Dodds, P.N.; Taylor, J.M. LOCALIZER: Subcellular Localization Prediction of Both Plant and Effector Proteins in the Plant Cell. Sci. Rep. 2017, 7, 44598. [Google Scholar] [CrossRef] [Green Version]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein Localization Predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [Green Version]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete Genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.; Cuzick, A.; Seager, J.; Wood, V.; Rutherford, K.; Venkatesh, S.Y.; De Silva, N.; Martinez, M.C.; Pedro, H.; Yates, A.D.; et al. PHI-Base: The Pathogen–Host Interactions Database. Nucleic Acids Res. 2020, 48, D613–D620. [Google Scholar] [CrossRef]
- Sperschneider, J.; Gardiner, D.M.; Dodds, P.N.; Tini, F.; Covarelli, L.; Singh, K.B.; Manners, J.M.; Taylor, J.M. EffectorP: Predicting Fungal Effector Proteins from Secretomes Using Machine Learning. New Phytol. 2016, 210, 743–761. [Google Scholar] [CrossRef] [PubMed]
- Sperschneider, J.; Dodds, P.N.; Gardiner, D.M.; Singh, K.B.; Taylor, J.M. Improved Prediction of Fungal Effector Proteins from Secretomes with EffectorP 2.0. Mol. Plant Pathol. 2018, 19, 2094–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dölfors, F.; Holmquist, L.; Dixelius, C.; Tzelepis, G. A LysM Effector Protein from the Basidiomycete Rhizoctonia solani Contributes to Virulence through Suppression of Chitin-Triggered Immunity. Mol. Genet. Genom. 2019, 294, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Feldman, D.; Yarden, O.; Hadar, Y. Seeking the Roles for Fungal Small-Secreted Proteins in Affecting Saprophytic Lifestyles. Front. Microbiol. 2020, 11, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böhnert, H.U.; Fudal, I.; Dioh, W.; Tharreau, D.; Notteghem, J.-L.; Lebrun, M.-H. A Putative Polyketide Synthase/Peptide Synthetase from Magnaporthe grisea Signals Pathogen Attack to Resistant Rice. Plant Cell 2004, 16, 2499–2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Song, T.; Zhang, X.; Yuan, H.; Su, L.; Li, W.; Xu, J.; Liu, S.; Chen, L.; Chen, T.; et al. Unconventionally Secreted Effectors of Two Filamentous Pathogens Target Plant Salicylate Biosynthesis. Nat. Commun. 2014, 5, 4686. [Google Scholar] [CrossRef] [Green Version]
- Gout, L.; Fudal, I.; Kuhn, M.-L.; Blaise, F.; Eckert, M.; Cattolico, L.; Balesdent, M.-H.; Rouxel, T. Lost in the Middle of Nowhere: The AvrLm1 Avirulence Gene of the Dothideomycete Leptosphaeria maculans. Mol. Microbiol. 2006, 60, 67–80. [Google Scholar] [CrossRef]
- Pazzagli, L.; Seidl-Seiboth, V.; Barsottini, M.; Vargas, W.A.; Scala, A.; Mukherjee, P.K. Cerato-Platanins: Elicitors and Effectors. Plant Sci. 2014, 228, 79–87. [Google Scholar] [CrossRef]
- Zhao, S.; Shang, X.; Bi, W.; Yu, X.; Liu, D.; Kang, Z.; Wang, X.; Wang, X. Genome-Wide Identification of Effector Candidates With Conserved Motifs From the Wheat Leaf Rust Fungus Puccinia triticina. Front. Microbiol. 2020, 11, 1188. [Google Scholar] [CrossRef]
- Jones, D.A.B.; Moolhuijzen, P.M.; Hane, J.K. Remote Homology Clustering Identifies Lowly Conserved Families of Effector Proteins in Plant-Pathogenic Fungi. Microb. Genomics 2021, 7, 000637. [Google Scholar] [CrossRef]
- Bertazzoni, S.; Williams, A.H.; Jones, D.A.; Syme, R.A.; Tan, K.-C.; Hane, J.K. Accessories Make the Outfit: Accessory Chromosomes and Other Dispensable DNA Regions in Plant-Pathogenic Fungi. Mol. Plant-Microbe Interact. 2018, 31, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Raffaele, S.; Kamoun, S. The Two-Speed Genomes of Filamentous Pathogens: Waltz with Plants. Curr. Opin. Genet. Dev. 2015, 35, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Raffaele, S.; Win, J.; Cano, L.M.; Kamoun, S. Analyses of Genome Architecture and Gene Expression Reveal Novel Candidate Virulence Factors in the Secretome of Phytophthora infestans. BMC Genom. 2010, 11, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croll, D.; McDonald, B.A. The Accessory Genome as a Cradle for Adaptive Evolution in Pathogens. PLoS Pathog. 2012, 8, e1002608. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Vallet, A.; Fouché, S.; Fudal, I.; Hartmann, F.E.; Soyer, J.L.; Tellier, A.; Croll, D. The Genome Biology of Effector Gene Evolution in Filamentous Plant Pathogens. Annu. Rev. Phytopathol. 2018, 56, 21–40. [Google Scholar] [CrossRef]
- Faino, L.; Seidl, M.F.; Shi-Kunne, X.; Pauper, M.; van den Berg, G.C.M.; Wittenberg, A.H.J.; Thomma, B.P.H.J. Transposons Passively and Actively Contribute to Evolution of the Two-Speed Genome of a Fungal Pathogen. Genome Res. 2016, 26, 1091–1100. [Google Scholar] [CrossRef] [Green Version]
- Rouxel, T.; Balesdent, M.-H. Life, Death and Rebirth of Avirulence Effectors in a Fungal Pathogen of Brassica Crops, Leptosphaeria maculans. New Phytol. 2017, 214, 526–532. [Google Scholar] [CrossRef] [Green Version]
- Plaumann, P.-L.; Schmidpeter, J.; Dahl, M.; Taher, L.; Koch, C. A Dispensable Chromosome Is Required for Virulence in the Hemibiotrophic Plant Pathogen Colletotrichum higginsianum. Front. Microbiol. 2018, 9, 1005. [Google Scholar] [CrossRef] [Green Version]
- Witte, T.E.; Villeneuve, N.; Boddy, C.N.; Overy, D.P. Accessory Chromosome-Acquired Secondary Metabolism in Plant Pathogenic Fungi: The Evolution of Biotrophs Into Host-Specific Pathogens. Front. Microbiol. 2021, 12, 664276. [Google Scholar] [CrossRef]
- Frantzeskakis, L.; Kracher, B.; Kusch, S.; Yoshikawa-Maekawa, M.; Bauer, S.; Pedersen, C.; Spanu, P.D.; Maekawa, T.; Schulze-Lefert, P.; Panstruga, R. Signatures of Host Specialization and a Recent Transposable Element Burst in the Dynamic One-Speed Genome of the Fungal Barley Powdery Mildew Pathogen. BMC Genom. 2018, 19, 381. [Google Scholar] [CrossRef] [Green Version]
- Stam, R.; Münsterkötter, M.; Pophaly, S.D.; Fokkens, L.; Sghyer, H.; Güldener, U.; Hückelhoven, R.; Hess, M. A New Reference Genome Shows the One-Speed Genome Structure of the Barley Pathogen Ramularia collo-cygni. Genome Biol. Evol. 2018, 10, 3243–3249. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Stam, R.; Cano, L.M.; Song, J.; Sklenar, J.; Yoshida, K.; Bozkurt, T.O.; Oliva, R.; Liu, Z.; Tian, M.; et al. Effector Specialization in a Lineage of the Irish Potato Famine Pathogen. Science 2014, 343, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Chavarro-Carrero, E.A.; Vermeulen, J.P.; Torres, D.E.; Usami, T.; Schouten, H.J.; Bai, Y.; Seidl, M.F.; Thomma, B.P.H.J. Comparative Genomics Reveals the in Planta-Secreted Verticillium dahliae Av2 Effector Protein Recognized in Tomato Plants That Carry the V2 Resistance Locus. Environ. Microbiol. 2021, 23, 1941–1958. [Google Scholar] [CrossRef] [PubMed]
- Chepsergon, J.; Motaung, T.E.; Moleleki, L.N. “Core” RxLR Effectors in Phytopathogenic Oomycetes: A Promising Way to Breeding for Durable Resistance in Plants? Virulence 2021, 12, 1921–1935. [Google Scholar] [CrossRef]
- Depotter, J.R.L.; Doehlemann, G. Target the Core: Durable Plant Resistance against Filamentous Plant Pathogens through Effector Recognition. Pest Manag. Sci. 2020, 76, 426–431. [Google Scholar] [CrossRef] [Green Version]
- Hemetsberger, C.; Mueller, A.N.; Matei, A.; Herrberger, C.; Hensel, G.; Kumlehn, J.; Mishra, B.; Sharma, R.; Thines, M.; Hückelhoven, R.; et al. The Fungal Core Effector Pep1 Is Conserved across Smuts of Dicots and Monocots. New Phytol. 2015, 206, 1116–1126. [Google Scholar] [CrossRef]
- Seitner, D.; Uhse, S.; Gallei, M.; Djamei, A. The Core Effector Cce1 Is Required for Early Infection of Maize by Ustilago maydis. Mol. Plant Pathol. 2018, 19, 2277–2287. [Google Scholar] [CrossRef] [Green Version]
- Lanver, D.; Tollot, M.; Schweizer, G.; Lo Presti, L.; Reissmann, S.; Ma, L.-S.; Schuster, M.; Tanaka, S.; Liang, L.; Ludwig, N.; et al. Ustilago maydis Effectors and Their Impact on Virulence. Nat. Rev. Microbiol. 2017, 15, 409–421. [Google Scholar] [CrossRef]
- Vargas, W.A.; Sanz-Martín, J.M.; Rech, G.E.; Armijos-Jaramillo, V.D.; Rivera, L.P.; Echeverria, M.M.; Díaz-Mínguez, J.M.; Thon, M.R.; Sukno, S.A. A Fungal Effector With Host Nuclear Localization and DNA-Binding Properties Is Required for Maize Anthracnose Development. Mol. Plant-Microbe Interact. 2016, 29, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Miao, J.; Shen, D.; Dou, D. Proteinaceous Effector Discovery and Characterization in Plant Pathogenic Colletotrichum Fungi. Front. Microbiol. 2022, 13, 1353–1376. [Google Scholar] [CrossRef]
- Wang, M.; Ji, Z.; Yan, H.; Xu, J.; Zhao, X.; Zhou, Z. Effector Sntf2 Interacted with Chloroplast-Related Protein Mdycf39 Promoting the Colonization of Colletotrichum gloeosporioides in Apple Leaf. Int. J. Mol. Sci. 2022, 23, 6379. [Google Scholar] [CrossRef]
- Tsushima, A.; Narusaka, M.; Gan, P.; Kumakura, N.; Hiroyama, R.; Kato, N.; Takahashi, S.; Takano, Y.; Narusaka, Y.; Shirasu, K. The Conserved Colletotrichum Spp. Effector Candidate CEC3 Induces Nuclear Expansion and Cell Death in Plants. Front. Microbiol. 2021, 12, 682155. [Google Scholar] [CrossRef]
- Stergiopoulos, I.; van den Burg, H.A.; Ökmen, B.; Beenen, H.G.; van Liere, S.; Kema, G.H.J.; de Wit, P.J.G.M. Tomato Cf Resistance Proteins Mediate Recognition of Cognate Homologous Effectors from Fungi Pathogenic on Dicots and Monocots. Proc. Natl. Acad. Sci. USA 2010, 107, 7610–7615. [Google Scholar] [CrossRef] [Green Version]
- Mesarich, C.H.; Ökmen, B.; Rovenich, H.; Griffiths, S.A.; Wang, C.; Karimi Jashni, M.; Mihajlovski, A.; Collemare, J.; Hunziker, L.; Deng, C.H.; et al. Specific Hypersensitive Response-Associated Recognition of New Apoplastic Effectors from Cladosporium fulvum in Wild Tomato. Mol. Plant-Microbe Interact. 2018, 31, 145–162. [Google Scholar] [CrossRef] [Green Version]
- Kohler, A.C.; Chen, L.-H.; Hurlburt, N.; Salvucci, A.; Schwessinger, B.; Fisher, A.J.; Stergiopoulos, I. Structural Analysis of an Avr4 Effector Ortholog Offers Insight into Chitin Binding and Recognition by the Cf-4 Receptor. Plant Cell 2016, 28, 1945–1965. [Google Scholar] [CrossRef] [Green Version]
- Santos Rezende, J.; Zivanovic, M.; Costa de Novaes, M.I.; Chen, Z.-Y. The AVR4 Effector Is Involved in Cercosporin Biosynthesis and Likely Affects the Virulence of Cercospora cf. flagellaris on Soybean. Mol. Plant Pathol. 2020, 21, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-H.; Kračun, S.K.; Nissen, K.S.; Mravec, J.; Jørgensen, B.; Labavitch, J.; Stergiopoulos, I. A Diverse Member of the Fungal Avr4 Effector Family Interacts with De-Esterified Pectin in Plant Cell Walls to Disrupt Their Integrity. Sci. Adv. 2021, 7, eabe0809. [Google Scholar] [CrossRef]
- Bolton, M.D.; van Esse, H.P.; Vossen, J.H.; de Jonge, R.; Stergiopoulos, I.; Stulemeijer, I.J.E.; van den Berg, G.C.M.; Borrás-Hidalgo, O.; Dekker, H.L.; de Koster, C.G.; et al. The Novel Cladosporium fulvum Lysin Motif Effector Ecp6 Is a Virulence Factor with Orthologues in Other Fungal Species. Mol. Microbiol. 2008, 69, 119–136. [Google Scholar] [CrossRef]
- De Jonge, R.; van Esse, H.P.; Kombrink, A.; Shinya, T.; Desaki, Y.; Bours, R.; van der Krol, S.; Shibuya, N.; Joosten, M.H.A.J.; Thomma, B.P.H.J. Conserved Fungal LysM Effector Ecp6 Prevents Chitin-Triggered Immunity in Plants. Science 2010, 329, 953–955. [Google Scholar] [CrossRef]
- Spanu, P.D. Cereal Immunity against Powdery Mildews Targets RNase-Like Proteins Associated with Haustoria (RALPH) Effectors Evolved from a Common Ancestral Gene. New Phytol. 2017, 213, 969–971. [Google Scholar] [CrossRef] [Green Version]
- De Guillen, K.; Ortiz-Vallejo, D.; Gracy, J.; Fournier, E.; Kroj, T.; Padilla, A. Structure Analysis Uncovers a Highly Diverse but Structurally Conserved Effector Family in Phytopathogenic Fungi. PLoS Pathog. 2015, 11, e1005228. [Google Scholar] [CrossRef]
- Lazar, N.; Mesarich, C.H.; Petit-Houdenot, Y.; Talbi, N.; de la Sierra-Gallay, I.L.; Zélie, E.; Blondeau, K.; Gracy, J.; Ollivier, B.; Blaise, F.; et al. A New Family of Structurally Conserved Fungal Effectors Displays Epistatic Interactions with Plant Resistance Proteins. PLoS Pathog. 2022, 18, e1010664. [Google Scholar] [CrossRef]
- Moolhuijzen, P.M.; See, P.T.; Oliver, R.P.; Moffat, C.S. Genomic Distribution of a Novel Pyrenophora tritici-repentis ToxA Insertion Element. PLoS ONE 2018, 13, e0206586. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Garcés, D.; Camborde, L.; Pel, M.J.C.; Jauneau, A.; Martinez, Y.; Néant, I.; Leclerc, C.; Moreau, M.; Dumas, B.; Gaulin, E. CRN13 Candidate Effectors from Plant and Animal Eukaryotic Pathogens Are DNA-Binding Proteins Which Trigger Host DNA Damage Response. New Phytol. 2016, 210, 602–617. [Google Scholar] [CrossRef] [Green Version]
- Frías, M.; Brito, N.; González, M.; González, C. The Phytotoxic Activity of the Cerato-Platanin BcSpl1 Resides in a Two-Peptide Motif on the Protein Surface. Mol. Plant Pathol. 2014, 15, 342–351. [Google Scholar] [CrossRef]
- Frías, M.; González, C.; Brito, N. BcSpl1, a Cerato-Platanin Family Protein, Contributes to Botrytis cinerea Virulence and Elicits the Hypersensitive Response in the Host. New Phytol. 2011, 192, 483–495. [Google Scholar] [CrossRef]
- Schouten, A.; Baarlen, P.V.; Kan, J.A.L.V. Phytotoxic Nep1-like Proteins from the Necrotrophic Fungus Botrytis cinerea Associate with Membranes and the Nucleus of Plant Cells. New Phytol. 2008, 177, 493–505. [Google Scholar] [CrossRef]
- Bae, H.; Kim, M.S.; Sicher, R.C.; Bae, H.-J.; Bailey, B.A. Necrosis- and Ethylene-Inducing Peptide from Fusarium oxysporum Induces a Complex Cascade of Transcripts Associated with Signal Transduction and Cell Death in Arabidopsis. Plant Physiol. 2006, 141, 1056–1067. [Google Scholar] [CrossRef] [Green Version]
- Bailey, B.A.; Jennings, J.C.; Anderson, J.D. The 24-KDa Protein from Fusarium oxysporum f.Sp. erythroxyli: Occurrence in Related Fungi and the Effect of Growth Medium on Its Production. Can. J. Microbiol. 1997, 43, 45–55. [Google Scholar] [CrossRef]
- Kleemann, J.; Rincon-Rivera, L.J.; Takahara, H.; Neumann, U.; Ver Loren van Themaat, E.; van Themaat, E.V.L.; van der Does, H.C.; Hacquard, S.; Stüber, K.; Will, I.; et al. Sequential Delivery of Host-Induced Virulence Effectors by Appressoria and Intracellular Hyphae of the Phytopathogen Colletotrichum higginsianum. PLoS Pathog. 2012, 8, e1002643. [Google Scholar] [CrossRef]
- Joosten, M.H.; Vogelsang, R.; Cozijnsen, T.J.; Verberne, M.C.; De Wit, P.J. The Biotrophic Fungus Cladosporium fulvum Circumvents Cf-4-Mediated Resistance by Producing Unstable AVR4 Elicitors. Plant Cell 1997, 9, 367–379. [Google Scholar] [CrossRef]
- Hemetsberger, C.; Herrberger, C.; Zechmann, B.; Hillmer, M.; Doehlemann, G. The Ustilago maydis Effector Pep1 Suppresses Plant Immunity by Inhibition of Host Peroxidase Activity. PLoS Pathog. 2012, 8, e1002684. [Google Scholar] [CrossRef] [Green Version]
- Saitoh, H.; Fujisawa, S.; Mitsuoka, C.; Ito, A.; Hirabuchi, A.; Ikeda, K.; Irieda, H.; Yoshino, K.; Yoshida, K.; Matsumura, H.; et al. Large-Scale Gene Disruption in Magnaporthe oryzae Identifies MC69, a Secreted Protein Required for Infection by Monocot and Dicot Fungal Pathogens. PLoS Pathog. 2012, 8, e1002711. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.G.; Casady, M.S.; Fee, R.A.; Vaughan, M.M.; Deb, D.; Fedkenheuer, K.; Huffaker, A.; Schmelz, E.A.; Tyler, B.M.; McDowell, J.M. Homologous Rxlr Effectors from Hyaloperonospora arabidopsidis and Phytophthora sojae Suppress Immunity in Distantly Related Plants. Plant J. Cell Mol. Biol. 2012, 72, 882–893. [Google Scholar] [CrossRef]
- Zhang, J.; Shao, F.; Li, Y.; Cui, H.; Chen, L.; Li, H.; Zou, Y.; Long, C.; Lan, L.; Chai, J.; et al. A Pseudomonas syringae Effector Inactivates MAPKs to Suppress PAMP-Induced Immunity in Plants. Cell Host Microbe 2007, 1, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Kelley, B.S.; Lee, S.-J.; Damasceno, C.M.B.; Chakravarthy, S.; Kim, B.-D.; Martin, G.B.; Rose, J.K.C. A Secreted Effector Protein (SNE1) from Phytophthora infestans Is a Broadly Acting Suppressor of Programmed Cell Death. Plant J. Cell Mol. Biol. 2010, 62, 357–366. [Google Scholar] [CrossRef]
- Catanzariti, A.-M.; Dodds, P.N.; Ve, T.; Kobe, B.; Ellis, J.G.; Staskawicz, B.J. The AvrM Effector from Flax Rust Has a Structured C-Terminal Domain and Interacts Directly with the M Resistance Protein. Mol. Plant-Microbe Interact. 2010, 23, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Ve, T.; Williams, S.J.; Catanzariti, A.-M.; Rafiqi, M.; Rahman, M.; Ellis, J.G.; Hardham, A.R.; Jones, D.A.; Anderson, P.A.; Dodds, P.N.; et al. Structures of the Flax-Rust Effector AvrM Reveal Insights into the Molecular Basis of Plant-Cell Entry and Effector-Triggered Immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 17594–17599. [Google Scholar] [CrossRef] [Green Version]
- Cesari, S.; Thilliez, G.; Ribot, C.; Chalvon, V.; Michel, C.; Jauneau, A.; Rivas, S.; Alaux, L.; Kanzaki, H.; Okuyama, Y.; et al. The Rice Resistance Protein Pair RGA4/RGA5 Recognizes the Magnaporthe oryzae Effectors AVR-Pia and AVR1-CO39 by Direct Binding. Plant Cell 2013, 25, 1463–1481. [Google Scholar] [CrossRef] [Green Version]
- Hammond-Kosack, K.E.; Tang, S.; Harrison, K.; Jones, J.D.G. The Tomato Cf-9 Disease Resistance Gene Functions in Tobacco and Potato to Confer Responsiveness to the Fungal Avirulence Gene Product Avr9. Plant Cell 1998, 10, 1251–1266. [Google Scholar] [CrossRef] [Green Version]
- Kanzaki, H.; Yoshida, K.; Saitoh, H.; Fujisaki, K.; Hirabuchi, A.; Alaux, L.; Fournier, E.; Tharreau, D.; Terauchi, R. Arms Race Co-Evolution of Magnaporthe Oryzae AVR-Pik and Rice Pik Genes Driven by Their Physical Interactions. Plant J. 2012, 72, 894–907. [Google Scholar] [CrossRef] [PubMed]
- Mosquera, G.; Giraldo, M.C.; Khang, C.H.; Coughlan, S.; Valent, B. Interaction Transcriptome Analysis Identifies Magnaporthe oryzae BAS1-4 as Biotrophy-Associated Secreted Proteins in Rice Blast Disease. Plant Cell 2009, 21, 1273–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Liu, L.; Wang, Y.; Wang, C.; Yan, J.; Liu, Y.; Wang, C.; Li, C. Overexpression of BAS1 in Rice Blast Fungus Can Promote Blast Fungus Growth, Sporulation and Virulence in Planta. Saudi J. Biol. Sci. 2017, 24, 1884–1893. [Google Scholar] [CrossRef] [PubMed]
- Pennington, H.G.; Jones, R.; Kwon, S.; Bonciani, G.; Thieron, H.; Chandler, T.; Luong, P.; Morgan, S.N.; Przydacz, M.; Bozkurt, T.; et al. The Fungal Ribonuclease-like Effector Protein CSEP0064/BEC1054 Represses Plant Immunity and Interferes with Degradation of Host Ribosomal RNA. PLoS Pathog. 2019, 15, 1007620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgos-Canul, Y.Y.; Canto-Canché, B.; Berezovski, M.V.; Mironov, G.; Loyola-Vargas, V.M.; Barba de Rosa, A.P.; Tzec-Simá, M.; Brito-Argáez, L.; Carrillo-Pech, M.; Grijalva-Arango, R.; et al. The Cell Wall Proteome from Two Strains of Pseudocercospora fijiensis with Differences in Virulence. World J. Microbiol. Biotechnol. 2019, 35, 105. [Google Scholar] [CrossRef]
- Sucheta Tripathy, S.B. A Tale of Effectors; Their Secretory Mechanisms and Computational Discovery in Pathogenic, Non-Pathogenic and Commensal Microbes. Mol. Biol. 2014, 03, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Laur, J.; Ramakrishnan, G.B.; Labbé, C.; Lefebvre, F.; Spanu, P.D.; Bélanger, R.R. Effectors Involved in Fungal–Fungal Interaction Lead to a Rare Phenomenon of Hyperbiotrophy in the Tritrophic System Biocontrol Agent–Powdery Mildew–Plant. New Phytol. 2018, 217, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating Signal Peptides from Transmembrane Regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Rabouille, C. Pathways of Unconventional Protein Secretion. Trends Cell Biol. 2017, 27, 230–240. [Google Scholar] [CrossRef]
- Boevink, P.C.; Wang, X.; McLellan, H.; He, Q.; Naqvi, S.; Armstrong, M.R.; Zhang, W.; Hein, I.; Gilroy, E.M.; Tian, Z.; et al. A Phytophthora infestans RXLR Effector Targets Plant PP1c Isoforms That Promote Late Blight Disease. Nat. Commun. 2016, 7, 10311. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Xu, J.-R. Effectors and Effector Delivery in Magnaporthe oryzae. PLoS Pathog. 2014, 10, e1003826. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, M.C.; Dagdas, Y.F.; Gupta, Y.K.; Mentlak, T.A.; Yi, M.; Martinez-Rocha, A.L.; Saitoh, H.; Terauchi, R.; Talbot, N.J.; Valent, B. Two Distinct Secretion Systems Facilitate Tissue Invasion by the Rice Blast Fungus Magnaporthe oryzae. Nat. Commun. 2013, 4, 1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kale, S.D.; Gu, B.; Capelluto, D.G.S.; Dou, D.; Feldman, E.; Rumore, A.; Arredondo, F.D.; Hanlon, R.; Fudal, I.; Rouxel, T.; et al. External Lipid PI3P Mediates Entry of Eukaryotic Pathogen Effectors into Plant and Animal Host Cells. Cell 2010, 142, 284–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonah, H.; Deshmukh, R.K.; Bélanger, R.R. Computational Prediction of Effector Proteins in Fungi: Opportunities and Challenges. Front. Plant Sci. 2016, 7, 00126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petre, B.; Lorrain, C.; Saunders, D.G.O.; Win, J.; Sklenar, J.; Duplessis, S.; Kamoun, S. Rust Fungal Effectors Mimic Host Transit Peptides to Translocate into Chloroplasts. Cell. Microbiol. 2016, 18, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Robin, G.P.; Kleemann, J.; Neumann, U.; Cabre, L.; Dallery, J.-F.; Lapalu, N.; O’Connell, R.J. Subcellular Localization Screening of Colletotrichum higginsianum Effector Candidates Identifies Fungal Proteins Targeted to Plant Peroxisomes, Golgi Bodies, and Microtubules. Front. Plant Sci. 2018, 9, 00562. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Lan, X.; Song, S.; Yin, L.; Dry, I.B.; Qu, J.; Xiang, J.; Lu, J. In Planta Functional Analysis and Subcellular Localization of the Oomycete Pathogen Plasmopara viticola Candidate RXLR Effector Repertoire. Front. Plant Sci. 2018, 9, 00286. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.; Seto, D.; Subramaniam, R.; Desveaux, D. Oh, the Places They’ll Go! A Survey of Phytopathogen Effectors and Their Host Targets. Plant J. 2018, 93, 651–663. [Google Scholar] [CrossRef]
- Scheibner, F.; Marillonnet, S.; Büttner, D. The Tal Effector Avrbs3 from Xanthomonas campestris pv. vesicatoria Contains Multiple Export Signals and Can Enter Plant Cells in the Absence of the Type III Secretion Translocon. Front. Microbiol. 2017, 8, 02180. [Google Scholar] [CrossRef]
- Rodríguez-Puerto, C.; Chakraborty, R.; Singh, R.; Rocha-Loyola, P.; Rojas, C.M. The Pseudomonas syringae Type III Effector HopG1 Triggers Necrotic Cell Death That Is Attenuated by AtNHR2B. Sci. Rep. 2022, 12, 5388. [Google Scholar] [CrossRef]
- Block, A.; Guo, M.; Li, G.; Elowsky, C.; Clemente, T.E.; Alfano, J.R. The Pseudomonas syringae Type III Effector HopG1 Targets Mitochondria, Alters Plant Development, and Suppresses Plant Innate Immunity. Cell. Microbiol. 2010, 12, 318–330. [Google Scholar] [CrossRef] [PubMed]
- Halane, M.K.; Kim, S.H.; Spears, B.J.; Garner, C.M.; Rogan, C.J.; Okafor, E.C.; Su, J.; Bhattacharjee, S.; Gassmann, W. The Bacterial Type III-Secreted Protein AvrRps4 Is a Bipartite Effector. PLoS Pathog. 2018, 14, e1006984. [Google Scholar] [CrossRef] [PubMed]
- Block, A.; Toruño, T.Y.; Elowsky, C.G.; Zhang, C.; Steinbrenner, J.; Beynon, J.; Alfano, J.R. The Pseudomonas syringae Type III Effector HopD1 Suppresses Effector-Triggered Immunity, Localizes to the Endoplasmic Reticulum, and Targets the Arabidopsis Transcription Factor NTL9. New Phytol. 2014, 201, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, A.; Rutter, W.; Wang, S.; Akhunova, A.; Bolus, S.; Chao, S.; Anderson, N.; De Soto, M.F.; Rouse, M.; Szabo, L.; et al. Variation in the AvrSr35 Gene Determines Sr35 Resistance against Wheat Stem Rust Race Ug99. Science 2017, 358, 1604–1606. [Google Scholar] [CrossRef] [Green Version]
- Manning, V.A.; Hardison, L.K.; Ciuffetti, L.M. Ptr ToxA Interacts with a Chloroplast-Localized Protein. Mol. Plant-Microbe Interact. 2007, 20, 168–177. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ni, H.; Du, X.; Wang, S.; Ma, X.-W.; Nürnberger, T.; Guo, H.-S.; Hua, C. The Verticillium-Specific Protein VdSCP7 Localizes to the Plant Nucleus and Modulates Immunity to Fungal Infections. New Phytol. 2017, 215, 368–381. [Google Scholar] [CrossRef] [Green Version]
- Ning, N.; Xie, X.; Yu, H.; Mei, J.; Li, Q.; Zuo, S.; Wu, H.; Liu, W.; Li, Z. Plant Peroxisome-Targeting Effector MoPtep1 Is Required for the Virulence of Magnaporthe oryzae. Int. J. Mol. Sci. 2022, 23, 2515. [Google Scholar] [CrossRef]
- Liu, T.; Ye, W.; Ru, Y.; Yang, X.; Gu, B.; Tao, K.; Lu, S.; Dong, S.; Zheng, X.; Shan, W.; et al. Two Host Cytoplasmic Effectors Are Required for Pathogenesis of Phytophthora sojae by Suppression of Host Defenses. Plant Physiol. 2011, 155, 490–501. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhang, M.; Shen, D.; Liu, T.; Chen, Y.; Zhou, J.-M.; Dou, D. A Phytophthora sojae Effector PsCRN63 Forms Homo-/Hetero-Dimers to Suppress Plant Immunity via an Inverted Association Manner. Sci. Rep. 2016, 6, 26951. [Google Scholar] [CrossRef] [Green Version]
- Jing, M.; Guo, B.; Li, H.; Yang, B.; Wang, H.; Kong, G.; Zhao, Y.; Xu, H.; Wang, Y.; Ye, W.; et al. A Phytophthora sojae Effector Suppresses Endoplasmic Reticulum Stress-Mediated Immunity by Stabilizing Plant Binding Immunoglobulin Proteins. Nat. Commun. 2016, 7, 11685. [Google Scholar] [CrossRef] [Green Version]
- Caillaud, M.-C.; Piquerez, S.J.M.; Fabro, G.; Steinbrenner, J.; Ishaque, N.; Beynon, J.; Jones, J.D.G. Subcellular Localization of the Hpa RxLR Effector Repertoire Identifies a Tonoplast-Associated Protein HaRxL17 That Confers Enhanced Plant Susceptibility. Plant J. 2012, 69, 252–265. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Zhong, X.; Shi, Y.; Liu, Z.; Jiang, N.; Liu, J.; Ding, B.; Li, Z.; Kang, H.; Ning, Y.; et al. A Fungal Effector Targets a Heat Shock-Dynamin Protein Complex to Modulate Mitochondrial Dynamics and Reduce Plant Immunity. Sci. Adv. 2020, 6, eabb7719. [Google Scholar] [CrossRef] [PubMed]
- Pandey, P.; Leary, A.Y.; Tumtas, Y.; Savage, Z.; Dagvadorj, B.; Duggan, C.; Yuen, E.L.; Sanguankiattichai, N.; Tan, E.; Khandare, V.; et al. An Oomycete Effector Subverts Host Vesicle Trafficking to Channel Starvation-Induced Autophagy to the Pathogen Interface. eLife 2021, 10, e65285. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Yang, J.; Zhang, Q.; Wang, W.; Feng, L.; Zhao, L.; An, B.; Wang, Q.; He, C.; Luo, H. The Effector Protein CgNLP1 of Colletotrichum gloeosporioides Affects Invasion and Disrupts Nuclear Localization of Necrosis-Induced Transcription Factor HbMYB8-Like to Suppress Plant Defense Signaling. Front. Microbiol. 2022, 13, 911479. [Google Scholar] [CrossRef]
- Dagvadorj, B.; Outram, M.A.; Williams, S.J.; Solomon, P.S. The Necrotrophic Effector ToxA from Parastagonospora nodorum Interacts with Wheat NHL Proteins to Facilitate Tsn1-Mediated Necrosis. Plant J. 2022, 110, 407–418. [Google Scholar] [CrossRef]
- Tang, L.; Yang, G.; Ma, M.; Liu, X.; Li, B.; Xie, J.; Fu, Y.; Chen, T.; Yu, Y.; Chen, W.; et al. An Effector of a Necrotrophic Fungal Pathogen Targets the Calcium-Sensing Receptor in Chloroplasts to Inhibit Host Resistance. Mol. Plant Pathol. 2020, 21, 686–701. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Shen, M.; Yang, J.; Xing, Y.; Chen, D.; Li, Z.; Zhao, W.; Zhang, Y. Peroxisomal Fission Is Induced during Appressorium Formation and Is Required for Full Virulence of the Rice Blast Fungus. Mol. Plant Pathol. 2016, 18, 222–237. [Google Scholar] [CrossRef]
- Launay, A.; Jolivet, S.; Clément, G.; Zarattini, M.; Dellero, Y.; Le Hir, R.; Jossier, M.; Hodges, M.; Expert, D.; Fagard, M. DspA/E-Triggered Non-Host Resistance against E. amylovora Depends on the Arabidopsis GLYCOLATE OXIDASE 2 Gene. Int. J. Mol. Sci. 2022, 23, 4224. [Google Scholar] [CrossRef]
- Tzelepis, G.; Dölfors, F.; Holmquist, L.; Dixelius, C. Plant Mitochondria and Chloroplasts Are Targeted by the Rhizoctonia solani RsCRP1 Effector. Biochem. Biophys. Res. Commun. 2021, 544, 86–90. [Google Scholar] [CrossRef]
- Nakano, M.; Mukaihara, T. The Type III Effector RipB from Ralstonia solanacearum RS1000 Acts as a Major Avirulence Factor in Nicotiana benthamiana and Other Nicotiana Species. Mol. Plant Pathol. 2019, 20, 1237–1251. [Google Scholar] [CrossRef] [Green Version]
- Washington, E.J.; Mukhtar, M.S.; Finkel, O.M.; Wan, L.; Banfield, M.J.; Kieber, J.J.; Dangl, J.L. Pseudomonas syringae Type III Effector HopAF1 Suppresses Plant Immunity by Targeting Methionine Recycling to Block Ethylene Induction. Proc. Natl. Acad. Sci. USA 2016, 113, E3577–E3586. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chen, Y.; Wang, J.; Zou, F.; Jia, Y.; Shen, D.; Zhang, Q.; Jing, M.; Dou, D.; Zhang, M. A Phytophthora capsici Virulence Effector Associates with NPR1 and Suppresses Plant Immune Responses. Phytopathol. Res. 2019, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, H.; Jing, M.; Zhu, J.; Guo, B.; Wang, Y.; Lin, Y.; Chen, H.; Kong, L.; Ma, Z.; et al. A Phytophthora Effector Recruits a Host Cytoplasmic Transacetylase into Nuclear Speckles to Enhance Plant Susceptibility. eLife 2018, 7, e40039. [Google Scholar] [CrossRef]
- Ma, L.; Djavaheri, M.; Wang, H.; Larkan, N.J.; Haddadi, P.; Beynon, E.; Gropp, G.; Borhan, M.H. Leptosphaeria maculans Effector Protein AvrLm1 Modulates Plant Immunity by Enhancing MAP Kinase 9 Phosphorylation. iScience 2018, 3, 177–191. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Wang, Y.; Wang, Y.; Du, Y.; Song, C.; Song, P.; Yang, Q.; He, F.; Bai, X.; Huang, L.; et al. Glycine-Serine-Rich Effector PstGSRE4 in Puccinia striiformis f. sp. tritici Inhibits the Activity of Copper Zinc Superoxide Dismutase to Modulate Immunity in Wheat. PLoS Pathog. 2022, 18, e1010702. [Google Scholar] [CrossRef] [PubMed]
- Plett, J.M.; Daguerre, Y.; Wittulsky, S.; Vayssières, A.; Deveau, A.; Melton, S.J.; Kohler, A.; Morrell-Falvey, J.L.; Brun, A.; Veneault-Fourrey, C.; et al. Effector MiSSP7 of the Mutualistic Fungus Laccaria bicolor Stabilizes the Populus JAZ6 Protein and Represses Jasmonic Acid (JA) Responsive Genes. Proc. Natl. Acad. Sci. USA 2014, 111, 8299–8304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blondeau, K.; Blaise, F.; Graille, M.; Kale, S.D.; Linglin, J.; Ollivier, B.; Labarde, A.; Lazar, N.; Daverdin, G.; Balesdent, M.-H.; et al. Crystal Structure of the Effector AvrLm4–7 of Leptosphaeria maculans Reveals Insights into Its Translocation into Plant Cells and Recognition by Resistance Proteins. Plant J. 2015, 83, 610–624. [Google Scholar] [CrossRef]
- Song, J.; Win, J.; Tian, M.; Schornack, S.; Kaschani, F.; Ilyas, M.; van der Hoorn, R.A.L.; Kamoun, S. Apoplastic Effectors Secreted by Two Unrelated Eukaryotic Plant Pathogens Target the Tomato Defense Protease Rcr3. Proc. Natl. Acad. Sci. USA 2009, 106, 1654–1659. [Google Scholar] [CrossRef] [Green Version]
- Mukhtar, M.S.; Carvunis, A.-R.; Dreze, M.; Epple, P.; Steinbrenner, J.; Moore, J.; Tasan, M.; Galli, M.; Hao, T.; Nishimura, M.T.; et al. Independently Evolved Virulence Effectors Converge onto Hubs in a Plant Immune System Network. Science 2011, 333, 596–601. [Google Scholar] [CrossRef] [Green Version]
- Ceulemans, E.; Ibrahim, H.M.M.; De Coninck, B.; Goossens, A. Pathogen Effectors: Exploiting the Promiscuity of Plant Signaling Hubs. Trends Plant Sci. 2021, 26, 780–795. [Google Scholar] [CrossRef]
- Kataria, R.; Kaundal, R. WeCoNET: A Host–Pathogen Interactome Database for Deciphering Crucial Molecular Networks of Wheat-Common Bunt Cross-Talk Mechanisms. Plant Methods 2022, 18, 73. [Google Scholar] [CrossRef] [PubMed]
- Kataria, R.; Kaundal, R. Deciphering the Host–Pathogen Interactome of the Wheat–Common Bunt System: A Step towards Enhanced Resilience in Next Generation Wheat. Int. J. Mol. Sci. 2022, 23, 2589. [Google Scholar] [CrossRef] [PubMed]
- González-Fuente, M.; Carrère, S.; Monachello, D.; Marsella, B.G.; Cazalé, A.-C.; Zischek, C.; Mitra, R.M.; Rezé, N.; Cottret, L.; Mukhtar, M.S.; et al. EffectorK, a Comprehensive Resource to Mine for Ralstonia, Xanthomonas, and Other Published Effector Interactors in the Arabidopsis Proteome. Mol. Plant Pathol. 2020, 21, 1257–1270. [Google Scholar] [CrossRef]
- Crua Asensio, N.; Muñoz Giner, E.; de Groot, N.S.; Torrent Burgas, M. Centrality in the Host-Pathogen Interactome Is Associated with Pathogen Fitness during Infection. Nat. Commun. 2017, 8, 14092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Groot, N.S.; Torrent Burgas, M. Bacteria Use Structural Imperfect Mimicry to Hijack the Host Interactome. PLoS Comput. Biol. 2020, 16, e1008395. [Google Scholar] [CrossRef]
- Mondal, S.I.; Mahmud, Z.; Elahi, M.; Akter, A.; Jewel, N.A.; Muzahidul Islam, M.; Ferdous, S.; Kikuchi, T. Study of Intra-Inter Species Protein-Protein Interactions for Potential Drug Targets Identification and Subsequent Drug Design for Escherichia coli O104:H4 C277-11. Silico Pharmacol. 2016, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Bauters, L.; Stojilković, B.; Gheysen, G. Pathogens Pulling the Strings: Effectors Manipulating Salicylic Acid and Phenylpropanoid Biosynthesis in Plants. Mol. Plant Pathol. 2021, 22, 1436–1448. [Google Scholar] [CrossRef]
- Chen, H.; Chen, J.; Li, M.; Chang, M.; Xu, K.; Shang, Z.; Zhao, Y.; Palmer, I.; Zhang, Y.; McGill, J.; et al. A Bacterial Type III Effector Targets the Master Regulator of Salicylic Acid Signaling, NPR1, to Subvert Plant Immunity. Cell Host Microbe 2017, 22, 777–788.e7. [Google Scholar] [CrossRef] [Green Version]
- Lal, N.K.; Thanasuwat, B.; Huang, P.-J.; Cavanaugh, K.A.; Carter, A.; Michelmore, R.W.; Dinesh-Kumar, S.P. Phytopathogen Effectors Use Multiple Mechanisms to Manipulate Plant Autophagy. Cell Host Microbe 2020, 28, 558–571.e6. [Google Scholar] [CrossRef]
- Kettles, G.J.; Bayon, C.; Sparks, C.A.; Canning, G.; Kanyuka, K.; Rudd, J.J. Characterization of an Antimicrobial and Phytotoxic Ribonuclease Secreted by the Fungal Wheat Pathogen Zymoseptoria tritici. New Phytol. 2018, 217, 320–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, C.; Ramachandran, S.R.; Zhai, Y.; Bu, C.; Pappu, H.R.; Hulbert, S.H. A Novel Fungal Effector from Puccinia graminis Suppressing RNA Silencing and Plant Defense Responses. New Phytol. 2019, 222, 1561–1572. [Google Scholar]
- Cui, C.; Wang, Y.; Liu, J.; Zhao, J.; Sun, P.; Wang, S. A Fungal Pathogen Deploys a Small Silencing RNA That Attenuates Mosquito Immunity and Facilitates Infection. Nat. Commun. 2019, 10, 4298. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Sun, Y.; Song, N.; Zhao, M.; Liu, R.; Feng, H.; Wang, X.; Kang, Z. Puccinia striiformis f. sp. tritici MicroRNA-like RNA 1 (Pst-MilR1), an Important Pathogenicity Factor of Pst, Impairs Wheat Resistance to Pst by Suppressing the Wheat Pathogenesis-Related 2 Gene. New Phytol. 2017, 215, 338–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Weiberg, A.; Lin, F.-M.; Thomma, B.; Huang, H.-D.; Jin, H. Bidirectional Cross-Kingdom RNAi and Fungal Uptake of External RNAs Confer Plant Protection. Nat. Plants 2016, 2, 16151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.; Gupta, V.K.; Jiang, Y.; Yang, B.; Gong, L.; Zhu, H. Cross-Kingdom Small RNAs among Animals, Plants and Microbes. Cells 2019, 8, 371. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhao, Y.-L.; Zhao, J.-H.; Wang, S.; Jin, Y.; Chen, Z.-Q.; Fang, Y.-Y.; Hua, C.-L.; Ding, S.-W.; Guo, H.-S. Cotton Plants Export MicroRNAs to Inhibit Virulence Gene Expression in a Fungal Pathogen. Nat. Plants 2016, 2, 16153. [Google Scholar] [CrossRef]
- Cai, Q.; Qiao, L.; Wang, M.; He, B.; Lin, F.-M.; Palmquist, J.; Huang, S.-D.; Jin, H. Plants Send Small RNAs in Extracellular Vesicles to Fungal Pathogen to Silence Virulence Genes. Science 2018, 360, 1126–1129. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Zhai, Y.; Feng, L.; Karimi, H.Z.; Rutter, B.D.; Zeng, L.; Choi, D.S.; Zhang, B.; Gu, W.; Chen, X.; et al. A Phytophthora Effector Suppresses Trans-Kingdom RNAi to Promote Disease Susceptibility. Cell Host Microbe 2019, 25, 153–165.e5. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cui, C.; Wang, G.; Li, Y.; Wang, S. Insects Defend against Fungal Infection by Employing MicroRNAs to Silence Virulence-Related Genes. Proc. Natl. Acad. Sci. USA 2021, 118, e2023802118. [Google Scholar] [CrossRef]
- Zanini, S.; Šečić, E.; Jelonek, L.; Kogel, K.-H. A Bioinformatics Pipeline for the Analysis and Target Prediction of RNA Effectors in Bidirectional Communication During Plant-Microbe Interactions. Front. Plant Sci. 2018, 9, 1212. [Google Scholar] [CrossRef] [Green Version]
- Derbyshire, M.; Mbengue, M.; Barascud, M.; Navaud, O.; Raffaele, S. Small RNAs from the Plant Pathogenic Fungus Sclerotinia sclerotiorum Highlight Host Candidate Genes Associated with Quantitative Disease Resistance. Mol. Plant Pathol. 2019, 20, 1279–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyman, M.C.; Solofoharivelo, M.-C.; Souza-Richards, R.; Stephan, D.; Murray, S.; Burger, J.T. The Use of High-Throughput Small RNA Sequencing Reveals Differentially Expressed MicroRNAs in Response to Aster Yellows Phytoplasma-Infection in Vitis vinifera cv. ‘Chardonnay’. PLoS ONE 2017, 12, e0182629. [Google Scholar] [CrossRef] [PubMed]
- Dunker, F.; Trutzenberg, A.; Rothenpieler, J.S.; Kuhn, S.; Pröls, R.; Schreiber, T.; Tissier, A.; Kemen, A.; Kemen, E.; Hückelhoven, R.; et al. Oomycete Small RNAs Bind to the Plant RNA-Induced Silencing Complex for Virulence. eLife 2020, 9, e56096. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.; Banerjee, S.; Surana, P.; Liu, M.; Fuerst, G.; Mathioni, S.; Meyers, B.C.; Nettleton, D.; Wise, R.P. Small RNA Discovery in the Interaction between Barley and the Powdery Mildew Pathogen. BMC Genom. 2019, 20, 610. [Google Scholar] [CrossRef] [Green Version]
- Scharf, D.H.; Heinekamp, T.; Brakhage, A.A. Human and Plant Fungal Pathogens: The Role of Secondary Metabolites. PLoS Pathog. 2014, 10, e1003859. [Google Scholar] [CrossRef] [Green Version]
- Devi, R.; Kaur, T.; Guleria, G.; Rana, K.L.; Kour, D.; Yadav, N.; Yadav, A.N.; Saxena, A.K. Chapter 9—Fungal Secondary Metabolites and Their Biotechnological Applications for Human Health. In New and Future Developments in Microbial Biotechnology and Bioengineering; Rastegari, A.A., Yadav, A.N., Yadav, N., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 147–161. ISBN 978-0-12-820528-0. [Google Scholar]
- Boruta, T. Uncovering the Repertoire of Fungal Secondary Metabolites: From Fleming’s Laboratory to the International Space Station. Bioengineered 2017, 9, 12–16. [Google Scholar] [CrossRef] [Green Version]
- Hassani, M.A.; Oppong-Danquah, E.; Feurtey, A.; Tasdemir, D.; Stukenbrock, E.H. Differential Regulation and Production of Secondary Metabolites among Isolates of the Fungal Wheat Pathogen Zymoseptoria tritici. Appl. Environ. Microbiol. 2022, 88, e02296-21. [Google Scholar] [CrossRef]
- Graham-Taylor, C.; Kamphuis, L.G.; Derbyshire, M.C. A Detailed in Silico Analysis of Secondary Metabolite Biosynthesis Clusters in the Genome of the Broad Host Range Plant Pathogenic Fungus Sclerotinia sclerotiorum. BMC Genom. 2020, 21, 7. [Google Scholar] [CrossRef] [Green Version]
- Muria-Gonzalez, M.J.; Chooi, Y.-H.; Breen, S.; Solomon, P.S. The Past, Present and Future of Secondary Metabolite Research in the Dothideomycetes. Mol. Plant Pathol. 2015, 16, 92–107. [Google Scholar] [CrossRef]
- Rangel, L.I.; Bolton, M.D. The Unsung Roles of Microbial Secondary Metabolite Effectors in the Plant Disease Cacophony. Curr. Opin. Plant Biol. 2022, 68, 102233. [Google Scholar] [CrossRef]
- Collemare, J.; Griffiths, S.; Iida, Y.; Karimi Jashni, M.; Battaglia, E.; Cox, R.J.; de Wit, P.J.G.M. Secondary Metabolism and Biotrophic Lifestyle in the Tomato Pathogen Cladosporium fulvum. PLoS ONE 2014, 9, e85877. [Google Scholar] [CrossRef]
- Baker, S.E.; Kroken, S.; Inderbitzin, P.; Asvarak, T.; Li, B.-Y.; Shi, L.; Yoder, O.C.; Turgeon, B.G. Two Polyketide Synthase-Encoding Genes Are Required for Biosynthesis of the Polyketide Virulence Factor, T-Toxin, by Cochliobolus heterostrophus. Mol. Plant-Microbe Interact. 2006, 19, 139–149. [Google Scholar] [CrossRef]
- Möbius, N.; Hertweck, C. Fungal Phytotoxins as Mediators of Virulence. Curr. Opin. Plant Biol. 2009, 12, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Brooks, D.M.; Bender, C.L.; Kunkel, B.N. The Pseudomonas syringae Phytotoxin Coronatine Promotes Virulence by Overcoming Salicylic Acid-Dependent Defences in Arabidopsis thaliana. Mol. Plant Pathol. 2005, 6, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Vergne, E.; Ballini, E.; Marques, S.; Sidi Mammar, B.; Droc, G.; Gaillard, S.; Bourot, S.; DeRose, R.; Tharreau, D.; Nottéghem, J.-L.; et al. Early and Specific Gene Expression Triggered by Rice Resistance Gene Pi33 in Response to Infection by ACE1 Avirulent Blast Fungus. New Phytol. 2007, 174, 159–171. [Google Scholar] [CrossRef]
- Thynne, E.; Mead, O.L.; Chooi, Y.-H.; McDonald, M.C.; Solomon, P.S. Acquisition and Loss of Secondary Metabolites Shaped the Evolutionary Path of Three Emerging Phytopathogens of Wheat. Genome Biol. Evol. 2019, 11, 890–905. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, X.; He, C.; Zhang, Q.-Y.; Zou, X.; Duan, K.; Gao, Q. Novel Fungal Pathogenicity and Leaf Defense Strategies Are Revealed by Simultaneous Transcriptome Analysis of Colletotrichum fructicola and Strawberry Infected by This Fungus. Front. Plant Sci. 2018, 9, 434. [Google Scholar] [CrossRef] [Green Version]
- Chaleckis, R.; Meister, I.; Zhang, P.; Wheelock, C.E. Challenges, Progress and Promises of Metabolite Annotation for LC-MS-Based Metabolomics. Curr. Opin. Biotechnol. 2019, 55, 44–50. [Google Scholar] [CrossRef]
- Pinu, F.R.; Goldansaz, S.A.; Jaine, J. Translational Metabolomics: Current Challenges and Future Opportunities. Metabolites 2019, 9, 108. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, R.E.; Bhatnagar, D.; Ganley, R.J.; Gillman, C.J.; Monahan, B.J.; Seconi, J.M. Dothistroma pini, a Forest Pathogen, Contains Homologs of Aflatoxin Biosynthetic Pathway Genes. Appl. Environ. Microbiol. 2002, 68, 2885–2892. [Google Scholar] [CrossRef] [Green Version]
- Rokas, A.; Mead, M.E.; Steenwyk, J.L.; Raja, H.A.; Oberlies, N.H. Biosynthetic Gene Clusters and the Evolution of Fungal Chemodiversity. Nat. Prod. Rep. 2020, 37, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Schwelm, A.; Bradshaw, R.E. Genetics of Dothistromin Biosynthesis of Dothistroma septosporum: An Update. Toxins 2010, 2, 2680–2698. [Google Scholar] [CrossRef] [PubMed]
- Joosten, M.; de Wit, P. The Tomato-Cladosporium fulvum Interaction: A Versatile Experimental System to Study Plant-Pathogen Interactions. Annu. Rev. Phytopathol. 1999, 37, 335–367. [Google Scholar] [CrossRef] [PubMed]
- De Wit, P.J.G.M. Cladosporium fulvum Effectors: Weapons in the Arms Race with Tomato. Annu. Rev. Phytopathol. 2016, 54, 1–23. [Google Scholar] [CrossRef] [Green Version]
- West, P.V.; Vleeshouwers, V.G. The Phytophthora infestans-Potato Interaction. In Plant Pathogen Interactions; Talbot, N.J., Ed.; Blackwell Scientific Publishers: Hoboken, NJ, USA, 2004. [Google Scholar]
- Yin, J.; Gu, B.; Huang, G.; Tian, Y.; Quan, J.; Lindqvist-Kreuze, H.; Shan, W. Conserved RXLR Effector Genes of Phytophthora infestans Expressed at the Early Stage of Potato Infection Are Suppressive to Host Defense. Front. Plant Sci. 2017, 8, 2155. [Google Scholar] [CrossRef] [Green Version]
- Xin, X.-F.; He, S.Y. Pseudomonas Syringae Pv. Tomato DC3000: A Model Pathogen for Probing Disease Susceptibility and Hormone Signaling in Plants. Annu. Rev. Phytopathol. 2013, 51, 473–498. [Google Scholar] [CrossRef]
- Morris, C.E.; Lamichhane, J.R.; Nikolić, I.; Stanković, S.; Moury, B. The Overlapping Continuum of Host Range among Strains in the Pseudomonas Syringae Complex. Phytopathol. Res. 2019, 1, 4. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Chen, X.; Kemppainen, M.; Pardo, A.G.; Veneault-Fourrey, C.; Kohler, A.; Martin, F.M. The Small Secreted Effector Protein MiSSP7.6 of Laccaria bicolor Is Required for the Establishment of Ectomycorrhizal Symbiosis. Environ. Microbiol. 2020, 22, 1435–1446. [Google Scholar] [CrossRef]
- Constantin, M.E.; Fokkens, L.; de Sain, M.; Takken, F.L.W.; Rep, M. Number of Candidate Effector Genes in Accessory Genomes Differentiates Pathogenic From Endophytic Fusarium oxysporum Strains. Front. Plant Sci. 2021, 12, 761740. [Google Scholar] [CrossRef]
- Collinge, D.B.; Jensen, B.; Jørgensen, H.J. Fungal Endophytes in Plants and Their Relationship to Plant Disease. Curr. Opin. Microbiol. 2022, 69, 102177. [Google Scholar] [CrossRef]
- Rovenich, H.; Boshoven, J.C.; Thomma, B.P.H.J. Filamentous Pathogen Effector Functions: Of Pathogens, Hosts and Microbiomes. Curr. Opin. Plant Biol. 2014, 20, 96–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, C.-M. Against Friend and Foe: Type 6 Effectors in Plant-Associated Bacteria. J. Microbiol. Seoul Korea 2015, 53, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Trunk, K.; Peltier, J.; Liu, Y.-C.; Dill, B.D.; Walker, L.; Gow, N.A.R.; Stark, M.J.R.; Quinn, J.; Strahl, H.; Trost, M.; et al. The Type VI Secretion System Deploys Antifungal Effectors against Microbial Competitors. Nat. Microbiol. 2018, 3, 920–931. [Google Scholar] [CrossRef]
- Hernandez, R.E.; Gallegos-Monterrosa, R.; Coulthurst, S.J. Type VI Secretion System Effector Proteins: Effective Weapons for Bacterial Competitiveness. Cell. Microbiol. 2020, 22, e13241. [Google Scholar] [CrossRef]
- Le, N.-H.; Pinedo, V.; Lopez, J.; Cava, F.; Feldman, M.F. Killing of Gram-Negative and Gram-Positive Bacteria by a Bifunctional Cell Wall-Targeting T6SS Effector. Proc. Natl. Acad. Sci. USA 2021, 118, e2106555118. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Guzmán, P.; Alemán-Duarte, M.I.; Delaye, L.; Herrera-Estrella, A.; Olmedo-Monfil, V. Identification of Effector-like Proteins in Trichoderma Spp. and Role of a Hydrophobin in the Plant-Fungus Interaction and Mycoparasitism. BMC Genet. 2017, 18, 16. [Google Scholar] [CrossRef] [Green Version]
- Horner, N.R.; Grenville-Briggs, L.J.; van West, P. The Oomycete Pythium Oligandrum Expresses Putative Effectors during Mycoparasitism of Phytophthora infestans and Is Amenable to Transformation. Fungal Biol. 2012, 116, 24–41. [Google Scholar] [CrossRef] [Green Version]
- Romero-Contreras, Y.J.; Ramírez-Valdespino, C.A.; Guzmán-Guzmán, P.; Macías-Segoviano, J.I.; Villagómez-Castro, J.C.; Olmedo-Monfil, V. Tal6 From Trichoderma atroviride Is a LysM Effector Involved in Mycoparasitism and Plant Association. Front. Microbiol. 2019, 10, 02231. [Google Scholar] [CrossRef]
- Snelders, N.C.; Rovenich, H.; Petti, G.C.; Rocafort, M.; van den Berg, G.C.M.; Vorholt, J.A.; Mesters, J.R.; Seidl, M.F.; Nijland, R.; Thomma, B.P.H.J. Microbiome Manipulation by a Soil-Borne Fungal Plant Pathogen Using Effector Proteins. Nat. Plants 2020, 6, 1365–1374. [Google Scholar] [CrossRef]
- Snelders, N.C.; Petti, G.C.; van den Berg, G.C.M.; Seidl, M.F.; Thomma, B.P.H.J. An Ancient Antimicrobial Protein Co-Opted by a Fungal Plant Pathogen for in Planta Mycobiome Manipulation. Proc. Natl. Acad. Sci. USA 2021, 118, e2110968118. [Google Scholar] [CrossRef]
- Sarkar, D.; Rovenich, H.; Jeena, G.; Nizam, S.; Tissier, A.; Balcke, G.U.; Mahdi, L.K.; Bonkowski, M.; Langen, G.; Zuccaro, A. The Inconspicuous Gatekeeper: Endophytic Serendipita vermifera Acts as Extended Plant Protection Barrier in the Rhizosphere. New Phytol. 2019, 224, 886–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravanakumar, K.; Fan, L.; Fu, K.; Yu, C.; Wang, M.; Xia, H.; Sun, J.; Li, Y.; Chen, J. Cellulase from Trichoderma harzianum Interacts with Roots and Triggers Induced Systemic Resistance to Foliar Disease in Maize. Sci. Rep. 2016, 6, 35543. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Ji, S.; Wang, Z.; Zhang, H.; Wang, Y.; Liu, Z. Trichoderma asperellum Xylanases Promote Growth and Induce Resistance in Poplar. Microbiol. Res. 2021, 248, 126767. [Google Scholar] [CrossRef] [PubMed]
- Vleeshouwers, V.G.A.A.; Raffaele, S.; Vossen, J.H.; Champouret, N.; Oliva, R.; Segretin, M.E.; Rietman, H.; Cano, L.M.; Lokossou, A.; Kessel, G.; et al. Understanding and Exploiting Late Blight Resistance in the Age of Effectors. Annu. Rev. Phytopathol. 2011, 49, 507–531. [Google Scholar] [CrossRef] [Green Version]
- Bart, R.; Cohn, M.; Kassen, A.; McCallum, E.J.; Shybut, M.; Petriello, A.; Krasileva, K.; Dahlbeck, D.; Medina, C.; Alicai, T.; et al. High-Throughput Genomic Sequencing of Cassava Bacterial Blight Strains Identifies Conserved Effectors to Target for Durable Resistance. Proc. Natl. Acad. Sci. USA 2012, 109, E1972–E1979. [Google Scholar] [CrossRef] [Green Version]
- Giannakopoulou, A.; Steele, J.F.C.; Segretin, M.E.; Bozkurt, T.O.; Zhou, J.; Robatzek, S.; Banfield, M.J.; Pais, M.; Kamoun, S. Tomato I2 Immune Receptor Can Be Engineered to Confer Partial Resistance to the Oomycete Phytophthora infestans in Addition to the Fungus Fusarium oxysporum. Mol. Plant-Microbe Interact. 2015, 28, 1316–1329. [Google Scholar] [CrossRef] [Green Version]
- Van de Wouw, A.P.; Idnurm, A. Biotechnological Potential of Engineering Pathogen Effector Proteins for Use in Plant Disease Management. Biotechnol. Adv. 2019, 37, 107387. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Distribution * | Effector Type | Organism | Effector Name and Uniprot ID | Function | Reference |
|---|---|---|---|---|---|
| Conserved: Different classes in Ascomycetes | Ceratoplatanin family | Botrytis cinerea | BcSpl1 (A0A384JBC5) | Phytotoxin, elicits systemic resistance (SAR) in tobacco. Necrosis (HR) in tobacco, tomato and Arabidopsis | [125,126] |
| Conserved: Wide distribution in Dothideomycetes class (Ascomycetes) | LysM domain | Cladosporium fulvum | Ecp6 (B3VBK9) | Virulence factor; binds to fungal chitin to prevent chitin triggered-immunity in host | [118] |
| Conserved: Colletotrichum genus (Glomerellales order), Fusarium, Trichoderma, (Hypocreales order) in Sordariomycetes class. Few species in Dothideomycetes class (Ascomycetes) | Necrosis inducing protein | Colletotrichum orbiculare | NIS1 (H7CE97) | Suppresses PAMP-triggered immunity, targets kinases BAK1 and BIK1 Recognition is followed by a plant cell death response (necrosis), potential avirulence effects | [41] |
| Conserved: Few genera in Sclerotiniaceae family (Ascomycetes phylum) | Necrosis and ethylene-inducing proteins (NLPs) | Botrytis cinerea | BcNep2 (Q079H3) | Induces cell death in dicotyledonous plants | [127] |
| Conserved: Fusarium genus (Hypocreales order), Bipolaris genus (Pleosporales order), Verticillium genus (Glomerellales order), in Sordariomycetes class (Ascomycetes phylum) | Necrosis and ethylene-inducing proteins (NLPs) | Fusarium oxysporum f. sp. erythroxyli | NEP1 (O42737) | Induces necrosis and ethylene emission in Erythroxylum coca leaves | [128,129] |
| Conserved: Many species in Colletotrichum genus (Glomerellales order) and Fusarium genus (Hypocreales order) | Necrosis inducing protein | Colletotrichum higginsianum | ChNLP1 (K7N7F9) | Induces necrosis in N. benthamiana | [130] |
| Conserved: Many species in Mycosphaerellaceae family, Dothideomycetes class | Chitin-binding type-2 | Cladosporium fulvum | Avr4 (Q00363) | Triggers a Cf-4-mediated hypersensitive response (HR) in tomato; protects fungal cell walls against hydrolysis by plant chitinases | [55,113,131] |
| Conserved: Species in Ustilaginaceae family | Pep1 | Ustilago maydis | Pep1 (G0X7E8) | Inhibitor of plant peroxidases; targets maize peroxidase POX12 to suppress plant immunity | [106,132] |
| Conserved: Few species in Magnaporthales, Sordariales, Xylariales, Hypocreales, Glomerellales orders | MC69 | Magnaporthe oryzae | MC69 (L7JHY1) | Development of invasive hyphae affected in mc69 mutant; reduced pathogenicity on host | [133] |
| Specific to genus | Avr | Phytophthora sojae | PsAvh163 (G1FRR2) | Suppresses PTI and ETI responses in Arabidopsis Activates immunity (HR response) in the Nicotiana genus. | [134] |
| Specific to genus | Hop | Pseudomonas syringae | HopAl1 (Q888W0) | Phosphothreonine lyase; inactivates MAPKs in Arabidopsis to overcome PTI | [135] |
| Specific to genus | Suppressor of necrosis 1 SNE1 | Phytophthora infestans | SNE1 (A2CLL0) | Suppresses cell death induced by NLP and Avr effectors in N. benthamiana and Solanum lycopersicum | [136] |
| Specific to genus | Avr | Melamspora lini | AvrM (Q2MV46) | Induces ETI in host; targets resistance protein M | [137,138] |
| Specific to genus | Avr | Magnaporthe oryzae | AvrPia (B9WZW9) | Induces ETI in host; targets resistance protein RGA5 | [139] |
| Species-specific | Avr | Cladosporium fulvum | Avr9 (P22287) | Induces ETI in host; Targets resistance protein of Cf-9 | [140] |
| Species-specific | Avr | Magnaporthe oryzae | AVR-Pik (C4B8B8) | Induces ETI in host; targets resistance protein Pik | [56,141] |
| Species-specific | Biotrophy-associated secreted (BAS) proteins | Magnaporthe oryzae | BAS1 (G5EHI7) | Triggers defense response in host; overexpression increases virulence, sporulation and reduces expression of host defense-related genes | [142,143] |
| Species-specific | Ribonuclease family | Blumeria graminis | BEC1054 (N1JJ94) | Ribonuclease like; binds to host ribosomes and inhibits the action of plant ribosome-inactivating proteins (RIPs) | [144] |
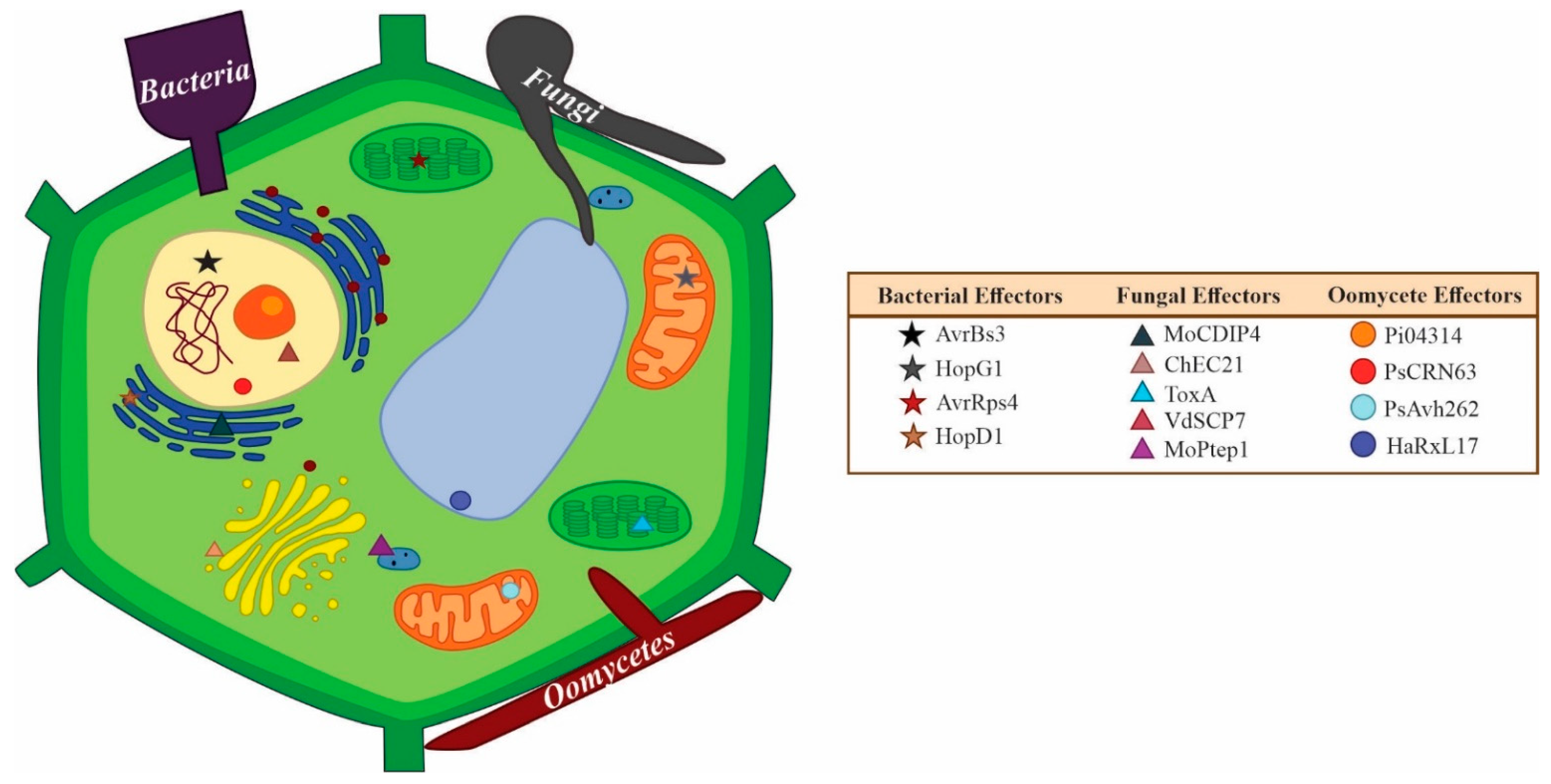
| Effector | Cell Localization | Microorganism | Kingdom | Lifestyle | Reference |
|---|---|---|---|---|---|
| CgEP1 | Nucleus | Colletotrichum graminicola | Fungi | Hemibiotrophic | [109] |
| ChECs (nine) | Nucleus, peroxisomes, and microtubules | Colletotrichum higginsianum | Fungi | Hemibiotrophic | [156] |
| MoCDIP4 | Mitochondria | Magnaporthe oryzae | Fungi | Hemibiotrophic | [172] |
| Rab8a | Golgi apparatus | Phytophthora infestans | Oomycete | Hemibiotrophic | [173] |
| CSEP0064/ BEC1054 | Cytoplasmic | Blumeria graminis | Fungi | Biotrophic | [144] |
| Sntf2 | Chloroplast | Colletotrichum gloeosporioides | Fungi | Hemibiotrophic | [111] |
| CgNLP1 | Nucleus | Colletotrichum gloeosporioides | Fungi | Hemibiotrophic | [174] |
| ToxA | Cytoplasm | Parastagonospora nodorum | Fungi | Necrotrophic | [175] |
| SsITL | Chloroplast | Sclerotinia sclerotiorum | Fungi | Necrotrophic | [176] |
| PEF1 | Peroxisomes | Magnaporthe oryzae | Fungi | Hemibiotrophic | [177] |
| DspA/E | Peroxisomes | Erwinia amylovora | Bacteria | Necrotrophic | [178] |
| RsCRP1 | Mitochondria and chloroplasts | Rhizoctonia solani | Fungi | Necrotrophic | [179] |
| RipAA | Chloroplasts | Ralstonia solanacearum | Bacteria | Necrotrophic | [180] |
| HopAF1 | Plasma membrane and cytoplasm | Pseudomonas syringae | Bacteria | Hemibiotrophic | [181] |
| PcRxLR48 | Nucleus | Phytophthora infestans | Oomycete | Hemibiotrophic | [182] |
| PsAvh52 | Cytoplasm and nucleus | Phytophthora infestans | Oomycete | Hemibiotrophic | [183] |
| AvrLm1 | Plasma membrane | Leptosphaeria maculans | Fungi | Hemibiotrophic | [184] |
| PstGSRE4 | Nucleus | Puccinia striiformis f. sp. tritici | Fungi | Biotrophic | [185] |
| MiSSP7 | Nucleus | Laccaria bicolor | Fungi | Ectomycorrhizal symbiont | [186] |
| RsCRP1 | Mitochondria and chloroplasts | Rhizoctonia solani | Fungi | Necrotrophic | [179] |
| PexRD54 | Golgi apparatus | Phytophthora infestans | Oomycete | Hemibiotrophic | [173] |
| Plants | ||||
|---|---|---|---|---|
| Host Target (Hub) (Targets in Different Hosts Targeted by Multiple Effectors) | Function in the Host | Targeted by (Selected Examples) | Microorganisms | References |
| Chorismate mutase (CM) | Salicylic acid (SA) biosynthesis | Cmu1, Mi-CM-3, AvrPtoB | Ustilago maydis (fungus), Meloidogyne javanica (nematode), Pseudomonas syringae (bacterium) | [197,198] |
| Isochorismatase (ICM) | Salicylic acid (SA) biosynthesis | VdIsc1, PsIsc1, HopI1 | Verticillium dahliae (fungus), Phytophthora sojae (oomycete), Pseudomonas syringae (bacterium) | [197] |
| JAZ6, negative regulator of JA induced transcription | Jasmonic acid (JA) biosynthesis | MiSSP7, HopZ1a, HopX1 | Laccaria bicolor and Golovinomyces orontii (fungi), Hyaloperonospora arabidopsidis (oomycete), Pseudomonas syringae (bacterium) | [197] |
| AuTophaGy (ATG) proteins | Autophagy | HrpZ1, HopF3, AvrPtoB, HrpZ1, HopO1-2, | Bremia lactucae, Hyaloperonospora arabidopsidis (oomycetes), Uromyces fabae (fungus), Globodera pallida (nematode), Pseudomonas syringae (bacterium) | [199] |
| Pathogen | ||||
| Pathogen hub (Multiple targets to one effector) | Functions | Targets in the host | Microorganisms where that effector has been identified | References |
| AvrPtoB | Interferes with salicylic acid (SA) biosynthesis and autophagy in plants | CM, ATG1 | Pseudomonas syringae | [198,199] |
| Zt6 | Phytotoxic and antimicrobial ribonuclease effector | Plant and microbial rRNA species | Zymoseptoria tritici | [200,201] |
| VdAMP2 | Soil colonization (antibacterial activity), and induction of necrosis in Nicotiana benthamiana | Unknown | Verticillium dahliae | [17] |
| VdAMP3 | Soil colonization (antifungal activity) | Unknown, mycobiome manipulation | Verticillium dahliae | [17] |
| VirD5 | Stabilizes VirF for Agrobacterium infection | Arabidopsis thaliana AT1G09270.1 AT1G43700.1 AT3G06720.1 AT5G59710.1 VIP2 | Agrobacterium tumefaciens | [193] |
| Avr2 | Inhibits tomato Rcr3 protease | Arabidopsis thaliana AT1G47128.1 AT3G19400.1 AT3G45310.1 AT4G35350.1 AT5G60360.1 | Cladosporium fulvum | [193] |
| HARXL106 | Suppresses transcriptional activation of salicylic acid (SA)-induced defense genes | Arabidopsis thaliana AT1G32230.1 AT2G35510.1 | Hyaloperonospora arabidopsidis | [193] |
| RXLR24 | Inhibitor of RABA GTPase-mediated vesicular secretion of antimicrobial PR-1 and PDF1.2 | Arabidopsis thaliana AT1G06400.1 AT1G09630.1 AT1G16920.1 AT2G30950.1 AT3G18820.1 AT3G46830.1 AT3G56940.1 AT4G18800.1 AT4G39990.1 AT5G45750.1 AT5G47960.1 AT5G59150.1 AT5G60860.1 AT5G65270.1 | Phytophthora brassicae | [193] |
| PAST | PRESENT |
|---|---|
| Effectors show no or limited sequence homology or conservation but may share structural properties | Some effectors are homologs of effectors in other microorganisms. Some orthologs display a high level of sequence conservation |
| Homologs are distributed in close phylogenetic relatives | Some effectors are distributed in phylogenetically related and distant organisms (core effectors) |
| Effectors are small proteins (<300 or 400 amino acids) | Limit in length of known current effectors is ~ 850 amino acids |
| Effectors are secreted proteins with signal Peptides | Many lack signal peptides and are secreted by little-understood non-conventional processes |
| Effector proteins lack TMDs | Some true effectors have one or two TMDs; the current limit for TMDs in a true effector is 6 |
| The majority of effectors initially discovered were extracellular, apoplastic proteins | Many effectors also target cytoplasmic and organellar host proteins. Some effectors may even target both apoplast and cytoplasm |
| Effector interactions follow the gene-for-gene model | Effector interactions encompass the gene-for-gene model, guard model, the decoy model, integrated decoy model etc. |
| Effectors are proteins (Enzymatic or non-enzymatic) | Effectors can be proteins, RNA or secondary metabolites |
| Effectors induce the hypersensitive response (local, visible lesion) on the host leaves | Not all effectors induce HR. Biotrophic effectors can interfere with downstream reactions triggered by effector-R cognate recognition and prevent development of visible lesions |
| MTI and ETI are independent immunity Pathways | MTI and ETI are interconnected |
| Effectors are produced by pathogens to target plant hosts | Produced by pathogens and non-pathogens with targets in a wide spectrum of organisms |
| Effector roles are associated with conquering the plant host | Effector roles documented in host and niche conquest; roles in shaping microbiomes and roles in reproduction and development of producer organism |
| Each effector has one target or cognate | Some effectors are multifunctional or have many different targets in the same host or different hosts; and vice versa, some host targets are targeted by multiple effectors from the same or different microorganisms |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Todd, J.N.A.; Carreón-Anguiano, K.G.; Islas-Flores, I.; Canto-Canché, B. Fungal Effectoromics: A World in Constant Evolution. Int. J. Mol. Sci. 2022, 23, 13433. https://doi.org/10.3390/ijms232113433
Todd JNA, Carreón-Anguiano KG, Islas-Flores I, Canto-Canché B. Fungal Effectoromics: A World in Constant Evolution. International Journal of Molecular Sciences. 2022; 23(21):13433. https://doi.org/10.3390/ijms232113433
Chicago/Turabian StyleTodd, Jewel Nicole Anna, Karla Gisel Carreón-Anguiano, Ignacio Islas-Flores, and Blondy Canto-Canché. 2022. "Fungal Effectoromics: A World in Constant Evolution" International Journal of Molecular Sciences 23, no. 21: 13433. https://doi.org/10.3390/ijms232113433
APA StyleTodd, J. N. A., Carreón-Anguiano, K. G., Islas-Flores, I., & Canto-Canché, B. (2022). Fungal Effectoromics: A World in Constant Evolution. International Journal of Molecular Sciences, 23(21), 13433. https://doi.org/10.3390/ijms232113433