
Antipsychotic Drug-Mediated Adverse Effects on Rat Testicles May Be Caused by Altered Redox and Hormonal Homeostasis

 ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
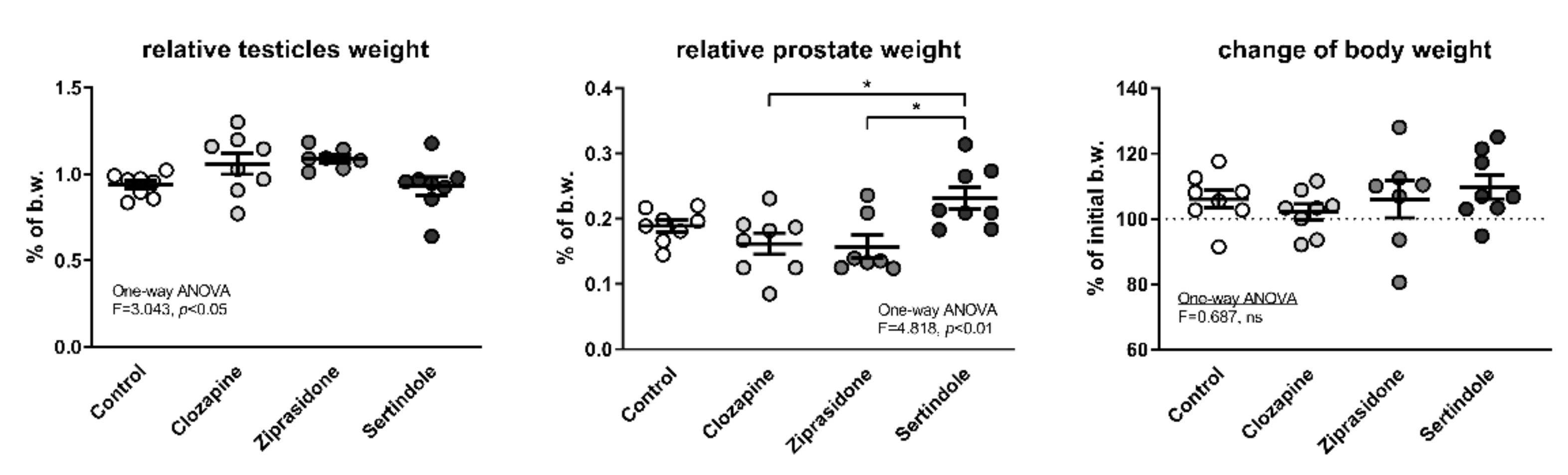
2.1. Relative Testicle and Prostate Weights
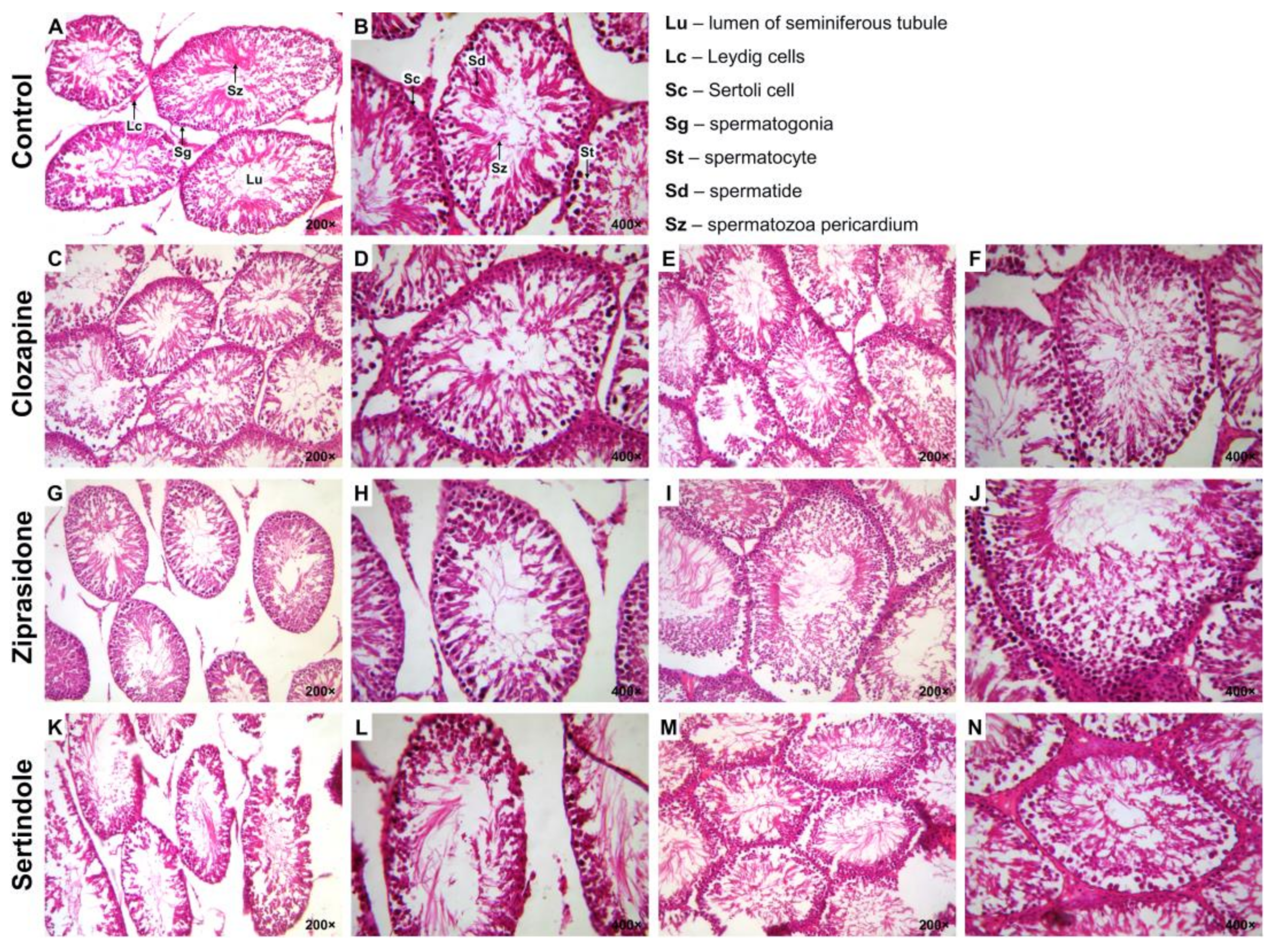
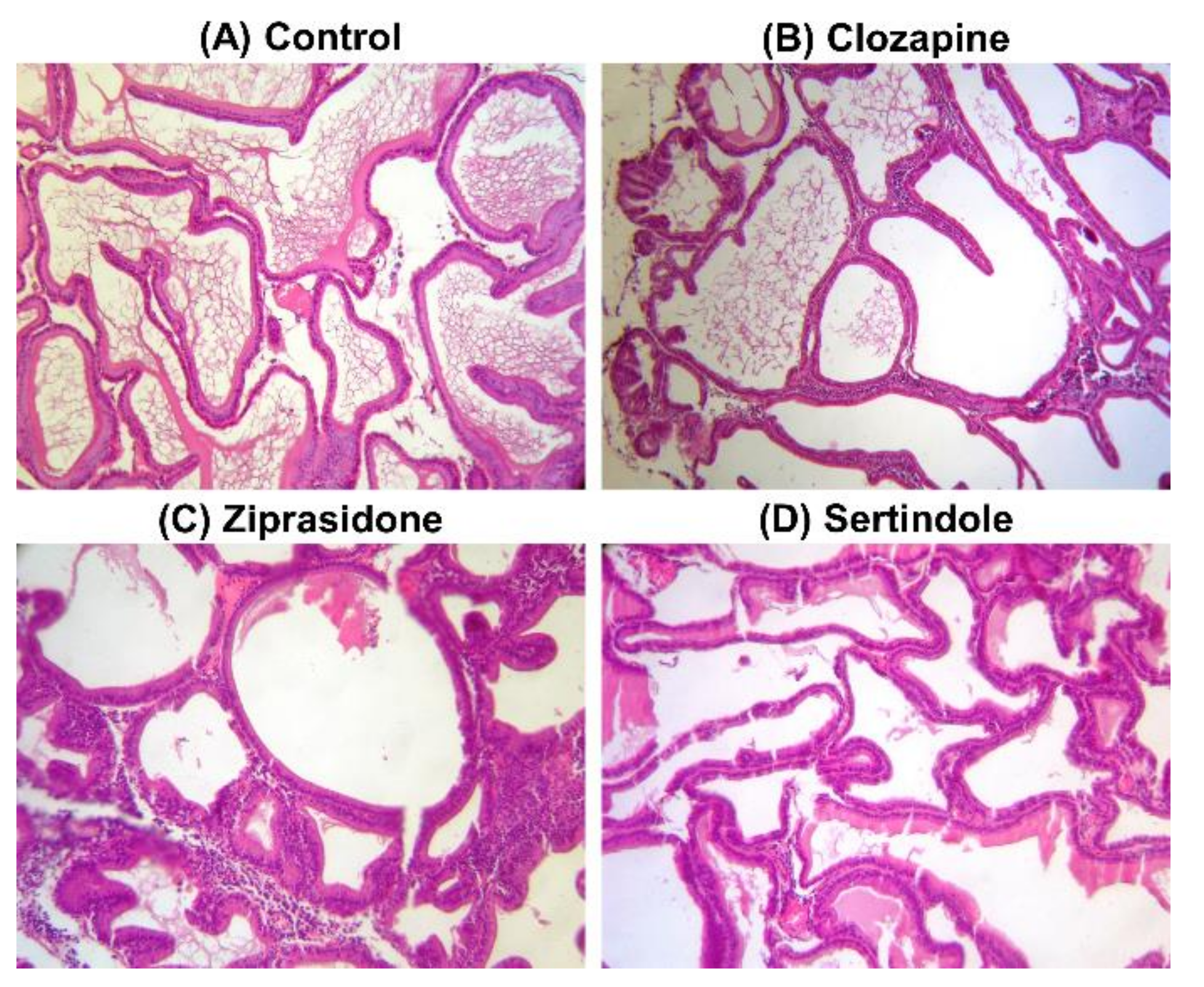
2.2. Histopathological Examination of the Testicular and Prostate Tissue
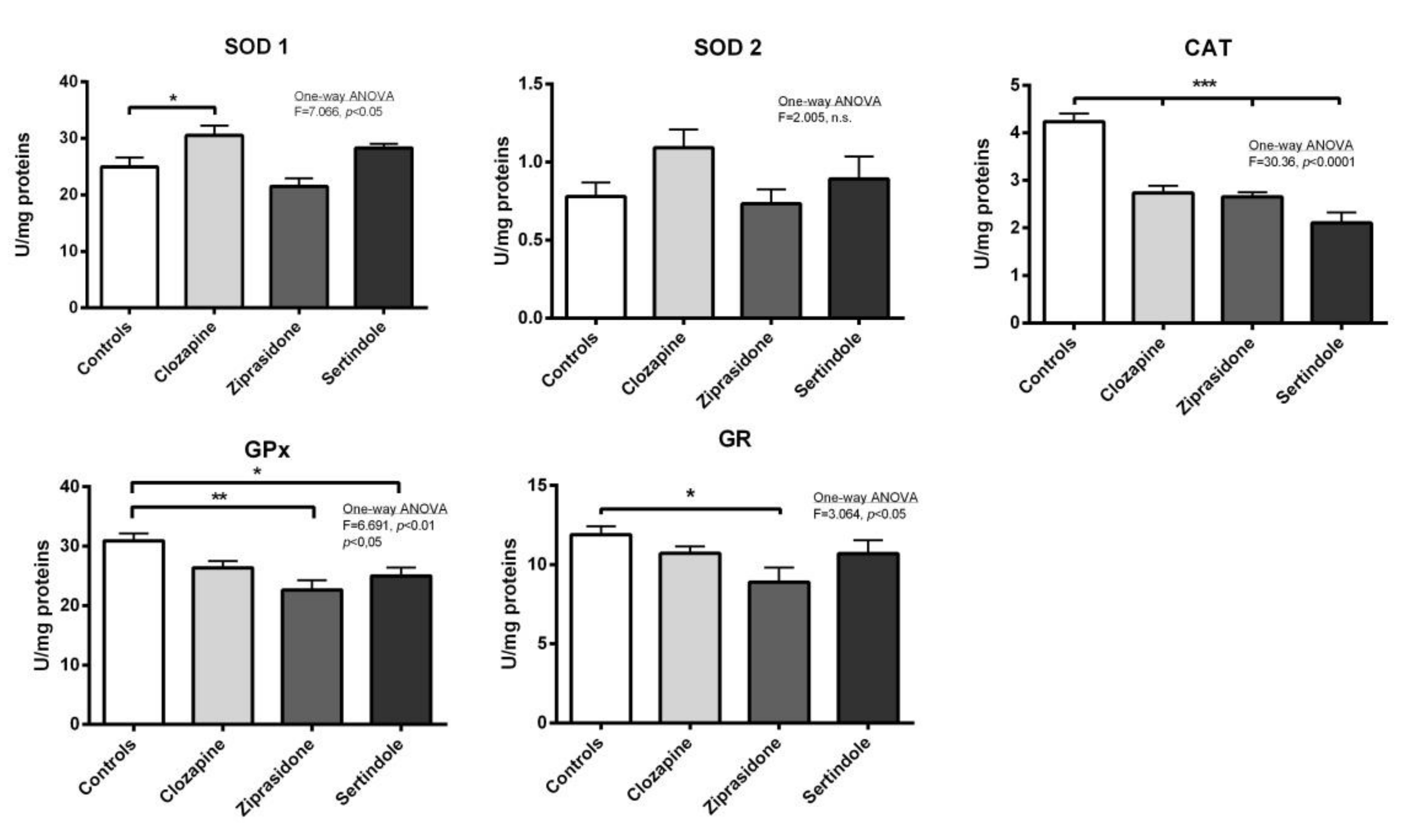
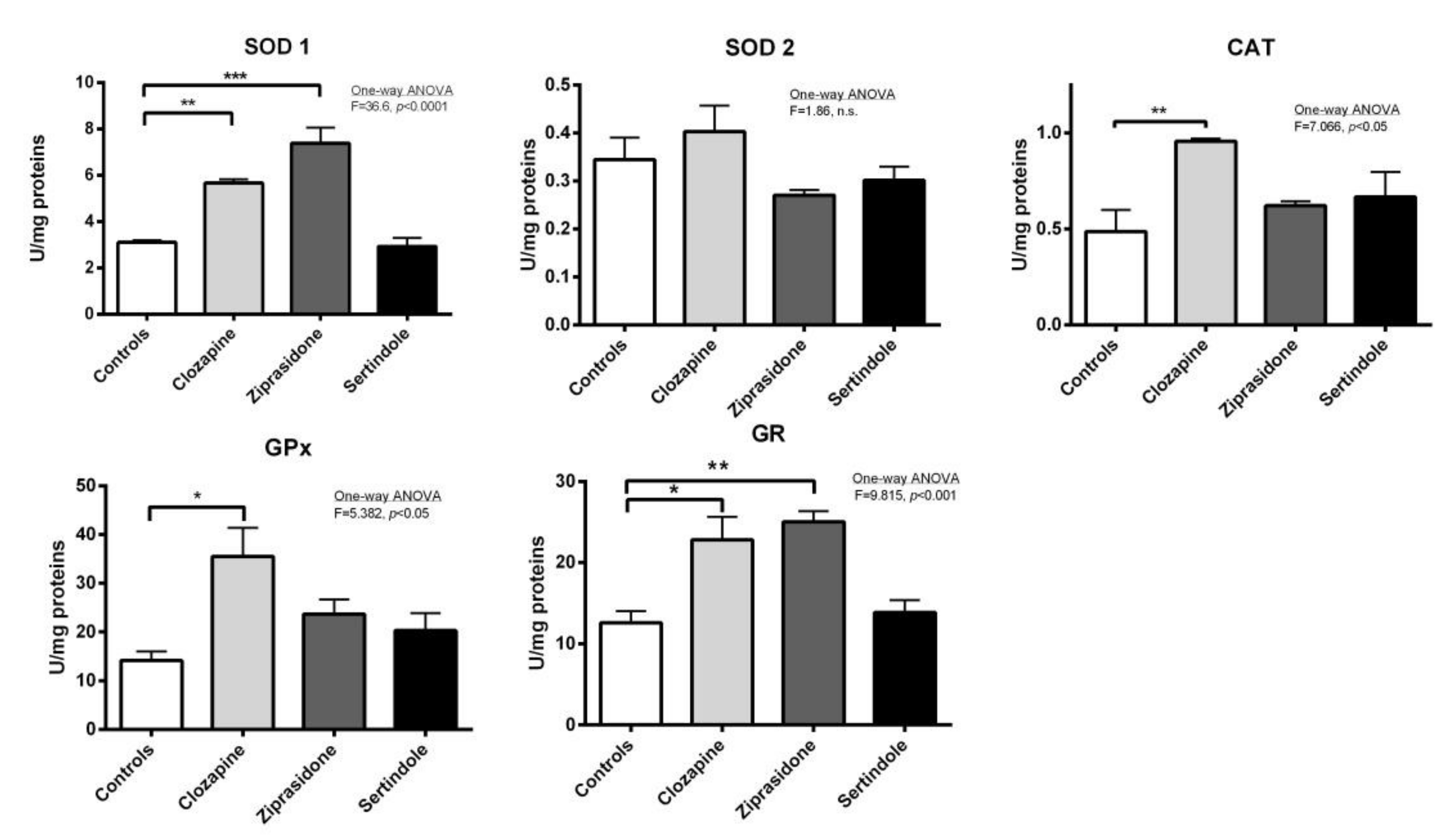
2.3. Activity of Antioxidant Enzymes in Testicles and Prostate
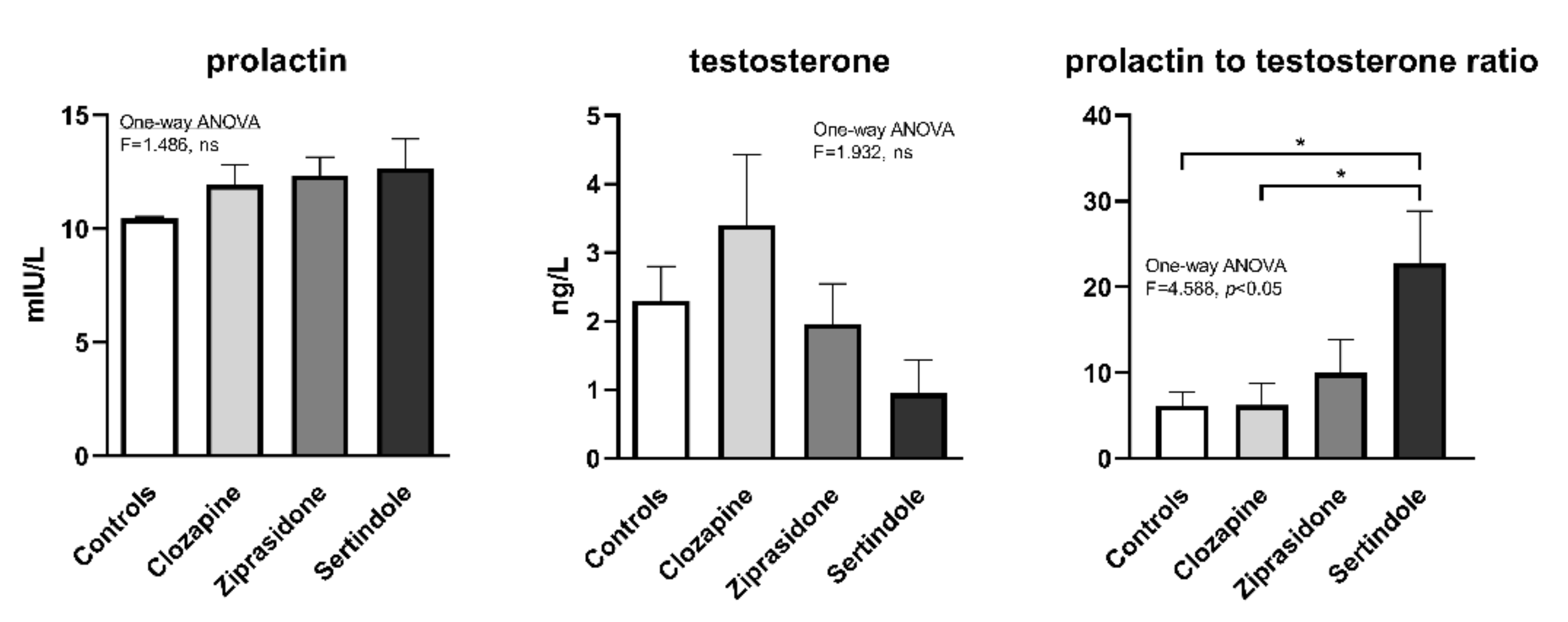
2.4. Hormone Levels in Plasma
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animals and Drug Treatment
4.3. Blood and Tissue Collection
4.4. Measurement of Prolactin, Testosterone, ALT and AST
4.5. Tissue Preparation and Determination of Antioxidant Enzyme Activities
4.6. Histopathological Examination
4.7. Histopathological Examination of Spermatogenesis (Visualisation of the Spermatogenic Cells)
4.8. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mauri, M.C.; Paletta, S.; Maffini, M.; Colasanti, A.; Dragogna, F.; Di Pace, C.; Altamura, A.C. Clinical pharmacology of atypical antipsychotics: An update. EXCLI J. 2014, 13, 1163–1191. [Google Scholar] [PubMed]
- Haupt, D.W. Differential metabolic effects of antipsychotic treatments. Eur. Neuro-Psychopharmacol. 2006, 216, S149–S155. [Google Scholar] [CrossRef] [PubMed]
- Glassman, A.H.; Bigger, J.T., Jr. Antipsychotic drugs: Prolonged QTc interval, torsade de pointes, and sudden death. Am. J. Psychiatry 2001, 158, 1774–17782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, Y.J.; Dixon, S.N.; Reiss, J.P.; Wald, R.; Parikh, C.R.; Gandhi, S.; Shariff, S.Z.; Pannu, N.; Nash, D.M.; Rehman, F.; et al. Atypical antipsychotic drugs and the risk for acute kidney injury and other adverse outcomes in older adults: A population-based cohort study. Ann. Intern. Med. 2014, 161, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Slim, M.; Medina-Caliz, I.; Gonzalez-Jimenez, A.; Cabello, M.R.; Mayoral-Cleries, F.; Lucena, M.I.; Andrade, R.J. Hepatic Safety of Atypical Antipsychotics: Current Evidence and Future Directions. Drug Saf. 2016, 39, 925–943. [Google Scholar] [CrossRef] [PubMed]
- Stroup, T.S.; Gray, N. Management of common adverse effects of antipsychotic medications. World Psychiatry 2018, 17, 341–356. [Google Scholar] [CrossRef]
- Montejo, A.L.; de Alarcón, R.; Prieto, N.; Acosta, J.M.; Buch, B.; Montejo, L. Management Strategies for Antipsychotic-Related Sexual Dysfunction: A Clinical Approach. J. Clin. Med. 2021, 10, 308. [Google Scholar] [CrossRef]
- Park, Y.W.; Kim, Y.; Lee, J.H. Antipsychotic-induced sexual dysfunction and its management. World J. Mens. Health. 2012, 30, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Cutler, A.J. Sexual dysfunction and antipsychotic treatment. Psychoneuroendocrinology 2003, 28, 69–82. [Google Scholar] [CrossRef]
- Samadi, A.; Isikhan, S.Y.; Ansari, M.H.K.; Samadi, M.; Sabuncuoglu, S. Effects of clozapine and haloperidol treatment on plasma concentrations of androgen hormones and androgen dependent organ changes in rats. Indian J. Pharmacol. 2019, 51, 269–275. [Google Scholar] [CrossRef]
- De Rosa, M.; Zarrilli, S.; Di Sarno, A.; Milano, N.; Gaccione, M.; Boggia, B.; Lombardi, G.; Colao, A. Hyperprolactinemia in men: Clinical and biochemical features and response to treatment. Endocrine 2003, 20, 75–82. [Google Scholar] [CrossRef]
- Howes, O.D.; Wheeler, M.J.; Pilowsky, L.S.; Landau, S.; Murray, R.M.; Smith, S. Sexual function and gonadal hormones in patients taking antipsychotic treatment for schizophrenia or schizoaffective disorder. J. Clin. Psychiatry 2007, 68, 361–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konarzewska, B.; Wołczyński, S.; Szulc, A.; Galińska, B.; Popławska, R.; Waszkiewicz, N. Effect of risperidone and olanzapine on reproductive hormones, psychopathology and sexual functioning in male patients with schizophrenia. Psychoneuroendocrinology 2009, 34, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Turrone, P.; Kapur, S.; Seeman, M.V.; Flint, A.J. Elevation of prolactin levels by atypical antipsychotics. Am. J. Psychiatry 2002, 159, 133–135. [Google Scholar] [CrossRef]
- Semet, M.; Paci, M.; Saïas-Magnan, J.; Metzler-Guillemain, C.; Boissier, R.; Lejeune, H.; Perrin, J. The impact of drugs on male fertility: A review. Andrology 2017, 5, 640–663. [Google Scholar] [CrossRef] [Green Version]
- Maroney, M. An update on current treatment strategies and emerging agents for the management of schizophrenia. Am. J. Manag. Care 2020, 26 (Suppl. S3), S55–S61. [Google Scholar] [CrossRef]
- Tremellen, K. Oxidative stress and male infertility-a clinical perspective. Hum. Reprod. Update 2008, 14, 243–258. [Google Scholar] [CrossRef]
- Platanić Arizanović, L.; Nikolić-Kokić, A.; Brkljačić, J.; Tatalović, N.; Miler, M.; Oreščanin-Dušić, Z.; Vidonja Uzelac, T.; Nikolić, M.; Milošević, V.; Blagojević, D.; et al. Effects of several atypical antipsychotics closapine, sertindole or ziprasidone on hepatic antioxidant enzymes: Possible role in drug-induced liver dysfunction. J. Toxicol. Environ. Health A 2021, 84, 173–182. [Google Scholar] [CrossRef]
- Nikolić-Kokić, A.; Mijušković, A.; Tatalović, N.; Nestorov, J.; Miler, M.; Oreščanin-Dušić, Z.; Nikolić, M.; Milošević, V.; Blagojević, D.; Spasić, M.; et al. Effects of antipsychotic drug administration on antioxidative defense enzymes in male rat kidney. J. Toxicol. Environ. Health A 2016, 79, 905–911. [Google Scholar] [CrossRef]
- Nikolić-Kokić, A.; Tatalović, N.; Nestorov, J.; Mijović, M.; Mijusković, A.; Miler, M.; Oreščanin-Dušić, Z.; Nikolić, M.; Milošević, V.; Blagojević, D.; et al. Clozapine, ziprasidone, and sertindole-induced morphological changes in the rat heart and their relationship to antioxidant enzymes function. J. Toxicol. Environ. Health A 2018, 81, 844–853. [Google Scholar] [CrossRef]
- Ardıç, C.M.; Ilgın, S.; Baysal, M.; Karaduman, A.B.; Kılıç, V.; Aydoğan-Kılıç, G.; Uçarcan, Ş.; Atlı-Eklioğlu, Ö. Olanzapine induced reproductive toxicity in male rats. Sci. Rep. 2021, 11, 4739. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Trocchia, S.; Abdel-Gawad, F.K.; Ciarcia, G. Roles of reactive oxygen species in the spermatogenesis regulation. Front. Endocrinol. 2014, 5, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parlaktas, B.S.; Ozyurt, B.; Ozyurt, H.; Tunc, A.T.; Akbas, A. Levels of oxidative stress parameters and the protective effects of melatonin in psychosis model rat testis. Asian J. Androl. 2008, 10, 259–265. [Google Scholar] [CrossRef]
- Choubey, M.; Ranjan, A.; Baltazar, F.; Martin, L.J.; Krishna, A. Role of adiponectin as a modulator of testicular function during aging in mice. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Choubey, M.; Ranjan, A.; Bora, P.S.; Baltazar, F.; Krishna, A. Direct actions of adiponectin on changes in reproductive, metabolic, and anti-oxidative enzymes status in the testis of adult mice. Gen. Comp. Endocrinol. 2019, 279, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.T.; Lysiak, J.J. Oxidative stress: A common factor in testicular dysfunction. J. Androl. 2008, 29, 488–498. [Google Scholar] [CrossRef] [Green Version]
- Görmüş, G.; Ilgın, S.; Baysal, M.; Karaduman, A.B.; Kılıç, V.; Aydoğan-Kılıç, G.; Karagöz, O.; Atlı-Eklioğlu, Ö. Risperidone induced reproductive toxicity in male rats targeting leydig cells and hypothalamic-pituitary-gonadal axis by inducing oxidative stress. Andrologia 2021, 53, e13813. [Google Scholar] [CrossRef]
- Mai, H.N.; Chung, Y.H.; Shin, E.-J.; Jeong, J.H.; Jung, T.W.; Sharma, N.; Lei, X.G.; Namh, S.-Y.; Jang, C.G.; Kim, D.J.; et al. Overexpression of glutathione peroxidase-1 attenuates cocaine-induced reproductive dysfunction in male mice by inhibiting nuclear factor κB. Chem. Biol. Interact. 2019, 307, 136–146. [Google Scholar] [CrossRef]
- Appasamy, M.; Muttukrishna, S.; Pizzey, A.R.; Ozturk, O.; Groome, N.P.; Serhal, P.; Jauniaux, E. Relationship between male reproductive hormones, sperm DNA damage and markers of oxidative stress in infertility. Reprod. Biomed. Online 2007, 14, 159–165. [Google Scholar] [CrossRef]
- Darbandi, M.; Darbandi, S.; Agarwal, A.; Sengupta, P.; Durairajanayagam, D.; Henkel, R.; Sadeghi, M.R. Reactive oxygen species and male reproductive hormones. Reprod. Biol. Endocrinol. 2018, 16, 87. [Google Scholar] [CrossRef]
- Just, M.J. The influence of atypical antipsychotic drugs on sexual function. Neuropsychiatry Dis. Treat. 2015, 11, 1655–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naelitz, B.; Shah, A.; Nowacki, A.S.; Bryk, D.J.; Farber, N.; Parekh, N.; Shoskes, D.; Hatipoglu, B.; Vij, S.C. Prolactin-to-Testosterone Ratio Predicts Pituitary Abnormalities in Mildly Hyperprolactinemic Men with Symptoms of Hypogonadism. J. Urol. 2021, 205, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Cooper, G.D.; Pickavance, L.C.; Wilding, J.P.H.; Harrold, J.A.; Halford, J.C.G.; Goudie, A.J. Effects of olanzapine in male rats: Enhanced adiposity in the absence of hyperphagia, weight gain or metabolic abnormalities Comparative Study. J. Psychopharmacol. 2007, 21, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Creasy, D.M. Pathogenesis of male reproductive toxicity. Toxicol. Pathol. 2001, 29, 64–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, T.T.; Bang, H.J.; Lysiak, J.J. Experimental testicular torsion: Reperfusion blood flow and subsequent testicular venous plasma testosterone concentrations. Urology 2005, 65, 390–394. [Google Scholar] [CrossRef]
- Hanukoglu, I. Antioxidant protective mechanisms against reactive oxygen species (ROS) generated by mitochondrial P450 systems in steroidogenic cells. Drug Metab. Rev. 2006, 38, 171–196. [Google Scholar] [CrossRef]
- Luo, L.; Chen, H.; Trush, M.A.; Show, M.D.; Anway, M.D.; Zirkin, B.R. Aging and the brown Norway rat leydig cell antioxidant defense system. J. Androl. 2006, 27, 240–247. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.J.; Baker, M.A.; Sawyer, D. Oxidative stress in the male germ line and its role in the aetiology of male infertility and genetic disease. Reprod. Biomed. Online 2003, 7, 65–70. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef]
- Beutler, E. Catalase: A Manual of Biochemical Methods. In Red Cell Metabolism; Beutler, E., Ed.; Grune and Stratton: New York, NY, USA, 1982; pp. 105–106. [Google Scholar]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 74–77. [Google Scholar]
- Glatzle, D.; Vuilleumier, J.P.; Weber, F.; Decker, K. Glutathione reductase test with whole blood a convenient procedure for the assessment of the riboflavin status in humans. Experientia 1974, 30, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.L.; Farr, A.L.; Randall, R.I. Protein measurement with Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Luna, L.G. Manual of Histologic Staining Methods of the Armed Forces, 3rd ed.; Blakiston Division, McGraw-Hill: New York, NY, USA, 1968; p. 258. [Google Scholar]
- Hinkle, E.D.; Wiersma, W.; Jurs, G.S. Applied Statistics for Behavioral Sciences, 2nd ed.; Houghton Mifflin Company: Boston, MA, USA, 1994. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Clozapine | Ziprasidone | Sertindole | |
|---|---|---|---|---|
| Plasma ALT (U/L) | 82.8 ± 6.4 | 69.6 ± 3.1 | 88.8 ± 10.2 | 72.1 ± 12.6 |
| Plasma AST (U/L) | 276 ± 21.5 | 244.8 ± 21.09 | 261.2 ± 29.2 | 258.2 ± 16.8 |
| Johnsen Score | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |||
| Control | N | 7 | n = 7 | |||||||||
| % | 100 | |||||||||||
| Clozapine | n | 4 | 2 | n = 6 | ||||||||
| % | 67 | 33 | ||||||||||
| Ziprasidone | n | 3 | 3 | n = 6 | ||||||||
| % | 50 | 50 | ||||||||||
| Sertindole | n | 2 | 6 | n = 8 | ||||||||
| % | 25 | 75 | ||||||||||
| χ2 = 57.77; p < 0.001 | ||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikolić-Kokić, A.; Tatalović, N.; Brkljačić, J.; Mijović, M.; Nestorović, V.; Mijušković, A.; Oreščanin-Dušić, Z.; Vidonja Uzelac, T.; Nikolić, M.; Spasić, S.; et al. Antipsychotic Drug-Mediated Adverse Effects on Rat Testicles May Be Caused by Altered Redox and Hormonal Homeostasis. Int. J. Mol. Sci. 2022, 23, 13698. https://doi.org/10.3390/ijms232213698
Nikolić-Kokić A, Tatalović N, Brkljačić J, Mijović M, Nestorović V, Mijušković A, Oreščanin-Dušić Z, Vidonja Uzelac T, Nikolić M, Spasić S, et al. Antipsychotic Drug-Mediated Adverse Effects on Rat Testicles May Be Caused by Altered Redox and Hormonal Homeostasis. International Journal of Molecular Sciences. 2022; 23(22):13698. https://doi.org/10.3390/ijms232213698
Chicago/Turabian StyleNikolić-Kokić, Aleksandra, Nikola Tatalović, Jelena Brkljačić, Milica Mijović, Vojkan Nestorović, Ana Mijušković, Zorana Oreščanin-Dušić, Teodora Vidonja Uzelac, Milan Nikolić, Snežana Spasić, and et al. 2022. "Antipsychotic Drug-Mediated Adverse Effects on Rat Testicles May Be Caused by Altered Redox and Hormonal Homeostasis" International Journal of Molecular Sciences 23, no. 22: 13698. https://doi.org/10.3390/ijms232213698
APA StyleNikolić-Kokić, A., Tatalović, N., Brkljačić, J., Mijović, M., Nestorović, V., Mijušković, A., Oreščanin-Dušić, Z., Vidonja Uzelac, T., Nikolić, M., Spasić, S., Blagojević, D., & Miljević, Č. (2022). Antipsychotic Drug-Mediated Adverse Effects on Rat Testicles May Be Caused by Altered Redox and Hormonal Homeostasis. International Journal of Molecular Sciences, 23(22), 13698. https://doi.org/10.3390/ijms232213698