
Maternal Dexamethasone Exposure Induces Sex-Specific Changes in Histomorphology and Redox Homeostasis of Rat Placenta

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Fetal and Placental Weight
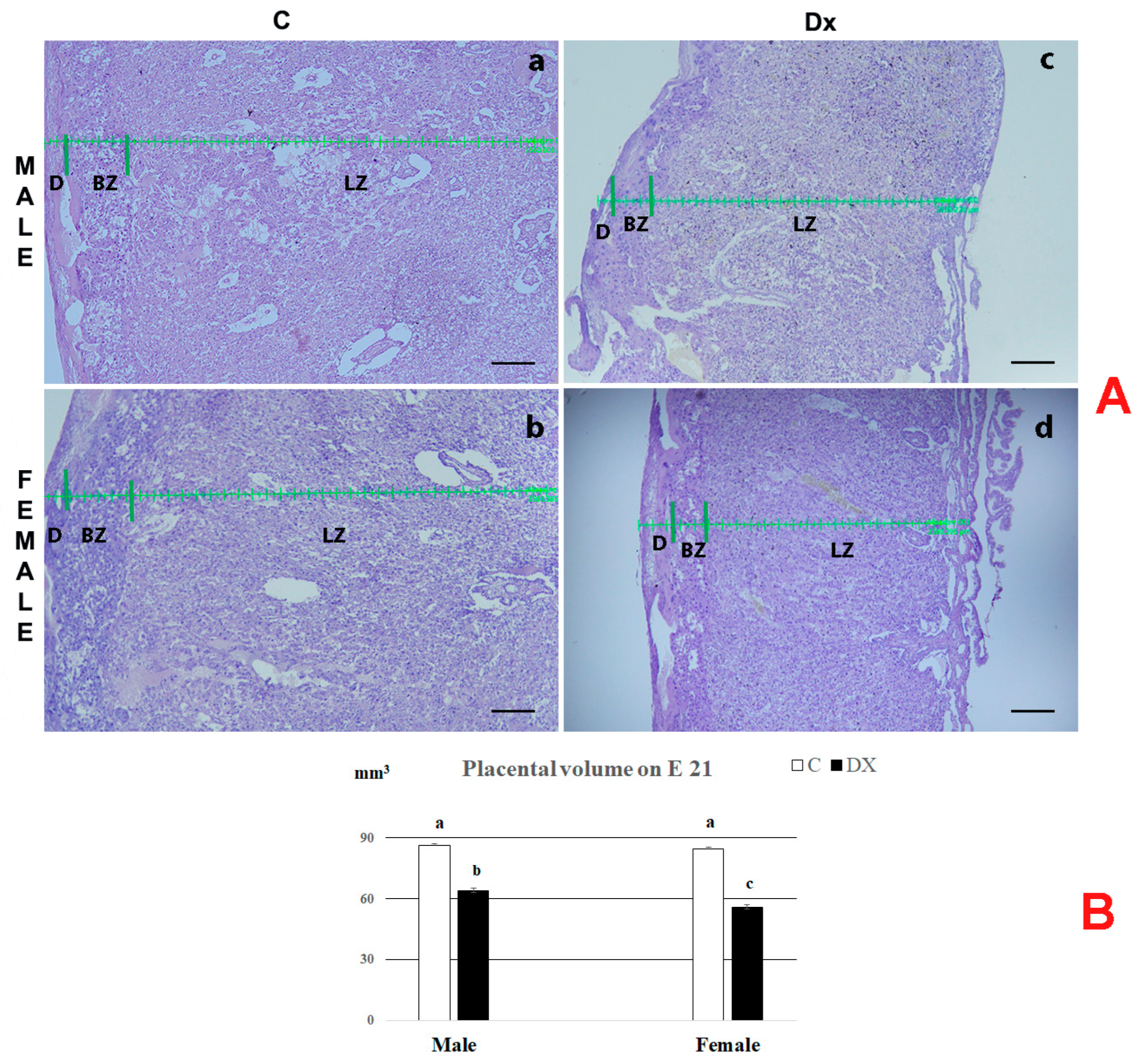
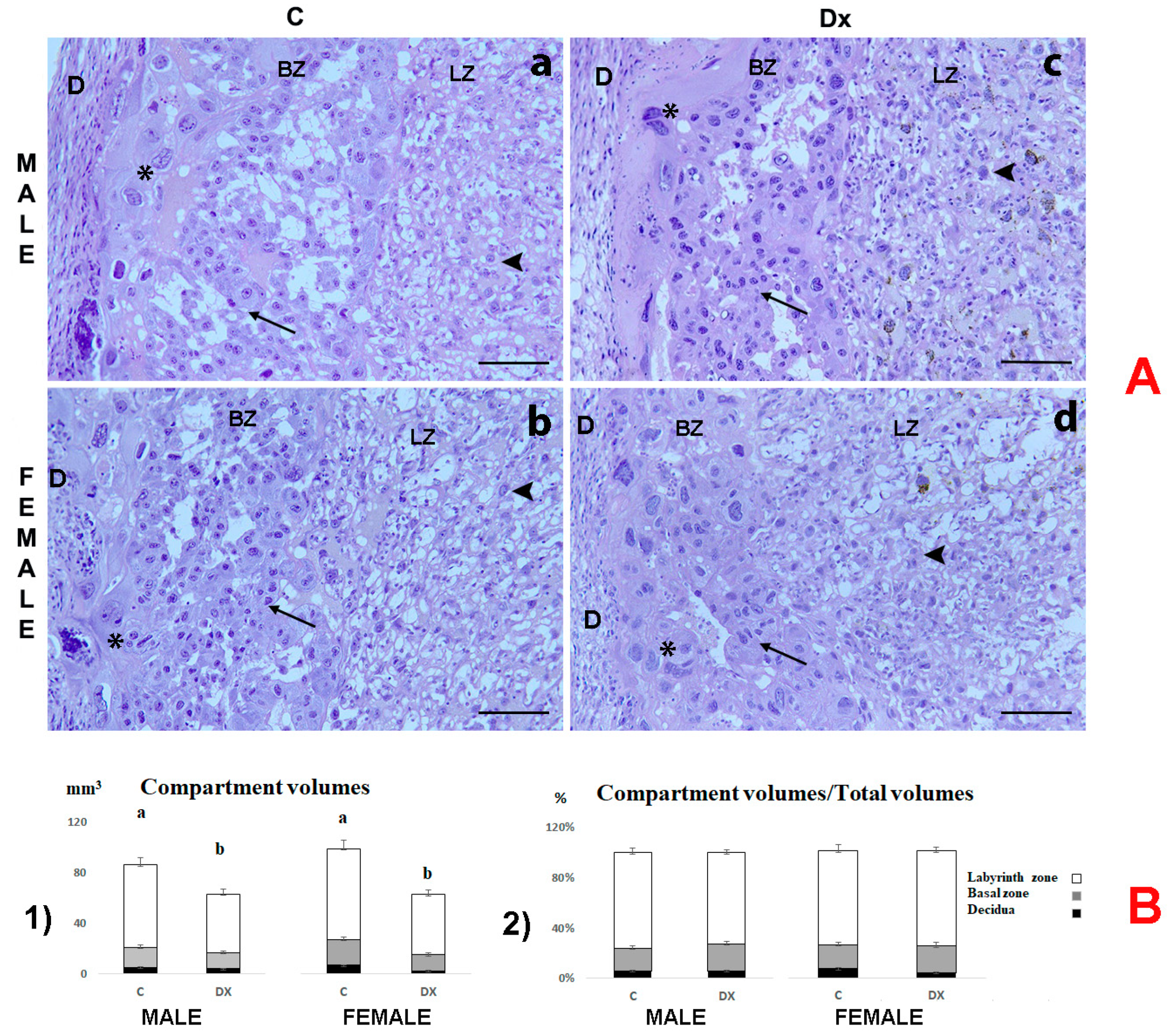
2.2. Histological and Stereological Analysis
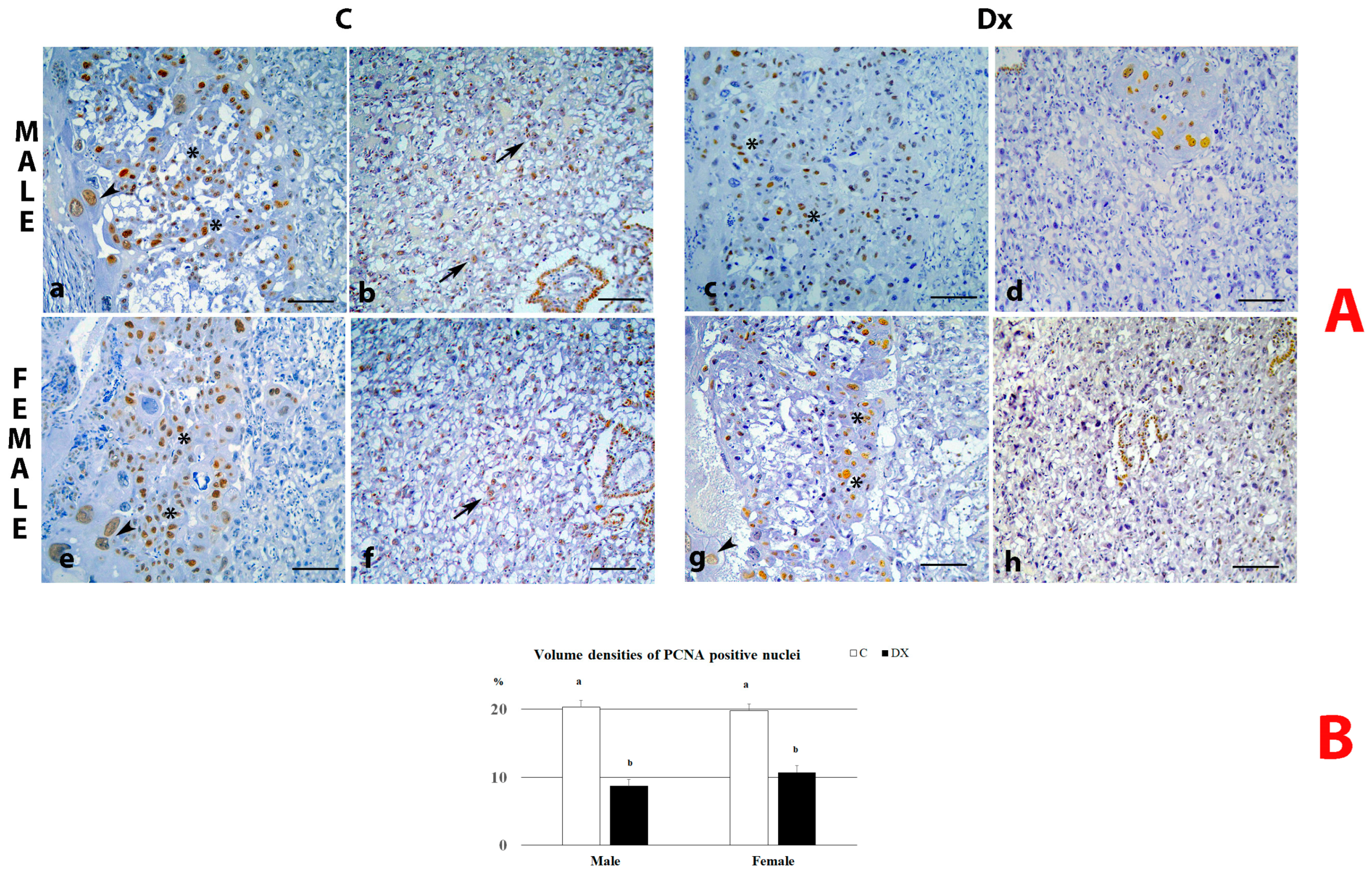
2.3. PCNA Immunostaining Analysis
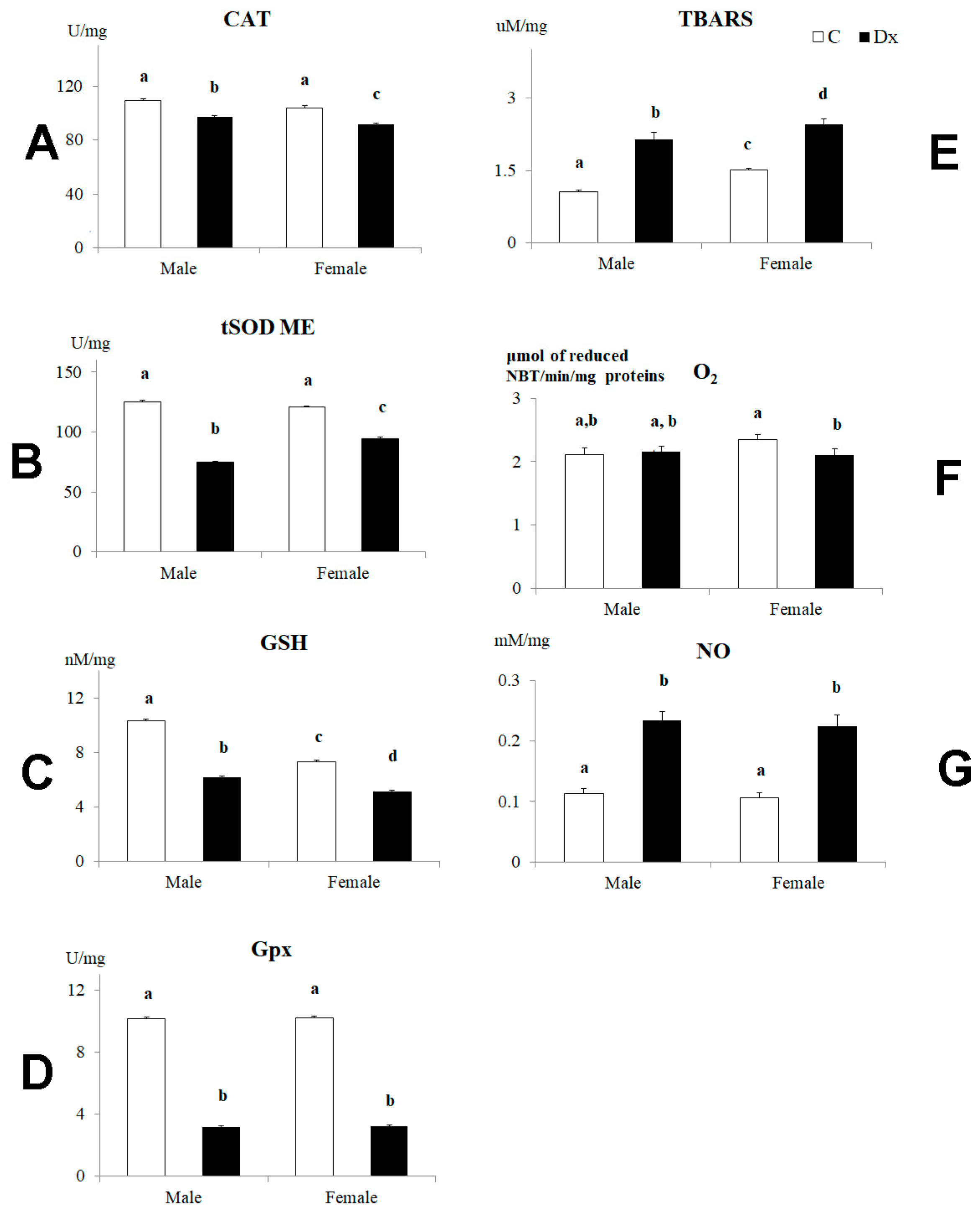
2.4. Oxidative Stress Analysis
3. Discussion
4. Materials and Methods
4.1. Histomorphology and Stereology Analyses
4.2. Determination of Oxidative/Nitrosative Status Parameters
4.3. Reagents
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Burton, G.J.; Fowden, A.L. The placenta: A multifaceted, transient organ. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ain, R.; Konno, T.; Canham, L.; Soares, M. Phenotypic Analysis of the Rat Placenta Methods in Molecular Medicine; Hunt © Humana Press Inc.: Totowa, NJ, USA, 2006; pp. 295–313. [Google Scholar]
- Vacher, C.M.; Lacaille, H.; O’Reilly, J.J.; Salzbank, J.; Bakalar, D.; Sebaoui, S.; Liere, P.; Clarkson-Paredes, C.; Sasaki, T.; Sathyanesan, A.; et al. Placental endocrine function shapes cerebellar development and social behavior. Nat. Neurosci. 2021, 24, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Bale, T.L. The placenta and neurodevelopment: Sex differences in prenatal vulnerability. Dialogues Clin. Neurosci. 2016, 18, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, C.S. Sex-Specific Placental Responses in Fetal Development. Endocrinology 2015, 156, 3422–3434. [Google Scholar] [CrossRef] [Green Version]
- Busada, J.T.; Cidlowski, J.A. Mechanisms of Glucocorticoid Action During Development. Curr. Top. Dev. Biol. 2017, 125, 147–170. [Google Scholar]
- Moisiadis, V.G.; Matthews, S.G. Glucocorticoids and fetal programming part 1: Outcomes. Nat. Rev. Endocrinol. 2014, 10, 391–402. [Google Scholar] [CrossRef]
- Alur, P. Sex Differences in Nutrition, Growth, and Metabolism in Preterm Infants. Front. Pediatr. 2019, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Takagi, Y.; Nikaido, T.; Toki, T.; Kita, N.; Kanai, M.; Ashida, T.; Satoshi Ohira, S.; Konishi, I. Levels of oxidative stress and redox-related molecules in the placenta in preeclampsia and fetal growth restriction. Virchows Arch. 2004, 444, 49–55. [Google Scholar] [CrossRef]
- Ristić, N.; Nestorović, N.; Manojlović-Stojanoski, M.; Trifunović, S.; Ajdžanović, V.; Filipović, B.; Pendovski, L.; Milošević, V. Adverse effect of dexamethasone on development of the fetal rat ovary. Fundam. Clin. Pharmacol. 2018, 33, 199–207. [Google Scholar] [CrossRef] [Green Version]
- Ristić, N.; Nestorović, N.; Manojlović-Stojanoski, M.; Trifunović, S.; Ajdžanović, V.; Filipović, B.; Pendovski, L.; Milošević, V. Prenatal dexamethasone exposure and developmental programming of the ovary of the offspring: A structural study in the rat. Reprod. Fertil. Dev. 2021, 33, 245–255. [Google Scholar] [CrossRef]
- Manojlovic-Stojanoski, M.; Lavrnja, I.; Stevanovic, I.; Trifunovic, S.; Ristic, N.; Nestorovic, N.; Sévigny, J.; Nedeljkovic, N.; Laketa, D. Antenatal Dexamethasone Treatment Induces Sex-dependent Upregulation of NTPDase1/CD39 and Ecto-5’-nucleotidase/CD73 in the Rat Fetal Brain. Cell. Mol. Neurobiol. 2021, 42, 1965–1981. [Google Scholar] [CrossRef] [PubMed]
- Dennery, P.A. Oxidative stress in development: Nature or nurture? Free Radic. Biol. Med. 2010, 49, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Arogbokun, O.; Rosen, E.; Keil, A.P.; Milne, G.L.; Barrett, E.; Nguyen, R.; Bush, N.R.; Swan, S.H.; Sathyanarayana, S.; Ferguson, K.K. Maternal Oxidative Stress Biomarkers in Pregnancy and Child Growth from Birth to Age 6. J. Clin. Endocrinol. Metab. 2021, 106, 1427–1436. [Google Scholar] [CrossRef] [PubMed]
- Duhig, K.; Chappell, L.C.; Shennan, A.H. Oxidative stress in pregnancy and reproduction. Obstet. Med. 2016, 9, 113–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, M.L.; Mark, P.J.; Lewis, J.L.; Mori, T.A.; Keelan, J.A.; Waddell, B.J. Antioxidant defenses in the rat placenta in late gestation: Increased labyrinthine expression of superoxide dismutase, glutathione peroxidase 3, and uncoupling protein 2. Biol. Reprod. 2010, 83, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Stark, M.J.; Hodyl, N.A.; Wright, I.M.R.; Clifton, V.L. Influence of sex and glucocorticoid exposure on preterm placental pro-oxidant-antioxidant balance. Placenta 2011, 32, 865–870. [Google Scholar] [CrossRef]
- Cuffe, J.S.M.; Saif, Z.; Perkins, A.V.; Moritz Clifton, V.L. Dexamethasone and sex regulate placental glucocorticoid receptor isoforms in mice. J. Endocrinol. 2017, 234, 89–100. [Google Scholar] [CrossRef]
- Chakraborty, S.; Islam, S.; Saha, S.; Ain, R. Dexamethasone-induced Intra-Uterine Growth Restriction impacts NOSTRIN and its downstream effector genes in the rat mesometrial uterus. Sci. Rep. 2018, 8, 8342. [Google Scholar] [CrossRef] [Green Version]
- Cuffe, J.S.; O’Sullivan, L.; Simmons, D.G.; Anderson, S.T.; Moritz, K.M. Maternal corticosterone exposure in the mouse has sex-specific effects on placental growth and mRNA expression. Endocrinology 2012, 153, 5500–5511. [Google Scholar] [CrossRef] [Green Version]
- Odermatt, A.; Kratschmar, D.V. Tissue-specific modulation of mineralocorticoid receptor function by 11β-hydroxysteroid dehydrogenases: An overview. Mol. Cell. Endocrinol. 2012, 350, 168–186. [Google Scholar] [CrossRef]
- Cuffe, J.S.M.; Dickinson, H.; Simmons, D.G.; Moritz, K.M. Sex specific changes in placental growth and MAPK following short term maternal dexamethasone exposure in the mouse. Placenta 2011, 32, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Tsuji, N.; Sugiyama, A. Morphology and physiology of rat placenta for toxicological evaluation. Review. J. Toxicol. Pathol. 2019, 32, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, S.; Tsuji, N.; Hayashi, S.; Abe, M.; Hagio, S.; Yamagishi, Y.; Kuroda, Y.; Sugiyama, A. Histomorphological comparison of rat placentas by different timing of chlorpromazine- administration. Exp. Toxicol. Pathol. 2015, 67, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Cain, D.W.; Cidlowski, J.A. Specificity and sensitivity of glucocorticoid signaling in health and disease. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 545–556. [Google Scholar] [CrossRef] [Green Version]
- Saif, Z.; Hodyl, N.A.; Stark, M.J.; Fuller, P.J.; Cole, T.; Lu, N.; Clifton, V.L. Expression of eight glucocorticoid receptor isoforms in the human preterm placenta vary with fetal sex and birthweight. Placenta 2015, 36, 723–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saif, Z.; Dyson, R.M.; Palliser, H.K.; Wright, I.M.; Lu, N.; Clifton, V.L. Identification of eight different isoforms of the glucocorticoid receptor in guinea pig placenta: Relationship to preterm delivery, sex and betamethasone exposure. PLoS ONE 2016, 1, e0148226. [Google Scholar] [CrossRef] [Green Version]
- Oakley, R.H.; Cidlowski, J.A. The biology of the glucocorticoid receptor: New signaling mechanisms in health and disease. J. Allergy Clin. Immunol. 2013, 132, 1033–1044. [Google Scholar] [CrossRef] [Green Version]
- Wu, I.; Shin, S.C.; Cao, Y.; Bender, I.K.; Jafari, N.; Feng, G.; Lin, S.; Cidlowski, J.A.; Schleimer, R.P.; Lu, N.Z. Selective glucocorticoid receptor translational isoforms reveal glucocorticoid-induced apoptotic transcriptomes. Cell Death Dis. 2013, 4, e453. [Google Scholar] [CrossRef]
- Myatt, L.; Cui, X. Oxidative stress in the placenta. Histochem. Cell Biol. 2004, 122, 369–382. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, F.C.; Fragoso, M.B.T.; Bueno, N.B.; Goulart, M.O.F.; Oliveira, A.C.M. Oxidative stress markers in preeclamptic placentas: A systematic review with meta-analysis. Placenta 2020, 99, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Little, B.D.; Hopkins, R.Z. Superoxide dismutases in biology and medicine: Essentials and recent advances. Reac. Oxyg. Species 2020, 9, 13–21. [Google Scholar] [CrossRef]
- Taysi, S.; Tascan, A.S.; Ugur, M.G.; Demir, M. Radicals, oxidative/nitrosative stress and preeclampsia. Mini Rev. Med. Chem. 2019, 19, 178–193. [Google Scholar] [CrossRef] [PubMed]
- You, J.-M.; Yun, S.-J.; Nam, K.N.; Kang, C.; Won, R.; Lee, E.H. Mechanism of glucocorticoid-induced oxidative stress in rat hippocampal slice cultures. Can. J. Physiol. Pharmacol. 2009, 87, 440–447. [Google Scholar] [CrossRef]
- Spiers, J.G.; Chen, H.J.; Cuffe, J.S.; Sernia, C.; Lavidis, N.A. Acute restraint stress induces rapid changes in central redox status and protective antioxidant genes in rats. Psychneuroendocrinology 2016, 67, 104–112. [Google Scholar] [CrossRef]
- Fowden, A.L.; Ward, J.W.; Wooding, F.P.; Forhead, A.J.; Constancia, M. Programming placental nutrient transport capacity. J. Physiol. 2006, 572, 5–15. [Google Scholar] [CrossRef]
- Schoots, M.H.; Gordijn, S.J.; Scherjon, S.A.; van Goor, H.; Hillebrands, J.L. Oxidative stress in placental pathology. Placenta 2018, 69, 153–161. [Google Scholar] [CrossRef]
- Biri, A.; Bozkurt, N.; Turp, A.; Kavutcu, M.; Himmetoglu, O.; Durak, I. Role of oxidative stress in intrauterine growth restriction. Gynecol. Obstet. Investig. 2007, 64, 187–192. [Google Scholar] [CrossRef]
- Gaccioli, F.; Lager, S.; Powell, T.L.; Jansson, T. Placental transport in response to altered maternal nutrition. J. Dev. Orig. Health Dis. 2013, 4, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Myatt, L.; Maloyan, A. Obesity and Placental Function. Semin. Reprod. Med. 2016, 34, 42–49. [Google Scholar]
- Baisden, B.; Sonne, S.; Joshi, R.M.; Ganapathy, V.; Shekhawat, P.S. Antenatal dexamethasone treatment leads to changes in gene expression in a murine late placenta. Placenta 2007, 28, 1082–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Dong, W.; Jiao, Z.; Liu, J.; Li, K.; Wang, H.; Xu, D. Prenatal Dexamethasone Exposure Induced Alterations in Neurobehavior and Hippocampal Glutamatergic System Balance in Female Rat Offspring. Toxicol. Sci. 2019, 31, kfz163. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, H.J. Stereology of arbitrary particles. A review of unbiased number and size estimators and the presentation of some new ones, in memory of William R. Thompson. J. Microsc. 1986, 43, 3–45. [Google Scholar] [CrossRef]
- Gundersen, H.J.; Jensen, E.B. The efficiency of systematic sampling in stereology and its prediction. J. Microsc. 1987, 147, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Dorph-Petersen, K.A.; Nyengaard, J.R.; Gundersen, H.J. Tissue shrinkage and unbiased stereological estimation of particle number and size. J. Microsc. 2001, 204, 232–246. [Google Scholar] [CrossRef]
- Takahashi, T.; Caviness, V.S., Jr. PCNA-binding to DNA at the G1/S tran-sition in proliferating cells of the developing cerebral wall. J. Neurocytol. 1993, 22, 1096–1102. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosenbrongh, N.J.; Farr, A.L.; Randal, R.J. Protein measurement with the folin phenol. reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Villacara, A.; Kumami, K.; Yamamoto, T.; Mrsulja, B.B.; Spatz, M. Ischemic modification of cerebrocortical membranes: 5-hydroxytryptamine receptors, fluidity, and inducible in vitro lipid peroxidation. J. Neurochem. 1989, 53, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Gonzálvez, J.A.; Garcia-Benayas, C.; Arenas, J. Semiautomated measurement of nitrate in biological fluids. Clin. Chem. 1998, 44, 679–681. [Google Scholar] [CrossRef] [Green Version]
- Auclair, C.; Voisin, E. Nitroblue tetrazolium reduction. In Handbook of Methods for Oxygen Radical Research; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 123–132. [Google Scholar]
- Sun, M.; Zigman, S. An important spectrophotometric assay for superoxide dismutase based on epinephrine auto-oxidation. Anal Biochem. 1978, 90, 81–89. [Google Scholar] [CrossRef]
- Góth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Maral, J.; Puget, K.; Michelson, A.M. Comparative study of superoxide dismutase, catalase and glutathione peroxidase levels in erythrocytes of different animals. Biochem. Biophys. Res. Commun. 1977, 77, 1525–1535. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.E. Tissue glutathione. In Handbook of Methods for Oxygen Radical Research; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 317–323. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Male -Control (g ± SD) | Female-Control (g ± SD) | Male-Dx Group (g ± SD) | Female-Dx Group (g ± SD) | |
|---|---|---|---|---|
| Fetal weight | 5 ± 0.04 | 4.9 ± 0.03 | 4.2 ± 0.4 a,b | 4.1 ± 0.04 a,b |
| Placental weight | 0.62 ± 0.06 | 0.59 ± 0.05 | 0.36 ± 0.03 a,b | 0.35 ± 0.03 a,b |
| Placental to fetal Weight ratio | 0.124 ± 0.002 | 0.120 ± 0.003 | 0.082 ± 0.0001 a,b | 0.085 ± 0.003 a,b |
| Factor | Treatment | Sex | Treatment × Sex | |||
|---|---|---|---|---|---|---|
| F (DFn, DFd) | P | F (DFn, DFd) | P | F (DFn, DFd) | P | |
| CAT | 682.3 (1, 24) | 0.000 * | 146.2 (1, 24) | 0.000 * | 0.0 (1, 24) | 0.98 |
| tSOD | 7963.2 (1, 24) | 0.000 * | 315.3 (1, 24) | 0.000 * | 772.7 (1, 24) | 0.000 * |
| GSH | 5471.7 (1, 24) | 0.000 * | 2260.3 (1, 24) | 0.000 * | 514.1 (1, 24) | 0.000 * |
| Gpx | 34,768.1 (1, 24) | 0.000 * | 0.6 (1, 24) | 0.444 | 0.0 (1, 24) | 0.912 |
| TBARS | 662 (1, 24) | 0.000 * | 96.3 (1, 24) | 0.000 * | 3.3 (1, 24) | 0.081 |
| O2•− | 8.5 (1, 24) | 0.008 | 5.94 (1, 24) | 0.023 | 17.8 (1, 24) | 0.000 * |
| NO | 479.3 (1, 24) | 0.000 * | 2.36 (1, 24) | 0.136 | 0.1 (1, 24) | 0.761 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trifunović, S.; Šošić Jurjević, B.; Ristić, N.; Nestorović, N.; Filipović, B.; Stevanović, I.; Begović-Kuprešanin, V.; Manojlović-Stojanoski, M. Maternal Dexamethasone Exposure Induces Sex-Specific Changes in Histomorphology and Redox Homeostasis of Rat Placenta. Int. J. Mol. Sci. 2023, 24, 540. https://doi.org/10.3390/ijms24010540
Trifunović S, Šošić Jurjević B, Ristić N, Nestorović N, Filipović B, Stevanović I, Begović-Kuprešanin V, Manojlović-Stojanoski M. Maternal Dexamethasone Exposure Induces Sex-Specific Changes in Histomorphology and Redox Homeostasis of Rat Placenta. International Journal of Molecular Sciences. 2023; 24(1):540. https://doi.org/10.3390/ijms24010540
Chicago/Turabian StyleTrifunović, Svetlana, Branka Šošić Jurjević, Nataša Ristić, Nataša Nestorović, Branko Filipović, Ivana Stevanović, Vesna Begović-Kuprešanin, and Milica Manojlović-Stojanoski. 2023. "Maternal Dexamethasone Exposure Induces Sex-Specific Changes in Histomorphology and Redox Homeostasis of Rat Placenta" International Journal of Molecular Sciences 24, no. 1: 540. https://doi.org/10.3390/ijms24010540
APA StyleTrifunović, S., Šošić Jurjević, B., Ristić, N., Nestorović, N., Filipović, B., Stevanović, I., Begović-Kuprešanin, V., & Manojlović-Stojanoski, M. (2023). Maternal Dexamethasone Exposure Induces Sex-Specific Changes in Histomorphology and Redox Homeostasis of Rat Placenta. International Journal of Molecular Sciences, 24(1), 540. https://doi.org/10.3390/ijms24010540