
Impact of JH Signaling on Reproductive Physiology of the Classical Insect Model, Rhodnius prolixus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
Subsections
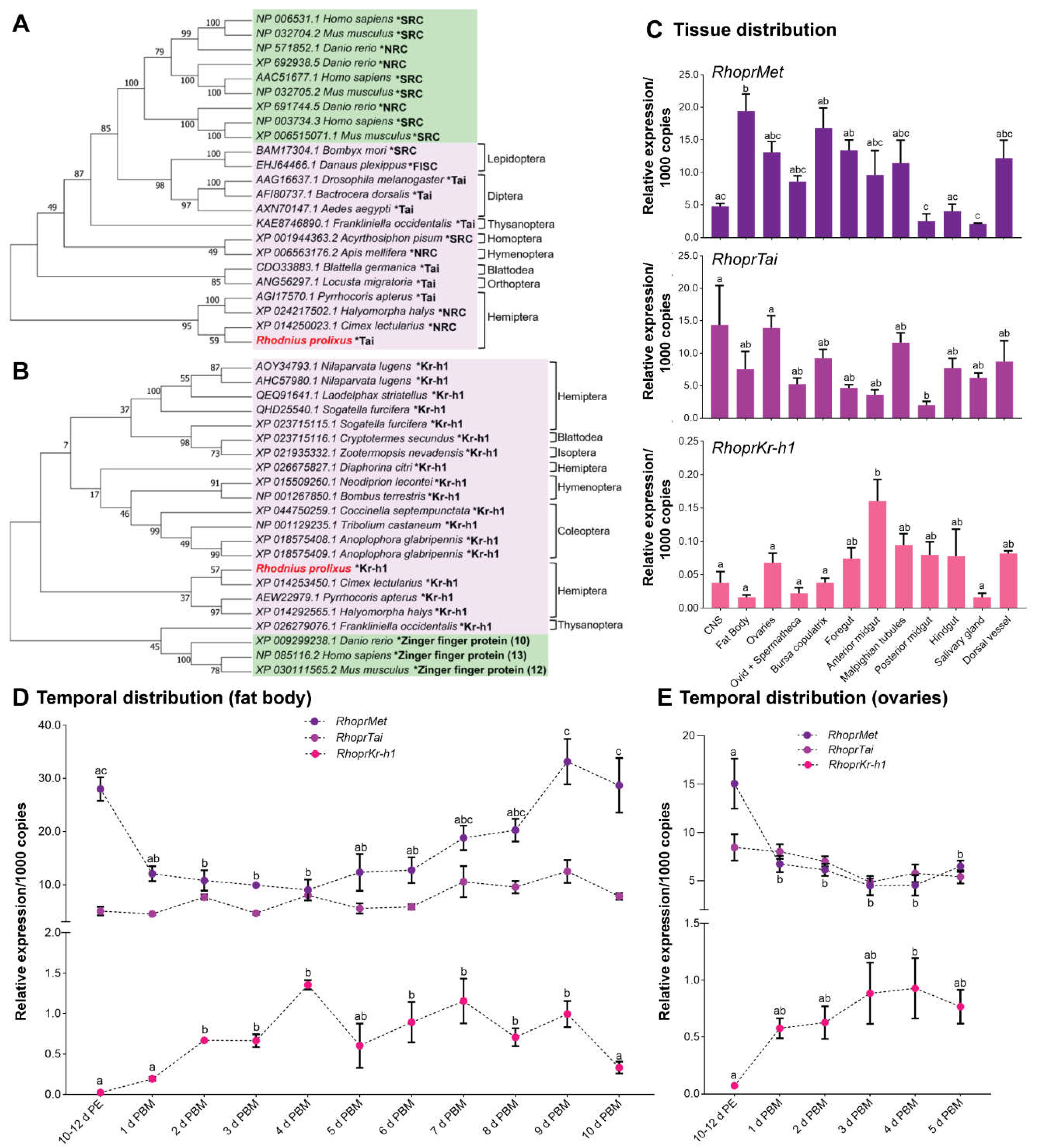
- RhoprMet and RhoprTai transcripts are constitutively expressed in R. prolixus but RhoprKr-h1 appears to be a nutritional response gene, as are the yolk protein precursors.
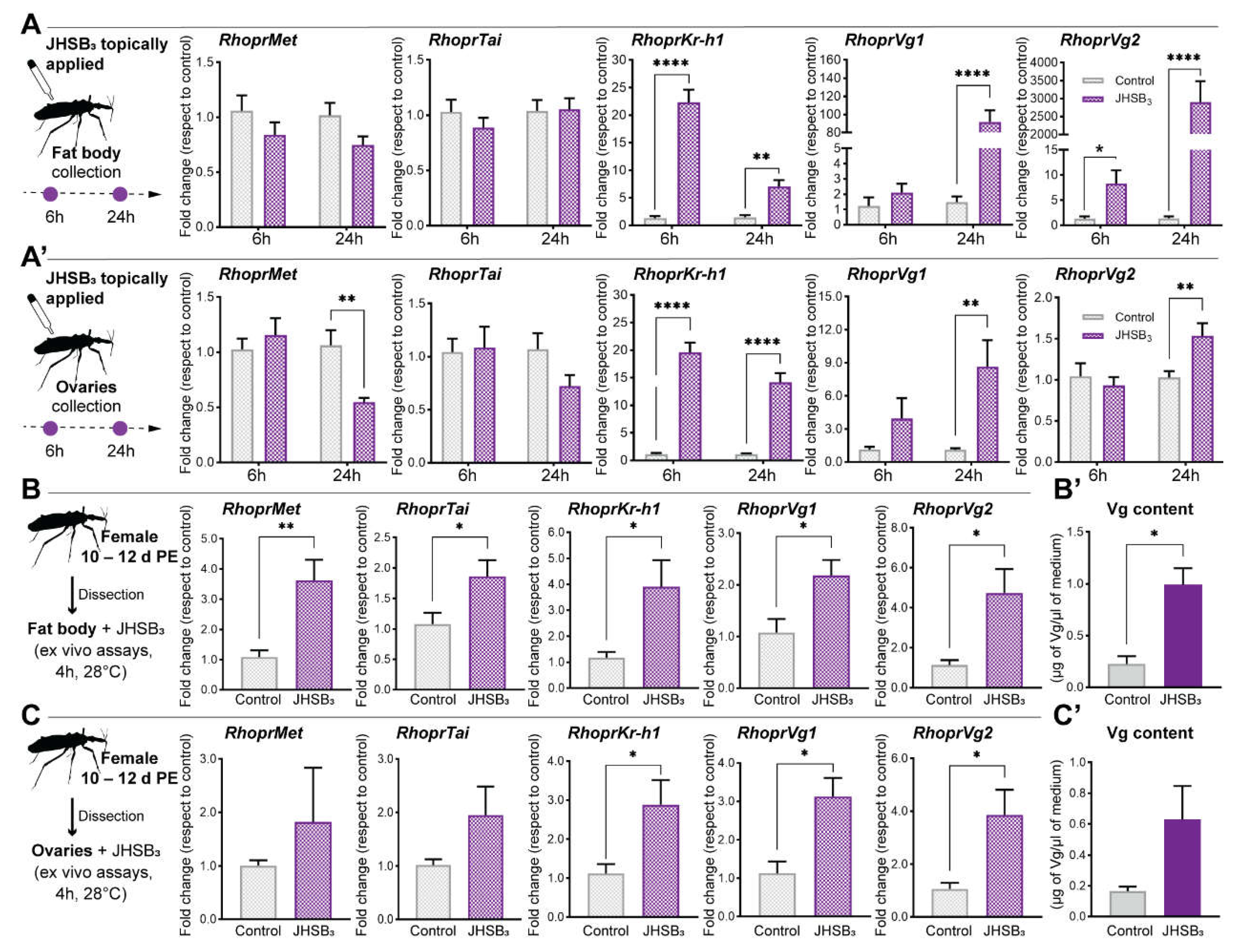
- RhoprKr-h1 mRNA is a JHSB3-dependent gene, as are the yolk protein precursors
- JHSB3 target genes are controlled through Met-Tai complex
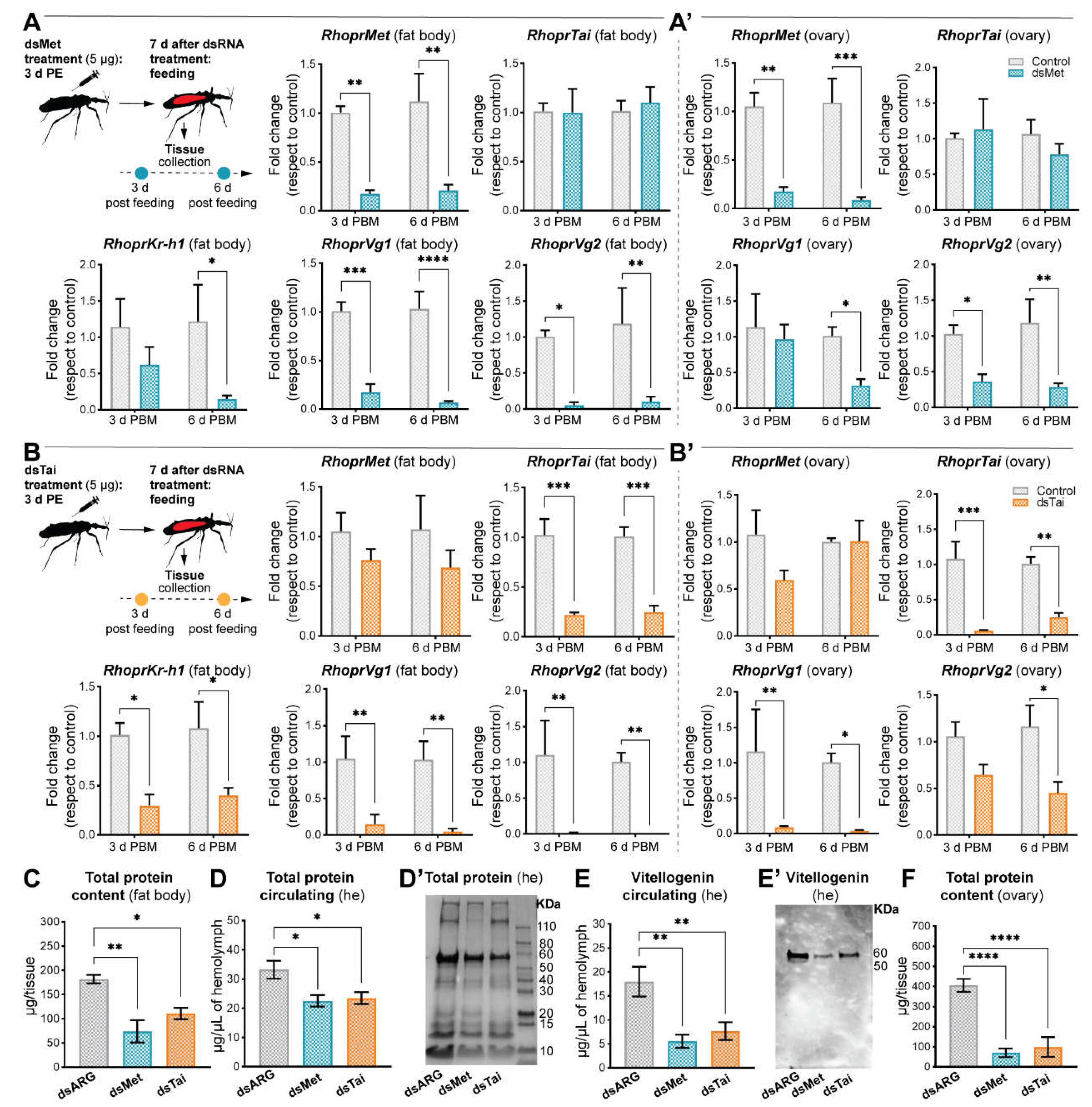
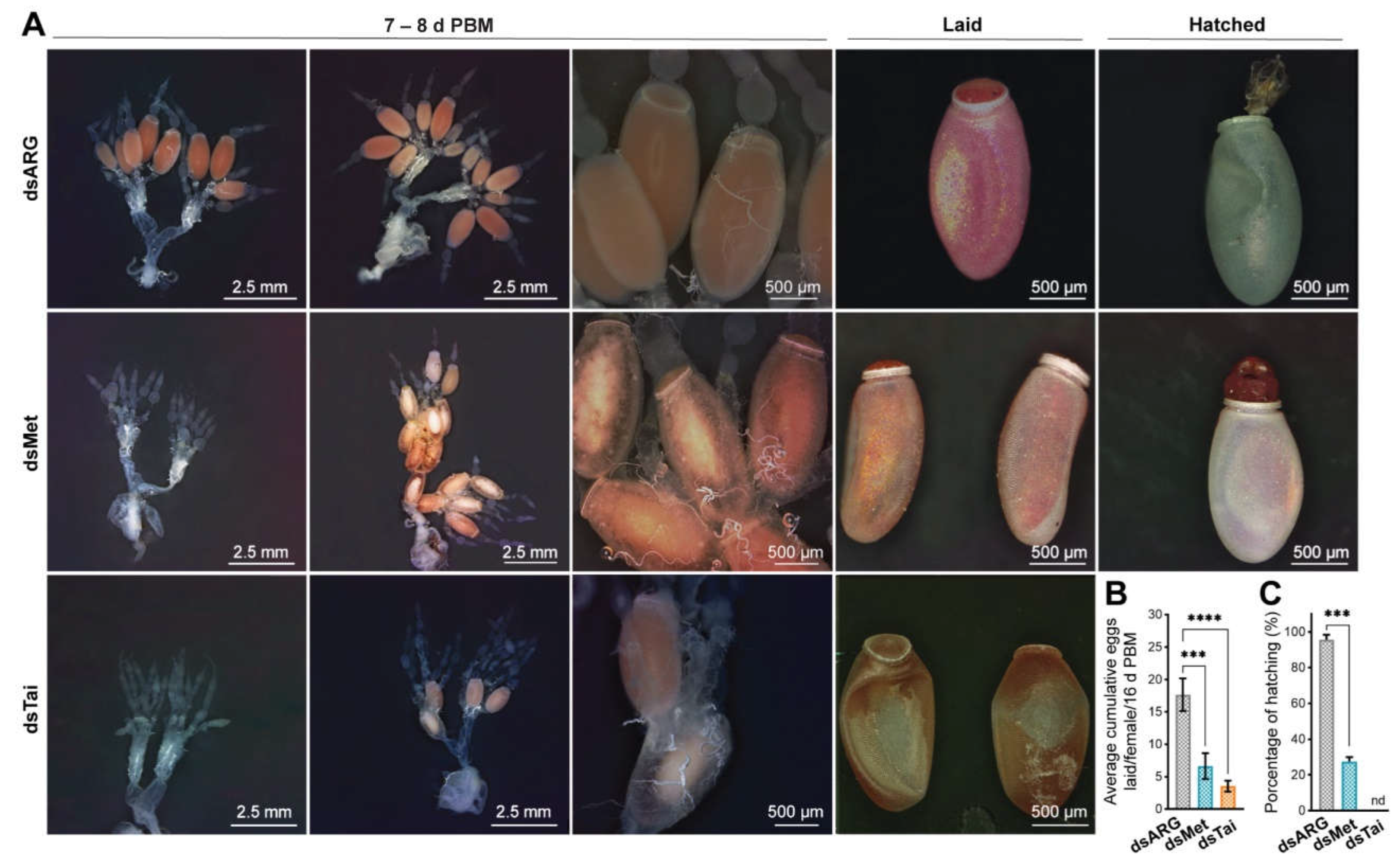
- Result of knockdown of RhoprMet and RhoprTai on the reproductive performance
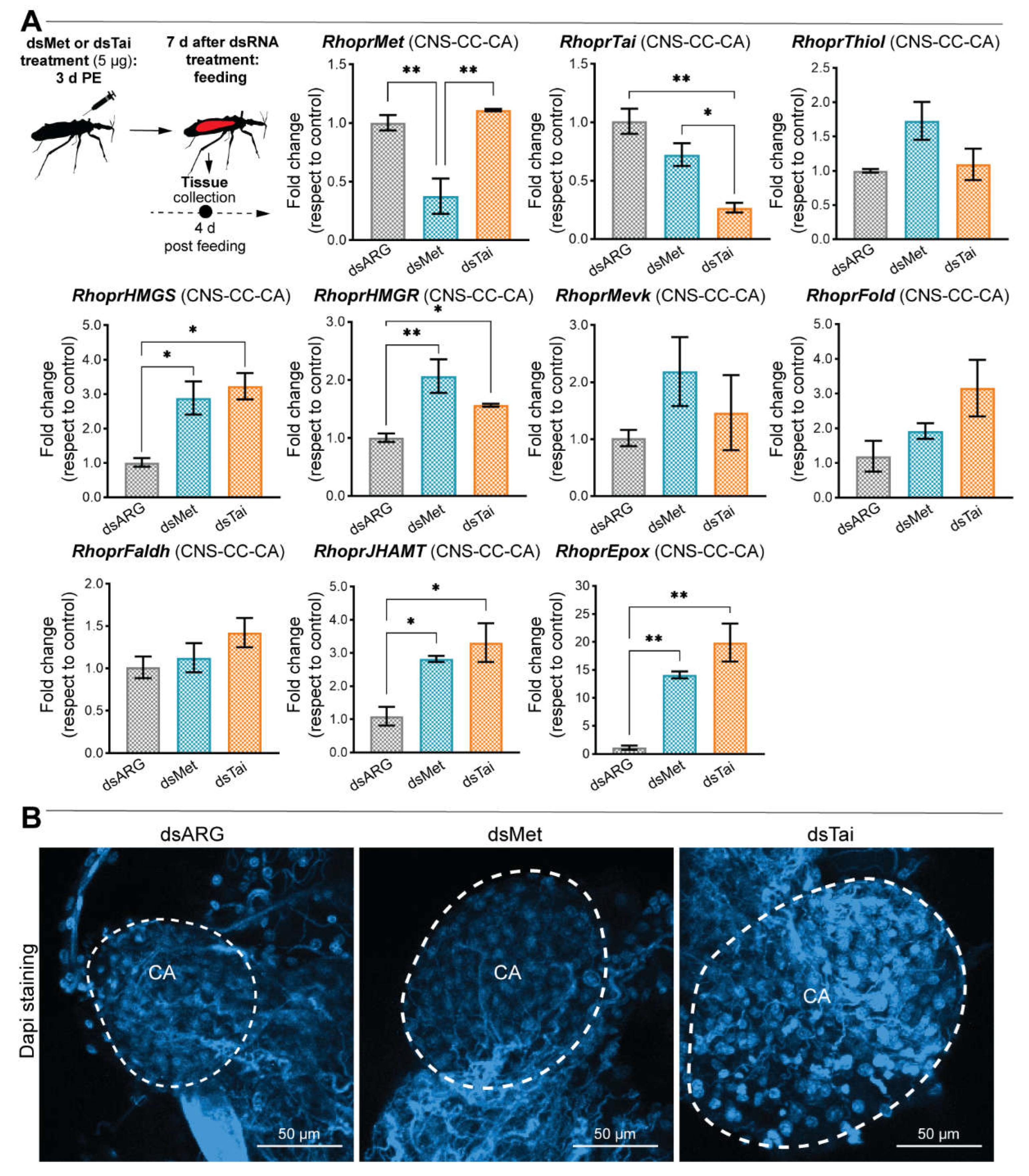
- RhoprMet and RhoprTai appear to regulate JHSB3 biosynthesis in the corpus allatum
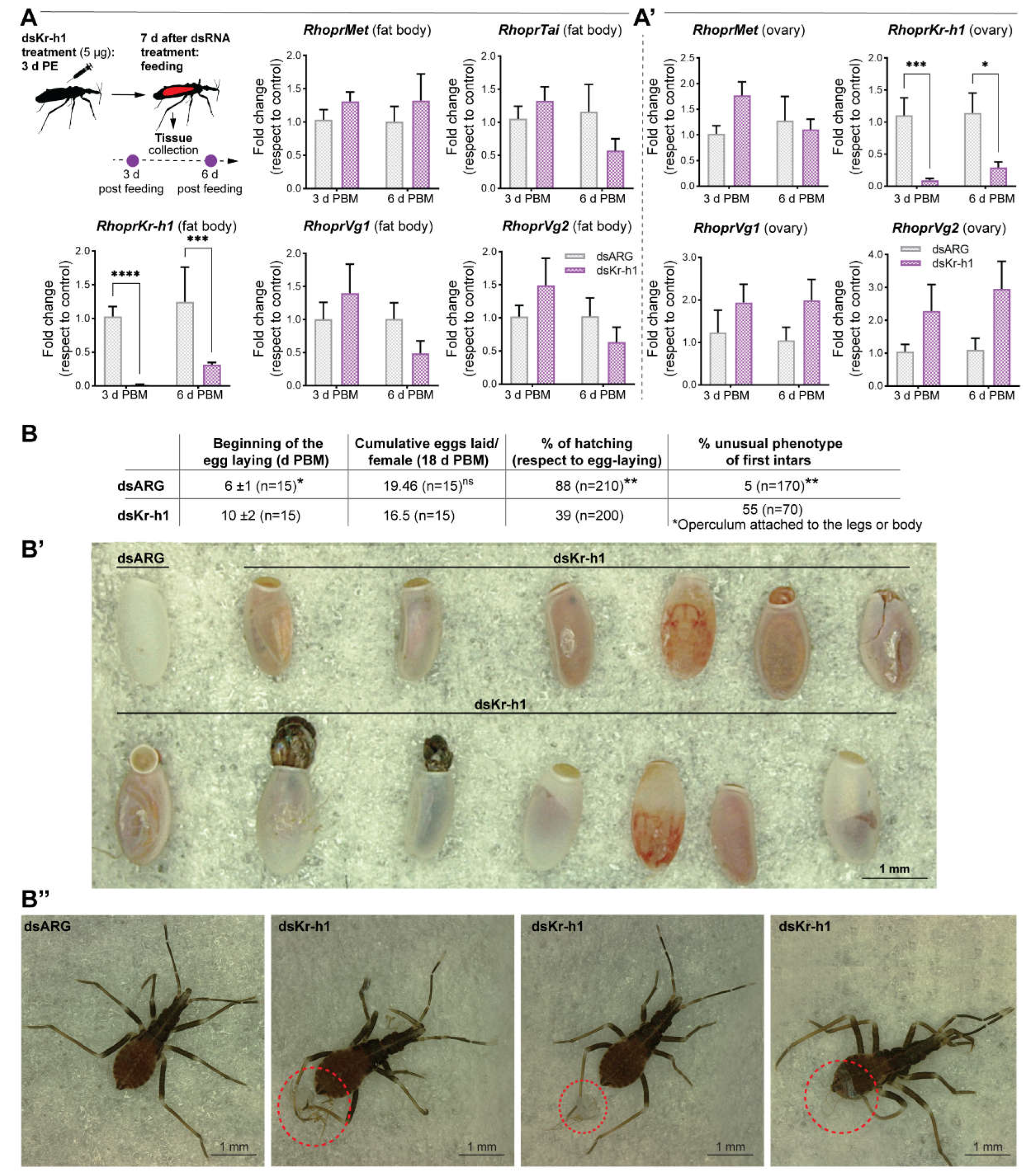
- RhoprKr-h1 does not appear to be involved in vitellogenesis but rather in successful hatching
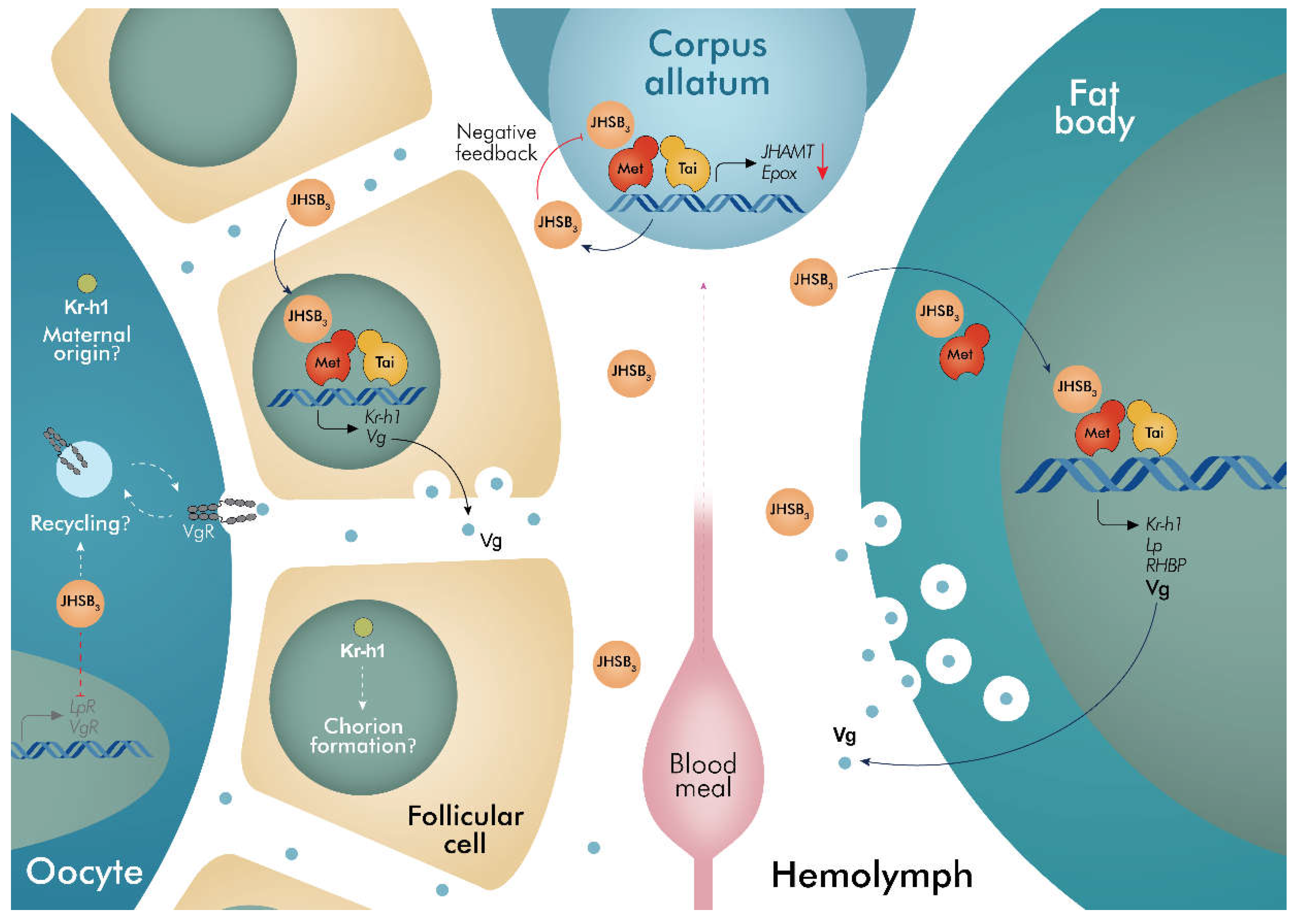
3. Discussion
- JHSB3 stimulates yolk protein synthesis and negatively modulates endocytic receptor transcript expression
- Both RhoprMet and RhoprTai are critically important for successful vitellogenesis
- RhoprMet and RhoprTai appear to regulate JHSB3 biosynthesis in the corpus allatum
- RhoprKr-h1 does not appear to be involved in vitellogenesis but rather in successful hatching
4. Materials and Methods
- Experimental Animals
- Phylogenetic Analysis of RhoprTai and RhoprKr-h1
- RNA extraction and reverse transcription/quantitative PCR (RT-qPCR)
- JHSB3 treatment: in vivo experiments
- JHSB3 treatment: ex vivo experiments
- RNAi assay
- Protein and lipid measurements
- SDS-PAGE and Western blot
- Quantification of vitellogenin by ELISA
- Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Wigglesworth, V.B. The function of the corpus allatum in the growth and reproduction of Rhodnius prolixus (Hemiptera). Q. J. Microsc. Sci. 1936, 79, 91–121. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. The physiology of ecdysis in Rhodnius prolixus (Hemiptera). II. Factors controlling moulting and ‘metamorphosis’. Q. J. Microsc. Sci. 1934, 77, 191–222. [Google Scholar] [CrossRef]
- Davey, K.G. The modes of action of juvenile hormones: Some questions we ought to ask. Insect Biochem. Mol. Biol. 2000, 30, 663–669. [Google Scholar] [CrossRef]
- Davey, K.G. The interaction of feeding and mating in the hormonal control of egg production in Rhodnius prolixus. J. Insect Physiol. 2007, 53, 208–215. [Google Scholar] [CrossRef]
- Tsang, S.; Law, S.; Li, C.; Qu, Z.; Bendena, W.G.; Tobe, S.S.; Hui, J.H.L. Diversity of Insect Sesquiterpenoid Regulation. Front. Genet. 2020, 11, 1027. [Google Scholar] [CrossRef]
- Villalobos-Sambucaro, M.J.; Nouzova, M.; Ramirez, C.E.; Alzugaray, M.E.; Fernandez-Lima, F.; Ronderos, J.R.; Noriega, F.G. The juvenile hormone described in Rhodnius prolixus by Wigglesworth is juvenile hormone III skipped bisepoxide. Sci. Rep. 2020, 10, 3091. [Google Scholar] [CrossRef] [Green Version]
- Wilson, T.G.; Fabian, J.A. Drosophila melanogaster mutant resistant to a chemical analog of juvenile hormone. Dev. Biol. 1996, 118, 190–201. [Google Scholar] [CrossRef]
- Konopova, B.; Jindra, M. Juvenile hormone resistance gene Methoprene-tolerant controls entry into metamorphosis in the beetle Tribolium castaneum. Proc. Natl. Acad. Sci. USA 2007, 104, 10488–10493. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Mead, E.A.; Zhu, J. Heterodimer of two bHLH-PAS proteins mediates juvenile hormone-induced gene expression. Proc. Natl. Acad. Sci. USA 2011, 108, 638–643. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Xu, J.; Sheng, Z.; Sui, Y.; Palli, S.R. Steroid receptor co-activator is required for juvenile hormone signal transduction through a bHLH-PAS transcription factor, methoprene tolerant. J. Biol. Chem. 2011, 286, 8437–8447. [Google Scholar] [CrossRef]
- Kayukawa, T.; Minakuchi, C.; Namiki, T.; Togawa, T.; Yoshiyama, M.; Kamimura, M.; Mita, K.; Imanishi, S.; Kiuchi, M.; Ishikawa, Y.; et al. Transcriptional regulation of juvenile hormone-mediated induction of Krüppel homolog 1, a repressor of insect metamorphosis. Proc. Natl. Acad. Sci. USA 2012, 109, 11729–11734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.L.; Liu, C.Y.; Liu, W.; Wang, D.; Wang, J.X.; Zhao, X.F. Methoprene-tolerant 1 regulates gene transcription to maintain insect larval status. J. Mol. Endocrinol. 2014, 53, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minakuchi, C.; Zhou, X.; Riddiford, L.M. Krüppel homolog 1 (Kr-h1) mediates juvenile hormone action during metamorphosis of Drosophila melanogaster. Mech. Dev. 2008, 125, 91–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Wu, Z.; Wang, Z.; Deng, S.; Zhou, S. Krüppel-homolog 1 mediates juvenile hormone action to promote vitellogenesis and oocyte maturation in the migratory locust. Insect Biochem. Mol. Biol. 2014, 52, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Yao, Y.; Wang, B. Methoprene-tolerant (Met) and Krüpple-homologue 1 (Kr-h1) are required for ovariole development and egg maturation in the brown plant hopper. Sci. Rep. 2016, 5, 18064. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Xu, Y.; Lin, X. Role of Broad-Complex (Br) and Krüppel homolog 1 (Krh1) in the ovary development of Nilaparvata lugens. Front. Physiol. 2017, 8, 1013. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; He, H.; Qu, X.; Cai, Y.; Ding, W.; Qiu, L.; Li, Y. RNA interference-mediated knockdown of the transcription factor Krüppel homologue 1 suppresses vitellogenesis in Chilo suppressalis. Insect Mol. Biol. 2000, 29, 183–192. [Google Scholar] [CrossRef]
- Yue, Y.; Yang, R.L.; Wang, W.P.; Zhou, Q.H.; Chen, E.H.; Yuan, G.R.; Wang, J.J.; Dou, W. Involvement of Met and Kr-h1 in JH-mediated reproduction of female Bactrocera dorsalis (Hendel). Front. Physiol. 2018, 9, 482. [Google Scholar] [CrossRef]
- Zhang, W.N.; Ma, L.; Liu, C.; Chen, L.; Xiao, H.J.; Liang, G.M. Dissecting the role of Krüppel homolog 1 in the metamorphosis and female reproduction of the cotton bollworm, Helicoverpa armigera. Insect Mol. Biol. 2018, 27, 492–504. [Google Scholar] [CrossRef]
- Konopova, B.; Smykal, V.; Jindra, M. Common and distinct roles of juvenile hormone signaling genes in metamorphosis of holometabolous and hemimetabolous insects. PLoS ONE 2011, 6, e28728. [Google Scholar] [CrossRef]
- Villalobos-Sambucaro, M.J.; Riccillo, F.L.; Calderón-Fernández, G.M.; Sterkel, M.; Diambra, L.A.; Ronderos, J.R. Genomic and functional characterization of a methoprene-tolerant gene in the kissing-bug Rhodnius prolixus. Gen. Comp. Endocrinol. 2015, 216, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Melo, A.C.; Valle, D.; Machado, E.A.; Salerno, A.P.; Paiva-Silva, G.O.; Cunha, E.; Silva, N.L.; de Souza, W.; Masuda, H. Synthesis of vitellogenin by the follicle cells of Rhodnius prolixus. Insect Biochem. Mol. Biol. 2000, 30, 549–557. [Google Scholar] [CrossRef]
- Lange, A.B.; Leyria, J.; Orchard, I. The hormonal and neural control of egg production in the historically important model insect, Rhodnius prolixus: A review, with new insights in this post-genomic era. Gen. Comp. Endocrinol. 2022, 321–322, 114030. [Google Scholar] [CrossRef]
- Eberhard, F.E.; Cunze, S.; Kochmann, J.; Klimpel, S. Modelling the climatic suitability of Chagas disease vectors on a global scale. eLife 2020, 9, e52072. [Google Scholar] [CrossRef] [PubMed]
- Minakuchi, C.; Namiki, T.; Shinoda, T. Krüppel homolog 1, an early juvenile hormone-response gene downstream of Methoprene-tolerant, mediates its anti-metamorphic action in the red flour beetle Tribolium castaneum. Dev. Biol. 2009, 325, 341–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fruttero, L.L.; Leyria, J.; Canavoso, L.E. Lipids in Insect Oocytes: From the storage pathways to their multiple functions. Results Probl. Cell Differ. 2017, 63, 403–434. [Google Scholar]
- Noriega, F.G. Juvenile Hormone biosynthesis in insects: What Is New, What Do We Know, and What Questions Remain? Int. Sch. Res. Not. 2014, 967361. [Google Scholar] [CrossRef] [Green Version]
- Atella, G.C.; Gondim, K.C.; Machado, E.A.; Medeiros, M.N.; Silva-Neto, M.A.; Masuda, H. Oogenesis and egg development in triatomines: A biochemical approach. An.Acad. Bras. Cienc. 2005, 77, 405–430. [Google Scholar] [CrossRef] [Green Version]
- Ramos, F.O.; Nouzova, M.; Fruttero, L.L.; Leyria, J.; Ligabue-Braun, R.; Noriega, F.G.; Canavoso, L. E Role of Methoprene-tolerant in the regulation of oogenesis in Dipetalogaster maxima. Sci. Rep. 2022, 12, 14195. [Google Scholar] [CrossRef]
- Tufail, M.; Takeda, M. Insect vitellogenin/lipophorin receptors: Molecular structures, role in oogenesis, and regulatory mechanisms. J. Insect Physiol. 2009, 55, 87–103. [Google Scholar] [CrossRef]
- Jing, Y.P.; Wen, X.; Li, L.; Zhang, S.; Zhang, C.; Zhou, S. The vitellogenin receptor functionality of the migratory locust depends on its phosphorylation by juvenile hormone. Proc. Natl. Acad. Sci. USA 2021, 118, e2106908118. [Google Scholar] [CrossRef] [PubMed]
- Bagamasbad, P.; Denver, R.J. Mechanisms and significance of nuclear receptor auto- and cross-regulation. Gen. Comp. Endocrinol. 2011, 170, 3–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gijbels, M.; Lenaerts, C.; Vanden Broeck, J.; Marchal, E. Juvenile Hormone receptor Met is essential for ovarian maturation in the Desert Locust, Schistocerca gregaria. Sci. Rep. 2019, 9, 10797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchal, E.; Hult, E.F.; Huang, J.; Pang, Z.; Stay, B.; Tobe, S.S. Methoprene-tolerant (Met) knockdown in the adult female cockroach, Diploptera punctata completely inhibits ovarian development. PLoS ONE 2014, 9, e106737. [Google Scholar] [CrossRef] [Green Version]
- Naghdi, M.; Maestro, J.L.; Belles, X.; Bandani, A. Transduction of the vitellogenic signal of juvenile hormone by Methoprene-tolerant in the cockroach Blattella germanica (L.) (Dictyoptera, Blattellidae). Arthropods 2016, 5, 130–137. [Google Scholar]
- Parthasarathy, R.; Sun, Z.; Bai, H.; Palli, S.R. Juvenile hormone regulation of vitellogenin synthesis in the red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 2010, 40, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Zhang, W.; Liu, C.; Chen, L.; Xu, Y.; Xiao, H.; Liang, G. Methoprene-Tolerant (Met) is indispensable for larval metamorphosis and female reproduction in the cotton bollworm Helicoverpa armigera. Front. Physiol. 2018, 9, 1601. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Fu, X.; Zhu, J. Juvenile hormone-regulated alternative splicing of the taiman gene primes the ecdysteroid response in adult mosquitoes. Proc. Natl. Acad. Sci. USA 2018, 115, E7738–E7747. [Google Scholar]
- Jindra, M.; Bellés, X.; Shinoda, T. Molecular basis of juvenile hormone signaling. Curr. Opin. Insect Sci. 2015, 11, 39–46. [Google Scholar] [CrossRef]
- Hahn, M.E. The aryl hydrocarbon receptor: A comparative perspective. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1998, 121, 23–53. [Google Scholar] [CrossRef]
- Smykal, V.; Bajgar, A.; Provaznik, J.; Fexova, S.; Buricova, M.; Hodkova, M.; Jindra, M.; Dolezel, D. Juvenile hormone signaling during reproduction and development of the linden bug, Pyrrhocoris apterus. Insect Biochem. Mol. Biol. 2014, 45, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Gujar, H.; Palli, S.R. Juvenile hormone regulation of female reproduction in the common bed bug, Cimex lectularius. Sci. Rep. 2016, 6, 35546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, J.; Uehara, Y.; Montell, D.J. Regulation of invasive cell behavior by taiman, a Drosophila protein related to AIB1, a steroid receptor coactivator amplified in breast cancer. Cell 2000, 103, 1047–1058. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, L.; Sun, G.; Raikhel, A.S. The competence factor beta Ftz-F1 potentiates ecdysone receptor activity via recruiting a p160/SRC coactivator. Mol. Cell Biol. 2006, 26, 9402–9412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozano, J.; Kayukawa, T.; Shinoda, T.; Belles, X. A role for Taiman in insect metamorphosis. PLoS Genet. 2014, 10, e1004769. [Google Scholar] [CrossRef]
- Noriega, F.G.; Nouzova, M. Approaches and Tools to Study the Roles of Juvenile Hormones in Controlling Insect Biology. Insects 2020, 11, 858. [Google Scholar] [CrossRef]
- Nouzova, M.; Edwards, M.J.; Michalkova, V.; Ramirez, C.E.; Ruiz, M.; Areiza, M.; DeGennaro, M.; Fernandez-Lima, F.; Feyereisen, R.; Jindra, M.; et al. Epoxidation of juvenile hormone was a key innovation improving insect reproductive fitness. Proc. Natl. Acad. Sci. USA 2021, 118, e2109381118. [Google Scholar] [CrossRef]
- Hejnikova, M.; Paroulek, M.; Hodkova, M. Decrease in Methoprene tolerant and Taiman expression reduces juvenile hormone effects and enhances the levels of juvenile hormone circulating in males of the linden bug Pyrrhocoris apterus. J. Insect Physiol. 2016, 93-94, 72–80. [Google Scholar] [CrossRef]
- Chiang, A.S.; Schal, C. Correlation among corpus allatum volume, cell size, and juvenile hormone biosynthesis in ovariectomized adult Blattella germanica. Arch. Insect Biochem. Physiol. 1991, 18, 37–44. [Google Scholar] [CrossRef]
- Liu, P.; Peng, H.J.; Zhu, J. Juvenile hormone-activated phospholipase C pathway enhances transcriptional activation by the methoprene-tolerant protein. Proc. Natl. Acad. Sci. USA 2015, 112, E1871–E1879. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Zhang, Y. Kr-h1, a cornerstone gene in insect life history. Front. Physiol. 2022, 13, 905441. [Google Scholar] [CrossRef] [PubMed]
- Avilés-Pagán, E.E.; Orr-Weaver, T.L. Activating embryonic development in Drosophila. Semin Cell Dev. Biol. 2018, 84, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Sedra, L.; Paluzzi, J.P.; Lange, A.B. Characterization and expression of a long neuropeptide F (NPF) receptor in the Chagas disease vector Rhodnius prolixus. PLoS ONE 2018, 13, e0202425. [Google Scholar] [CrossRef]
- Fernandez-Nicolas, A.; Belles, X. Juvenile hormone signaling in short germ-band hemimetabolan embryos. Development 2017, 144, 4637–4644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orchard, I.; Leyria, J.; Al-Dailami, A.; Lange, A.B. Fluid Secretion by Malpighian Tubules of Rhodnius prolixus: Neuroendocrine Control With New Insights From a Transcriptome Analysis. Front. Endocrinol. 2021, 12, 722487. [Google Scholar] [CrossRef] [PubMed]
- Leyria, J.; Orchard, I.; Lange, A.B. The involvement of insulin/ToR signaling pathway in reproductive performance of Rhodnius prolixus. Insect Biochem. Mol. Biol. 2021, 130, 103526. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genet-ics Analysis across Computing Platforms. Mo.l Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quanti-tative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ramos, F.O.; Leyria, J.; Nouzova, M.; Fruttero, L.L.; Noriega, F.G.; Canavoso, L.E. Juvenile hormone mediates lipid storage in the oocytes of Dipetalogaster maxima. Insect Biochem. Mol. Biol. 2021, 133, 103499. [Google Scholar] [CrossRef] [PubMed]
- Comas, D.; Piulachs, M.D.; Bellés, X. Induction of vitellogenin gene transcription in vitro by juvenile hormone in Blattella germanica. Mol. Cell Endocrinol. 2001, 183, 93–100. [Google Scholar] [CrossRef]
- Leyria, J.; El-Mawed, H.; Orchard, I.; Lange, A.B. Regulation of a Trehalose-Specific Facilitated Transporter (TRET) by Insulin and Adipokinetic Hormone in Rhodnius prolixus, a Vector of Chagas Disease. Front. Physiol. 2021, 12, 624165. [Google Scholar] [CrossRef] [PubMed]
- Leyria, J.; Orchard, I.; Lange, A.B. What happens after a blood meal? A transcriptome analysis of the main tissues involved in egg production in Rhodnius prolixus, an insect vector of Chagas disease. PLoS Negl. Trop Dis. 2020, 14, e0008516. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, S.A.; Frede, S.; Rubiolo, E.R.; Canavoso, L.E. Vitellogenesis in the hematophagous Dipetalogaster maxima (Hemiptera: Reduviidae), a vector of Chagas’ disease. J. Insect Physiol. 2008, 54, 393–402. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leyria, J.; Orchard, I.; Lange, A.B. Impact of JH Signaling on Reproductive Physiology of the Classical Insect Model, Rhodnius prolixus. Int. J. Mol. Sci. 2022, 23, 13832. https://doi.org/10.3390/ijms232213832
Leyria J, Orchard I, Lange AB. Impact of JH Signaling on Reproductive Physiology of the Classical Insect Model, Rhodnius prolixus. International Journal of Molecular Sciences. 2022; 23(22):13832. https://doi.org/10.3390/ijms232213832
Chicago/Turabian StyleLeyria, Jimena, Ian Orchard, and Angela B. Lange. 2022. "Impact of JH Signaling on Reproductive Physiology of the Classical Insect Model, Rhodnius prolixus" International Journal of Molecular Sciences 23, no. 22: 13832. https://doi.org/10.3390/ijms232213832
APA StyleLeyria, J., Orchard, I., & Lange, A. B. (2022). Impact of JH Signaling on Reproductive Physiology of the Classical Insect Model, Rhodnius prolixus. International Journal of Molecular Sciences, 23(22), 13832. https://doi.org/10.3390/ijms232213832