
The Ros/MucR Zinc-Finger Protein Family in Bacteria: Structure and Functions

Abstract
:1. Introduction
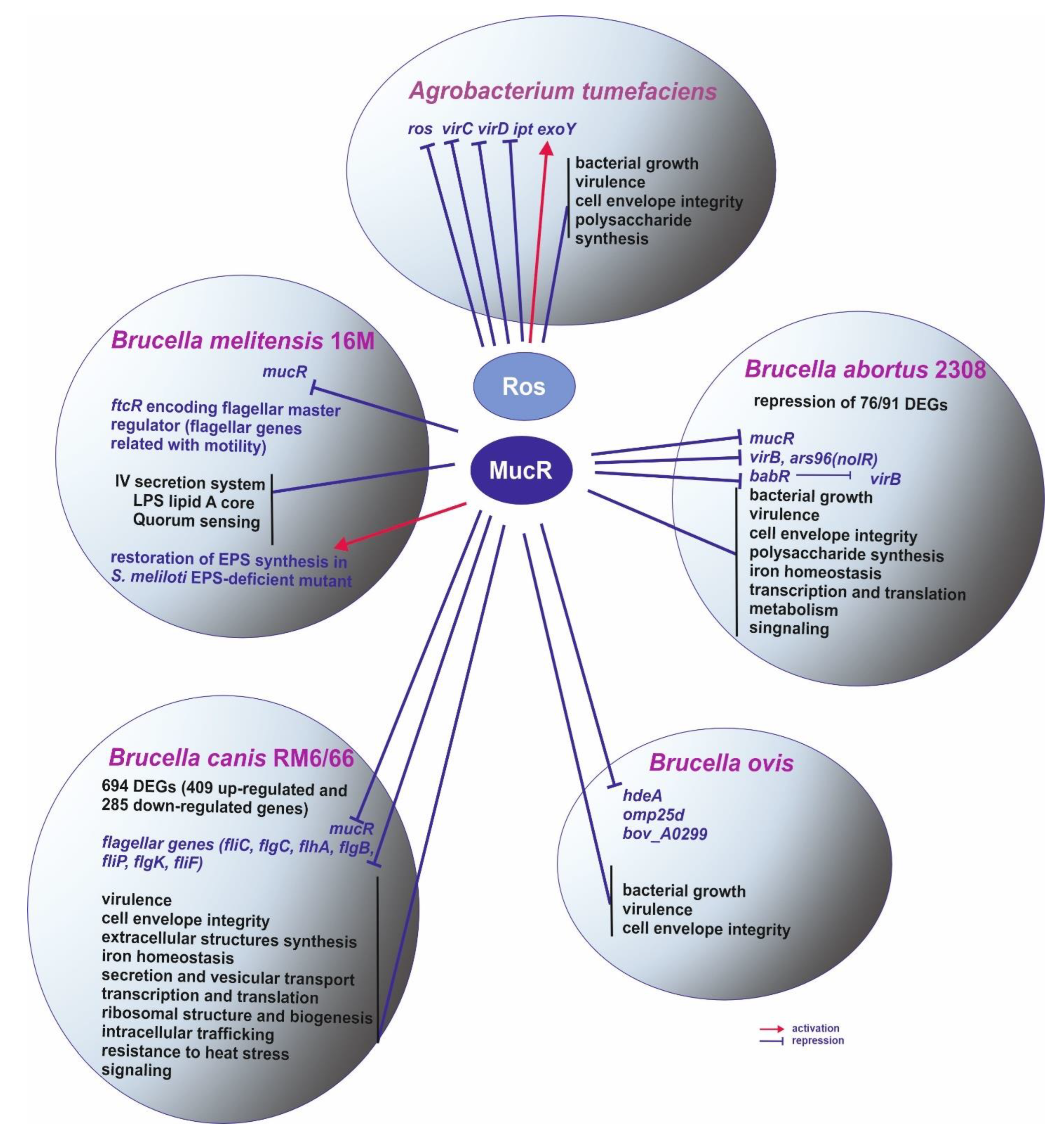
2. Pathogenic Bacteria
2.1. Agrobacterium Tumefaciens
2.2. Brucella spp.
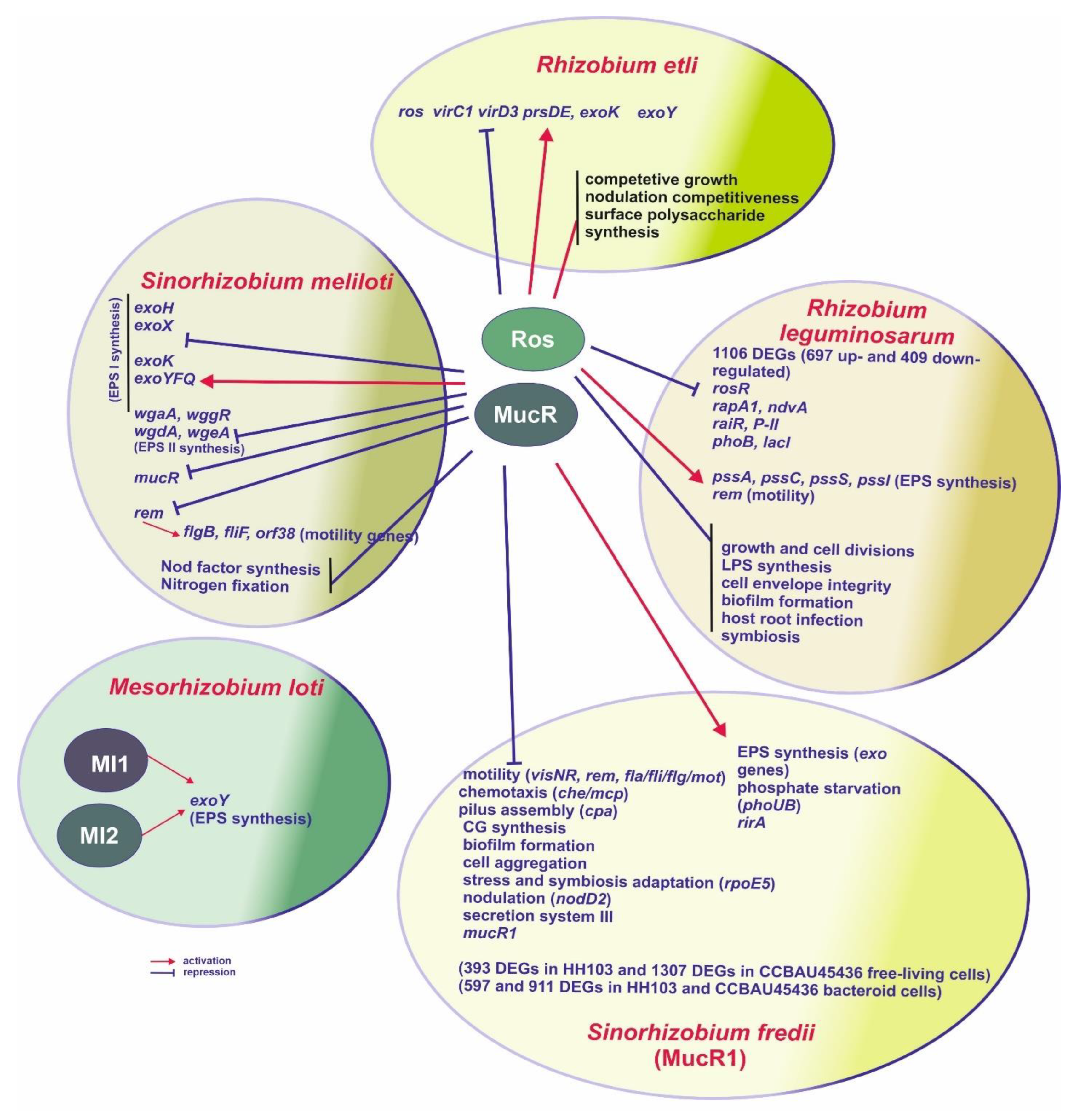
3. Symbiotic Bacteria
3.1. Sinorhizobium Meliloti
3.2. Rhizobium Etli and Rhizobium Leguminosarum
3.3. Mesorhizobium Loti
3.4. Sinorhizobium Fredii
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| aa | amino acids |
| H | histidine |
| C | cysteine |
| A | adenine |
| T | thymine |
| ZF | zinc-finger |
| C2H2 | Cys2His2 |
| ORF | open reading frame |
| IR | inverted repeats |
| CRP | catabolite regulatory protein |
| EPS | exopolysaccharide |
| LPS | lipopolysaccharide |
| CG | cyclic glucans |
| H-NS | Histone–like Nucleoid–Structuring |
| RNAP | RNA polymerase |
| UP | upstream promoter |
| COG | clusters of orthologous genes |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| DEG | differentially expressed genes |
| T3SS | type three secretion system |
References
- Close, T.J.; Tait, R.C.; Kado, C.I. Regulation of Ti plasmid virulence genes by a chromosomal locus of Agrobacterium tumefaciens. J. Bacteriol. 1985, 164, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.; Roxlau, A.; Weng, W.M.; Schmidt, M.; Quandt, J.; Niehaus, K.; Jording, D.; Arnold, W.; Puhler, A. Molecular analysis of the Rhizobium meliloti mucR gene regulating the biosynthesis of the exopolysaccharides succinoglycan and galactoglucan. Mol. Plant Microbe Interact. 1995, 8, 267–277. [Google Scholar] [CrossRef]
- Janczarek, M.; Skorupska, A. The Rhizobium leguminosarum bv. trifolii RosR: Transcriptional regulator involved in exopolysaccharide production. Mol. Plant Microbe Interact. 2007, 20, 867–881. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Jurado, S.; Alias-Villegas, C.; Navarro-Gómez, P.; Zehner, S.; Murdoch, P.D.; Rodríguez-Carvajal, M.A.; Soto, M.J.; Ollero, F.-J.; Ruiz-Sainz, J.E.; Göttfert, M.; et al. The Sinorhizobium fredii HH103 MucR1 Global Regulator Is Connected with the nod Regulon and Is Required for Efficient Symbiosis With Lotus burttii and Glycine max cv. Williams. Mol. Plant Microbe Interact. 2016, 29, 700–712. [Google Scholar] [CrossRef] [Green Version]
- Jiao, J.; Wu, L.J.; Zhang, B.; Hu, Y.; Li, Y.; Zhang, X.X.; Guo, H.J.; Liu, L.X.; Chen, W.X.; Zhang, Z.; et al. MucR Is Required for Transcriptional Activation of Conserved Ion Transporters to Support Nitrogen Fixation of Sinorhizobium fredii in Soybean Nodules. Mol. Plant Microbe Interact. 2016, 29, 352–361. [Google Scholar] [CrossRef] [Green Version]
- Pirone, L.; Pitzer, J.E.; D’Abrosca, G.; Fattorusso, R.; Malgieri, G.; Pedone, E.M.; Pedone, P.V.; Ii, R.M.R.; Baglivo, I. Identifying the region responsible for Brucella abortus MucR higher-order oligomer formation and examining its role in gene regulation. Sci. Rep. 2018, 8, 17238. [Google Scholar] [CrossRef] [Green Version]
- Laity, J.H.; Lee, B.M.; Wright, P.E. Zinc finger proteins: New insights into structural and functional diversity. Curr. Opin. Struct. Biol. 2001, 11, 39–46. [Google Scholar] [CrossRef]
- Gamsjaeger, R.; Liew, C.K.; Loughlin, F.E.; Crossley, M.; Mackay, J.P. Sticky fingers: Zinc-fingers as protein-recognition motifs. Trends Biochem. Sci. 2007, 32, 63–70. [Google Scholar] [CrossRef]
- Chou, A.Y.; Archdeacon, J.; Kado, C.I. Agrobacterium transcriptional regulator Ros is a prokaryotic zinc finger protein that regulates the plant oncogene ipt. Proc. Natl. Acad. Sci. USA 1998, 95, 5293–5298. [Google Scholar] [CrossRef] [Green Version]
- Isernia, C.; Malgieri, G.; Russo, L.; D’Abrosca, G.; Baglivo, I.; Pedone, P.V.; Fattorusso, R. 12. Zinc Fingers. In Transition Metals and Sulfur—A Strong Relationship for Life; De Gruyter: Boston, MA, USA, 2020; pp. 415–436. [Google Scholar] [CrossRef]
- Palmieri, M.; Malgieri, G.; Russo, L.; Baglivo, I.; Esposito, S.; Netti, F.; Del Gatto, A.; de Paola, I.; Zaccaro, L.; Pedone, P.V.; et al. Structural Zn(II) implies a switch from fully cooperative to partly downhill folding in highly homologous proteins. J. Am. Chem. Soc. 2013, 135, 5220–5228. [Google Scholar] [CrossRef]
- Kado, C.I. Origin and evolution of plasmids. Antonie Van Leeuwenhoek 1998, 73, 117–126. [Google Scholar] [CrossRef]
- D’Souza-Ault, M.R.; Cooley, M.B.; Kado, C.I. Analysis of the Ros repressor of Agrobacterium virC and virD operons: Molecular intercommunication between plasmid and chromosomal genes. J. Bacteriol. 1993, 175, 3486–3490. [Google Scholar] [CrossRef] [Green Version]
- Kado, C.I. Negative transcriptional regulation of virulence and oncogenes of the Ti plasmid by Ros bearing a conserved C2H2-zinc finger motif. Plasmid 2002, 48, 179–185. [Google Scholar] [CrossRef]
- Cooley, M.B.; D’Souza, M.R.; Kado, C.I. The virC and virD operons of the Agrobacterium Ti plasmid are regulated by the ros chromosomal gene: Analysis of the cloned ros gene. J. Bacteriol. 1991, 173, 2608–2616. [Google Scholar] [CrossRef] [Green Version]
- Close, T.J.; Rogowsky, P.M.; Kado, C.I.; Winans, S.C.; Yanofsky, M.F.; Nester, E.W. Dual control of Agrobacterium tumefaciens Ti plasmid virulence genes. J. Bacteriol. 1987, 169, 5113–5118. [Google Scholar] [CrossRef] [Green Version]
- Tiburtius, A.; de Luca, N.G.; Hussain, H.; Johnston, A.W. Expression of the exoY gene, required for exopolysaccharide synthesis in Agrobacterium, is activated by the regulatory ros gene. Microbiology 1996, 142, 2621–2629. [Google Scholar] [CrossRef] [Green Version]
- Cooley, M.B.; Kado, C.I. Mapping of the ros virulence regulatory gene of Agrobacterium tumefaciens. Mol. Gen. Genet. 1991, 230, 24–27. [Google Scholar] [CrossRef]
- Esposito, S.; Baglivo, I.; Malgieri, G.; Russo, L.; Zaccaro, L.; D’Andrea, L.D.; Mammucari, M.; Di Blasio, B.; Isernia, C.; Fattorusso, R.; et al. A novel type of zinc finger DNA binding domain in the Agrobacterium tumefaciens transcriptional regulator Ros. Biochemistry 2006, 45, 10394–10405. [Google Scholar] [CrossRef]
- Malgieri, G.; Russo, L.; Esposito, S.; Baglivo, I.; Zaccaro, L.; Pedone, E.M.; Di Blasio, B.; Isernia, C.; Pedone, P.V.; Fattorusso, R. The prokaryotic Cys2His2 zinc-finger adopts a novel fold as revealed by the NMR structure of Agrobacterium tumefaciens Ros DNA-binding domain. Proc. Natl. Acad. Sci. USA 2007, 104, 17341–17346. [Google Scholar] [CrossRef]
- Netti, F.; Malgieri, G.; Esposito, S.; Palmieri, M.; Baglivo, I.; Isernia, C.; Omichinski, J.G.; Pedone, P.V.; Lartillot, N.; Fattorusso, R. An experimentally tested scenario for the structural evolution of eukaryotic Cys2His2 zinc fingers from eubacterial ros homologs. Mol. Biol. Evol. 2013, 30, 1504–1513. [Google Scholar] [CrossRef] [Green Version]
- Baglivo, I.; Pirone, L.; Pedone, E.M.; Pitzer, J.E.; Muscariello, L.; Marino, M.M.; Malgieri, G.; Freschi, A.; Chambery, A.; Ii, R.-M.R.; et al. Ml proteins from Mesorhizobium loti and MucR from Brucella abortus: An AT-rich core DNA-target site and oligomerization ability. Sci. Rep. 2017, 7, 15805. [Google Scholar] [CrossRef] [Green Version]
- Grainger, D.C. Structure and function of bacterial H-NS protein. Biochem. Soc. Trans. 2016, 44, 1561–1569. [Google Scholar] [CrossRef]
- D’Abrosca, G.; Paladino, A.; Baglivo, I.; Russo, L.; Sassano, M.; Grazioso, R.; Iacovino, R.; Pirone, L.; Pedone, E.M.; Pedone, P.V.; et al. Structural insight of the full-length Ros protein: A prototype of the prokaryotic zinc-finger family. Sci. Rep. 2020, 10, 9283. [Google Scholar] [CrossRef]
- Grazioso, R.; García-Viñuales, S.; D’Abrosca, G.; Baglivo, I.; Pedone, P.V.; Milardi, D.; Fattorusso, R.; Isernia, C.; Russo, L.; Malgieri, G. The change of conditions does not affect Ros87 downhill folding mechanism. Sci. Rep. 2020, 10, 21067. [Google Scholar] [CrossRef]
- Moreno, E.; Stackebrandt, E.; Dorsch, M.; Wolters, J.; Busch, M.; Mayer, H. Brucella abortus 16S rRNA and lipid A reveal a phylogenetic relationship with members of the alpha-2 subdivision of the class Proteobacteria. J. Bacteriol. 1990, 172, 3569–3576. [Google Scholar] [CrossRef] [Green Version]
- Pappas, G.; Papadimitriou, P.; Akritidis, N.; Christou, L.; Tsianos, E.V. The new global map of human brucellosis. Lancet Infect. Dis. 2006, 6, 91–99. [Google Scholar] [CrossRef]
- Mirabella, A.; Terwagne, M.; Zygmunt, M.S.; Cloeckaert, A.; De Bolle, X.; Letesson, J.J. Brucella melitensis MucR, an orthologue of Sinorhizobium meliloti MucR, is involved in resistance to oxidative, detergent, and saline stresses and cell envelope modifications. J. Bacteriol. 2013, 195, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Pei, J.; Turse, C.; Ficht, T.A. Mariner mutagenesis of Brucella melitensis reveals genes with previously uncharacterized roles in virulence and survival. BMC Microbiol. 2006, 6, 102. [Google Scholar] [CrossRef] [Green Version]
- Mirabella, A.; Yañez Villanueva, R.M.; Delrue, R.M.; Uzureau, S.; Zygmunt, M.S.; Cloeckaert, A.; De Bolle, X.; Letesson, J.J. The two-component system PrlS/PrlR of Brucella melitensis is required for persistence in mice and appears to respond to ionic strength. Microbiology 2012, 158, 2642–2651. [Google Scholar] [CrossRef]
- Dong, H.; Liu, W.; Peng, X.; Jing, Z.; Wu, Q. The effects of MucR on expression of type IV secretion system, quorum sensing system and stress responses in Brucella melitensis. Vet. Microbiol. 2013, 166, 535–542. [Google Scholar] [CrossRef]
- Caswell, C.C.; Elhassanny, A.E.M.; Planchin, E.E.; Roux, C.M.; Weeks-Gorospe, J.N.; Ficht, T.A.; Dunman, P.M.; Roop, R.M. Diverse genetic regulon of the virulence-associated transcriptional regulator MucR in Brucella abortus 2308. Infect. Immun. 2013, 81, 1040–1051. [Google Scholar] [CrossRef] [Green Version]
- Borriello, G.; Russo, V.; Paradiso, R.; Riccardi, M.G.; Criscuolo, D.; Verde, G.; Marasco, R.; Pedone, P.V.; Galiero, G.; Baglivo, I. Different impacts of MucR binding to the babR and virB promoters on gene expression in Brucella abortus 2308. Biomolecules 2020, 10, 788. [Google Scholar] [CrossRef] [PubMed]
- Baglivo, I.; Pirone, L.; Malgieri, G.; Fattorusso, R.; Roop, R.M.; Marasco, R.; Pedone, P.V.; Galiero, G.; Baglivo, I. MucR binds multiple target sites in the promoter of its own gene and is a heat-stable protein: Is MucR a H-NS-like protein? FEBS Open Biol. 2018, 8, 711–718. [Google Scholar] [CrossRef] [Green Version]
- Castang, S.; Dove, S.L. High-order oligomerization is required for the function of the H-NS family member MvaT in Pseudomonas aeruginosa. Mol. Microbiol. 2010, 78, 916–931. [Google Scholar] [CrossRef] [Green Version]
- Gordon, B.R.; Li, Y.; Wang, L.; Sintsova, A.; van Bakel, H.; Tian, S.; Navarre, W.W.; Xia, B.; Liu, J. Lsr2 is a nucleoid-associated protein that targets AT-rich sequences and virulence genes in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2010, 107, 5154–5159. [Google Scholar] [CrossRef] [Green Version]
- Navarre, W.W.; McClelland, M.; Libby, S.J.; Fang, F.C. Silencing of xenogeneic DNA by H-NS–Facilitation of lateral gene transfer in bacteria by a defense system that recognizes foreign DNA. Genes Dev. 2007, 21, 1456–1471. [Google Scholar] [CrossRef] [Green Version]
- Ding, P.; McFarland, K.A.; Jin, S.; Tong, G.; Duan, B.; Yang, A.; Hughes, T.R.; Liu, J.; Dove, S.L.; Navarre, W.W.; et al. A Novel AT-Rich DNA Recognition Mechanism for Bacterial Xenogeneic Silencer MvaT. PLoS Pathog. 2015, 11, e1004967. [Google Scholar] [CrossRef]
- Will, W.R.; Navarre, W.W.; Fang, F.C. Integrated circuits: How transcriptional silencing and counter-silencing facilitate bacterial evolution. Curr. Opin. Microbiol. 2015, 23, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Ulissi, U.; Fabbretti, A.; Sette, M.; Giuliodori, A.M.; Spurio, R. Time-resolved assembly of a nucleoprotein complex between Shigella flexneri virF promoter and its transcriptional repressor H-NS. Nucleic Acids Res. 2014, 42, 13039–13050. [Google Scholar] [CrossRef]
- Sun, J.; Dong, H.; Peng, X.; Liu, Y.; Jiang, H.; Feng, Y. Deletion of the Transcriptional Regulator MucR in Brucella canis Affects Stress Responses and Bacterial Virulence. Front. Vet. Sci. 2021, 8, 650942. [Google Scholar] [CrossRef]
- Tartilán-Choya, B.; Sidhu-Muñoz, R.S.; Vizcaíno, N. The Transcriptional Regulator MucR, but Not Its Controlled Acid-Activated Chaperone HdeA, Is Essential for Virulence and Modulates Surface Architecture and Properties in Brucella ovis PA. Front. Vet. Sci. 2022, 8, 814752. [Google Scholar] [CrossRef] [PubMed]
- Fumeaux, C.; Radhakrishnan, S.K.; Ardissone, S.; Théraulaz, L.; Frandi, A.; Martins, D.; Nesper, J.; Abel, S.; Jenal, U.; Viollier, P.H. Cell cycle transition from S-phase to G1 in Caulobacter is mediated by ancestral virulence regulators. Nat. Commun. 2014, 5, 4081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.T.; Chen, W.F.; Tian, C.F.; Young, J.P.W.; Chen, W.X. Ecology and evolution of Rhizobia. Principles and Applications; Springer Nature Singapore Pte Ltd.: Singapore, 2019; ISBN 978-981-32-9555-1. [Google Scholar] [CrossRef]
- Mendoza-Suárez, M.; Andersen, S.U.; Poole, P.S.; Sánchez-Cañizares, C. Competition, Nodule Occupancy, and Persistence of Inoculant Strains: Key Factors in the Rhizobium-Legume Symbioses. Front. Plant Sci. 2021, 12, 690567. [Google Scholar] [CrossRef] [PubMed]
- López-Baena, F.J.; Ruiz-Sainz, J.E.; Rodríguez-Carvajal, M.A.; Vinardell, J.M. Bacterial Molecular Signals in the Sinorhizobium fredii-Soybean Symbiosis. Int. J. Mol. Sci. 2016, 17, 755. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Jurado, S.; Fuentes-Romero, F.; Ruiz-Sainz, J.-E.; Janczarek, M.; Vinardell, J.-M. Rhizobial exopolysaccharides: Genetic regulation of their synthesis and relevance in symbiosis with Legumes. Int. J. Mol. Sci. 2021, 22, 6233. [Google Scholar] [CrossRef] [PubMed]
- Janczarek, M.; Rachwał, K.; Marzec, A.; Grządziel, J.; Palusińska-Szysz, M. Signal molecules and cell-surface components involved in early stages of the legume–rhizobium interactions. Appl. Soil Ecol. 2015, 85, 94–113. [Google Scholar] [CrossRef]
- Downie, J.A. The roles of extracellular proteins, polysaccharides and signals in the interactions of rhizobia with legume roots. FEMS Microbiol. Rev. 2010, 34, 150–170. [Google Scholar] [CrossRef]
- Bertram-Drogatz, P.A.; Rüberg, S.; Becker, A.; Pühler, A. The regulatory protein MucR binds to a short DNA region located upstream of the mucR coding region in Rhizobium meliloti. Mol. Gen. Genet. 1997, 254, 529–538. [Google Scholar] [CrossRef]
- Bertram-Drogatz, P.A.; Quester, I.; Becker, A.; Pühler, A. The Sinorhizobium meliloti MucR protein, which is essential for the production of high-molecular-weight succinoglycan exopolysaccharide, binds to short DNA regions upstream of exoH and exoY. Mol. Gen. Genet. 1998, 257, 433–441. [Google Scholar] [CrossRef]
- Janczarek, M. Environmental signals and regulatory pathways that influence exopolysaccharide production in rhizobia. Int. J. Mol. Sci. 2011, 12, 7898–7933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahlawane, C.; Baumgarth, B.; Serrania, J.; Rüberg, S.; Becker, A. Fine-tuning of galactoglucan biosynthesis in Sinorhizobium meliloti by differential WggR (ExpG)-, PhoB-, and MucR-dependent regulation of two promoters. J. Bacteriol. 2008, 190, 3456–3466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, K.; González, J.E. Complex regulation of symbiotic functions is coordinated by MucR and quorum sensing in Sinorhizobium meliloti. J. Bacteriol. 2011, 193, 485–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloret, J.; Martín, M.; Oruezabal, R.I.; Bonilla, I.; Rivilla, R. MucR and mucS activate exp genes transcription and galactoglucan production in Sinorhizobium meliloti EFB1. Mol. Plant Microbe Interact. 2002, 15, 54–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahlawane, C.; McIntosh, M.; Krol, E.; Becker, A. Sinorhizobium meliloti regulator MucR couples exopolysaccharide synthesis and motility. Mol. Plant Microbe Interact. 2008, 21, 1498–1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittinger, M.A.; Milner, J.L.; Saville, B.J.; Handelsman, J. rosR, a determinant of nodulation competitiveness in Rhizobium etli. Mol. Plant Microbe Interact. 1997, 10, 180–186. [Google Scholar] [CrossRef] [Green Version]
- Araujo, R.S.; Robleto, E.A.; Handelsman, J. A hydrophobic mutant of Rhizobium etli altered in nodulation competitiveness and growth in the rhizosphere. Appl. Environ. Microbiol. 1994, 60, 1430–1436. [Google Scholar] [CrossRef] [Green Version]
- Bittinger, M.A.; Handelsman, J. Identification of genes in the RosR regulon of Rhizobium etli. J. Bacteriol. 2000, 182, 1706–1713. [Google Scholar] [CrossRef]
- Downie, J.A. Legume nodulation. Curr. Biol. 2014, 24, R184–R190. [Google Scholar] [CrossRef] [Green Version]
- Kozieł, M.; Kalita, M.; Janczarek, M. Genetic diversity of microsymbionts nodulating Trifolium pratense in subpolar and temperate climate regions. Sci. Rep. 2022, 12, 12144. [Google Scholar] [CrossRef]
- Janczarek, M.; Kalita, M.; Skorupska, A. New taxonomic markers for identification of Rhizobium leguminosarum and discrimination between closely related species. Arch. Microbiol. 2009, 191, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Janczarek, M.; Urbanik-Sypniewska, T. Expression of the Rhizobium leguminosarum bv. trifolii pssA gene involved in exopolysaccharide synthesis is regulated by RosR, phosphate and the carbon source. J. Bacteriol. 2013, 195, 3412–3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janczarek, M.; Rachwał, K. Mutation in the pssA gene involved in exopolysaccharide synthesis leads to several physiological and symbiotic defects in Rhizobium leguminosarum bv. trifolii. Int. J. Mol. Sci. 2013, 14, 3711–3735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janczarek, M.; Skorupska, A. Rhizobium leguminosarum bv. trifolii rosR gene expression is regulated by catabolic repression. FEMS Microbiol. Lett. 2009, 291, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Janczarek, M.; Skorupska, A. Modulation of rosR expression and exopolysaccharide production in Rhizobium leguminosarum bv. trifolii by phosphate and clover root exudates. Int. J. Mol. Sci. 2011, 12, 4132–4155. [Google Scholar] [CrossRef] [Green Version]
- Rachwał, K.; Lipa, P.; Wojda, I.; Vinardell, J.-M.; Janczarek, M. Regulatory elements located in the upstream region of the Rhizobium leguminosarum rosR global regulator are essential for its transcription and mRNA stability. Genes 2017, 8, 388. [Google Scholar] [CrossRef] [Green Version]
- Browning, D.F.; Busby, S.J. The regulation of bacterial transcription initiation. Nat. Rev. Microbiol. 2004, 2, 57–65. [Google Scholar] [CrossRef]
- Janczarek, M.; Skorupska, A. Regulation of pssA and pssB gene expression in R. leguminosarum bv. trifolii in response to environmental factors. Antonie Van Leeuwenhoek 2004, 85, 217–227. [Google Scholar] [CrossRef]
- Emory, S.A.; Bouvet, P.; Belasco, J.G. A 50 -terminal stem-loop structure can stabilize mRNA in Escherichia coli. Genes Dev. 1992, 6, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Rachwał, K.; Matczyńska, E.; Janczarek, M. Transcriptome profiling of a Rhizobium leguminosarum bv. trifolii rosR mutant reveals the role of the transcriptional regulator RosR in motility, synthesis of cell-surface components, and other cellular processes. BMC Genom. 2015, 16, 1111. [Google Scholar] [CrossRef] [Green Version]
- Rachwał, K.; Boguszewska, A.; Kopcińska, J.; Karaś, M.; Tchórzewski, M.; Janczarek, M. The regulatory protein RosR affects Rhizobium leguminosarum bv. trifolii protein profiles, cell surface properties, and symbiosis with clover. Front. Microbiol. 2016, 7, 1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janczarek, M.; Rachwał, K.; Kopcińska, J. Genetic characterization of the Pss region and the role of PssS in exopolysaccharide production and symbiosis of Rhizobium leguminosarum bv. trifolii with clover. Plant Soil 2015, 396, 257–275. [Google Scholar] [CrossRef] [Green Version]
- Janczarek, M.; Rachwal, K.; Turska-Szewczuk, A. A mutation in pssE affects exopolysaccharide synthesis by Rhizobium leguminosarum bv. trifolii, its surface properties, and symbiosis with clover. Plant Soil 2017, 417, 331–347. [Google Scholar] [CrossRef] [Green Version]
- Gottesman, S. Bacterial regulation: Global regulatory networks. Annu. Rev. Genet. 1984, 18, 415–444. [Google Scholar] [CrossRef]
- Janczarek, M.; Kutkowska, J.; Piersiak, T.; Skorupska, A. Rhizobium leguminosarum bv. trifolii rosR is required for interaction with clover, biofilm formation and adaptation to the environment. BMC Microbiol. 2010, 10, 284. [Google Scholar] [CrossRef] [Green Version]
- Cieśla, J.; Kopycińska, M.; Łukowska, M.; Bieganowski, A.; Janczarek, M. Surface properties of wild-type Rhizobium leguminosarum bv. trifolii strain 24.2 and its derivatives with different extracellular polysaccharide content. PLoS ONE 2016, 11, e0165080. [Google Scholar] [CrossRef] [Green Version]
- Lipa, M.; Janczarek, M. Phosphorylation systems in symbiotic nitrogen-fixing bacteria and their role in bacterial adaptation to various environmental stresses. Peer J. 2020, 8, e8466. [Google Scholar] [CrossRef] [Green Version]
- Janczarek, M.; Rachwał, K.; Cieśla, J.; Ginalska, G.; Bieganowski, J. Production of exopolysaccharide by Rhizobium leguminosarum bv. trifolii and its role in bacterial attachment and surface properties. Plant Soil 2015, 388, 211–227. [Google Scholar] [CrossRef]
- Janczarek, M.; Jaroszuk-Ściseł, J.; Skorupska, A. Multiple copies of rosR and pssA genes enhance exopolysaccharide production, symbiotic competitiveness and clover nodulation in Rhizobium leguminosarum bv. trifolii. Antonie Van Leeuwenhoek 2009, 96, 471–486. [Google Scholar] [CrossRef]
- Kopycińska, M.; Lipa, P.; Cieśla, J.; Kozieł, M.; Janczarek, M. Extracellular polysaccharide protects Rhizobium leguminosarum cells against zinc stress in vitro and during symbiosis with clover. Environ. Microbiol. Rep. 2018, 10, 355–368. [Google Scholar] [CrossRef]
- Jaszek, M.; Janczarek, M.; Kuczyński, K.; Piersiak, T.; Grzywnowicz, K. The response of the Rhizobium leguminosarum bv. trifolii wild-type and exopolysaccharide-deficient mutants to oxidative stress. Plant Soil 2014, 376, 75–94. [Google Scholar] [CrossRef] [Green Version]
- Baglivo, I.; Russo, L.; Esposito, S.; Malgieri, G.; Renda, M.; Salluzzo, A.; Di Blasio, B.; Isernia, C.; Fattorusso, R.; Pedone, P.V. The structural role of the zinc ion can be dispensable in prokaryotic zinc-finger domains. Proc. Natl. Acad. Sci. USA 2009, 106, 6933–6938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, L.; Palmieri, M.; Baglivo, I.; Esposito, L.; Isernia, C.; Malgieri, G.; Pedone, P.V.; Fattorusso, R. NMR assignments of the DNA binding domain of Ml4 protein from Mesorhizobium loti. Biomol. NMR Assign. 2010, 4, 55–57. [Google Scholar] [CrossRef] [PubMed]
- D’Abrosca, G.; Russo, L.; Palmieri, M.; Baglivo, I.; Netti, F.; de Paola, I.; Zaccaro, L.; Farina, B.; Iacovino, R.; Pedone, P.V.; et al. The (unusual) aspartic acid in the metal coordination sphere of the prokaryotic zinc finger domain. J. Inorg. Biochem. 2016, 161, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Jurado, S.; Navarro-Gómez, P.; Murdoch, P.; Crespo-Rivas, J.C.; Jie, S.; Cuesta-Berrio, L.; Ruiz-Sainz, J.E.; Rodríguez-Carvajal, M.Á.; Vinardell, J.M. Exopolysaccharide Production by Sinorhizobium fredii HH103 Is Repressed by Genistein in a NodD1-Dependent Manner. PLoS ONE 2016, 11, e0160499. [Google Scholar] [CrossRef] [Green Version]
- Vinardell, J.-M.; Acosta-Jurado, S.; Zehner, S.; Göttfert, M.; Becker, A.; Baena, I.; Blom, J.; Crespo-Rivas, J.C.; Goesmann, A.; Jaenicke, S.; et al. The Sinorhizobium fredii HH103 genome: A comparative analysis with S. fredii strains differing in their symbiotic behavior with soybean. Mol. Plant Microbe Interact. 2015, 28, 811–824. [Google Scholar] [CrossRef] [Green Version]
- Weidner, S.; Becker, A.; Bonilla, I.; Jaenicke, S.; Lloret, J.; Margaret, I.; Pühler, A.; Ruiz-Sainz, J.E.; Schneiker-Bekel, S.; Szczepanowski, R.; et al. Genome sequence of the soybean symbiont Sinorhizobium fredii HH103. J. Bacteriol. 2012, 194, 1617–1618. [Google Scholar] [CrossRef] [Green Version]
- Alías-Villegas, C.; Fuentes-Romero, F.; Cuéllar, V.; Navarro-Gómez, P.; Soto, M.J.; Vinardell, J.-M.; Sebastián Acosta-Jurado, S. Surface Motility Regulation of Sinorhizobium fredii HH103 by Plant Flavonoids and the NodD1, TtsI, NolR, and MucR1 Symbiotic Bacterial Regulators Int. J. Mol. Sci. 2022, 23, 7698. [Google Scholar] [CrossRef]
- Li, M.L.; Jiao, J.; Zhang, B.; Shi, W.T.; Yu, W.H.; Tian, C.F. Global transcriptional repression of diguanylate cyclases by MucR1 is essential for Sinorhizobium-soybean symbiosis. mBio 2021, 12, e0119221. [Google Scholar] [CrossRef]
- Jiménez-Guerrero, I.; Acosta-Jurado, S.; del Cerro, P.; Navarro-Gómez, P.; López-Baena, F.J.; Ollero, F.J.; Vinardell, J.M.; Pérez-Montaño, F. Transcriptomic studies of the effect of nod gene-inducing molecules in rhizobia: Different weapons, one purpose. Genes 2018, 9, 1. [Google Scholar] [CrossRef] [Green Version]
- Jiao, J.; Zhang, J.; Li, M.-L.; Zhang, Z.; Tian, C.-F. The zinc-finger bearing xenogeneic silencer MucR in α-proteobacteria balances adaptation and regulatory integrity. ISME J. 2022, 16, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Warman, E.A.; Singh, S.S.; Gubieda, A.G.; Grainger, D.C. A non-canonical promoter element drives spurious transcription of horizontally acquired bacterial genes. Nucleic Acids Res. 2020, 48, 4891–4901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, J.; Tian, C.-F. Ancestral zinc-finger bearing protein MucR in alpha-proteobacteria: A novel xenogeneic silencer? Comput. Struct. Biotechnol. J. 2020, 18, 3623–3631. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, B.; Jiao, J.; Dai, S.-Q.; Chen, W.-X.; Tian, C.-F. Modulation of symbiotic compatibility by rhizobial zinc starvation machinery. mBio 2020, 11, e03193-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Jiao, J.; Zhang, P.; Cui, W.J.; Zhang, Z.; Tian, C.F. Comparative analysis of core and accessory genes in coexpression network. Methods Mol. Biol. 2021, 2242, 45–58. [Google Scholar] [CrossRef]
- Doolittle, W.F.; Nesbo, C.L.; Bapteste, E.; Zhaxybayeva, O. Lateral gene transfer. In Evolutionary Genomics and Proteomics; Pagel, M., Pomiankowski, A., Eds.; Sinauer Associates Inc.: Sunderland, UK, 2008; pp. 45–80. [Google Scholar]
- Zhao, R.; Liu, L.X.; Zhang, Y.Z.; Jiao, J.; Cui, W.J.; Zhang, B.; Wang, X.L.; Li, M.L.; Chen, Y.; Xiong, Z.Q.; et al. Adaptive evolution of rhizobial symbiotic compatibility mediated by co-evolved insertion sequences. ISME J. 2018, 12, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Lucchini, S.; Rowley, G.; Goldberg, M.D.; Hurd, D.; Harrison, M.; Hinton, J.C.D. H-NS mediates the silencing of laterally acquired genes in bacteria. PLoS Pathog. 2006, 2, 746–752. [Google Scholar] [CrossRef]
- Piña-Iturbe, A.; Suazo, I.D.; Hoppe-Elsholz, G.; Ulloa-Allendes, D.; González, P.A.; Kalergis, A.M.; Bueno, S.M. Horizontally acquired homologs of xenogeneic silencers: Modulators of gene expression encoded by plasmids, phages and genomic islands. Genes 2020, 11, 142. [Google Scholar] [CrossRef]
- Lamberte, L.E.; Baniulyte, G.; Singh, S.S.; Stringer, A.M.; Bonocora, R.P.; Stracy, M.; Kapanidis, A.N.; Wade, J.T.; Grainger, D.C. Horizontally acquired AT-rich genes in Escherichia coli cause toxicity by sequestering RNA polymerase. Nat. Microbiol. 2017, 2, 16249. [Google Scholar] [CrossRef]
- Yun, C.S.; Suzuki, C.; Naito, K.; Takeda, T.; Takahashi, Y.; Sai, F.; Terabayashi, T.; Miyakoshi, M.; Shintani, M.; Nishida, H.; et al. Pmr, a histone-like protein H1 (H-NS) family protein encoded by the IncP-7 plasmid pCAR1, is a key global regulator that alters host function. J. Bacteriol. 2010, 192, 4720–4731. [Google Scholar] [CrossRef] [Green Version]
- Pfeifer, E.; Hünnefeld, M.; Popa, O.; Frunzke, J. Impact of xenogeneic silencing on phage-host interactions. J. Mol. Biol. 2019, 431, 4670–4683. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.T.; Zhang, B.; Li, M.L.; Liu, K.H.; Jiao, J.; Tian, C.F. The convergent xenogeneic silencer MucR predisposes α-proteobacteria to integrate AT-rich symbiosis genes. Nucleic Acids Res. 2022, 50, 8580–8598. [Google Scholar] [CrossRef] [PubMed]
- Dame, R.T.; Luijsterburg, M.S.; Krin, E.; Bertin, P.N.; Wagner, R.; Wuite, G.J.L. DNA Bridging: A property shared among H-NS-like proteins. J. Bacteriol. 2005, 187, 1845–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Chen, H.; Kenney, L.J.; Yan, J. A divalent switch drives H-NS/DNA-binding conformations between stiffening and bridging modes. Genes Dev. 2010, 24, 339–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Valk, R.A.; Vreede, J.; Qin, L.; Moolenaar, G.F.; Hofmann, A.; Goosen, N.; Dame, R.T. Mechanism of environmentally driven conformational changes that modulate H-NS DNA-bridging activity. Elife 2017, 6, e27369. [Google Scholar] [CrossRef] [PubMed]
- Navarre, W.W.; Porwollik, S.; Wang, Y.; McClelland, M.; Rosen, H.; Libby, S.J.; Fang, F.C. Selective silencing of foreign DNA with low GC content by the H-NS protein in Salmonella. Science 2006, 313, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Tendeng, C.; Soutourina, O.A.; Danchin, A.; Bertin, P.N. MvaT proteins in Pseudomonas spp.: A novel class of H-NS-like proteins. Microbiology 2003, 149, 3047–3050. [Google Scholar] [CrossRef] [Green Version]
- Duan, B.; Ding, P.; Navarre, W.W.; Liu, J.; Xia, B. Xenogeneic silencing and bacterial genome evolution: Mechanisms for DNA recognition imply multifaceted roles of xenogeneic silencers. Mol. Biol. Evol. 2021, 38, 4135–4148. [Google Scholar] [CrossRef]
- Gordon, B.R.G.; Imperial, R.; Wang, L.; Navarre, W.W.; Liu, J. Lsr2 of Mycobacterium represents a novel class of H-NS-like proteins. J. Bacteriol. 2008, 190, 7052–7059. [Google Scholar] [CrossRef] [Green Version]
- Colangeli, R.; Helb, D.; Vilchèze, C.; Hazbón, M.H.; Lee, C.G.; Safi, H.; Sayers, B.; Sardone, I.; Jones, M.B.; Fleischmann, R.D.; et al. Transcriptional regulation of multi-drug tolerance and antibiotic-induced responses by the histone-like protein Lsr2 in M. tuberculosis. PLoS Pathog. 2007, 3, 780–793. [Google Scholar] [CrossRef]
- Seid, C.A.; Smith, J.L.; Grossman, A.D. Genetic and biochemical interactions between the bacterial replication initiator DnaA and the nucleoid-associated protein Rok in Bacillus subtilis. Mol. Microbiol. 2017, 103, 798–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, W.K.; Grossman, A.D. The transcriptional regulator Rok binds A+T-rich DNA and is involved in repression of a mobile genetic element in Bacillus subtilis. PLoS Genet. 2010, 6, e1001207. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Milstein, J.N.; Navarre, W.W. Xenogeneic silencing and its impact on bacterial genomes. Annu. Rev. Microbiol. 2016, 70, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Ni, M.; Zhang, B.; Zhang, Z.; Young, J.P.W.; Chan, T.-F.; Chen, W.X.; Lam, H.-M.; Tian, C.F. Coordinated regulation of core and accessory genes in the multipartite genome of Sinorhizobium fredii. PLoS Genet. 2018, 14, e1007428. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Protein | Length (aa) | Bacterial Species | Identity (%) | Query Cover (%) | E-Value | Accession Number |
|---|---|---|---|---|---|---|
| MucR | 143 | Sinorhizobium meliloti 2011 | 100 | 100 | 9 × 10−87 | WP_003527383.1 |
| MucR | 143 | Sinorhizobium medicae | 99.30 | 100 | 3 × 10−86 | WP_011974816.1 |
| MucR1 | 143 | Sinorhizobium fredii HH103 | 96.50 | 100 | 2 × 10−84 | WP_014327705.1 |
| MucR1 | 143 | Sinorhizobium fredii CCBAU 45436 | 96.50 | 100 | 2 × 10−84 | AWI62033 |
| MucR | 143 | Ensifer aridi | 93.01 | 100 | 1 × 10−81 | WP_026615989.1 |
| MucR | 143 | Rhizobium herbae | 89.51 | 100 | 1 × 10−77 | WP_220372103.1 |
| MucR | 143 | Rhizobium giardinii | 88.11 | 100 | 2 × 10−77 | WP_018329544.1 |
| MucR | 143 | Pararhizobium sp. YC-54 | 87.41 | 100 | 3 × 10−76 | WP_264219505.1 |
| MucR | 143 | Pararhizobium polonicum | 86.71 | 100 | 1 × 10−75 | WP_068952849.1 |
| MucR | 144 | Rhizobium herbae | 85.92 | 99 | 3 × 10−75 | WP_209853764.1 |
| MucR | 142 | Agrobacterium rhizogenes | 85.71 | 97 | 2 × 10−74 | WP_047463113.1 |
| MucR | 143 | Rhizobium lusitanum | 82.52 | 100 | 2 × 10−72 | QND49923.1 |
| MucR | 143 | Rhizobium sullae | 81.82 | 99 | 1 × 10−71 | WP_132559880.1 |
| RosR | 143 | Rhizobium leguminosarum | 81.12 | 99 | 7 × 10−71 | AAT92553 |
| Ros | 143 | Rhizobium etli | 80.12 | 99 | 1 × 10−71 | AAC44878 |
| Ros | 143 | Agrobacterium tumefaciens | 82.52 | 100 | 3 × 10−74 | WP_132517515.1 |
| Protein | Length (aa) | Bacterial Species | Identity (%) | Query Cover (%) | E-Value | Accession Number |
|---|---|---|---|---|---|---|
| MucR | 142 | Brucella melitensis 16M | 100 | 100 | 1 × 10−80 | WP_006266678.1 |
| MucR | 161 | Brucella ovis ATCC 25840 | 99.30 | 100 | 4 × 10−80 | ABQ60939.1 |
| MucR | 142 | Brucella abortus | 99.30 | 100 | 4 × 10−80 | USC10419 |
| MucR | 142 | Brucella inopinata | 98.59 | 100 | 7 × 10−80 | WP_008508511.1 |
| MucR | 142 | Brucella oryzae | 94.37 | 100 | 2 × 10−76 | WP_104755423.1 |
| MucR | 142 | Brucella pecoris | 93.66 | 100 | 7 × 10−77 | WP_140019901.1 |
| MucR | 143 | Paramesorhizobium deserti | 82.61 | 97 | 5 × 10−64 | WP_068879566.1 |
| MucR | 142 | Phyllobacterium phragmitis | 80.71 | 98 | 2 × 10−61 | WP_105740081.1 |
| MucR | 142 | Brucella endophytica | 78.87 | 100 | 6 × 10−62 | WP_188826054.1 |
| MucR | 143 | Mesorhizobium ephedrae | 71.83 | 100 | 8 × 10−57 | WP_106775715.1 |
| Ml1 | 149 | Mesorhizobium loti | 72.14 | 98 | 2 × 10−55 | Q989W1 |
| Ml2 | 141 | Mesorhizobium loti | 71.89 | 98 | 5 × 10−56 | Q985J6 |
| MucR | 142 | Mesorhizobium tamadayense | 72.14 | 98 | 6 × 10−56 | WP_125005267.1 |
| MucR | 142 | Mesorhizobium sp. ORS 3428 | 71.43 | 98 | 1 × 10−55 | WP_071100028.1 |
| MucR | 141 | Mesorhizobium soli | 71.43 | 98 | 3 × 10−55 | WP_106723787.1 |
| MucR | 141 | Mesorhizobium alhagi | 70.00 | 98 | 9 × 10−55 | WP_008839483.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janczarek, M. The Ros/MucR Zinc-Finger Protein Family in Bacteria: Structure and Functions. Int. J. Mol. Sci. 2022, 23, 15536. https://doi.org/10.3390/ijms232415536
Janczarek M. The Ros/MucR Zinc-Finger Protein Family in Bacteria: Structure and Functions. International Journal of Molecular Sciences. 2022; 23(24):15536. https://doi.org/10.3390/ijms232415536
Chicago/Turabian StyleJanczarek, Monika. 2022. "The Ros/MucR Zinc-Finger Protein Family in Bacteria: Structure and Functions" International Journal of Molecular Sciences 23, no. 24: 15536. https://doi.org/10.3390/ijms232415536
APA StyleJanczarek, M. (2022). The Ros/MucR Zinc-Finger Protein Family in Bacteria: Structure and Functions. International Journal of Molecular Sciences, 23(24), 15536. https://doi.org/10.3390/ijms232415536