

The Role of Ectodysplasin A on the Ocular Surface Homeostasis

 , ,
, ,  ,
, 
Abstract
:1. Introduction
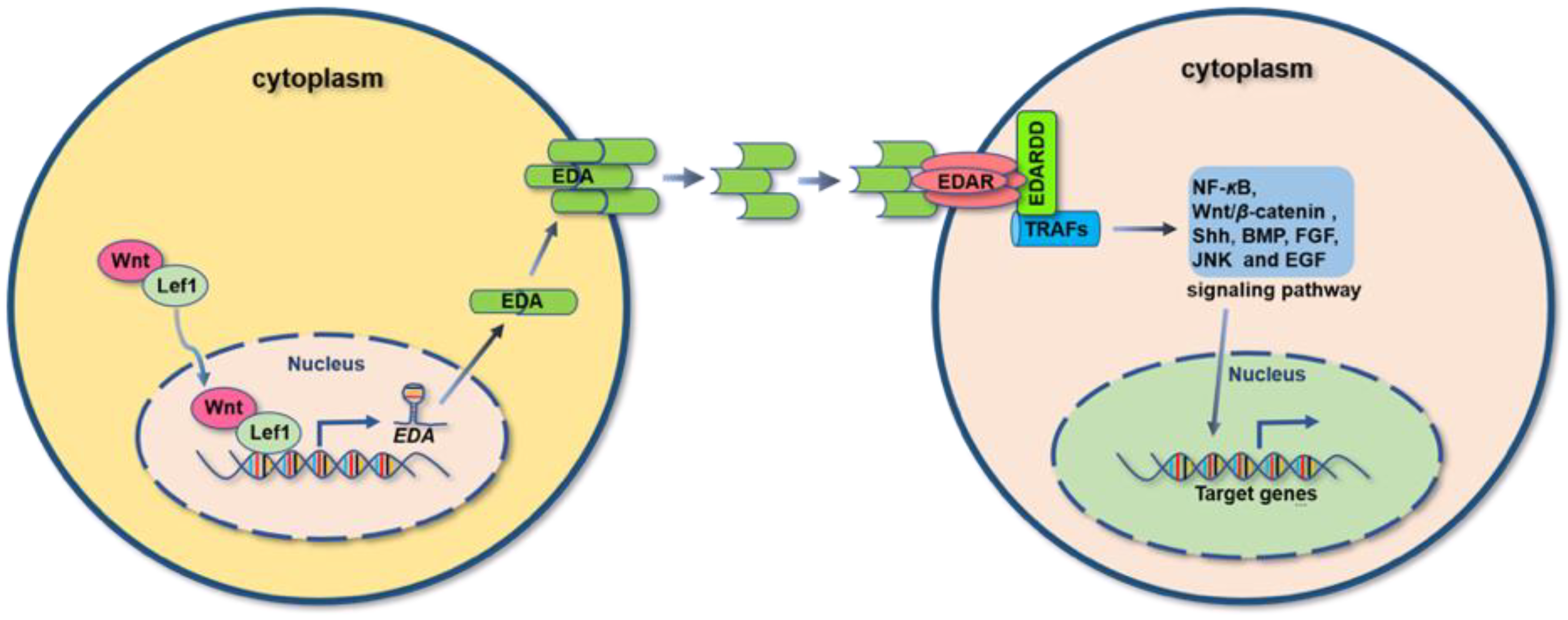
2. EDA/EDA Receptors System
3. EDA-EDAR-Dependent Signaling Pathways
3.1. NF-κB Signaling Pathway
3.2. Wnt/β-Catenin Signaling Pathway
3.3. BMP Signaling Pathway
3.4. c-Jun N-Terminal Kinase Signaling Pathway
4. Function of EDA in Physiology and Pathology
5. The Homeostasis of Ocular Surface
6. The Role of EDA in the Development of Ocular Surface
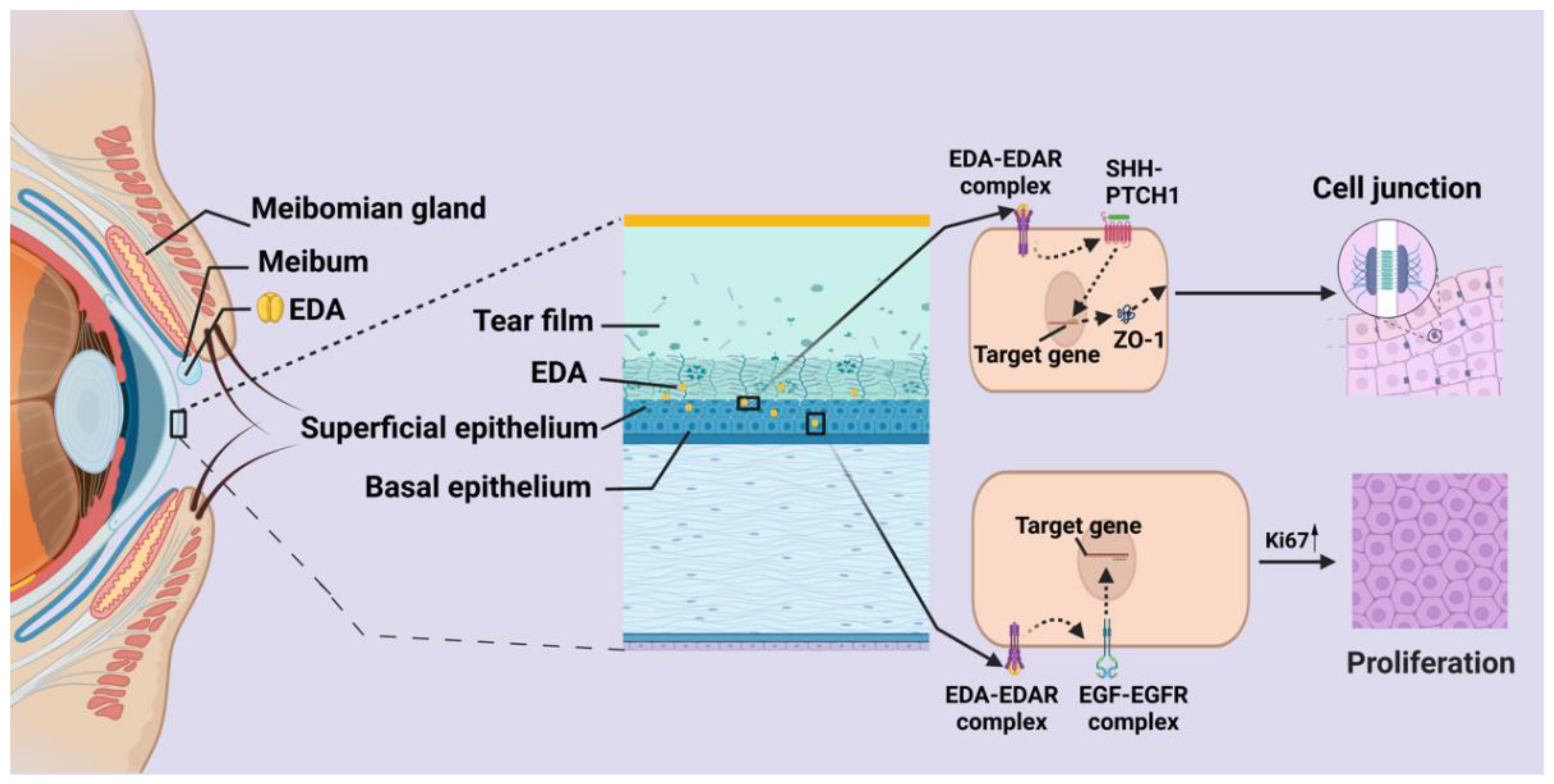
7. The Role of EDA in Ocular Surface Homeostasis
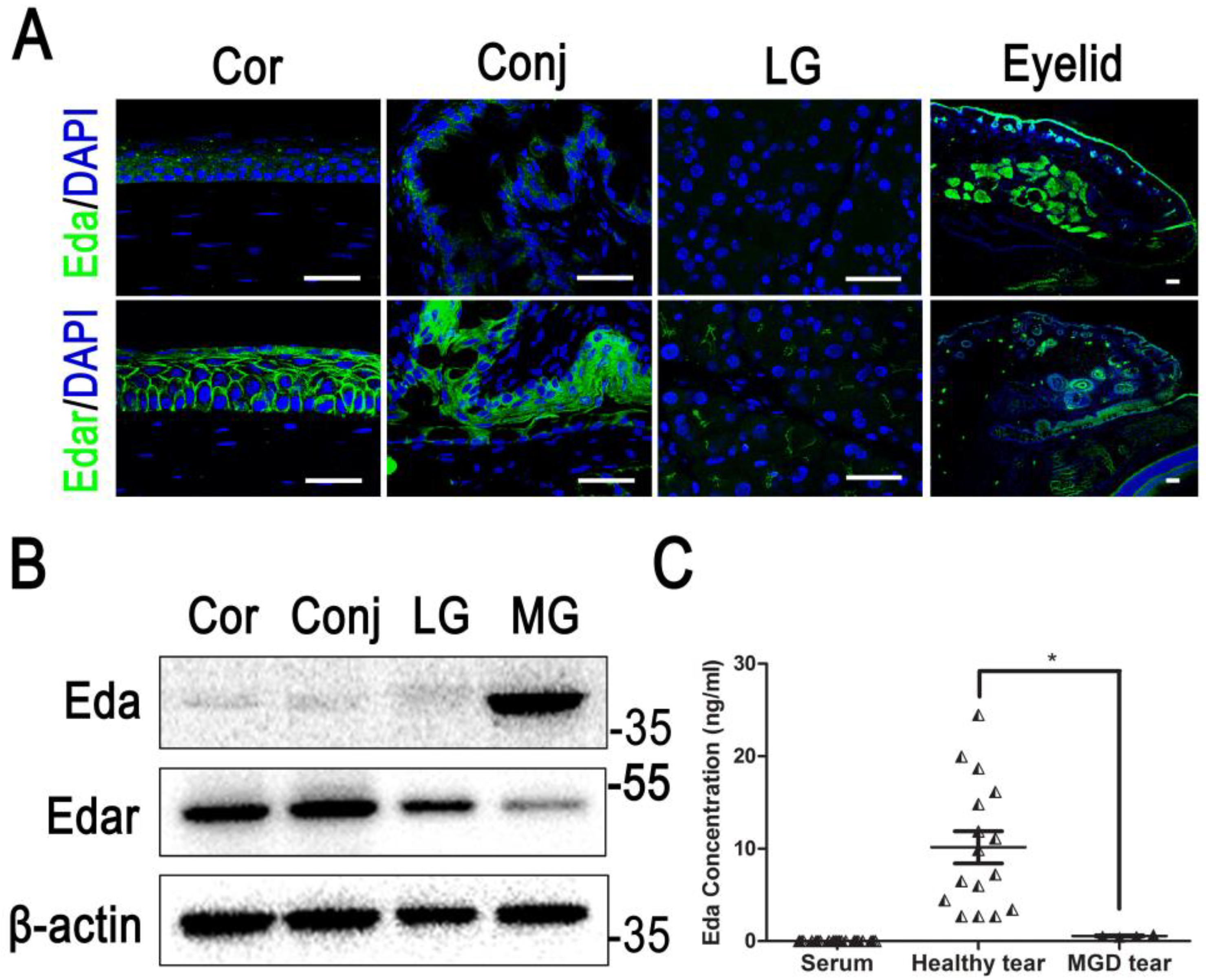
7.1. Meibomian Gland
7.2. Lacrimal Gland
7.3. Cornea
8. Therapeutic Efficiency of Recombinant EDA
9. Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cosman, D. A family of ligands for the TNF receptor superfamily. Stem Cells 1994, 12, 440–455. [Google Scholar] [CrossRef] [PubMed]
- Ezer, S.; Bayés, M.; Elomaa, O.; Schlessinger, D.; Kere, J. Ectodysplasin is a collagenous trimeric type II membrane protein with a tumor necrosis factor-like domain and co-localizes with cytoskeletal structures at lateral and apical surfaces of cells. Hum. Mol. Genet 1999, 8, 2079–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadier, A.; Viriot, L.; Pantalacci, S.; Laudet, V. The ectodysplasin pathway: From diseases to adaptations. Trends Genet 2014, 30, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Schneider, P.; Street, S.L.; Gaide, O.; Hertig, S.; Tardivel, A.; Tschopp, J.; Runkel, L.; Alevizopoulos, K.; Ferguson, B.M.; Zonana, J. Mutations leading to X-linked hypohidrotic ectodermal dysplasia affect three major functional domains in the tumor necrosis factor family member ectodysplasin-A. J. Biol. Chem. 2001, 276, 18819–18827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayes, M.; Hartung, A.J.; Ezer, S.; Pispa, J.; Thesleff, I.; Srivastava, A.K.; Kere, J. The anhidrotic ectodermal dysplasia gene (EDA) undergoes alternative splicing and encodes ectodysplasin-A with deletion mutations in collagenous repeats. Hum. Mol. Genet. 1998, 7, 1661–1669. [Google Scholar] [CrossRef] [Green Version]
- Yan, M.; Wang, L.C.; Hymowitz, S.G.; Schilbach, S.; Lee, J.; Goddard, A.; de Vos, A.M.; Gao, W.Q.; Dixit, V.M. Two-amino acid molecular switch in an epithelial morphogen that regulates binding to two distinct receptors. Science 2000, 290, 523–527. [Google Scholar] [CrossRef]
- Brosh, R.; Sarig, R.; Bar Natan, E.; Molchadsky, A.; Madar, S.; Bornstein, C.; Buganim, Y.; Shapira, T.; Goldfinger, N.; Paus, R.; et al. p53-dependent transcriptional regulation of EDA2R and its involvement in chemotherapy-induced hair loss. Febs Lett. 2010, 584, 2473–2477. [Google Scholar] [CrossRef]
- Pinheiro, M.; Freiremaia, N. Ectodermal Dysplasias—A Clinical Classification and a Causal Review. Am. J. Med. Genet. 1994, 53, 153–162. [Google Scholar] [CrossRef]
- Hymowitz, S.G.; Compaan, D.M.; Yan, M.; Wallweber, H.J.; Dixit, V.M.; Starovasnik, M.A.; de Vos, A.M. The crystal structures of EDA-A1 and EDA-A2: Splice variants with distinct receptor specificity. Structure 2003, 11, 1513–1520. [Google Scholar] [CrossRef] [Green Version]
- Chassaing, N.; Cluzeau, C.; Bal, E.; Guigue, P.; Vincent, M.C.; Viot, G.; Ginisty, D.; Munnich, A.; Smahi, A.; Calvas, P. Mutations in EDARADD account for a small proportion of hypohidrotic ectodermal dysplasia cases. Br. J. Derm. 2010, 162, 1044–1048. [Google Scholar] [CrossRef]
- Kojima, T.; Morikawa, Y.; Copeland, N.G.; Gilbert, D.J.; Jenkins, N.A.; Senba, E.; Kitamura, T. TROY, a newly identified member of the tumor necrosis factor receptor superfamily, exhibits a homology with Edar and is expressed in embryonic skin and hair follicles. J. Biol. Chem. 2000, 275, 20742–20747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, A.; Yoshida, H.; Nishioka, E.; Satoh, M.; Azuma, S.; Yamamoto, T.; Nishikawa, S.-i.; Inoue, J.-i. TRAF6-deficient mice display hypohidrotic ectodermal dysplasia. Proc. Natl. Acad. Sci. USA 2002, 99, 8766–8771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morlon, A.; Munnich, A.; Smahi, A. TAB2, TRAF6 and TAK1 are involved in NF-kappaB activation induced by the TNF-receptor, Edar and its adaptator Edaradd. Hum. Mol. Genet. 2005, 14, 3751–3757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanikawa, C.; Ri, C.; Kumar, V.; Nakamura, Y.; Matsuda, K. Crosstalk of EDA-A2/XEDAR in the p53 signaling pathway. Mol. Cancer Res. 2010, 8, 855–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, S.K.; Zachariah, S.; Quiñones, H.I.; Shindo, M.; Chaudhary, P.M. Role of TRAF3 and -6 in the activation of the NF-kappa B and JNK pathways by X-linked ectodermal dysplasia receptor. J. Biol. Chem. 2002, 277, 44953–44961. [Google Scholar] [CrossRef] [Green Version]
- Lindfors, P.H.; Voutilainen, M.; Mikkola, M.L. Ectodysplasin/NF-κB signaling in embryonic mammary gland development. J. Mammary Gland Biol. Neoplasia 2013, 18, 165–169. [Google Scholar] [CrossRef]
- Courtney, J.M.; Blackburn, J.; Sharpe, P.T. The Ectodysplasin and NFkappaB signalling pathways in odontogenesis. Arch. Oral. Biol. 2005, 50, 159–163. [Google Scholar] [CrossRef]
- Botchkarev, V.A.; Fessing, M.Y. Edar signaling in the control of hair follicle development. J. Investig. Derm. Symp. Proc. 2005, 10, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.Y.; Durmowicz, M.; Ottolenghi, C.; Hashimoto, T.; Griggs, B.; Srivastava, A.K.; Schlessinger, D. Inducible mEDA-A1 transgene mediates sebaceous gland hyperplasia and differential formation of two types of mouse hair follicles. Hum. Mol. Genet. 2003, 12, 2931–2940. [Google Scholar] [CrossRef]
- Zhang, Y.; Tomann, P.; Andl, T.; Gallant, N.M.; Huelsken, J.; Jerchow, B.; Birchmeier, W.; Paus, R.; Piccolo, S.; Mikkola, M.L.; et al. Reciprocal requirements for EDA/EDAR/NF-kappaB and Wnt/beta-catenin signaling pathways in hair follicle induction. Dev. Cell 2009, 17, 49–61. [Google Scholar] [CrossRef]
- Laurikkala, J.; Mikkola, M.; Mustonen, T.; Aberg, T.; Koppinen, P.; Pispa, J.; Nieminen, P.; Galceran, J.; Grosschedl, R.; Thesleff, I. TNF signaling via the ligand-receptor pair ectodysplasin and edar controls the function of epithelial signaling centers and is regulated by Wnt and activin during tooth organogenesis. Dev. Biol. 2001, 229, 443–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sima, J.; Piao, Y.; Chen, Y.; Schlessinger, D. Molecular dynamics of Dkk4 modulates Wnt action and regulates meibomian gland development. Development 2016, 143, 4723–4735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voutilainen, M.; Lindfors, P.H.; Trela, E.; Lönnblad, D.; Shirokova, V.; Elo, T.; Rysti, E.; Schmidt-Ullrich, R.; Schneider, P.; Mikkola, M.L. Ectodysplasin/NF-κB Promotes Mammary Cell Fate via Wnt/β-catenin Pathway. PLoS Genet. 2015, 11, e1005676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Liang, Y.; Chai, X.; Chen, S.; Ye, Z.; Li, R.; Li, X.; Kong, G.; Li, Y.; Zhang, X.; et al. Ectodysplasin A receptor (EDAR) promotes colorectal cancer cell proliferation via regulation of the Wnt/β-catenin signaling pathway. Exp. Cell Res. 2020, 395, 112170. [Google Scholar] [CrossRef]
- Mou, C.; Jackson, B.; Schneider, P.; Overbeek, P.A.; Headon, D.J. Generation of the primary hair follicle pattern. Proc. Natl. Acad. Sci. USA 2006, 103, 9075–9080. [Google Scholar] [CrossRef] [Green Version]
- Voutilainen, M.; Lindfors, P.H.; Lefebvre, S.; Ahtiainen, L.; Fliniaux, I.; Rysti, E.; Murtoniemi, M.; Schneider, P.; Schmidt-Ullrich, R.; Mikkola, M.L. Ectodysplasin regulates hormone-independent mammary ductal morphogenesis via NF-κB. Proc. Natl. Acad. Sci. USA 2012, 109, 5744–5749. [Google Scholar] [CrossRef] [Green Version]
- Koppinen, P.; Pispa, J.; Laurikkala, J.; Thesleff, I.; Mikkola, M.L. Signaling and subcellular localization of the TNF receptor Edar. Exp. Cell Res. 2001, 269, 180–192. [Google Scholar] [CrossRef]
- Awazawa, M.; Gabel, P.; Tsaousidou, E.; Nolte, H.; Krüger, M.; Schmitz, J.; Ackermann, P.J.; Brandt, C.; Altmüller, J.; Motameny, S.; et al. A microRNA screen reveals that elevated hepatic ectodysplasin A expression contributes to obesity-induced insulin resistance in skeletal muscle. Nat. Med. 2017, 23, 1466–1473. [Google Scholar] [CrossRef]
- Kumar, A.; Eby, M.T.; Sinha, S.; Jasmin, A.; Chaudhary, P.M. The ectodermal dysplasia receptor activates the nuclear factor-kappaB, JNK, and cell death pathways and binds to ectodysplasin A. J. Biol. Chem. 2001, 276, 2668–2677. [Google Scholar] [CrossRef] [Green Version]
- Pummila, M.; Fliniaux, I.; Jaatinen, R.; James, M.J.; Laurikkala, J.; Schneider, P.; Thesleff, I.; Mikkola, M.L. Ectodysplasin has a dual role in ectodermal organogenesis: Inhibition of Bmp activity and induction of Shh expression. Development 2007, 134, 117–125. [Google Scholar] [CrossRef]
- Elo, T.; Lindfors, P.H.; Lan, Q.; Voutilainen, M.; Trela, E.; Ohlsson, C.; Huh, S.H.; Ornitz, D.M.; Poutanen, M.; Howard, B.A.; et al. Ectodysplasin target gene Fgf20 regulates mammary bud growth and ductal invasion and branching during puberty. Sci. Rep. 2017, 7, 5049. [Google Scholar] [CrossRef]
- Isaacs, K.; Brown, G.; Moore, G.P. Interactions between epidermal growth factor and the Tabby mutation in skin. Exp. Derm. 1998, 7, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Li, S.M.; Zhou, J.; Bu, J.H.; Ning, K.; Zhang, L.Y.; Li, J.; Guo, Y.L.; He, X.; He, H.; Cai, X.X.; et al. Ectodysplasin A protein promotes corneal epithelial cell proliferation. J. Biol. Chem. 2017, 292, 13391–13401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elomaa, O.; Pulkkinen, K.; Hannelius, U.; Mikkola, M.; Saarialho-Kere, U.; Kere, J. Ectodysplasin is released by proteolytic shedding and binds to the EDAR protein. Hum. Mol. Genet. 2001, 10, 953–962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, C.Y.; Schlessinger, D. EDA signaling and skin appendage development. Cell Cycle 2006, 5, 2477–2483. [Google Scholar] [CrossRef] [PubMed]
- Kere, J.; Srivastava, A.K.; Montonen, O.; Zonana, J.; Thomas, N.; Ferguson, B.; Munoz, F.; Morgan, D.; Clarke, A.; Baybayan, P.; et al. X-linked anhidrotic (hypohidrotic) ectodermal dysplasia is caused by mutation in a novel transmembrane protein. Nat. Genet. 1996, 13, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Brennan, E.P.; Mohan, M.; McClelland, A.; Tikellis, C.; Ziemann, M.; Kaspi, A.; Gray, S.P.; Pickering, R.; Tan, S.M.; Ali-Shah, S.T.; et al. Lipoxins Regulate the Early Growth Response-1 Network and Reverse Diabetic Kidney Disease. J. Am. Soc. Nephrol. 2018, 29, 1437–1448. [Google Scholar] [CrossRef] [Green Version]
- Duan, F.; Zhai, Y.; Kong, X. [Clinical and genetic analysis of a child with X-linked hypohidrotic ectodermal dysplasia]. Zhonghua Yi Xue Yi Chuan Xue Za Zhi 2021, 38, 469–471. [Google Scholar] [CrossRef]
- Bayliss, J.; Ooi, G.J.; De Nardo, W.; Shah, Y.J.H.; Montgomery, M.K.; McLean, C.; Kemp, W.; Roberts, S.K.; Brown, W.A.; Burton, P.R.; et al. Ectodysplasin A Is Increased in Non-Alcoholic Fatty Liver Disease, But Is Not Associated With Type 2 Diabetes. Front. Endocrinol. 2021, 12, 642432. [Google Scholar] [CrossRef]
- Yan, M.; Zhang, Z.; Brady, J.R.; Schilbach, S.; Fairbrother, W.J.; Dixit, V.M. Identification of a Novel Death Domain-Containing Adaptor Molecule for Ectodysplasin-A Receptor that Is Mutated in crinkled Mice. Curr. Biol. 2002, 12, 409–413. [Google Scholar] [CrossRef]
- Pispa, J.; Mikkola, M.L.; Mustonen, T.; Thesleff, I. Ectodysplasin, Edar and TNFRSF19 are expressed in complementary and overlapping patterns during mouse embryogenesis. Gene Expr. Patterns 2003, 3, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Rawlins, E.L.; Hogan, B.L. Intercellular growth factor signaling and the development of mouse tracheal submucosal glands. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2005, 233, 1378–1385. [Google Scholar] [CrossRef] [PubMed]
- Ostedgaard, L.S.; Price, M.P.; Whitworth, K.M.; Abou Alaiwa, M.H.; Fischer, A.J.; Warrier, A.; Samuel, M.; Spate, L.D.; Allen, P.D.; Hilkin, B.M.; et al. Lack of airway submucosal glands impairs respiratory host defenses. Elife 2020, 9, e59653. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhou, J.; Zhang, L.; Li, J.; Yu, J.; Ning, K.; Qu, Y.; He, H.; Chen, Y.; Reinach, P.S.; et al. Ectodysplasin A regulates epithelial barrier function through sonic hedgehog signalling pathway. J. Cell. Mol. Med. 2018, 22, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Mauldin, E.A.; Gaide, O.; Schneider, P.; Casal, M.L. Neonatal treatment with recombinant ectodysplasin prevents respiratory disease in dogs with X-linked ectodermal dysplasia. Am. J. Med. Genet. A 2009, 149a, 2045–2049. [Google Scholar] [CrossRef] [Green Version]
- Dietz, J.; Kaercher, T.; Schneider, A.T.; Zimmermann, T.; Huttner, K.; Johnson, R.; Schneider, H. Early respiratory and ocular involvement in X-linked hypohidrotic ectodermal dysplasia. Eur. J. Pediatr. 2013, 172, 1023–1031. [Google Scholar] [CrossRef]
- Marrero, E.; Attal, N.; Nimeri, A.; McGee, R.M.; Benbow, J.H.; Thompson, K.J.; Schrum, L.W.; McKillop, I.H. Ectodysplasin-A mRNA in exosomes released from activated hepatic stellate cells stimulates macrophage response. Exp. Cell Res. 2022, 419, 113297. [Google Scholar] [CrossRef]
- Garcin, C.L.; Huttner, K.M.; Kirby, N.; Schneider, P.; Hardman, M.J. Ectodysplasin A Pathway Contributes to Human and Murine Skin Repair. J. Investig. Derm. 2016, 136, 1022–1030. [Google Scholar] [CrossRef]
- Mustonen, T.; Ilmonen, M.; Pummila, M.; Kangas, A.T.; Laurikkala, J.; Jaatinen, R.; Pispa, J.; Gaide, O.; Schneider, P.; Thesleff, I.; et al. Ectodysplasin A1 promotes placodal cell fate during early morphogenesis of ectodermal appendages. Development 2004, 131, 4907–4919. [Google Scholar] [CrossRef] [Green Version]
- Kamberov, Y.G.; Wang, S.; Tan, J.; Gerbault, P.; Wark, A.; Tan, L.; Yang, Y.; Li, S.; Tang, K.; Chen, H.; et al. Modeling recent human evolution in mice by expression of a selected EDAR variant. Cell 2013, 152, 691–702. [Google Scholar] [CrossRef]
- Wahlbuhl, M.; Schuepbach-Mallepell, S.; Kowalczyk-Quintas, C.; Dick, A.; Fahlbusch, F.B.; Schneider, P.; Schneider, H. Attenuation of Mammary Gland Dysplasia and Feeding Difficulties in Tabby Mice by Fetal Therapy. J. Mammary Gland. Biol. Neoplasia 2018, 23, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Kuony, A.; Ikkala, K.; Kalha, S.; Magalhães, A.C.; Pirttiniemi, A.; Michon, F. Ectodysplasin-A signaling is a key integrator in the lacrimal gland-cornea feedback loop. Development 2019, 146, dev176693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.C.; Li, S.; Chen, X.; Ma, B.; He, H.; Liu, T.; Yu, J.; Zhang, L.; Chen, Y.; Liu, Z.; et al. Meibomian Gland Absence Related Dry Eye in Ectodysplasin A Mutant Mice. Am. J. Pathol. 2016, 186, 32–42. [Google Scholar] [CrossRef]
- Zhang, X.; Jeyalatha, M.J.; Qu, Y.; He, X.; Ou, S.; Bu, J.; Jia, C.; Wang, J.; Wu, H.; Liu, Z.; et al. Dry Eye Management: Targeting the Ocular Surface Microenvironment. Int. J. Mol. Sci. 2017, 18, 1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gipson, I.K. Goblet cells of the conjunctiva: A review of recent findings. Prog. Retin. Eye Res. 2016, 54, 49–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirayama, M. Advances in Functional Restoration of the Lacrimal Glands. Investig. Ophthalmol. Vis. Sci. 2018, 59, Des174–Des182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butovich, I.A. Meibomian glands, meibum, and meibogenesis. Exp. Eye Res. 2017, 163, 2–16. [Google Scholar] [CrossRef]
- Tawfik, H.A.; Abdulhafez, M.H.; Fouad, Y.A.; Dutton, J.J. Embryologic and Fetal Development of the Human Eyelid. Ophthalmic. Plast. Reconstr. Surg. 2016, 32, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.Y.; Kunisada, M.; Esibizione, D.; Douglass, E.G.; Schlessinger, D. Analysis of the temporal requirement for eda in hair and sweat gland development. J. Investig. Derm. 2009, 129, 984–993. [Google Scholar] [CrossRef] [Green Version]
- Beahrs, J.O.; Lillington, G.A.; Rosan, R.C.; Russin, L.; Lindgren, J.A.; Rowley, P.T. Anhidrotic ectodermal dysplasia: Predisposition to bronchial disease. Ann. Intern. Med. 1971, 74, 92–96. [Google Scholar] [CrossRef]
- Clarke, A.; Phillips, D.I.; Brown, R.; Harper, P.S. Clinical aspects of X-linked hypohidrotic ectodermal dysplasia. Arch. Dis. Child. 1987, 62, 989–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Söderholm, A.L.; Kaitila, I. Expression of X-linked hypohidrotic ectodermal dysplasia in six males and in their mothers. Clin. Genet. 1985, 28, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Gilgenkrantz, S.; Blanchet-Bardon, C.; Nazzaro, V.; Formiga, L.; Mujica, P.; Alembik, Y. Hypohidrotic ectodermal dysplasia. Clinical study of a family of 30 over three generations. Hum. Genet. 1989, 81, 120–122. [Google Scholar] [CrossRef] [PubMed]
- Kaercher, T. Ocular symptoms and signs in patients with ectodermal dysplasia syndromes. Graefes Arch. Clin. Exp. Ophthalmol. 2004, 242, 495–500. [Google Scholar] [CrossRef]
- Casal, M.L.; Lewis, J.R.; Mauldin, E.A.; Tardivel, A.; Ingold, K.; Favre, M.; Paradies, F.; Demotz, S.; Gaide, O.; Schneider, P. Significant correction of disease after postnatal administration of recombinant ectodysplasin A in canine X-linked ectodermal dysplasia. Am. J. Hum. Genet. 2007, 81, 1050–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margolis, C.A.; Schneider, P.; Huttner, K.; Kirby, N.; Houser, T.P.; Wildman, L.; Grove, G.L.; Schneider, H.; Casal, M.L. Prenatal Treatment of X-Linked Hypohidrotic Ectodermal Dysplasia using Recombinant Ectodysplasin in a Canine Model. J. Pharm. Exp. Ther. 2019, 370, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.Y.; Kunisada, M.; Piao, Y.; Childress, V.; Ko, M.S.; Schlessinger, D. Dkk4 and Eda regulate distinctive developmental mechanisms for subtypes of mouse hair. PLoS ONE 2010, 5, e10009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaide, O.; Schneider, P. Permanent correction of an inherited ectodermal dysplasia with recombinant EDA. Nat. Med. 2003, 9, 614–618. [Google Scholar] [CrossRef] [Green Version]
- Kuramoto, T.; Yokoe, M.; Hashimoto, R.; Hiai, H.; Serikawa, T. A rat model of hypohidrotic ectodermal dysplasia carries a missense mutation in the Edaradd gene. BMC Genet. 2011, 12, 91. [Google Scholar] [CrossRef] [Green Version]
- Cui, C.Y.; Smith, J.A.; Schlessinger, D.; Chan, C.C. X-linked anhidrotic ectodermal dysplasia disruption yields a mouse model for ocular surface disease and resultant blindness. Am. J. Pathol. 2005, 167, 89–95. [Google Scholar] [CrossRef]
- Klenkler, B.; Sheardown, H.; Jones, L. Growth factors in the tear film: Role in tissue maintenance, wound healing, and ocular pathology. Ocul. Surf. 2007, 5, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Sabeti, S.; Kheirkhah, A.; Yin, J.; Dana, R. Management of meibomian gland dysfunction: A review. Surv. Ophthalmol. 2020, 65, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Wohlfart, S.; Meiller, R.; Hammersen, J.; Park, J.; Menzel-Severing, J.; Melichar, V.O.; Huttner, K.; Johnson, R.; Porte, F.; Schneider, H. Natural history of X-linked hypohidrotic ectodermal dysplasia: A 5-year follow-up study. Orphanet. J. Rare Dis. 2020, 15, 7. [Google Scholar] [CrossRef] [PubMed]
- Craig, J.P.; Nichols, K.K.; Akpek, E.K.; Caffery, B.; Dua, H.S.; Joo, C.K.; Liu, Z.; Nelson, J.D.; Nichols, J.J.; Tsubota, K.; et al. TFOS DEWS II Definition and Classification Report. Ocul. Surf. 2017, 15, 276–283. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Kruse, F.E.; Pohl, J.; Volcker, H.E. Bone morphogenetic proteins and growth and differentiation factors in the human cornea. Investig. Ophthalmol. Vis. Sci. 1999, 40, 296–311. [Google Scholar]
- Yoon, K.C.; Park, C.S.; You, I.C.; Choi, H.J.; Lee, K.H.; Im, S.K.; Park, H.Y.; Pflugfelder, S.C. Expression of CXCL9, -10, -11, and CXCR3 in the tear film and ocular surface of patients with dry eye syndrome. Investig. Ophthalmol. Vis. Sci. 2010, 51, 643–650. [Google Scholar] [CrossRef]
- Wilson, F.M., 2nd; Grayson, M.; Pieroni, D. Corneal changes in ectodermal dysplasia. Case report, histopathology, and differential diagnosis. Am. J. Ophthalmol. 1973, 75, 17–27. [Google Scholar] [CrossRef]
- Baum, J.L.; Bull, M.J. Ocular manifestations of the ectrodactyly, ectodermal dysplasia, cleft lip-palate syndrome. Am. J. Ophthalmol. 1974, 78, 211–216. [Google Scholar] [CrossRef]
- Headon, D.J.; Emmal, S.A.; Ferguson, B.M.; Tucker, A.S.; Justice, M.J.; Sharpe, P.T.; Zonana, J.; Overbeek, P.A. Gene defect in ectodermal dysplasia implicates a death domain adapter in development. Nature 2001, 414, 913–916. [Google Scholar] [CrossRef]
- Kowalczyk, C.; Dunkel, N.; Willen, L.; Casal, M.L.; Mauldin, E.A.; Gaide, O.; Tardivel, A.; Badic, G.; Etter, A.L.; Favre, M.; et al. Molecular and therapeutic characterization of anti-ectodysplasin A receptor (EDAR) agonist monoclonal antibodies. J. Biol. Chem. 2011, 286, 30769–30779. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Reali, J.; Mendoza-Ramos, M.I.; Garrido-Guerrero, E.; Mendez-Catala, C.F.; Mendez-Cruz, A.R.; Pozo-Molina, G. Hypohidrotic ectodermal dysplasia: Clinical and molecular review. Int. J. Derm. 2018, 57, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Hermes, K.; Schneider, P.; Krieg, P.; Dang, A.; Huttner, K.; Schneider, H. Prenatal therapy in developmental disorders: Drug targeting via intra-amniotic injection to treat X-linked hypohidrotic ectodermal dysplasia. J. Investig. Derm. 2014, 134, 2985–2987. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Physiology | Pathology | |
|---|---|---|
| Heart | Expression [11] | NA |
| Kidey | Expression in kidney epithelial cells [37] | Polycystic kidney dysplasia [38], diabetic kidneys [37]. |
| Pancreas | Expression [39,40] | Insulin resistance [39] |
| Brain | Expression [41] | NA |
| Lung | Expression in distal tracheal regions and the distal lung [42]. The development of submucosal glands [43]. | Lung infection [44,45], high prevalence of asthma-like symptoms [46] |
| Liver | Expression in hepatic stellate cells [28,47] | Increases in non-alcoholic fatty liver disease and insulin resistance [28] |
| Skeletal muscle | Expression in muscle cells [28] | Insulin resistance [28] |
| Skin and skin appendages | Expression in epithelium. The formation of skin appendages [35] and skin repair [48] | Defective formation and further morphogenesis dysfunction of hair follicles, sweat glands and teeth [35], delay in healing [48] |
| Mammary glands | Expression in mammary epithelium. Mammary placode formation and branching morphogenesis [31,49,50] | Smaller ductal trees [16,51] |
| Ocular surface | Expression in meibomian gland epithelium. Meibomian gland formation [19] and lacrimal gland morphogenesis [52] | Dry eye [53], delay in healing [33] |
| EDA | EDAR | Function | Pathway | Pathology | |
|---|---|---|---|---|---|
| Cornea | + | +++ | Proliferation Barrier function | EGF [33] SHH [70] ERK [33] | Corneal defect, keratitis, decrease in corneal epithelial proliferation and delayed corneal wound healing [33] |
| Conjunctiva | + | ++ | NA | NA | |
| Meibomian gland | +++ | ++ | Development | WNT [22] | Abnormal development of the meibomian gland [33] |
| Lacrimal gland | +/− | ++ | Development Lacrimation | Cxcl10 [52,71] | The terminal differentiation of cells was abnormal and a decrease in tear production [52] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ou, S.; Jeyalatha, M.V.; Mao, Y.; Wang, J.; Chen, C.; Zhang, M.; Liu, X.; Liang, M.; Lin, S.; Wu, Y.; et al. The Role of Ectodysplasin A on the Ocular Surface Homeostasis. Int. J. Mol. Sci. 2022, 23, 15700. https://doi.org/10.3390/ijms232415700
Ou S, Jeyalatha MV, Mao Y, Wang J, Chen C, Zhang M, Liu X, Liang M, Lin S, Wu Y, et al. The Role of Ectodysplasin A on the Ocular Surface Homeostasis. International Journal of Molecular Sciences. 2022; 23(24):15700. https://doi.org/10.3390/ijms232415700
Chicago/Turabian StyleOu, Shangkun, Mani Vimalin Jeyalatha, Yi Mao, Junqi Wang, Chao Chen, Minjie Zhang, Xiaodong Liu, Minghui Liang, Sijie Lin, Yiming Wu, and et al. 2022. "The Role of Ectodysplasin A on the Ocular Surface Homeostasis" International Journal of Molecular Sciences 23, no. 24: 15700. https://doi.org/10.3390/ijms232415700
APA StyleOu, S., Jeyalatha, M. V., Mao, Y., Wang, J., Chen, C., Zhang, M., Liu, X., Liang, M., Lin, S., Wu, Y., Li, Y., & Li, W. (2022). The Role of Ectodysplasin A on the Ocular Surface Homeostasis. International Journal of Molecular Sciences, 23(24), 15700. https://doi.org/10.3390/ijms232415700