
Anti-Psoriatic Effect of Rheum palmatum L. and Its Underlying Molecular Mechanisms



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
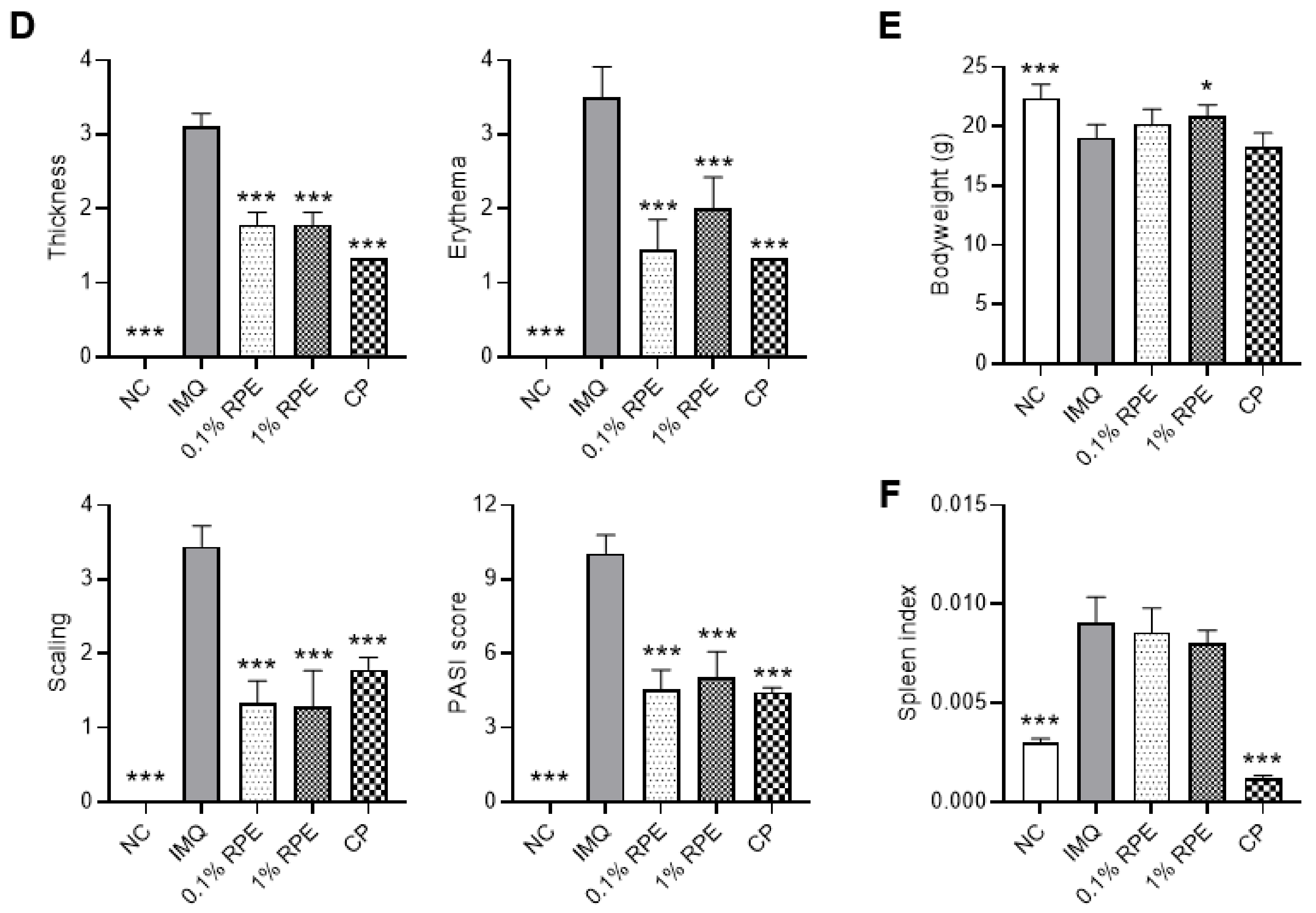
2.1. Effects of RPE on Psoriasis-like Symptoms in IMQ-Induced Mice
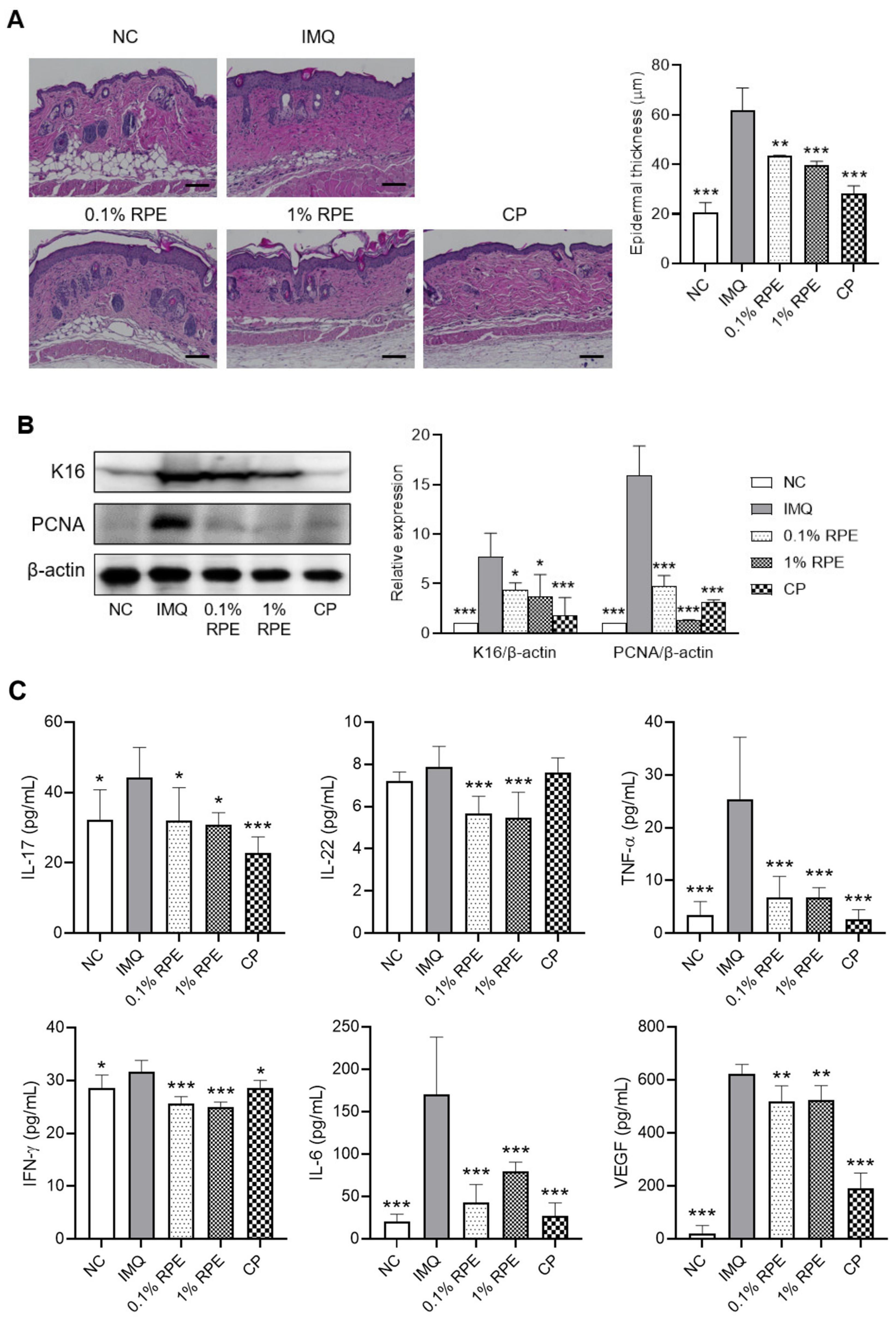
2.2. Effects of RPE on Epidermal Hyperplasia and the Expression of Proliferation Markers and Inflammatory Cytokines in IMQ-Induced Mice
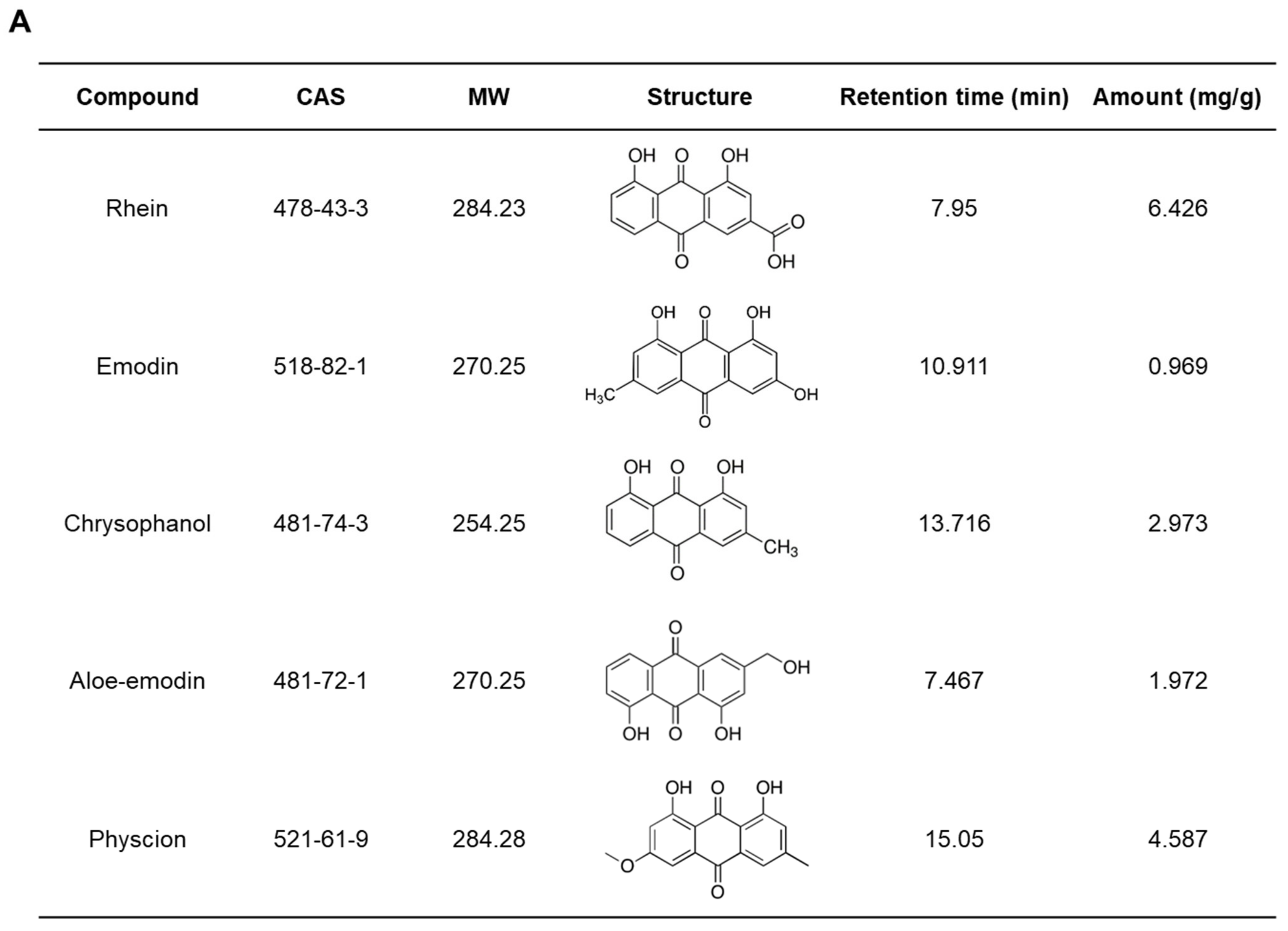
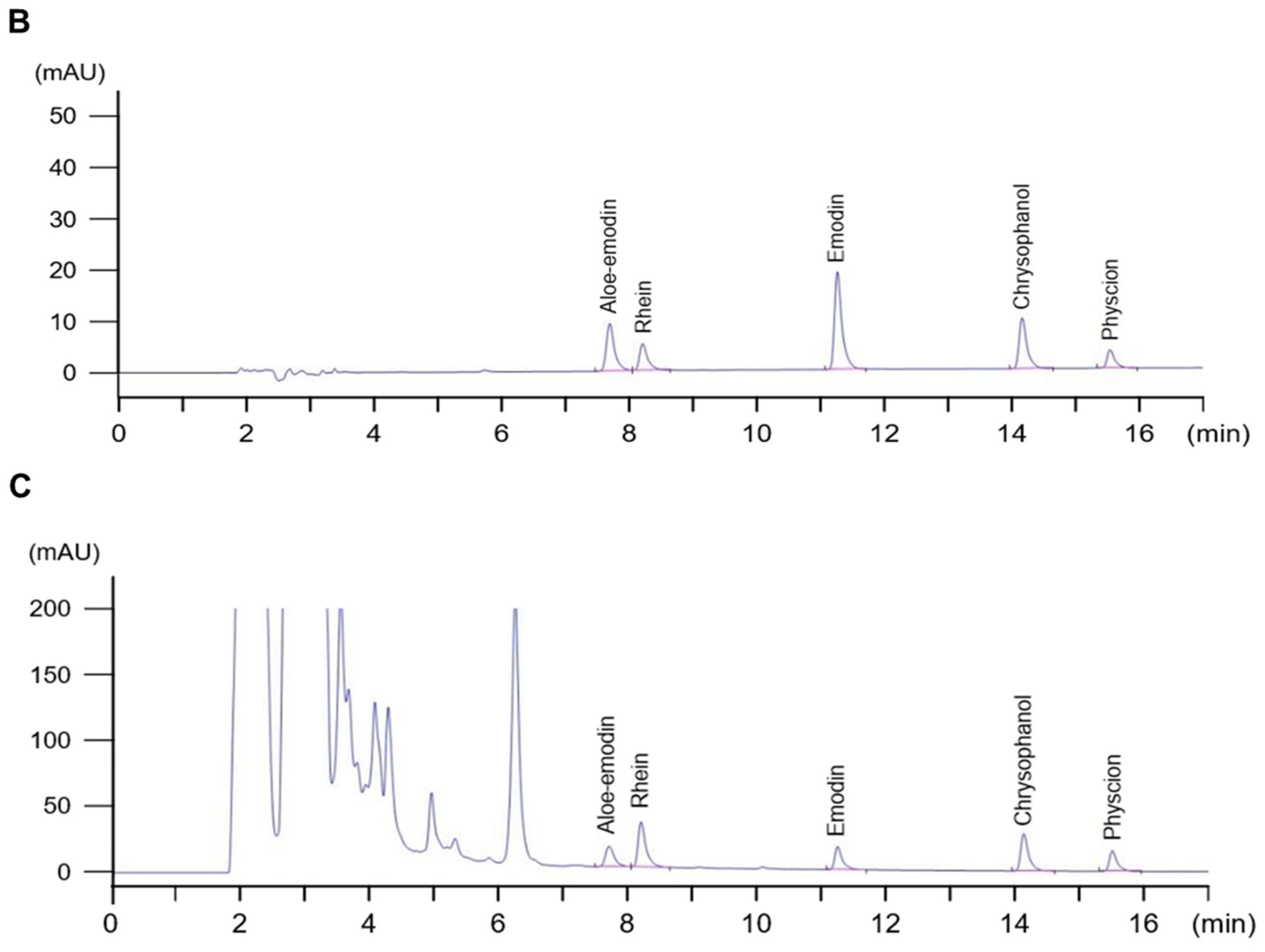
2.3. Quantitative Assessment of the Chemical Composition of RPE
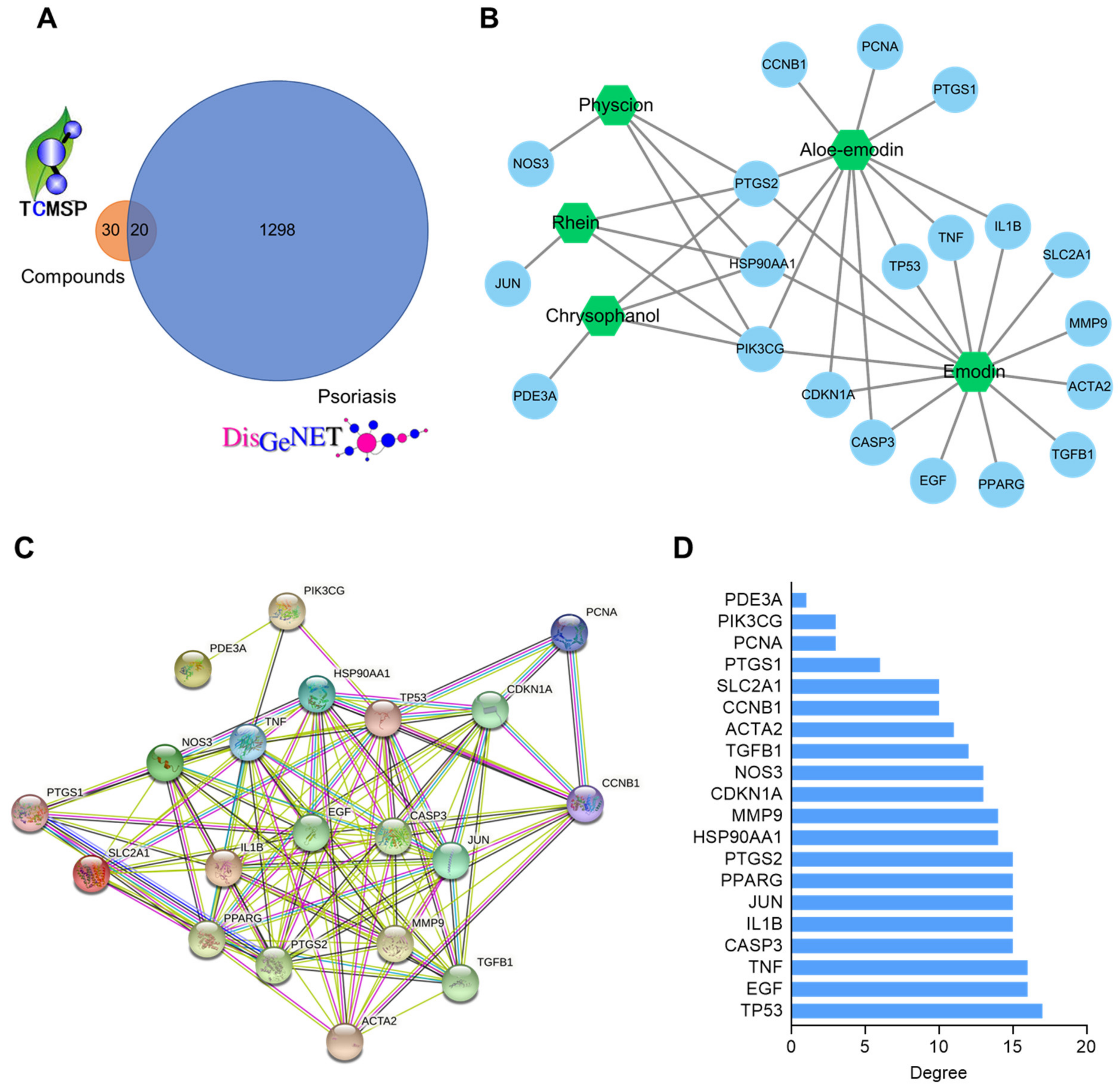
2.4. Compound–Target and Protein–Protein Interaction (PPI) Network Construction
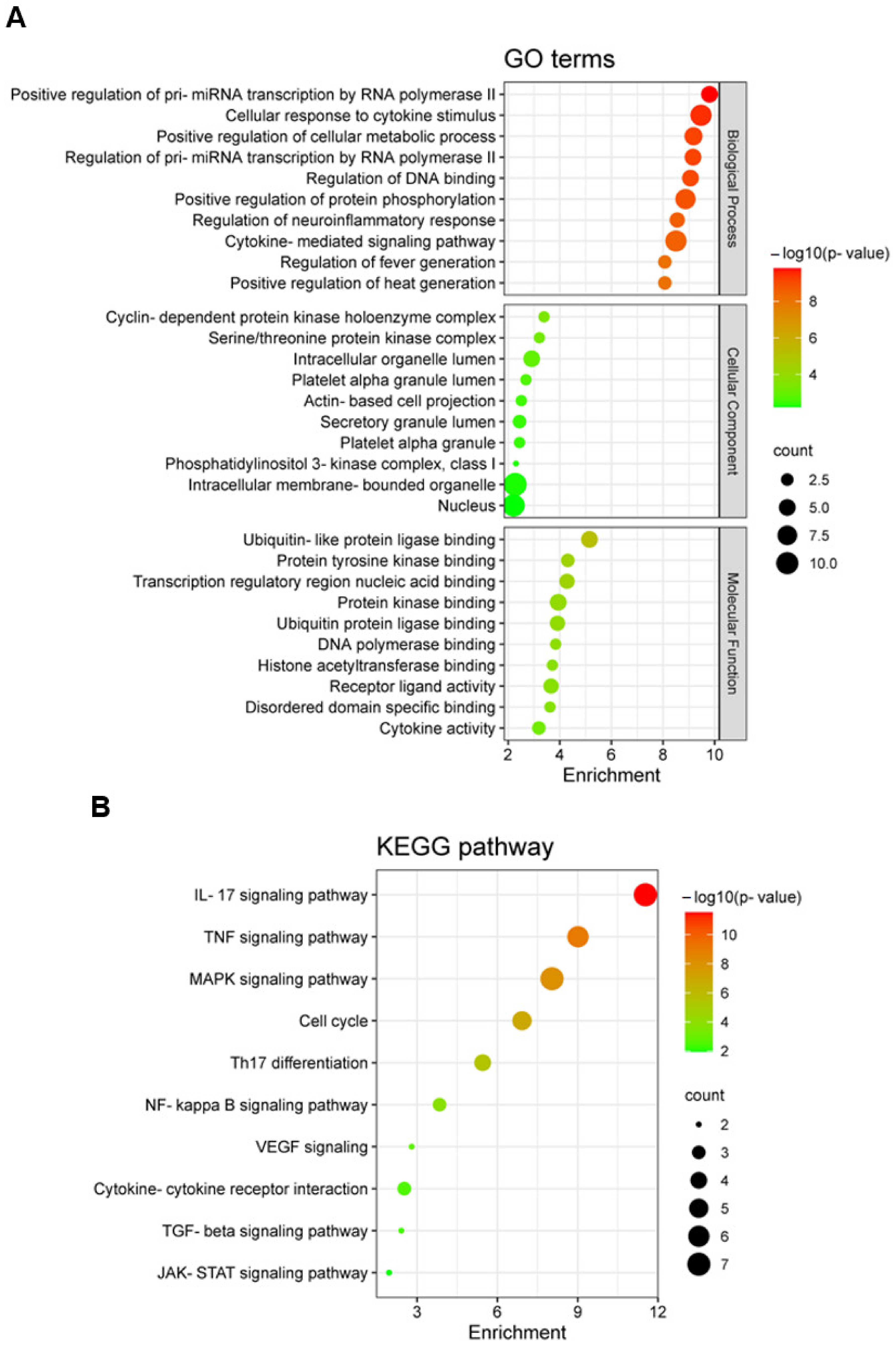
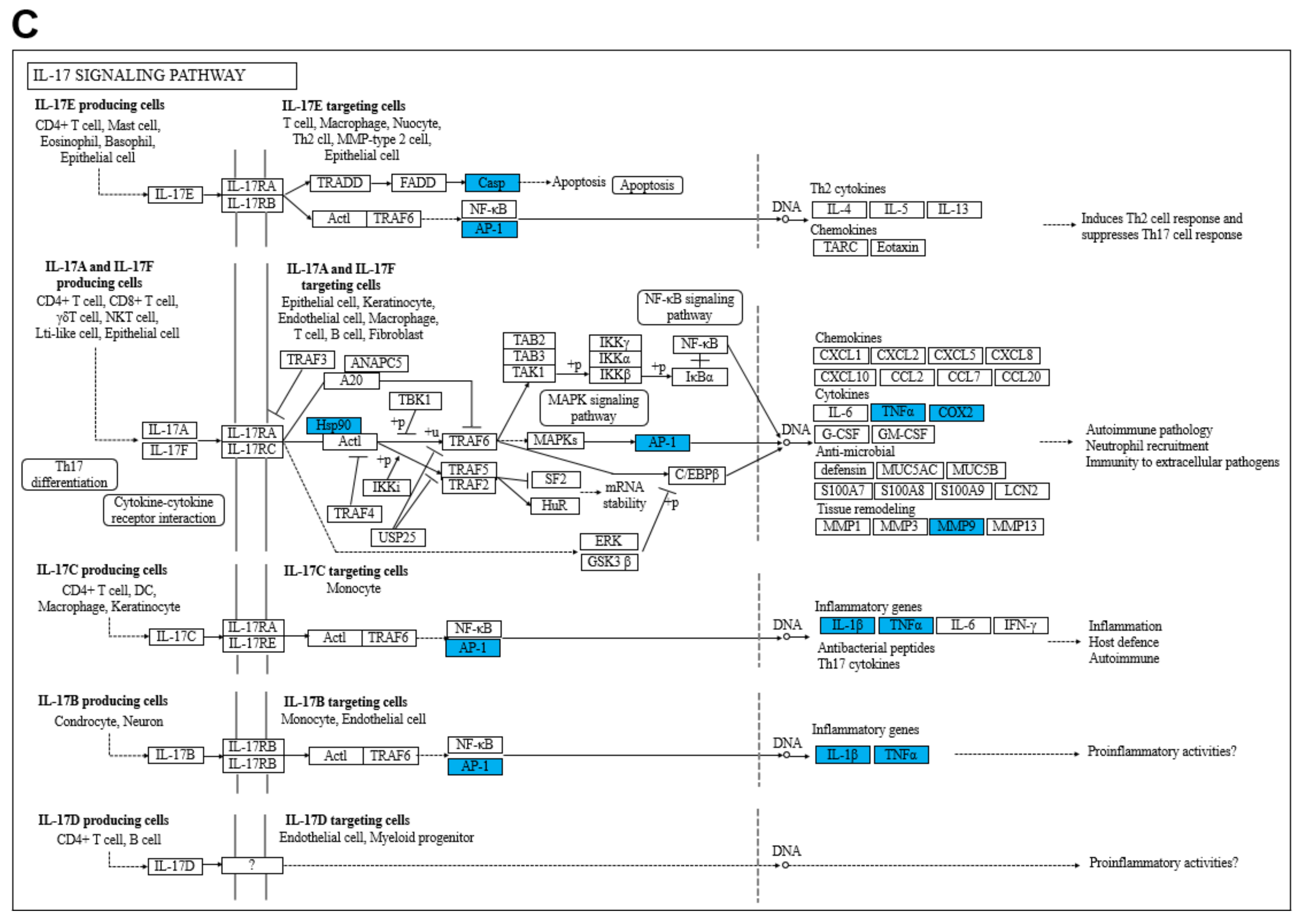
2.5. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis
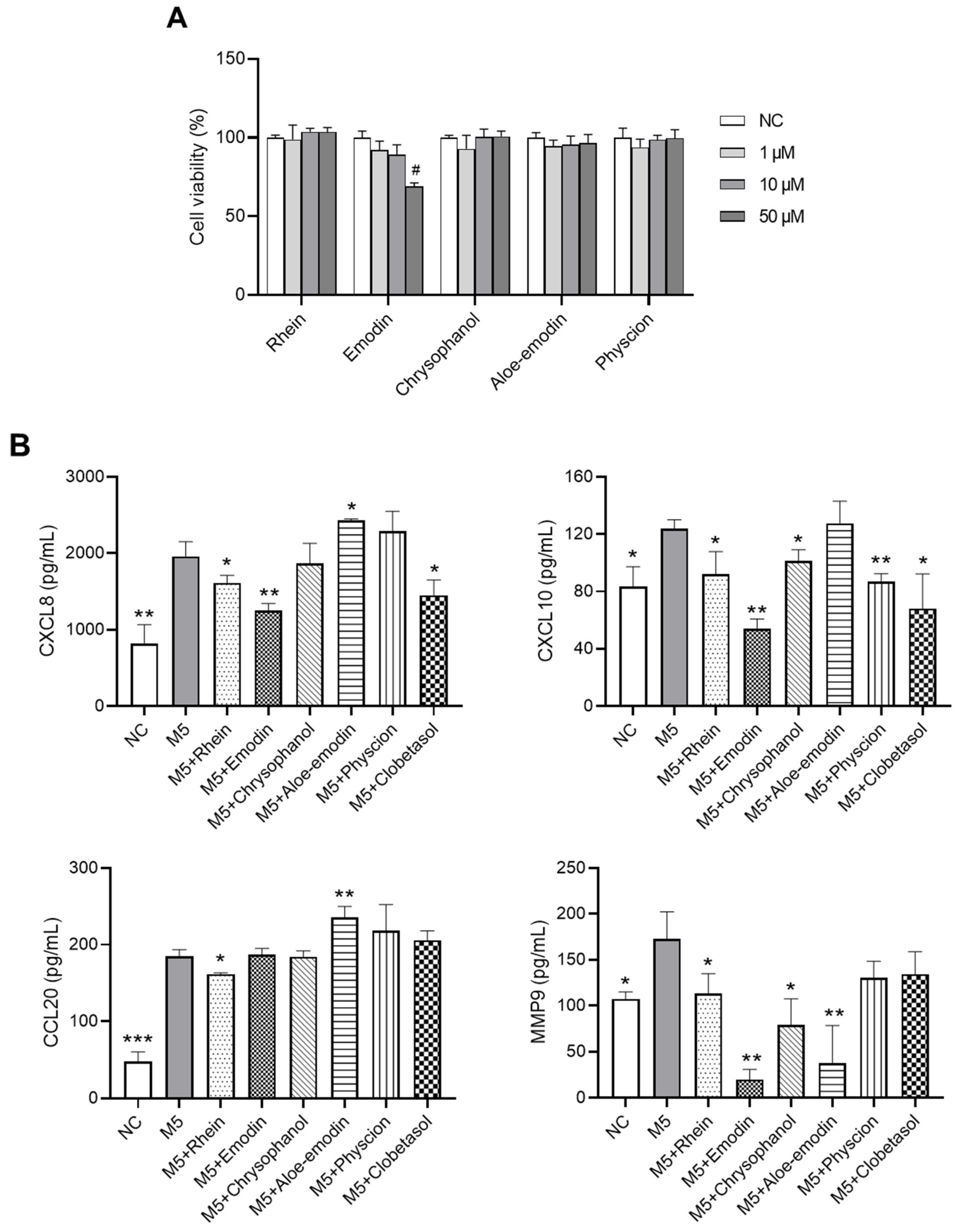
2.6. Effects of RPE Compounds on the Inflammatory Response in PI-Stimulated EL-4 Cells
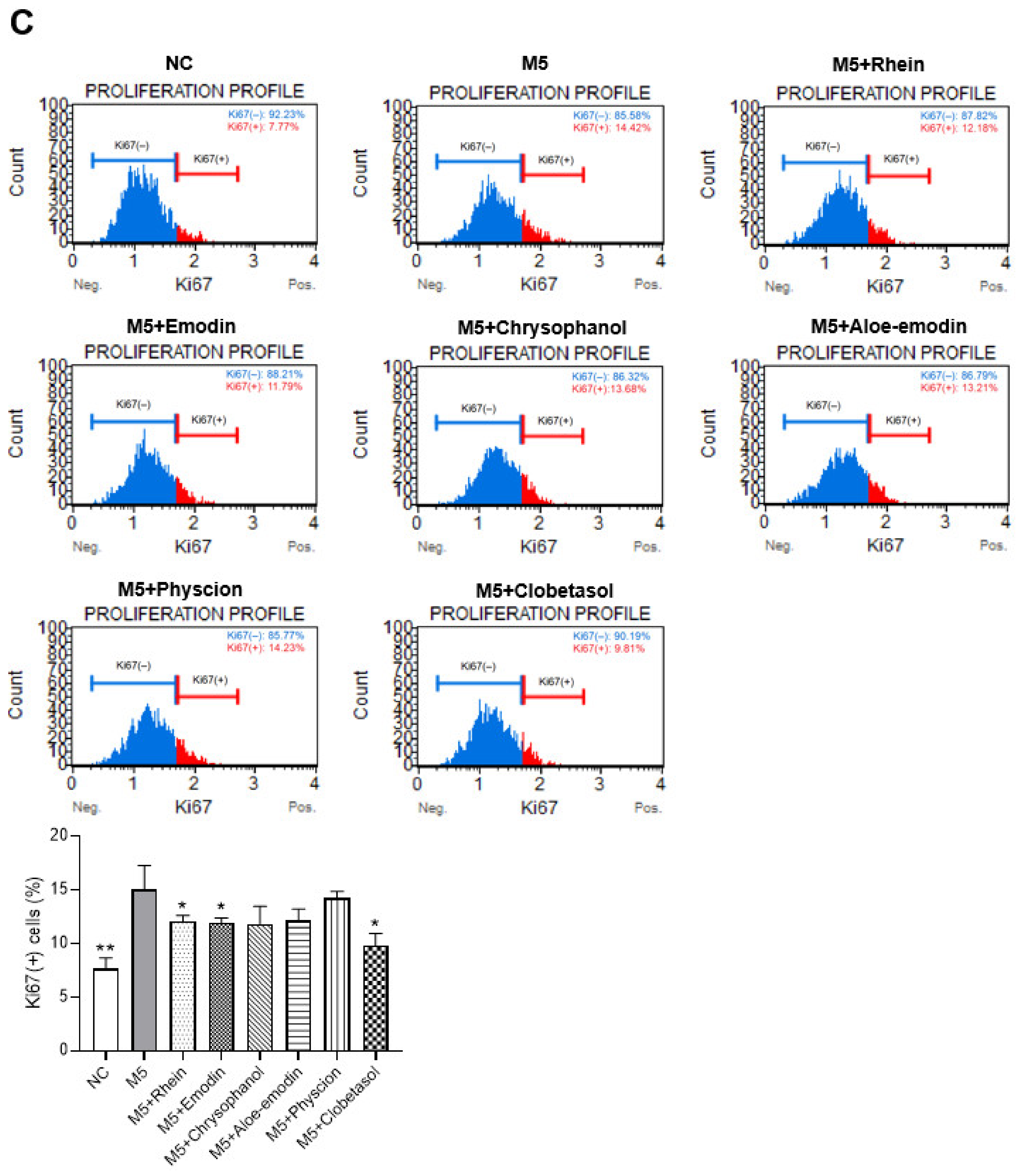
2.7. Effects of RPE Compounds on the Inflammatory Response and Proliferation in M5-Stimulated HaCaT Cells
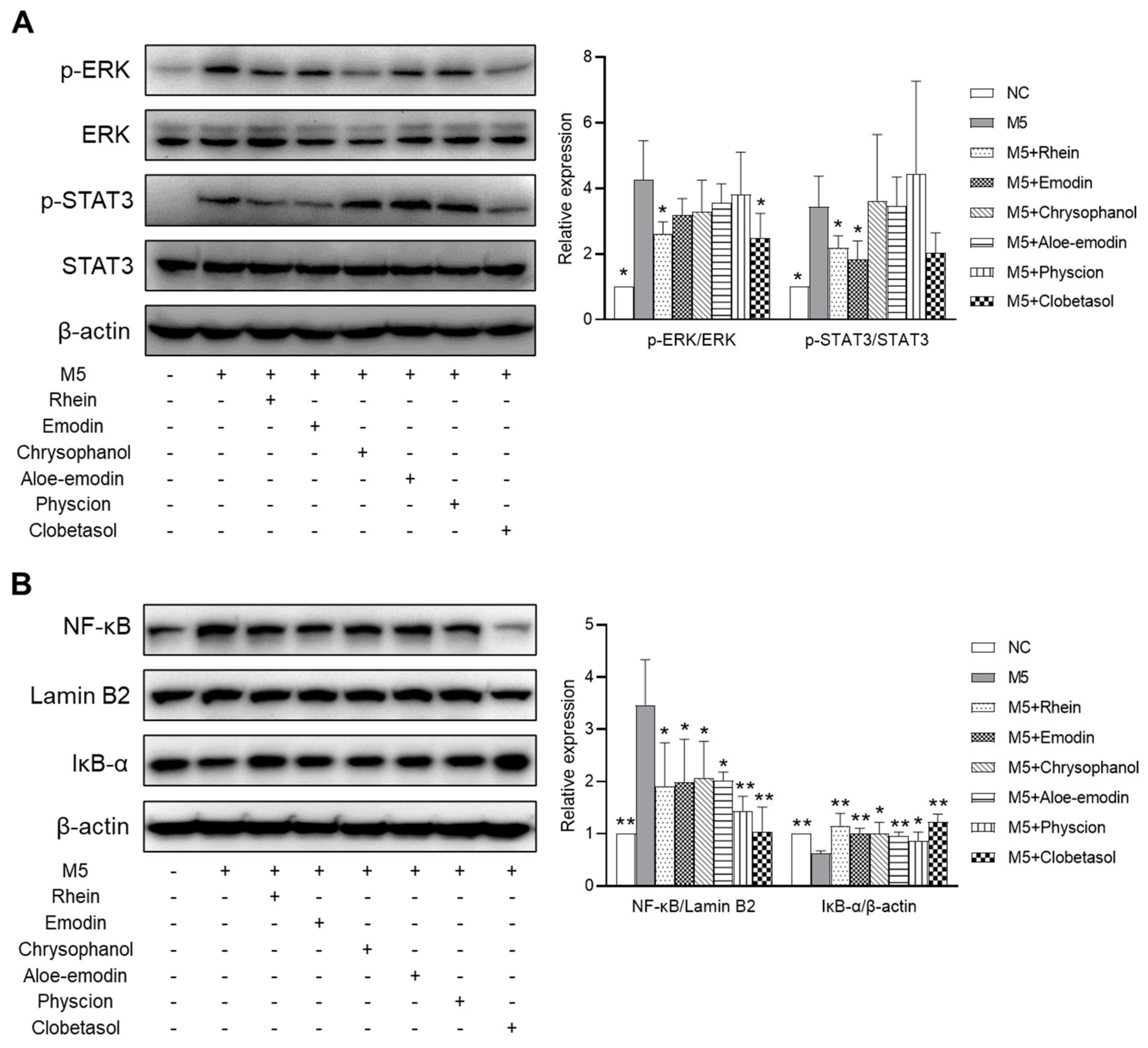
2.8. Effects of RPE Compounds on the Activation of the ERK, STAT3, and NF-κB Signaling Pathways in M5-Stimulated HaCaT Cells
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Animal Grouping and Treatments
4.3. Histological Assessment
4.4. High-Performance Liquid Chromatography (HPLC) Analysis
4.5. Network Pharmacology
4.6. Cell Culture
4.7. Cell Viability
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. Ki67 Proliferation Assay
4.10. Western Blot Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Nickoloff, B.J.; Nestle, F.O. Recent insights into the immunopathogenesis of psoriasis provide new therapeutic opportunities. J. Clin. Investig. 2004, 113, 1664–1675. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Debbaneh, M.G.; Liao, W. Genetic Epidemiology of Psoriasis. Curr. Dermatol. Rep. 2014, 3, 61–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, S.; Liu, X.; Fang, L.; Chen, X.; Guo, T.; Zhang, J. The cytokine milieu in the interplay of pathogenic Th1/Th17 cells and regulatory T cells in autoimmune disease. Cell Mol. Immunol. 2010, 7, 182–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liuzzo, G.; Trotta, F.; Pedicino, D. Interleukin-17 in atherosclerosis and cardiovascular disease: The good, the bad, and the unknown. Eur. Heart J. 2013, 34, 556–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albanesi, C.; Madonna, S.; Gisondi, P.; Girolomoni, G. The Interplay Between Keratinocytes and Immune Cells in the Pathogenesis of Psoriasis. Front. Immunol. 2018, 9, 1549. [Google Scholar] [CrossRef] [Green Version]
- Uva, L.; Miguel, D.; Pinheiro, C.; Antunes, J.; Cruz, D.; Ferreira, J.; Filipe, P. Mechanisms of action of topical corticosteroids in psoriasis. Int. J. Endocrinol. 2012, 2012, 561018. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Chen, J.; Kuai, L.; Zhang, Y.; Ding, X.; Luo, Y.; Ru, Y.; Xing, M.; Li, H.; Sun, X.; et al. Chinese Herbal Medicine for Psoriasis: Evidence From 11 High-Quality Randomized Controlled Trials. Front. Pharmacol. 2021, 11, 599433. [Google Scholar] [CrossRef]
- Lai, F.; Zhang, Y.; Xie, D.-P.; Mai, S.-T.; Weng, Y.-N.; Du, J.-D.; Wu, G.-P.; Zheng, J.-X.; Han, Y. A Systematic Review of Rhubarb (a Traditional Chinese Medicine) Used for the Treatment of Experimental Sepsis. Evid. Based Complement. Alternat. Med. 2015, 2015, 131283. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Miao, X. The topical application of Queyin tincture on psoriasis. Liaoning J. Trad. Chin. Med. 2004, 31, 394. [Google Scholar]
- Wang, J.; Zhu, M.; Xiang, L.; Xiao, Y. Chinese medicinal bath for psoriasis vulgaris. J. Chin. Phys. 2002, 4, 96–97. [Google Scholar]
- Shraibom, N.; Madaan, A.; Joshi, V.; Verma, R.; Chaudhary, A.; Mishra, G.; Awasthi, A.; Singh, A.T.; Jaggi, M. Evaluation of in vitro anti-psoriatic activity of a novel polyherbal formulation by multiparametric analysis. Antiinflamm. Antiallergy Agents Med. Chem. 2017, 16, 94–111. [Google Scholar] [CrossRef]
- Cheon, M.-S.; Yoon, T.-S.; Choi, G.-Y.; Kim, S.-J.; Lee, A.; Moon, B.-C.; Choo, B.-K.; Kim, H.-K. Comparative study of extracts from rhubarb on inflammatory activity in RAW 264.7 cells. Korean J. Med. Crop Sci. 2009, 17, 109–114. [Google Scholar]
- Zhou, P.; Zhang, J.; Xu, Y.; Zhang, P.; Xiao, Y.; Liu, Y. Simultaneous quantification of anthraquinone glycosides, aglycones, and glucuronic acid metabolites in rat plasma and tissues after oral administration of raw and steamed rhubarb in blood stasis rats by UHPLC-MS/MS. J. Sep. Sci. 2022, 45, 529–541. [Google Scholar] [CrossRef]
- Yang, T.Z.; Liu, Y.; Liu, Y.Y.; Ding, X.F.; Chen, J.X.; Kou, M.J.; Zou, X.J. The Use of Rheum palmatum L. In the Treatment of Acute Respiratory Distress Syndrome: A Meta-Analysis of Randomized, Controlled Trials. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 334–347. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.Z.; Wang, J.S.; Wang, P.; Jiang, C.Y.; Deng, B.X.; Li, P.; Zhao, Y.M.; Liu, W.L.; Qu, X.; Chen, W.W.; et al. Distribution and development of the TCM syndromes in psoriasis vulgaris. J. Tradit. Chin. Med. 2009, 29, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Qin, W.; Wu, L.; Yang, B.; Wang, Q.; Kuang, H.; Cheng, G. A review of Chinese medicine for the treatment of psoriasis: Principles, methods and analysis. Chin. Med. 2021, 16, 138. [Google Scholar] [CrossRef]
- Meng, S.; Lin, Z.; Wang, Y.; Wang, Z.; Li, P.; Zheng, Y. Psoriasis therapy by Chinese medicine and modern agents. Chin. Med. 2018, 13, 16. [Google Scholar] [CrossRef] [Green Version]
- Choi, T.Y.; Jun, J.H.; Park, B.; Lee, J.A.; You, S.S.; Jung, J.Y.; Lee, M.S. Concept of blood stasis in Chinese medical textbooks: A systematic review. Eur. J. Integr. Med. 2016, 8, 158–164. [Google Scholar] [CrossRef]
- Li, W.; Man, X.Y.; Chen, J.Q.; Zhou, J.; Cai, S.Q.; Zheng, M. Targeting VEGF/VEGFR in the Treatment of Psoriasis. Discov. Med. 2014, 18, 97–104. [Google Scholar]
- Guilloteau, K.; Paris, I.; Pedretti, N.; Boniface, K.; Juchaux, F.; Huguier, V.; Guillet, G.; Bernard, F.X.; Lecron, J.C.; Morel, F. Skin Inflammation Induced by the Synergistic Action of IL-17A, IL-22, Oncostatin M, IL-1α, and TNF-α Recapitulates Some Features of Psoriasis. J. Immunol. 2010, 184, 5263–5270. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, R.; Chugh, S.; Bansal, S. General measures and quality of life issues in psoriasis. Indian Dermatol. Online J. 2016, 7, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Gao, S.; Han, W.; Gao, Z.; Wei, Y.; Wu, G.; Qiqiu, W.; Chen, L.; Feng, Y.; Yue, S.; et al. Potential effects and mechanisms of Chinese herbal medicine in the treatment of psoriasis. J. Ethnopharmacol. 2022, 294, 115275. [Google Scholar] [CrossRef] [PubMed]
- Van der Fits, L.; Mourits, S.; Voerman, J.S.A.; Kant, M.; Boon, L.; Laman, J.D.; Cornelissen, F.; Mus, A.M.; Florencia, E.; Prens, E.P.; et al. Imiquimod-Induced Psoriasis-like Skin Inflammation in Mice Is Mediated via the IL-23/IL-17 Axis. J. Immunol. 2009, 182, 5836–5845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuellar, M.J.; Giner, R.M.; Recio, M.C.; Manez, S.; Rios, J.L. Topical anti-inflammatory activity of some Asian medicinal plants used in dermatological disorders. Fitoterapia 2001, 72, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef]
- Austin, L.M.; Ozawa, M.; Kikuchi, T.; Walters, I.B.; Krueger, J.G. The majority of epidermal T cells in psoriasis vulgaris lesions can produce type 1 cytokines, interferon-γ, interleukin-2, and tumor necrosis factor-α, defining TC1 (Cytotoxic T Lymphocyte) and TH1 effector populations: 1 a type 1 differentiation bias is also measured in circulating blood T cells in psoriatic patients. J. Investig. Dermatol. 1999, 113, 752–759. [Google Scholar]
- Moos, S.; Mohebiany, A.N.; Waisman, A.; Kurschus, F.C. Imiquimod-Induced Psoriasis in Mice Depends on the IL-17 Signaling of Keratinocytes. J. Investig. Dermatol. 2019, 139, 1110–1117. [Google Scholar] [CrossRef] [Green Version]
- Benhadou, F.; Mintoff, D.; del Marmol, V. Psoriasis: Keratinocytes or Immune Cells—Which Is the Trigger? Dermatology 2019, 235, 91–100. [Google Scholar] [CrossRef]
- Tonel, G.; Conrad, C. Interplay between keratinocytes and immune cells-Recent insights into psoriasis pathogenesis. Int. J. Biochem. Cell B. 2009, 41, 963–968. [Google Scholar] [CrossRef]
- Li, Q.; Laumonnier, Y.; Syrovets, T.; Simmet, T. Recruitment of CCR6-expressing Th17 cells by CCL20 secreted from plasmin-stimulated macrophages. Acta Biochim. Biophys. Sin. 2013, 45, 593–600. [Google Scholar] [CrossRef] [Green Version]
- Mezentsev, A.; Nikolaev, A.; Bruskin, S. Matrix metalloproteinases and their role in psoriasis. Gene 2014, 540, 1–10. [Google Scholar] [CrossRef]
- Gao, J.; Chen, F.; Fang, H.; Mi, J.; Qi, Q.; Yang, M. Daphnetin inhibits proliferation and inflammatory response in human HaCaT keratinocytes and ameliorates imiquimod-induced psoriasis-like skin lesion in mice. Biol. Res. 2020, 53, 48. [Google Scholar] [CrossRef]
- Li, C.; Xiao, L.; Jia, J.; Li, F.; Wang, X.; Duan, Q.; Jing, H.; Yang, P.; Chen, C.; Wang, Q.; et al. Cornulin Is Induced in Psoriasis Lesions and Promotes Keratinocyte Proliferation via Phosphoinositide 3-Kinase/Akt Pathways. J. Investig. Dermatol. 2019, 139, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Wu, N.; Duan, Q.; Yang, H.; Wang, X.; Yang, P.; Zhang, M.; Liu, J.; Liu, Z.; Shao, Y. C10orf99 contributes to the development of psoriasis by promoting the proliferation of keratinocytes. Sci. Rep. 2018, 8, 8590. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.H.; Si, X.Q. Rapamycin ameliorates psoriasis by regulating the expression and methylation levels of tropomyosin via ERK1/2 and mTOR pathways in vitro and in vivo. Exp. Dermatol. 2018, 27, 1112–1119. [Google Scholar] [CrossRef]
- Miyoshi, K.; Takaishi, M.; Nakajima, K.; Ikeda, M.; Kanda, T.; Tarutani, M.; Iiyama, T.; Asao, N.; DiGiovanni, J.; Sano, S. Stat3 as a Therapeutic Target for the Treatment of Psoriasis: A Clinical Feasibility Study with STA-21, a Stat3 Inhibitor. J. Investig. Dermatol. 2011, 131, 108–117. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Yu, P.; Liu, M.; Deng, Y.; Dong, Y.; Liu, Q.; Zhang, J.; Wu, T. ERK inhibitor JSI287 alleviates imiquimod-induced mice skin lesions by ERK/IL-17 signaling pathway. Int. Immunopharmacol. 2019, 66, 236–241. [Google Scholar] [CrossRef]
- Goldminz, A.M.; Au, S.C.; Kim, N.; Gottlieb, A.B.; Lizzul, P.F. NF-κB: An essential transcription factor in psoriasis. J. Dermatol. Sci. 2013, 69, 89–94. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Gaynor, R.B. IκB kinases: Key regulators of the NF-κB pathway. Trends Biochem. Sci. 2004, 29, 72–79. [Google Scholar] [CrossRef]
- Moorchung, N.; Kulaar, J.S.; Chatterjee, M.; Vasudevan, B.; Tripathi, T.; Dutta, V. Role of NF-κB in the pathogenesis of psoriasis elucidated by its staining in skin biopsy specimens. Int. J. Dermatol. 2014, 53, 570–574. [Google Scholar] [CrossRef]
- Andrés, R.M.; Montesinos, M.C.; Navalón, P.; Payá, M.; Terencio, M.C. NF-κB and STAT3 inhibition as a therapeutic strategy in psoriasis: In vitro and in vivo effects of BTH. J. Investig. Dermatol. 2013, 133, 2362–2371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imura, C.; Ueyama, A.; Sasaki, Y.; Shimizu, M.; Furue, Y.; Tai, N.; Tsujii, K.; Katayama, K.; Okuno, T.; Shichijo, M. A novel RORγt inhibitor is a potential therapeutic agent for the topical treatment of psoriasis with low risk of thymic aberrations. J. Dermatol. Sci. 2019, 93, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Rai, V.K.; Sinha, P.; Yadav, K.S.; Shukla, A.; Saxena, A.; Bawankule, D.U.; Tandon, S.; Khan, F.; Chanotiya, C.S.; Yadav, N.P. Anti-psoriatic effect of Lavandula angustifolia essential oil and its major components linalool and linalyl acetate. J. Ethnopharmacol. 2020, 261, 113127. [Google Scholar] [CrossRef] [PubMed]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y.; et al. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 2014, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Piñero, J.; Bravo, À.; Queralt-Rosinach, N.; Gutiérrez-Sacristán, A.; Deu-Pons, J.; Centeno, E.; García-García, J.; Sanz, F.; Furlong, L.I. DisGeNET: A comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res. 2017, 45, D833–D839. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Syafrizayanti; Betzen, C.; Hoheisel, J.D.; Kastelic, D. Methods for analyzing and quantifying protein–protein interaction. Expert Rev. Proteomics 2014, 11, 107–120. [Google Scholar] [CrossRef]
- Timalsina, P.; Charles, K.; Mondal, A.M. STRING PPI score to characterize protein subnetwork biomarkers for human diseases and pathways. In Proceedings of the 2014 IEEE International Conference on Bioinformatics and Bioengineering, Boca Raton, FL, USA, 10–12 November 2014. [Google Scholar]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, L.T.H.; Ahn, S.-H.; Shin, H.-M.; Yang, I.-J. Anti-Psoriatic Effect of Rheum palmatum L. and Its Underlying Molecular Mechanisms. Int. J. Mol. Sci. 2022, 23, 16000. https://doi.org/10.3390/ijms232416000
Nguyen LTH, Ahn S-H, Shin H-M, Yang I-J. Anti-Psoriatic Effect of Rheum palmatum L. and Its Underlying Molecular Mechanisms. International Journal of Molecular Sciences. 2022; 23(24):16000. https://doi.org/10.3390/ijms232416000
Chicago/Turabian StyleNguyen, Ly Thi Huong, Sang-Hyun Ahn, Heung-Mook Shin, and In-Jun Yang. 2022. "Anti-Psoriatic Effect of Rheum palmatum L. and Its Underlying Molecular Mechanisms" International Journal of Molecular Sciences 23, no. 24: 16000. https://doi.org/10.3390/ijms232416000
APA StyleNguyen, L. T. H., Ahn, S. -H., Shin, H. -M., & Yang, I. -J. (2022). Anti-Psoriatic Effect of Rheum palmatum L. and Its Underlying Molecular Mechanisms. International Journal of Molecular Sciences, 23(24), 16000. https://doi.org/10.3390/ijms232416000