
Opioids and Sepsis: Elucidating the Role of the Microbiome and microRNA-146

Abstract
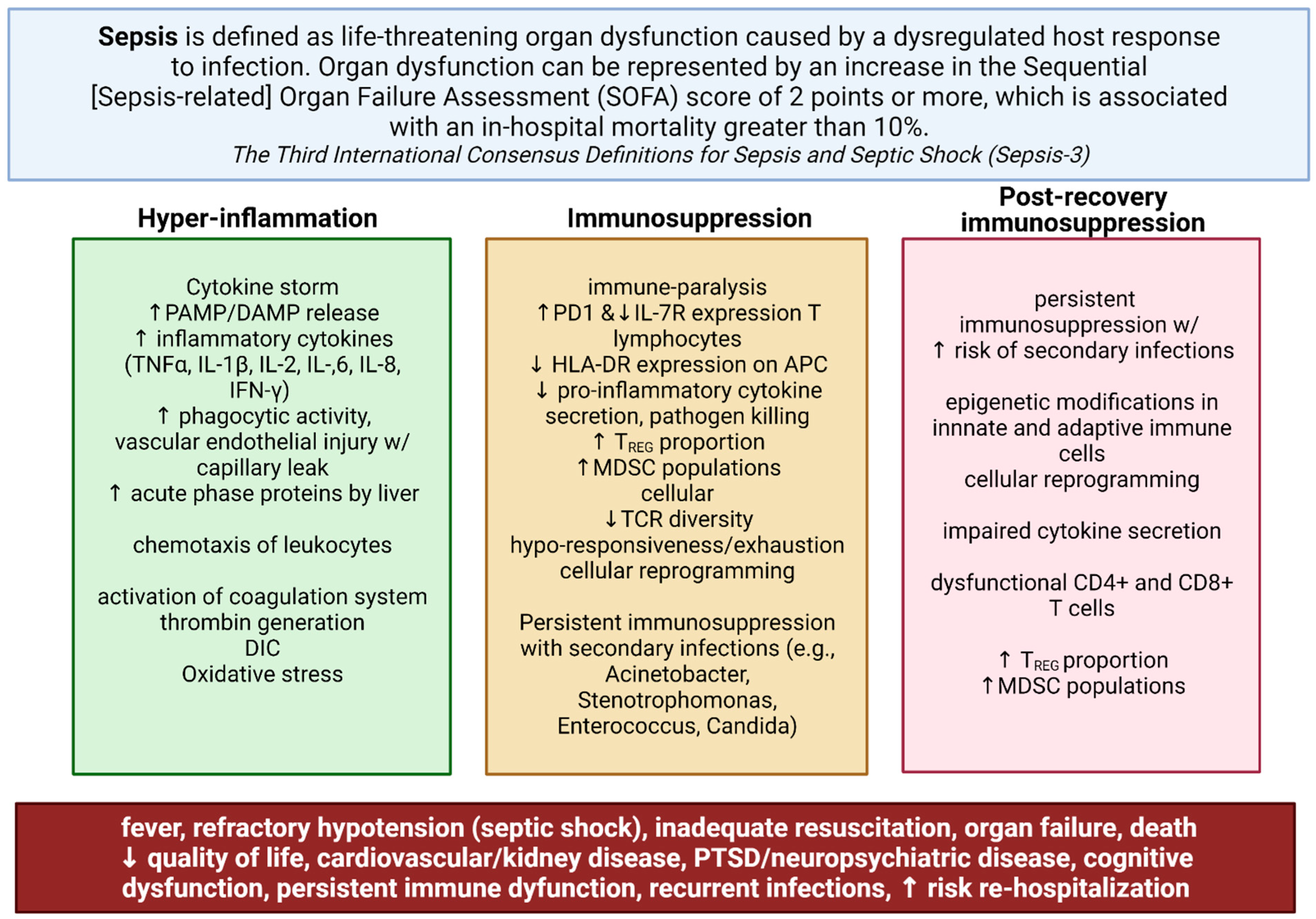
:1. Introduction
2. The Role of the Gut Microbiota in Sepsis
2.1. Intestinal Microbial Homeostasis and Resistance to Pathogen Colonization
2.2. Gut Microbial Disruption by Sepsis: A Bi-Directional Relationship
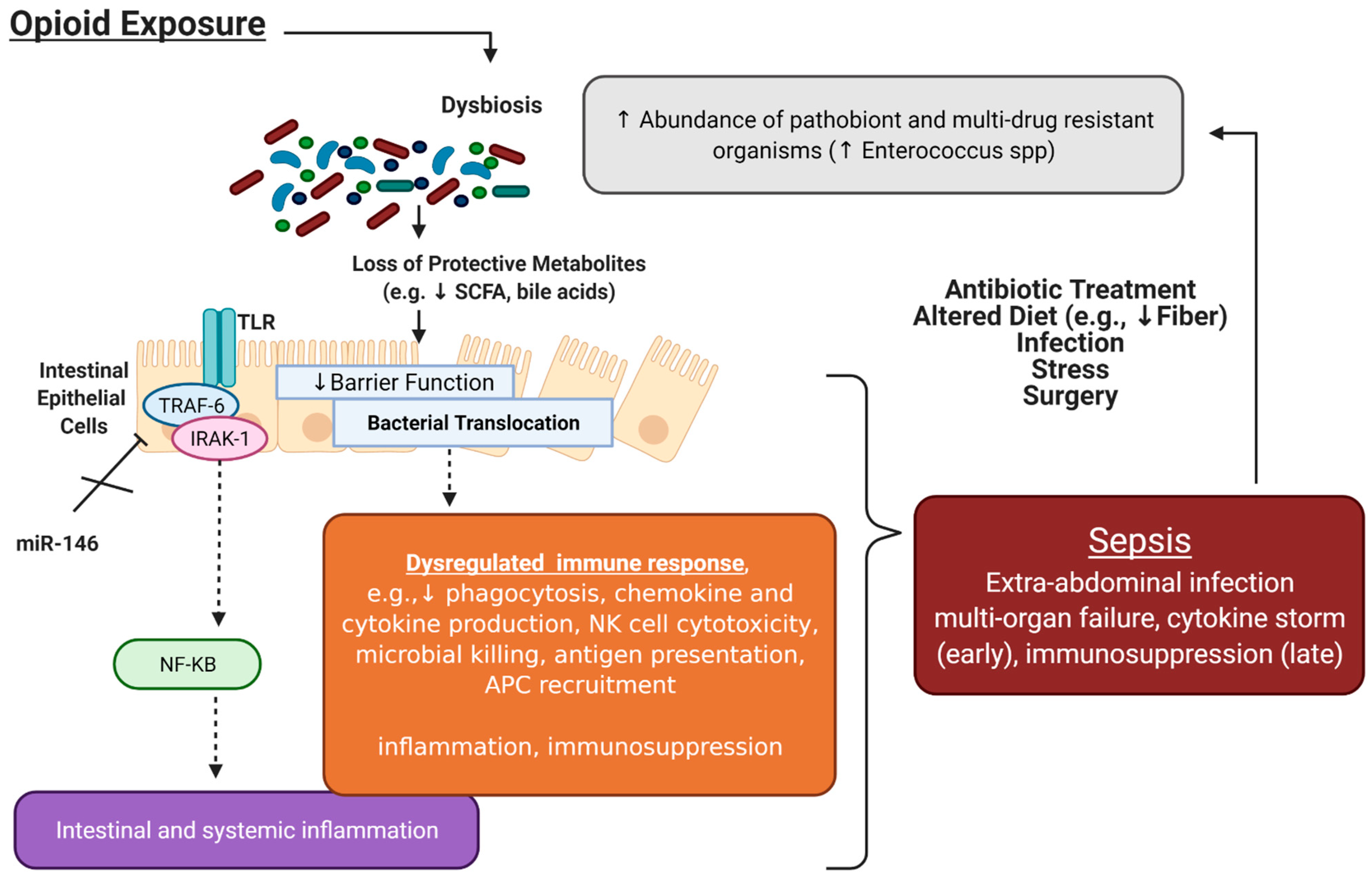
3. Interactions between Opioid Use and Sepsis: The Role of the Microbiome and microRNAs
3.1. Opioids Increase the Risk of Sepsis in Clinical and Preclinical Models
3.2. Opioids Induce Gut Microbial Dysbiosis, Which Is Associated with Increased Sepsis Risk
3.3. Opioid Modulation of Immune Cells may Increase Sepsis Risk
4. MicroRNA Modulation of Sepsis
5. Therapeutic Modulation of Sepsis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The third international consensus definitions for sepsis and septic shock (sepsis-3). JAMA J. Am. Med. Assoc. 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Rhee, C.; Dantes, R.; Epstein, L.; Murphy, D.J.; Seymour, C.W.; Iwashyna, T.J.; Kadri, S.S.; Angus, D.C.; Danner, R.L.; Fiore, A.E.; et al. Incidence and trends of sepsis in US hospitals using clinical vs claims data, 2009–2014. JAMA J. Am. Med. Assoc. 2017, 318, 1241–1249. [Google Scholar] [CrossRef]
- Hotchkiss, R.S.; Monneret, G.; Payen, D. Sepsis-induced immunosuppression: From cellular dysfunctions to immunotherapy. Nat. Rev. Immunol. 2013, 13, 862–874. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, M.; Marik, P.E. Management of sepsis during the early “golden hours”. J. Emerg. Med. 2006, 31, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Angus, D.C.; Linde-Zwirble, W.T.; Lidicker, J.; Clermont, G.; Carcillo, J.; Pinsky, M.R. Epidemiology of severe sepsis in the United States: Analysis of incidence, outcome, and associated costs of care. Crit. Care Med. 2001, 29, 1303–1310. [Google Scholar] [CrossRef]
- Haak, B.W.; Wiersinga, W.J. The role of the gut microbiota in sepsis. Lancet Gastroenterol. Hepatol. 2017, 2, 135–143. [Google Scholar] [CrossRef]
- Prescott, H.C.; Osterholzer, J.J.; Langa, K.M.; Angus, D.C.; Iwashyna, T.J. Late mortality after sepsis: Propensity matched cohort study. BMJ 2016, 353, i2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gigliotti, J.C.; Okusa, M.D. The Spleen: The Forgotten Organ in Acute Kidney Injury of Critical Illness. Nephron Clin. Pract. 2014, 127, 153–157. [Google Scholar] [CrossRef] [Green Version]
- Funahashi, Y.; Kato, N.; Masuda, T.; Nishio, F.; Kitai, H.; Ishimoto, T.; Kosugi, T.; Tsuboi, N.; Matsuda, N.; Maruyama, S.; et al. miR-146a targeted to splenic macrophages prevents sepsis-induced multiple organ injury. Lab. Investig. 2019, 99, 1130–1142. [Google Scholar] [CrossRef]
- Williams, D.L.; Ha, T.; Li, C.; Kalbfleisch, J.H.; Schweitzer, J.; Vogt, W.; Browder, I.W. Modulation of tissue Toll-like receptor 2 and 4 during the early phases of polymicrobial sepsis correlates with mortality. Crit. Care Med. 2003, 31, 1808–1818. [Google Scholar] [CrossRef]
- Gao, M.; Ha, T.; Zhang, X.; Liu, L.; Wang, X.; Kelley, J.; Gao, X.; Williams, D.; Li, C. TLR3 plays a central role in cardiac dysfunction during polymicrobial sepsis. Crit. Care Med. 2012, 40, 2390–2399. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, N.; Hattori, Y.; Takahashi, Y.; Nishihira, J.; Jesmin, S.; Kobayashi, M.; Gando, S. Therapeutic effect of in vivo transfection of transcription factor decoy to NF-κB on septic lung in mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 287, 1248–1255. [Google Scholar] [CrossRef] [Green Version]
- Van Der Poll, T.; Van De Veerdonk, F.L.; Scicluna, B.P.; Netea, M.G. The immunopathology of sepsis and potential therapeutic targets. Nat. Rev. Immunol. 2017, 17, 407–420. [Google Scholar] [CrossRef]
- Alrawashdeh, M.; Klompas, M.; Kimmel, S.; Larochelle, M.R.; Gokhale, R.H.; Dantes, R.B.; Hoots, B.; Hatfield, K.M.; Reddy, S.C.; Fiore, A.E.; et al. Epidemiology, Outcomes, and Trends of Patients With Sepsis and Opioid-Related Hospitalizations in U.S. Hospitals. Crit. Care Med. 2021, 49, 2102–2111. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.P.; Wall, M.J.; Novack, L.; Marshall, J.; Hsu, D.J.; Howell, M.D. The critical care crisis of opioid overdoses in the United States. Ann. Am. Thorac. Soc. 2017, 14, 1803–1809. [Google Scholar] [CrossRef]
- SAMSHA. Key Substance Use and Mental Health Indicators in the United States: Results from the 2019 National Survey on Drug Use and Health; HHS Publ. No. PEP19-5068, NSDUH Ser. H-54; Center for Behavioral Health Statistics and Quality, Substance Abuse and Mental Health Services Administration: Rockville, MD, USA, 2019; Volume 170, pp. 51–58.
- Berterame, S.; Erthal, J.; Thomas, J.; Fellner, S.; Vosse, B.; Clare, P.; Hao, W.; Johnson, D.T.; Mohar, A.; Pavadia, J.; et al. Use of and barriers to access to opioid analgesics: A worldwide, regional, and national study. Lancet 2016, 387, 1644–1656. [Google Scholar] [CrossRef]
- Adelman, M.W.; Woodworth, M.H.; Langelier, C.; Busch, L.M.; Kempker, J.A.; Kraft, C.S.; Kraft, C.S.; Martin, G.S.; Martin, G.S. The gut microbiome’s role in the development, maintenance, and outcomes of sepsis. Crit. Care 2020, 24, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kullberg, R.F.J.; Wiersinga, W.J.; Haak, B.W. Gut microbiota and sepsis: From pathogenesis to novel treatments. Curr. Opin. Gastroenterol. 2021, 37, 578–585. [Google Scholar] [CrossRef]
- Miller, W.D.; Keskey, R.; Alverdy, J.C. Sepsis and the microbiome: A vicious cycle. J. Infect. Dis. 2021, 223, S264–S269. [Google Scholar] [CrossRef]
- Tourelle, K.M.; Boutin, S.; Weigand, M.A.; Schmitt, F.C.F. Sepsis and the human microbiome. Just another kind of organ failure? A review. J. Clin. Med. 2021, 10, 4831. [Google Scholar] [CrossRef]
- Wu, H.J.; Wu, E. The role of gut microbiota in immune homeostasis and autoimmunity. Gut Microbes 2012, 3, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Kamada, N.; Seo, S.U.; Chen, G.Y.; Núñez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef]
- Pamer, E.G. Resurrecting the intestinal microbiota to combat antibiotic- resistant pathogens. Science 2016, 352, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Venegas, D.P.; De La Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Schuijt, T.J.; van der Poll, T.; de Vos, W.M.; Wiersinga, W.J. The intestinal microbiota and host immune interactions in the critically ill. Trends Microbiol. 2013, 21, 221–229. [Google Scholar] [CrossRef]
- Ayres, J.S.; Trinidad, N.J.; Vance, R.E. Lethal inflammasome activation by a multidrug-resistant pathobiont upon antibiotic disruption of the microbiota. Nat. Med. 2012, 18, 799–806. [Google Scholar] [CrossRef] [Green Version]
- Lankelma, J.M.; Cranendonk, D.R.; Belzer, C.; De Vos, A.F.; De Vos, W.M.; Van Der Poll, T.; Wiersinga, W.J. Antibiotic-induced gut microbiota disruption during human endotoxemia: A randomised controlled study. Gut 2017, 66, 1623–1630. [Google Scholar] [CrossRef]
- Lankelma, J.M.; van Vught, L.A.; Belzer, C.; Schultz, M.J.; van der Poll, T.; de Vos, W.M.; Wiersinga, W.J. Critically ill patients demonstrate large interpersonal variation in intestinal microbiota dysregulation: A pilot study. Intensive Care Med. 2017, 43, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.; Marshall, J.; Anzueto, A.; Martin, C.D.; Gomersall, C. International Study of the Prevalence and Outcomes of Infection in Intensive Care Units. JAMA 2009, 302, 2323–2329. [Google Scholar] [CrossRef] [Green Version]
- Rampelli, S.; Candela, M.; Turroni, S.; Biagi, E.; Collino, S.; Toole, P.W.O.; Brigidi, P. Functional metagenomic profiling of intestinal microbiome in extreme ageing. Aging 2013, 5, 902–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agudelo-Ochoa, G.M.; Valdés-Duque, B.E.; Giraldo-Giraldo, N.A.; Jaillier-Ramírez, A.M.; Giraldo-Villa, A.; Acevedo-Castaño, I.; Yepes-Molina, M.A.; Barbosa-Barbosa, J.; Benítez-Paéz, A. Gut microbiota profiles in critically ill patients, potential biomarkers and risk variables for sepsis. Gut Microbes 2020, 12, 1707610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Keefe, S.J. Effect of fiber supplementation on the microbiota in critically ill patients. World J. Gastrointest. Pathophysiol. 2011, 2, 138. [Google Scholar] [CrossRef]
- Bassetti, M.; Bandera, A.; Gori, A. Therapeutic Potential of the Gut Microbiota in the Management of Sepsis. Crit. Care 2020, 24, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fay, K.T.; Klingensmith, N.J.; Chen, C.W.; Zhang, W.; Sun, Y.; Morrow, K.N.; Liang, Z.; Burd, E.M.; Ford, M.L.; Coopersmith, C.M. The gut microbiome alters immunophenotype and survival from sepsis. FASEB J. 2019, 33, 11258–11269. [Google Scholar] [CrossRef]
- Dickson, R.P. The microbiome and critical illness. Lancet Respir. Med. 2016, 4, 59–72. [Google Scholar] [CrossRef] [Green Version]
- Hyoju, S.K.; Zaborin, A.; Keskey, R.; Sharma, A.; Arnold, W.; Van Den Berg, F.; Kim, S.M.; Gottel, N.; Bethel, C.; Charnot-Katsikas, A.; et al. Mice fed an obesogenic western diet, administered antibiotics, and subjected to a sterile surgical procedure develop lethal septicemia with multidrug-resistant pathobionts. MBio 2019, 10, e00903-19. [Google Scholar] [CrossRef] [Green Version]
- Napier, B.A.; Andres-Terre, M.; Massis, L.M.; Hryckowian, A.J.; Higginbottom, S.K.; Cumnock, K.; Casey, K.M.; Haileselassie, B.; Lugo, K.A.; Schneider, D.S.; et al. Western diet regulates immune status and the response to LPS-driven sepsis independent of diet-associated microbiome. Proc. Natl. Acad. Sci. USA 2019, 116, 3688–3694. [Google Scholar] [CrossRef] [Green Version]
- Taur, Y.; Xavier, J.B.; Lipuma, L.; Ubeda, C.; Goldberg, J.; Gobourne, A.; Lee, Y.J.; Dubin, K.A.; Socci, N.D.; Viale, A.; et al. Intestinal domination and the risk of bacteremia in patients undergoing allogeneic hematopoietic stem cell transplantation. Clin. Infect. Dis. 2012, 55, 905–914. [Google Scholar] [CrossRef]
- Prescott, H.C.; Dickson, R.P.; Rogers, M.A.M.; Langa, K.M.; Iwashyna, T.J. Hospitalization type and subsequent severe sepsis. Am. J. Respir. Crit. Care Med. 2015, 192, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Baggs, J.; Jernigan, J.A.; Halpin, A.L.; Lauren, H.; Hatfield, K.M.; Mcdonald, L.C. Risk of Subsequent Sepsis within 90 Days of a Previous Hospital Stay by Type of Antibiotic Exposure. Clin. Infect. Dis 2018, 66, 1004–1012. [Google Scholar] [CrossRef] [Green Version]
- Freedberg, D.E.; Zhou, M.J.; Cohen, M.E.; Annavajhala, M.K.; Khan, S.; Moscoso, D.; Brooks, C.; Whittier, S.; Chong, D.H.; Uhlemann, A.-C.; et al. Pathogen colonization of the gastrointestinal microbiome at intensive care unit admission and risk for subsequent death or infection. Intensive Care Med. 2018, 44, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Cheng, M.; Li, J.; Zhang, P.; Fan, H.; Hu, Q.; Han, M.; Su, L.; He, H.; Tong, Y.; et al. Classification of the Gut Microbiota of Patients in Intensive Care Units During Development of Sepsis and Septic Shock. Genom. Proteom. Bioinform. 2020, 18, 696–707. [Google Scholar] [CrossRef] [PubMed]
- Wirbel, J.; Pyl, P.T.; Kartal, E.; Zych, K.; Kashani, A.; Milanese, A.; Fleck, J.S.; Voigt, A.Y.; Palleja, A.; Ponnudurai, R.; et al. Meta-analysis of fecal metagenomes reveals global microbial signatures that are specific for colorectal cancer. Nat. Med. 2019, 25, 679–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojima, M.; Motooka, D.; Shimizu, K.; Gotoh, K.; Shintani, A.; Yoshiya, K.; Nakamura, S.; Ogura, H.; Iida, T.; Shimazu, T. Metagenomic Analysis Reveals Dynamic Changes of Whole Gut Microbiota in the Acute Phase of Intensive Care Unit Patients. Dig. Dis. Sci. 2016, 61, 1628–1634. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, F.C.F.; Brenner, T.; Uhle, F.; Loesch, S.; Hackert, T.; Ulrich, A.; Hofer, S.; Dalpke, A.H.; Weigand, M.A.; Boutin, S. Gut microbiome patterns correlate with higher postoperative complication rates after pancreatic surgery. BMC Microbiol. 2019, 19, 42. [Google Scholar] [CrossRef] [Green Version]
- Gaither, J.R.; Goulet, J.L.; Becker, W.C.; Crystal, S.; Edelman, E.J.; Gordon, K.; Kerns, R.D.; Rimland, D.; Skanderson, M.; Justice, A.C.; et al. The Association Between Receipt of Guideline-Concordant Long-Term Opioid Therapy and All-Cause Mortality. J. Gen. Intern. Med. 2016, 31, 492–501. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Meng, J.; Lian, Q.; Chen, X.; Bauman, B.; Chu, H.; Segura, B.; Roy, S. Prescription opioids are associated with higher mortality in patients diagnosed with sepsis: A retrospective cohort study using electronic health records. PLoS ONE 2018, 13, e0190362. [Google Scholar] [CrossRef]
- Devlin, J.W.; Roberts, R.J. Pharmacology of Commonly Used Analgesics and Sedatives in the ICU: Benzodiazepines, Propofol, and Opioids. Crit. Care Clin. 2009, 25, 431–449. [Google Scholar] [CrossRef]
- Glattard, E.; Welters, I.D.; Lavaux, T.; Muller, A.H.; Laux, A.; Zhang, D.; Schmidt, A.R.; Delalande, F.; Laventie, B.J.; Dirrig-Grosch, S.; et al. Endogenous morphine levels are increased in sepsis: A partial implication of neutrophils. PLoS ONE 2010, 5, e8791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.; Banerjee, S.; Li, D.; Sindberg, G.M.; Wang, F.; Ma, J.; Roy, S. Opioid exacerbation of gram-positive sepsis, induced by gut microbial modulation, is rescued by IL-17A neutralization. Sci. Rep. 2015, 5, 10918. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Charboneau, R.G.; Barke, R.A. Morphine synergizes with lipopolysaccharide in a chronic endotoxemia model. J. Neuroimmunol. 1999, 95, 107–114. [Google Scholar] [CrossRef]
- Ocasio, F.M.; Jiang, Y.; House, S.D.; Chang, S.L. Chronic morphine accelerates the progression of lipopolysaccharide-induced sepsis to septic shock. J. Neuroimmunol. 2004, 149, 90–100. [Google Scholar] [CrossRef]
- Hilburger, M.E.; Adler, M.W.; Truant, A.L.; Meissler, J.J.; Satishchandran, V.; Rogers, T.J.; Eisenstein, T.K. Morphine induces sepsis in mice. J. Infect. Dis. 1997, 176, 183–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Barke, R.A.; Charboneau, R.; Roy, S. Morphine Impairs Host Innate Immune Response and Increases Susceptibility to Streptococcus pneumoniae Lung Infection. J. Immunol. 2005, 174, 426–434. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Barke, R.A.; Ma, J.; Charboneau, R.; Roy, S. Opiate abuse, innate immunity, and bacterial infectious diseases. Arch. Immunol. Ther. Exp. 2008, 56, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Ninkovic, J.; Banerjee, S.; Charboneau, R.; Das, S.; Dutta, R.; Kirchner, V.; Koodie, L.; Ma, J.; Meng, J. Opioid Drug Abuse and Modulation of Immune Function: Consequences in the Susceptibility to Opportunistic Infections. J. Neuroimmune Pharmacol. 2011, 6, 442–465. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Yu, H.; Ma, J.; Wang, J.; Banerjee, S.; Charboneau, R.; Barke, R.A.; Roy, S. Morphine Induces Bacterial Translocation in Mice by Compromising Intestinal Barrier Function in a TLR-Dependent Manner. PLoS ONE 2013, 8, e54040. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Meng, J.; Zhang, L.; Johnson, T.; Chen, C.; Roy, S. Morphine induces changes in the gut microbiome and metabolome in a morphine dependence model. Sci. Rep. 2018, 8, 3596. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Sindberg, G.; Wang, F.; Meng, J.; Sharma, U.; Zhang, L.; Dauer, P.; Chen, C.; Dalluge, J.; Johnson, T.; et al. Opioid-induced gut microbial disruption and bile dysregulation leads to gut barrier compromise and sustained systemic inflammation. Mucosal Immunol. 2016, 9, 1418–1428. [Google Scholar] [CrossRef] [Green Version]
- Greeneltch, K.M.; Haudenschild, C.C.; Keegan, A.D. The opioid antagonist naltrexone blocks acute endotoxic shock by inhibiting tumor necrosis factor- a production. Brain Behav. Immun. 2004, 18, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Babrowski, T.; Holbrook, C.; Moss, J.; Gottlieb, L.; Valuckaite, V.; Zaborin, A.; Poroyko, V.; Liu, D.C.; Zaborina, O.; Alverdy, J.C. Pseudomonas aeruginosa virulence expression is directly activated by morphine and is capable of causing lethal gut-derived sepsis in mice during chronic morphine administration. Ann. Surg. 2012, 255, 386–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and inflammation Yasmine. Cell 2015, 157, 121–141. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Meng, J.; Das, S.; Krishnan, A.; Haworth, J.; Charboneau, R.; Zeng, Y.; Ramakrishnan, S.; Roy, S. Morphine induced exacerbation of sepsis is mediated by tempering endotoxin tolerance through modulation of miR-146a. Sci. Rep. 2013, 3, 1977. [Google Scholar] [CrossRef]
- Plein, L.M.; Rittner, H.L. Opioids and the immune system – friend or foe. Br. J. Pharmacol. 2018, 175, 2717–2725. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, J.; Xue, Y.; Zhao, J.; Li, D.; Zhang, S.; Li, K.; Hou, Y.; Fan, H. GSKJ4 protects mice against early sepsis via reducing proinflammatory factors and up-regulating MiR-146a. Front. Immunol. 2018, 9, 2272. [Google Scholar] [CrossRef]
- Mehta, A.; Baltimore, D. MicroRNAs as regulatory elements in immune system logic. Nat. Rev. Immunol. 2016, 16, 279–294. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- Toyama, K.; Kiyosawa, N.; Watanabe, K.; Ishizuka, H. Identification of circulating miRNAs differentially regulated by opioid treatment. Int. J. Mol. Sci. 2017, 18, 1991. [Google Scholar] [CrossRef] [Green Version]
- Anzola, A.; González, R.; Gámez-Belmonte, R.; Ocón, B.; Aranda, C.J.; Martínez-Moya, P.; López-Posadas, R.; Hernández-Chirlaque, C.; Sánchez de Medina, F.; Martínez-Augustin, O. miR-146a regulates the crosstalk between intestinal epithelial cells, microbial components and inflammatory stimuli. Sci. Rep. 2018, 8, 17350. [Google Scholar] [CrossRef]
- Nahid, M.A.; Pauley, K.M.; Satoh, M.; Chan, E.K.L. miR-146a is critical for endotoxin-induced tolerance: Implication in innate immunity. J. Biol. Chem. 2009, 284, 34590–34599. [Google Scholar] [CrossRef] [Green Version]
- Taganov, K.D.; Boldin, M.P.; Chang, K.-J.; Baltimore, D. NF-B-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [Green Version]
- Saba, R.; Sorensen, D.L.; Booth, S.A. MicroRNA-146a: A dominant, negative regulator of the innate immune response. Front. Immunol. 2014, 5, 578. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, W.; Xia, B.; Zhang, M.; Wang, Y. MicroRNA-146a attenuates the development of morphine analgesic tolerance in a rat model. Neurol. Res. 2020, 42, 415–421. [Google Scholar] [CrossRef]
- Jiang, S. Recent findings regarding let-7 in immunity. Cancer Lett. 2018, 434, 130–131. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulos, D.; Hirsch, H.A.; Struhl, K. An Epigenetic Switch Involving NF-$κ$B, Lin28, Let-7 MicroRNA, and IL6 Links Inflammation to Cell Transformation. Cell 2009, 139, 693–706. [Google Scholar] [CrossRef] [Green Version]
- Dave, R.S.; Khalili, K. Morphine treatment of human monocyte-derived macrophages induces differential miRNA and protein expression: Impact on inflammation and oxidative stress in the central nervous system. J. Cell. Biochem. 2010, 110, 834–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.G.; Becattini, S.; Moody, T.U.; Shliaha, P.V.; Littmann, E.R.; Seok, R.; Gjonbalaj, M.; Eaton, V.; Fontana, E.; Amoretti, L.; et al. Microbiota-derived lantibiotic restores resistance against vancomycin-resistant Enterococcus. Nature 2019, 572, 665–669. [Google Scholar] [CrossRef]
- Sabharwala, H.; Cichona, C.; Ölschlägerb, T.A.; Sonnenbornc, U.; Schmidta, M.A. Interleukin-8, CXCL1, and MicroRNA miR-146a Responses to Probiotic Escherichia coli Nissle 1917 and Enteropathogenic E. coli in Human Intestinal Epithelial T84 and Monocytic THP-1 Cells after Apical or Basolateral Infection. Infect. Immun. 2016, 84, 2482–2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Dou, H.; Li, X.; Zhao, X.; Li, Y.; Liu, D.; Ji, J.; Liu, F.; Ding, L.; Ni, Y.; et al. Exosomal miR-146a Contributes to the Enhanced Therapeutic Efficacy of Interleukin-1β-Primed Mesenchymal Stem Cells Against Sepsis. Stem Cells 2017, 35, 1208–1221. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yan, Y.; Meng, J.; Girotra, M.; Ramakrishnan, S.; Roy, S. Immune Modulation Mediated by Extracellular Vesicles of Intestinal Organoids is Disrupted by Opioids. Mucosal Immunol. 2021, 14, 887–898. [Google Scholar] [CrossRef]
- Robbins, P.D.; Morelli, A.E. Regulation of immune responses by extracellular vesicles. Nat. Rev. Immunol. 2014, 14, 195–208. [Google Scholar] [CrossRef] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essandoh, K.; Fan, G.C. Role of extracellular and intracellular microRNAs in sepsis. Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 2155–2162. [Google Scholar] [CrossRef] [Green Version]
- Vergadi, E.; Vaporidi, K.; Tsatsanis, C. Regulation of endotoxin tolerance and compensatory anti-inflammatory response syndrome by non-coding RNAs. Front. Immunol. 2018, 9, 2705. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Zhang, J.; Cao, M.; Han, S.; Liu, Y.; Wang, K.; Han, F.; Li, X.; Jia, Y.; Wang, X.; et al. MicroRNA-146a protects against LPS-induced organ damage by inhibiting Notch1 in macrophage. Int. Immunopharmacol. 2018, 63, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Zhang, L.; Fan, X.; Dong, X.; Zhang, Z.; Fan, W. MicroRNA-146a improves sepsis-induced cardiomyopathy by regulating the TLR-4/NF-κB signaling pathway. Exp. Ther. Med. 2019, 779–785. [Google Scholar] [CrossRef] [Green Version]
- Du, C.T.; Gao, W.; Ma, K.; Yu, S.X.; Li, N.; Yan, S.Q.; Zhou, F.H.; Liu, Z.Z.; Chen, W.; Lei, L.C.; et al. MicroRNA-146a deficiency protects against listeria monocytogenes infection by modulating the gut microbiota. Int. J. Mol. Sci. 2018, 19, 993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Da Cunha, A.P.; Rezende, R.M.; Cialic, R.; Wei, Z.; Bry, L.; Comstock, L.E.; Gandhi, R.; Weiner, H.L. The Host Shapes the Gut Microbiota via Fecal MicroRNA. Cell Host Microbe 2016, 19, 32–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero, S.; Kim, S.; Carter, R.A.; Leiner, I.M.; Sušac, B.; Miller, L.; Kim, G.J.; Ling, L.; Pamer, E.G. Cooperating Commensals Restore Colonization Resistance to Vancomycin-Resistant Enterococcus faecium. Cell Host Microbe 2017, 21, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, H.C.; Chen, C.; Zeng, J.; Wang, Q.; Zheng, L.; Yu, H. Du Differential expression of plasma miR-146a in sepsis patients compared with non-sepsis-SIRS patients. Exp. Ther. Med. 2013, 5, 1101–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Y.; Li, J.; Cai, Y.; Xie, Y.; Ma, G.; Li, Y.; Chen, Y.; Liu, G.; Zhao, B.; Cui, L.; et al. The Functional Polymorphisms of miR-146a Are Associated with Susceptibility to Severe Sepsis in the Chinese Population. Mediators Inflamm. 2014, 2014. [Google Scholar] [CrossRef]
- Oostdijk, E.A.N.; De Smet, A.M.G.A.; Blok, H.E.M.; Thieme Groen, E.S.; Van Asselt, G.J.; Benus, R.F.J.; Bernards, S.A.T.; Frénay, I.H.M.E.; Jansz, A.R.; De Jongh, B.M.; et al. Ecological effects of selective decontamination on resistant gram-negative bacterial colonization. Am. J. Respir. Crit. Care Med. 2010, 181, 452–457. [Google Scholar] [CrossRef]
- Huttner, B.; Haustein, T.; Uçkay, I.; Renzi, G.; Stewardson, A.; Schaerrer, D.; Agostinho, A.; Andremont, A.; Schrenzel, J.; Pittet, D.; et al. Decolonization of intestinal carriage of extended-spectrum Β-lactamase-producing Enterobacteriaceae with oral colistin and neomycin: A randomized, double-blind, placebo-controlled trial. J. Antimicrob. Chemother. 2013, 68, 2375–2382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Xu, K.; Gui, Q.; Chen, Y.; Chen, D.; Yang, Y. Probiotic pre-administration reduces mortality in a mouse model of cecal ligation and puncture-induced sepsis. Exp. Ther. Med. 2016, 12, 1836–1842. [Google Scholar] [CrossRef] [Green Version]
- Panigrahi, P.; Parida, S.; Nanda, N.C.; Satpathy, R.; Pradhan, L.; Chandel, D.S.; Baccaglini, L.; Mohapatra, A.; Mohapatra, S.S.; Misra, P.R.; et al. A randomized synbiotic trial to prevent sepsis among infants in rural India. Nature 2017, 548, 407–412. [Google Scholar] [CrossRef]
- Costeloe, K.; Hardy, P.; Juszczak, E.; Wilks, M.; Millar, M.R. Bifidobacterium breve BBG-001 in very preterm infants: A randomised controlled phase 3 trial. Lancet 2016, 387, 649–660. [Google Scholar] [CrossRef]
- Morowitz, M.J.; Di Caro, V.; Pang, D.; Cummings, J.; Firek, B.; Rogers, M.B.; Ranganathan, S.; Clark, R.S.B.; Aneja, R.K. Dietary supplementation with nonfermentable fiber alters the gut microbiota and confers protection in murine models of sepsis. Crit. Care Med. 2017, 45, e516–e523. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, C.; Tang, C.; He, Q.; Zhao, X.; Li, N.; Li, J. Successful treatment of severe sepsis and diarrhea after vagotomy utilizing fecal microbiota transplantation: A case report. Crit. Care 2015, 19, 37. [Google Scholar] [CrossRef] [Green Version]
- Gopalsamy, S.N.; Sherman, A.; Woodworth, M.H.; Lutgring, J.D.; Kraft, C.S. Fecal Microbiota Transplant for Multidrug-Resistant Organism Decolonization Administered during Septic Shock. Infect. Control. Hosp. Epidemiol. 2018, 39, 490–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Yang, J.; Wang, J.; Yang, Y.; Huang, J.; Gong, H.; Cui, H.; Chen, D. Successful treatment with fecal microbiota transplantation in patients with multiple organ dysfunction syndrome and diarrhea following severe sepsis. Crit. Care 2016, 20, 332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Link, A.; Becker, V.; Goel, A.; Wex, T.; Malfertheiner, P. Feasibility of fecal microRNAs as novel biomarkers for pancreatic cancer. PLoS ONE 2012, 7, e42933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, F.E.; Jeffries, C.D.; Vos, P.W.; Flake, G.; Nuovo, G.J.; Sinar, D.R.; Naziri, W.; Marcuard, S.P. Diagnostic microRNA markers for screening sporadic human colon cancer and active ulcerative colitis in stool and tissue. Cancer Genom. Proteom. 2009, 6, 281–295. [Google Scholar]



{kind=link}
{kind=link}
| Opioid Modulation of miR-146a | ||
|---|---|---|
| Model | Key Findings | Reference |
| Murine | morphine ↓ endotoxin/LPS induced miR-146a and 155 expression in macrophages; only miR-146a overexpression, not miR-155, abrogates morphine-mediated hyper-inflammation; antagonizing miR-146a ↑ severity of morphine-mediated hyper-inflammation | [11] |
| Murine | ↓ miR-146a in spinal cord in a morphine-tolerance rat model; lentiviral overexpression of miR-146a fails to attenuate the development of morphine analgesic tolerance | [80] |
| Murine | miR-146a levels unchanged in the extracellular vesicles after morphine treatment | [81] |
| Human | ↑ plasma Let-7 family and miR-146a expression after 24 h of hydromorphone or oxycodone treatment | [82] |
| Human | morphine ↑ miR-146a expression in human monocyte-derived macrophages | [19] |
| Role of miR-146a in Sepsis and SIRS Models | ||
|---|---|---|
| Model | Key Findings | Reference |
| Murine | ↑ miR-146a in macrophages protective against LPS-induced organ damage | [10] |
| Murine | miR-146a ↓ hyper-inflammation and prevents multiple organ failure in sepsis; miR-146a ↓ production of pro-inflammatory cytokines in RAW264.7 macrophage cells after LPS stimulation | [28] |
| Murine | miR-146a ↓ development of sepsis-induced cardiomyopathy by regulating TLR4/NF-κβ signaling pathway | [91] |
| Murine | miR-146a-deficient mice more resistant to L. monocytogenes infection via modulation of the gut microbiota | [22] |
| Murine | IL-1β-primed mesenchymal stem cells, which produce miR-146a, ↓ hyper-inflammation in murine sepsis models | [86] |
| Murine | inhibition of JMJD3, which modulates miR-146a transcription, protects mice against early septic death | [65] |
| Human | ↓ plasma miR-146a in sepsis patients compared to non-sepsis SIRS patients | [92] |
| Human | mutations in the miR-146a gene associated with an ↑ risk of sepsis | [84] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abu, Y.; Vitari, N.; Yan, Y.; Roy, S. Opioids and Sepsis: Elucidating the Role of the Microbiome and microRNA-146. Int. J. Mol. Sci. 2022, 23, 1097. https://doi.org/10.3390/ijms23031097
Abu Y, Vitari N, Yan Y, Roy S. Opioids and Sepsis: Elucidating the Role of the Microbiome and microRNA-146. International Journal of Molecular Sciences. 2022; 23(3):1097. https://doi.org/10.3390/ijms23031097
Chicago/Turabian StyleAbu, Yaa, Nicolas Vitari, Yan Yan, and Sabita Roy. 2022. "Opioids and Sepsis: Elucidating the Role of the Microbiome and microRNA-146" International Journal of Molecular Sciences 23, no. 3: 1097. https://doi.org/10.3390/ijms23031097
APA StyleAbu, Y., Vitari, N., Yan, Y., & Roy, S. (2022). Opioids and Sepsis: Elucidating the Role of the Microbiome and microRNA-146. International Journal of Molecular Sciences, 23(3), 1097. https://doi.org/10.3390/ijms23031097