
Deletion of Specific Conserved Motifs from the N-Terminal Domain of αB-Crystallin Results in the Activation of Chaperone Functions

 ,
,  ,
,  and
and 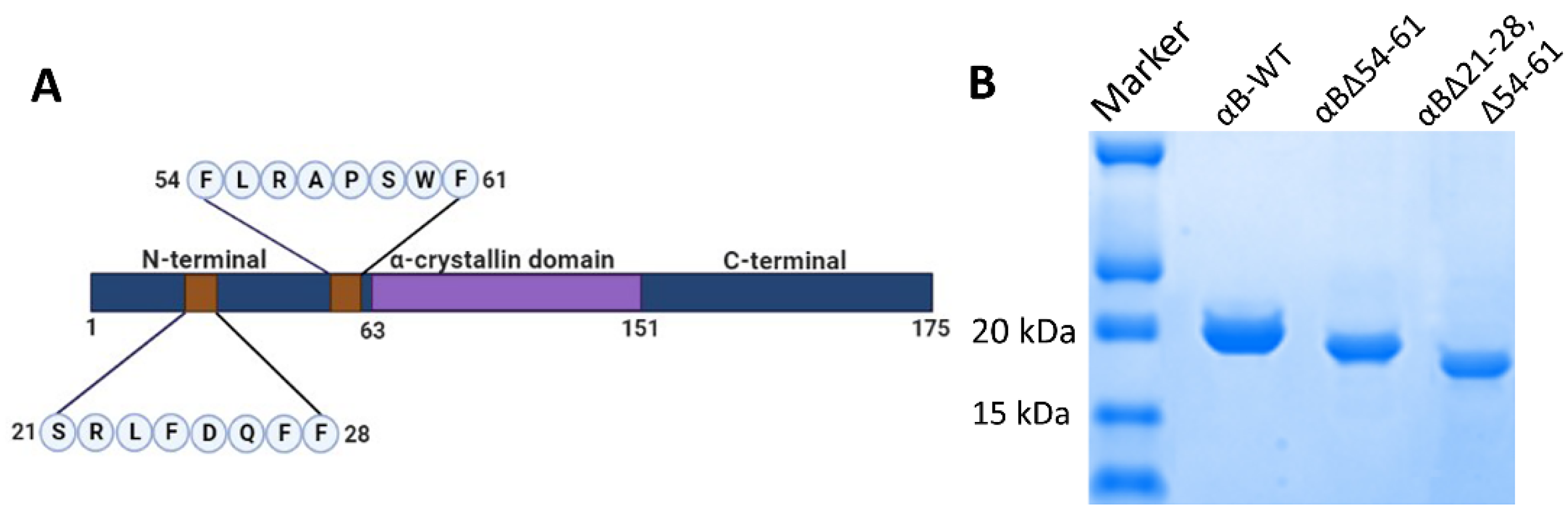{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
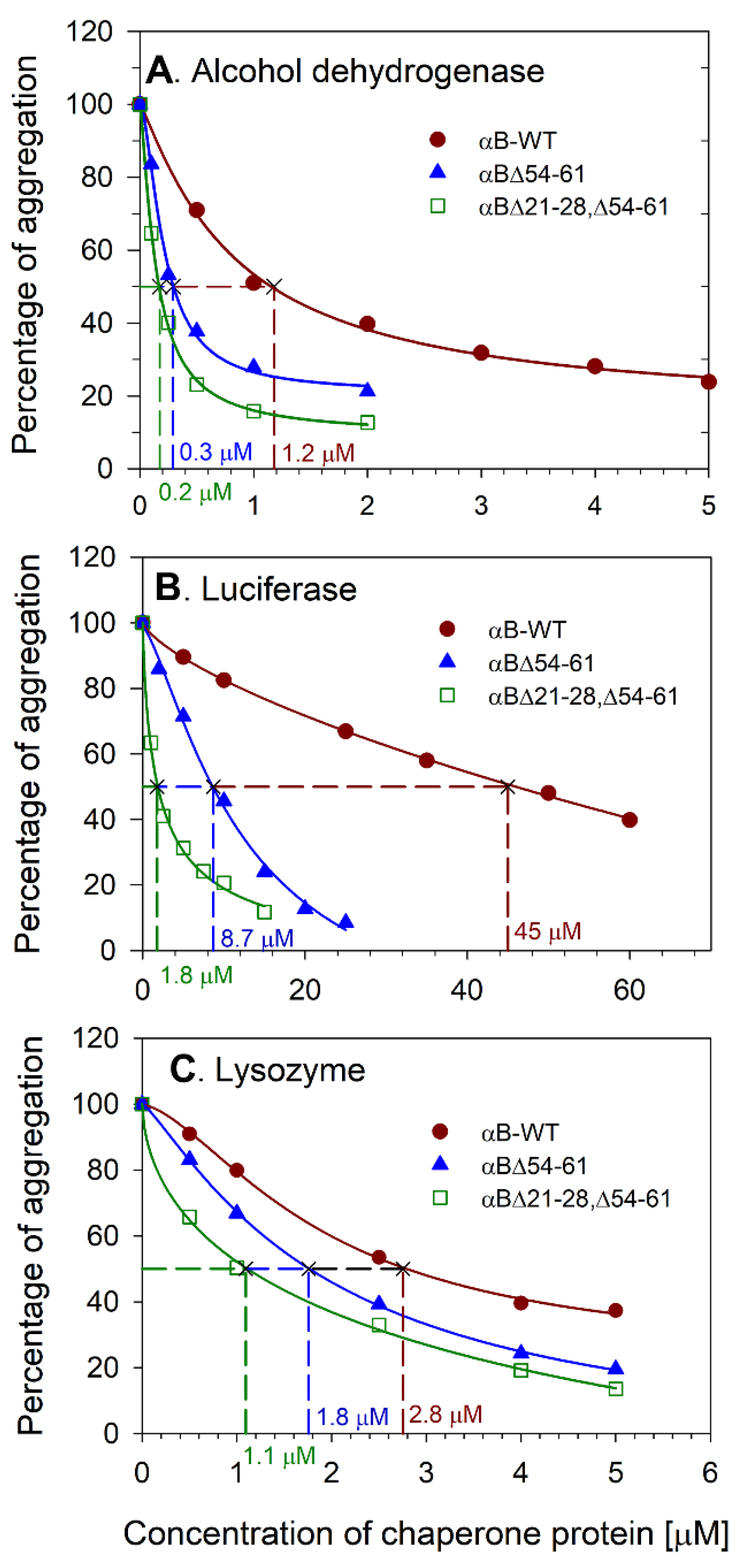
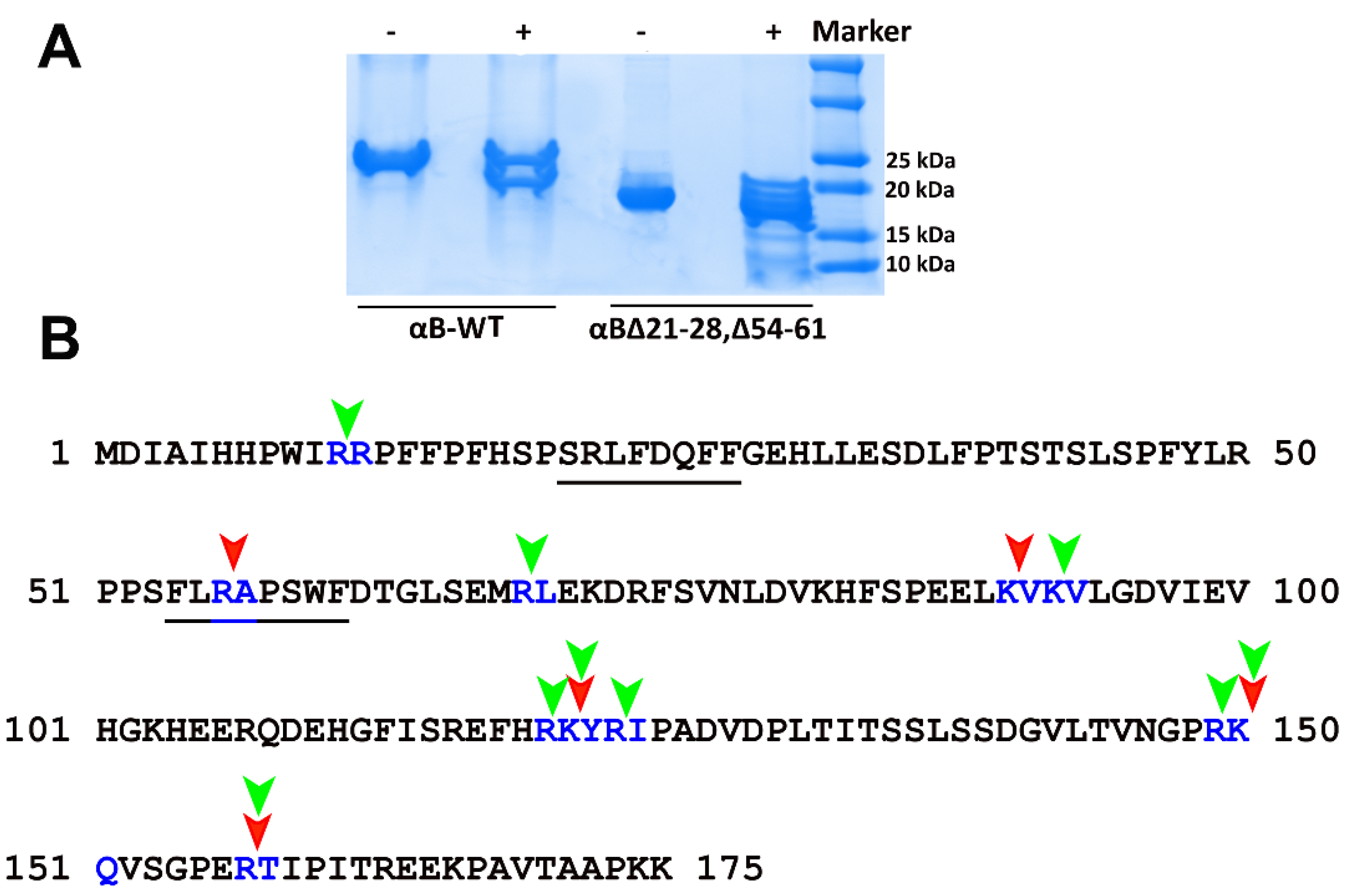
2.1. Deleting 21–28 and 54–61 Residues from αB-Crystallin Results in Increased Chaperone Activity
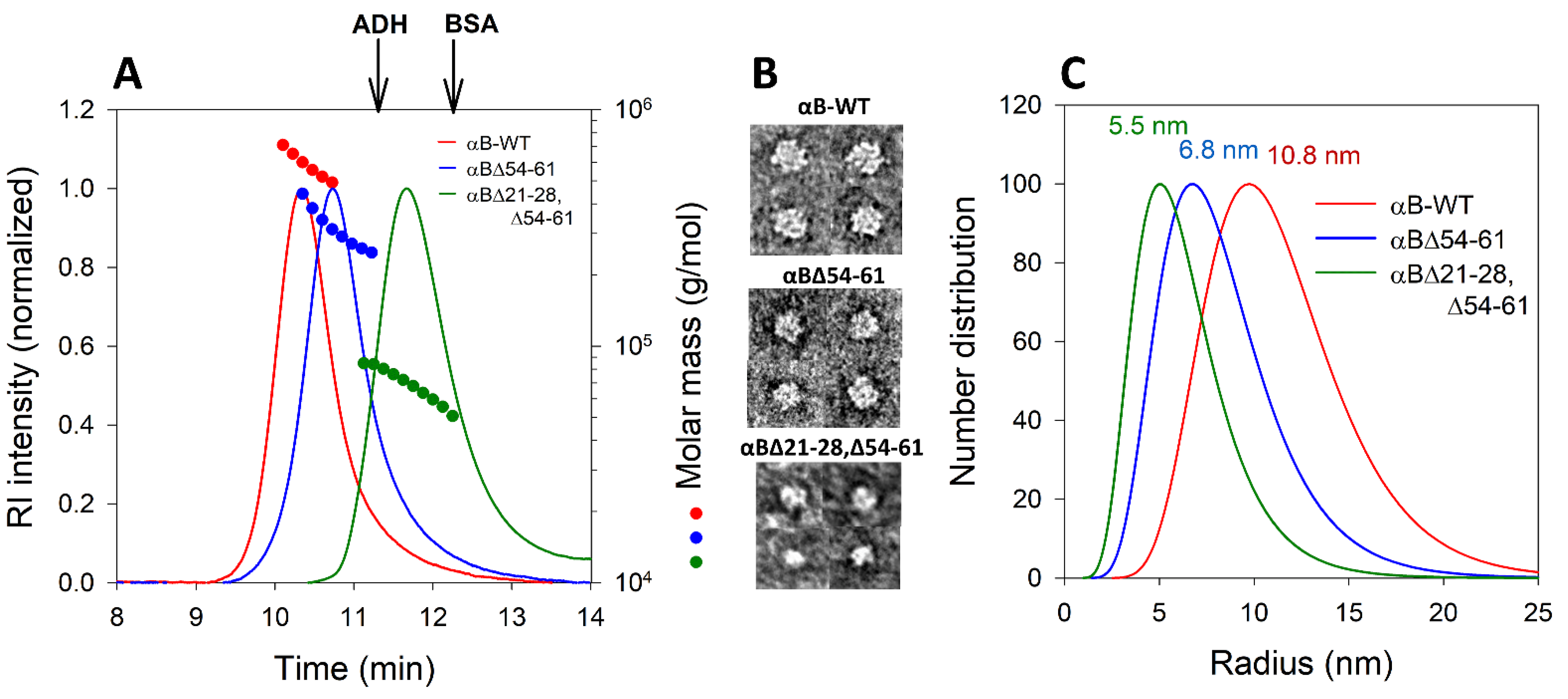
2.2. Deletion of 21–28 and 54–61 Residues in αB-Crystallin Leads to the Smaller Homo-Oligomers
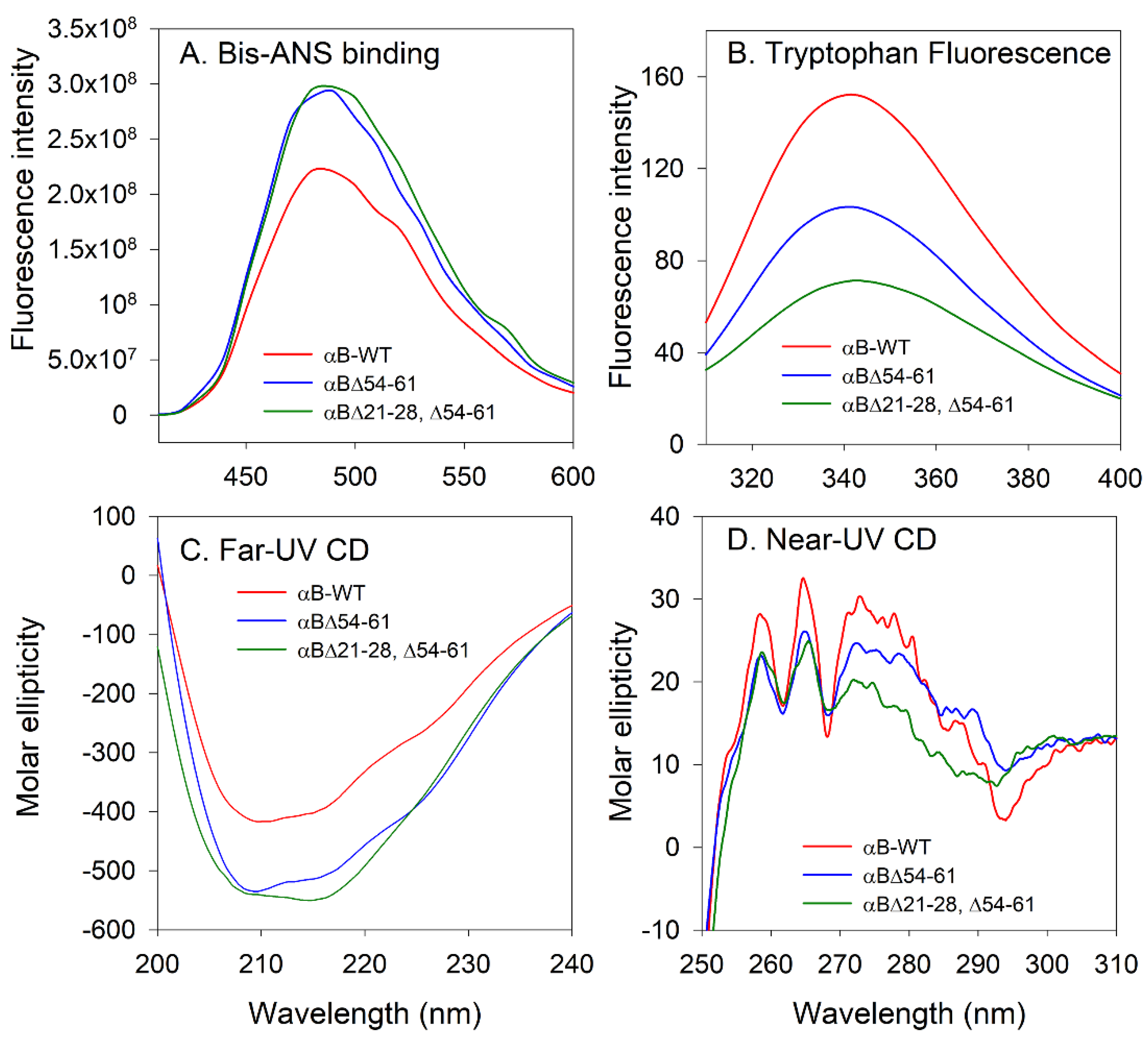
2.3. Structural Characterization of the αB∆21–28, ∆54–61 Showed Significant Differences with αB-WT and αB∆54–61
2.4. αB∆21–28, ∆54–61 Is More Susceptible to Cleavage by Trypsin than the aB-WT
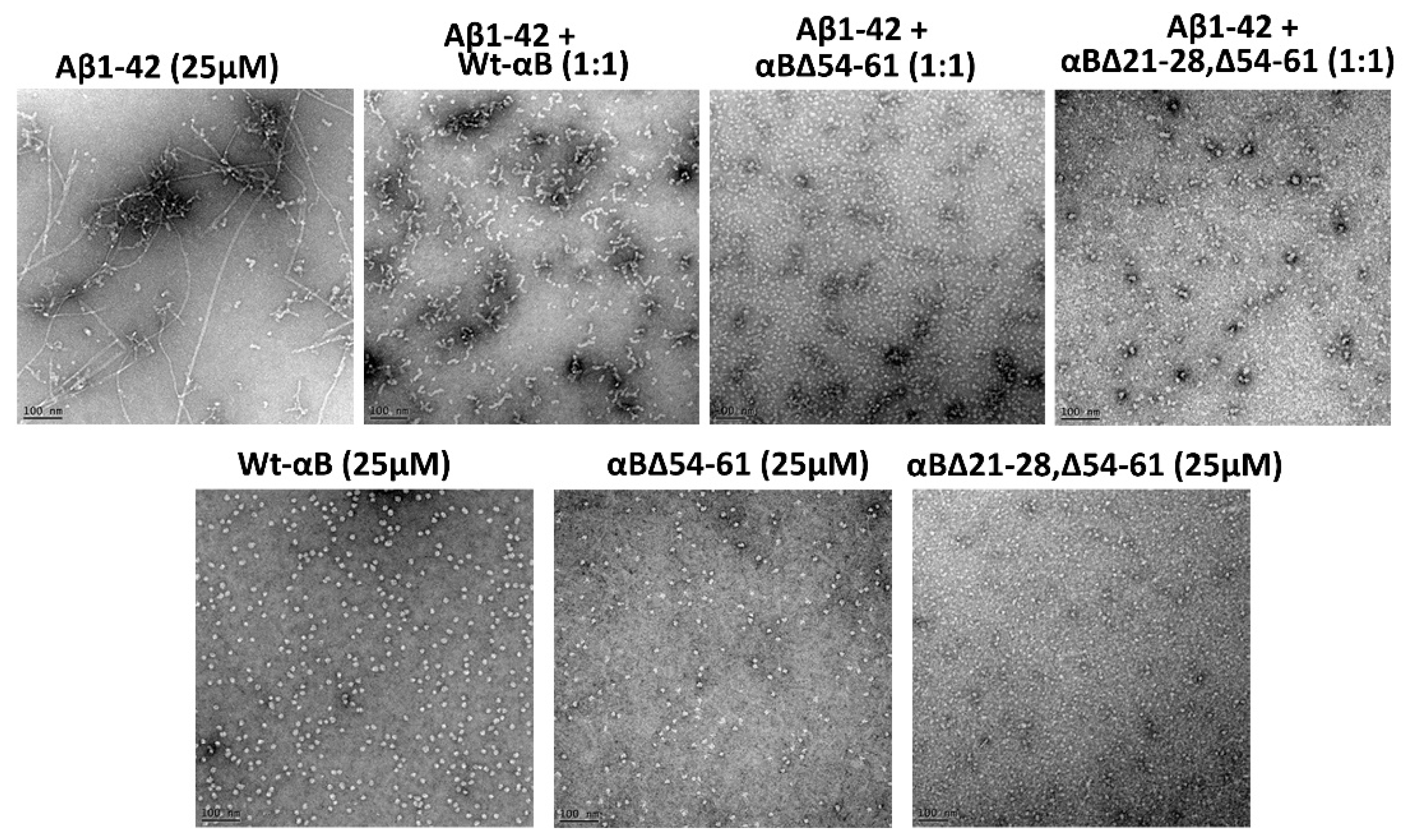
2.5. αB∆21–28, ∆54–61 Shows Increased Anti-Amyloidogenic Potential In Vitro
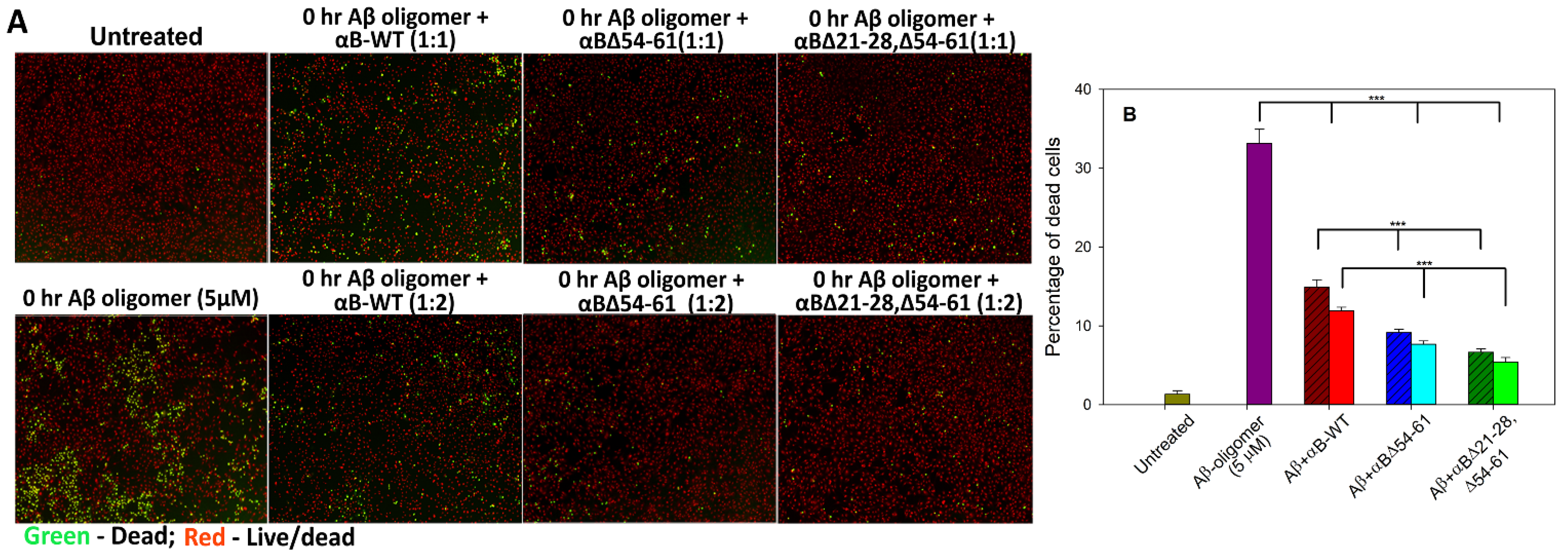
2.6. αBΔ21–28, Δ54–61 Crystallin Suppresses the Aβ1–42 Peptide-Induced Toxicity in ARPE-19 Cells
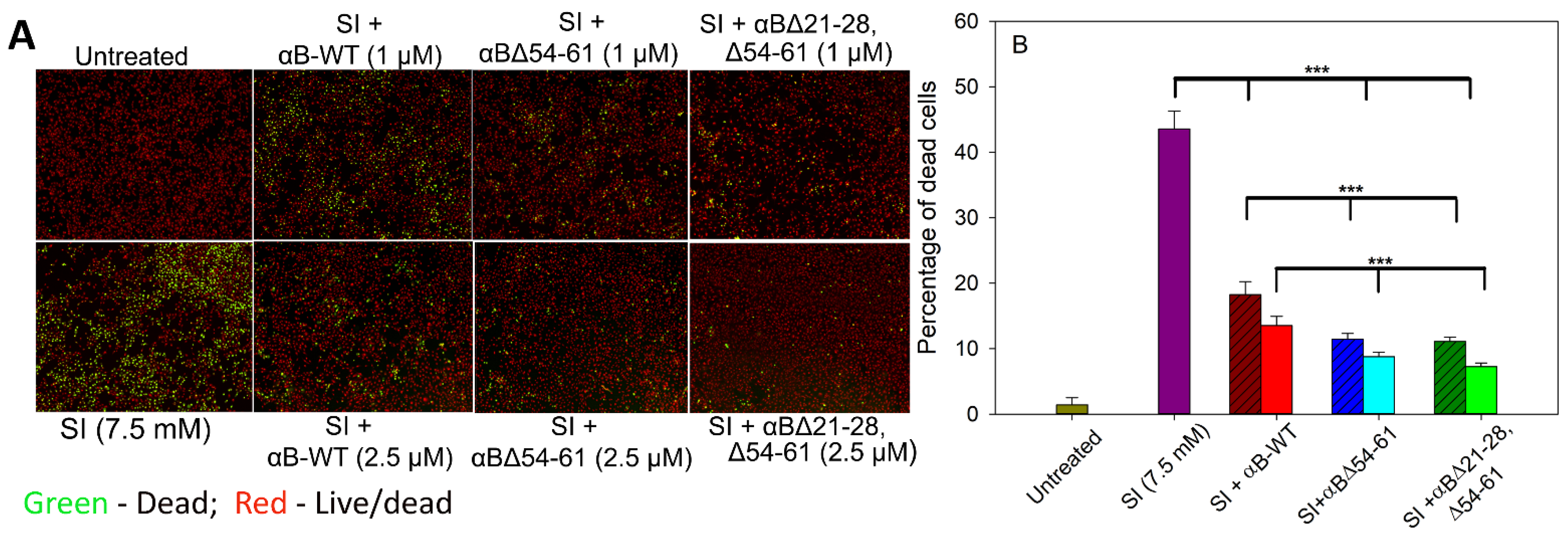
2.7. αBΔ21–28, Δ54–61 Crystallin Blocks the Cytotoxic Action of Sodium Iodate and Increases Cell Viability on ARPE-19 Cells
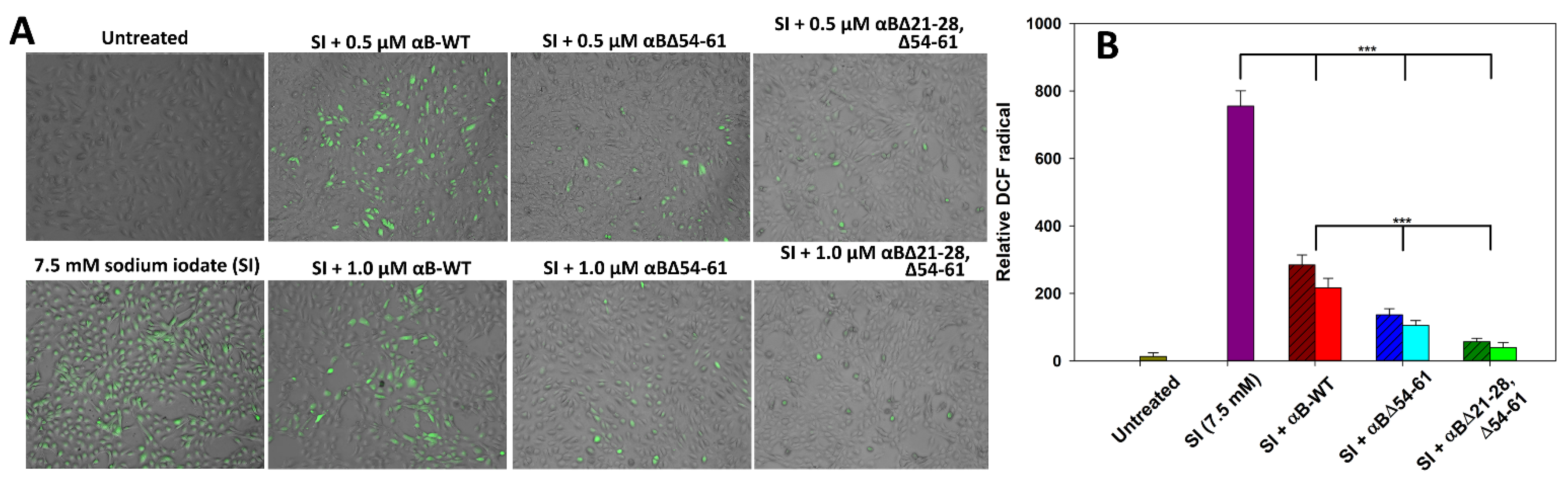
2.8. αBΔ21–28, Δ54–61 Attenuates Sodium Iodate-Induced Oxidative Stress in ARPE-19 Cells
3. Discussion
4. Materials and Methods
4.1. Construction of Plasmid DNA Expressing αB∆21–28, ∆54–61
4.2. Overexpression and Purification of Wild-Type and αB-Crystallin Mutants
4.3. Chaperone Assays
4.4. Structural Characterization of the αBΔ21–28, Δ54–61 Mutant
4.4.1. Multi-Angle Light Scattering Analysis of αBΔ21–28, Δ54–61 Crystallin
4.4.2. Transmission Electron Microscopy
4.4.3. Fluorescence Measurements
4.4.4. Far- and Near-UV CD Spectra
4.5. Limited Proteolysis of αB∆21–28, ∆54–61 Using Trypsin
4.6. Anti-Amyloidogenic Potential of αB∆21–28, ∆54–61 Crystallin
4.7. Suppression of β-Amyloid Cytotoxicity on ARPE-19 Cells by αB∆21–28, ∆54–61 Crystallin
4.8. Effect of αB∆21–28, ∆54–61 Crystallin on Sodium Iodate-Induced Oxidative Stress and Cytotoxicity on ARPE-19 Cells
4.9. Anti-Oxidant Action of αB∆21–28, ∆54–61 Crystallin on ARPE-19 Cells
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Horwitz, J. Alpha-crystallin can function as a molecular chaperone. Proc. Natl. Acad. Sci. USA 1992, 89, 10449–10453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raman, B.; Ban, T.; Sakai, M.; Pasta, S.Y.; Ramakrishna, T.; Naiki, H.; Goto, Y.; Rao, C.M. AlphaB-crystallin, a small heat-shock protein, prevents the amyloid fibril growth of an amyloid beta-peptide and beta2-microglobulin. Biochem. J. 2005, 392, 573–581. [Google Scholar] [CrossRef] [Green Version]
- Ecroyd, H.; Meehan, S.; Horwitz, J.; Aquilina, J.A.; Benesch, J.L.; Robinson, C.V.; Macphee, C.E.; Carver, J.A. Mimicking phosphorylation of alphaB-crystallin affects its chaperone activity. Biochem. J. 2007, 401, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Derham, B.K.; Harding, J.J. Effects of modifications of alpha-crystallin on its chaperone and other properties. Biochem. J. 2002, 364, 711–717. [Google Scholar] [CrossRef]
- Peschek, J.; Braun, N.; Franzmann, T.M.; Georgalis, Y.; Haslbeck, M.; Weinkauf, S.; Buchner, J. The eye lens chaperone alpha-crystallin forms defined globular assemblies. Proc. Natl. Acad. Sci. USA 2009, 106, 13272–13277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derham, B.K.; Harding, J.J. Alpha-crystallin as a molecular chaperone. Prog. Retin. Eye Res. 1999, 18, 463–509. [Google Scholar] [CrossRef]
- Santhoshkumar, P.; Murugesan, R.; Sharma, K.K. Deletion of (54)FLRAPSWF(61) residues decreases the oligomeric size and enhances the chaperone function of alphaB-crystallin. Biochemistry 2009, 48, 5066–5073. [Google Scholar] [CrossRef] [Green Version]
- Mahalingam, S.; Karmakar, S.; Santhoshkumar, P.; Sharma, K.K. Effect of Structural Changes Induced by Deletion of 54FLRAPSWF61 Sequence in αB-crystallin on Chaperone Function and Anti-Apoptotic Activity. Int. J. Mol. Sci. 2021, 22, 10771. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.B.; Sun, Y.; Smith, D.L.; Green, B. Identification of the posttranslational modifications of bovine lens alpha B-crystallins by mass spectrometry. Protein Sci. 1992, 1, 601–608. [Google Scholar] [CrossRef]
- Ahmad, M.F.; Raman, B.; Ramakrishna, T.; Rao Ch, M. Effect of phosphorylation on alpha B-crystallin: Differences in stability, subunit exchange and chaperone activity of homo and mixed oligomers of alpha B-crystallin and its phosphorylation-mimicking mutant. J. Mol. Biol. 2008, 375, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Ciano, M.; Allocca, S.; Ciardulli, M.C.; Della Volpe, L.; Bonatti, S.; D’Agostino, M. Differential phosphorylation-based regulation of αB-crystallin chaperone activity for multipass transmembrane proteins. Biochem. Biophys. Res. Commun. 2016, 479, 325–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mühlhofer, M.; Peters, C.; Kriehuber, T.; Kreuzeder, M.; Kazman, P.; Rodina, N.; Reif, B.; Haslbeck, M.; Weinkauf, S.; Buchner, J. Phosphorylation activates the yeast small heat shock protein Hsp26 by weakening domain contacts in the oligomer ensemble. Nat. Commun. 2021, 12, 6697. [Google Scholar] [CrossRef]
- Jehle, S.; Vollmar, B.S.; Bardiaux, B.; Dove, K.K.; Rajagopal, P.; Gonen, T.; Oschkinat, H.; Klevit, R.E. N-terminal domain of alphaB-crystallin provides a conformational switch for multimerization and structural heterogeneity. Proc. Natl. Acad. Sci. USA 2011, 108, 6409–6414. [Google Scholar] [CrossRef] [Green Version]
- Kundu, M.; Sen, P.C.; Das, K.P. Structure, stability, and chaperone function of αA-crystallin: Role of N-terminal region. Biopolymers 2007, 86, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Pasta, S.Y.; Raman, B.; Ramakrishna, T.; Rao, C.M. Role of the conserved SRLFDQFFG region of alpha-crystallin, a small heat shock protein. Effect on oligomeric size, subunit exchange, and chaperone-like activity. J. Biol. Chem. 2003, 278, 51159–51166. [Google Scholar] [CrossRef]
- Dasari, B.; Prasanthi, J.R.; Marwarha, G.; Singh, B.B.; Ghribi, O. The oxysterol 27-hydroxycholesterol increases β-amyloid and oxidative stress in retinal pigment epithelial cells. BMC Ophthalmol. 2010, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Butterfield, D.A.; Boyd-Kimball, D. Amyloid β-Peptide(1-42) Contributes to the Oxidative Stress and Neurodegeneration Found in Alzheimer Disease Brain. Brain Pathol. 2004, 14, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Kiuchi, K.; Yoshizawa, K.; Shikata, N.; Moriguchi, K.; Tsubura, A. Morphologic characteristics of retinal degeneration induced by sodium iodate in mice. Curr. Eye Res. 2002, 25, 373–379. [Google Scholar] [CrossRef]
- Yang, Y.; Ng, T.K.; Ye, C.; Yip, Y.W.Y.; Law, K.; Chan, S.-O.; Pang, C.P. Assessing Sodium Iodate–Induced Outer Retinal Changes in Rats Using Confocal Scanning Laser Ophthalmoscopy and Optical Coherence Tomography. Investig. Ophthalmol. Vis. Sci. 2014, 55, 1696–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibazawa, A.; Nagaoka, T.; Takahashi, A.; Omae, T.; Tani, T.; Sogawa, K.; Yokota, H.; Yoshida, A. Optical Coherence Tomography Angiography in Diabetic Retinopathy: A Prospective Pilot Study. Am. J. Ophthalmol. 2015, 160, 35–44.e1. [Google Scholar] [CrossRef] [Green Version]
- Diwanji, N.; Bergmann, A. An unexpected friend—ROS in apoptosis-induced compensatory proliferation: Implications for regeneration and cancer. Semin. Cell Dev. Biol. 2018, 80, 74–82. [Google Scholar] [CrossRef]
- Ghosh, J.G.; Clark, J.I. Insights into the domains required for dimerization and assembly of human alphaB crystallin. Protein Sci. 2005, 14, 684–695. [Google Scholar] [CrossRef] [Green Version]
- Shroff, N.P.; CheriaŽ Shaw, M.; Bera, S.; Abraham, E.C. Mutation of R116C results in highly oligomerized alpha A-crystallin with modified structure and defective chaperone-like function. Biochemistry 2000, 39, 1420–1426. [Google Scholar] [CrossRef]
- Phadte, A.S.; Mahalingam, S.; Santhoshkumar, P.; Sharma, K.K. Functional Rescue of Cataract-Causing alphaA-G98R-Crystallin by Targeted Compensatory Suppressor Mutations in Human alphaA-Crystallin. Biochemistry 2019, 58, 4148–4158. [Google Scholar] [CrossRef]
- Shatov, V.M.; Weeks, S.D.; Strelkov, S.V.; Gusev, N.B. The Role of the Arginine in the Conserved N-Terminal Domain RLFDQxFG Motif of Human Small Heat Shock Proteins HspB1, HspB4, HspB5, HspB6, and HspB8. Int. J. Mol. Sci. 2018, 19, 2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feil, I.K.; Malfois, M.; Hendle, J.; van Der Zandt, H.; Svergun, D.I. A novel quaternary structure of the dimeric alpha-crystallin domain with chaperone-like activity. J. Biol. Chem. 2001, 276, 12024–12029. [Google Scholar] [CrossRef] [Green Version]
- Ohto-Fujita, E.; Fujita, Y.; Atomi, Y. Analysis of the alphaB-crystallin domain responsible for inhibiting tubulin aggregation. Cell Stress Chaperones 2007, 12, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosinski-Collins, M.S.; Flaugh, S.L.; King, J. Probing folding and fluorescence quenching in human γD crystallin Greek key domains using triple tryptophan mutant proteins. Protein Sci. 2004, 13, 2223–2235. [Google Scholar] [CrossRef] [PubMed]
- Sreelakshmi, Y.; Sharma, K.K. The interaction between alphaA- and alphaB-crystallin is sequence-specific. Mol. Vis. 2006, 12, 581–587. [Google Scholar] [PubMed]
- Reiss, A.B.; Arain, H.A.; Stecker, M.M.; Siegart, N.M.; Kasselman, L.J. Amyloid toxicity in Alzheimer’s disease. Rev. Neurosci. 2018, 29, 613–627. [Google Scholar] [CrossRef]
- Bao, F.; Wicklund, L.; Lacor, P.N.; Klein, W.L.; Nordberg, A.; Marutle, A. Different β-amyloid oligomer assemblies in Alzheimer brains correlate with age of disease onset and impaired cholinergic activity. Neurobiol. Aging 2012, 33, 825.e1–825.e13. [Google Scholar] [CrossRef]
- Cleary, J.P.; Walsh, D.M.; Hofmeister, J.J.; Shankar, G.M.; Kuskowski, M.A.; Selkoe, D.J.; Ashe, K.H. Natural oligomers of the amyloid-β protein specifically disrupt cognitive function. Nat. Neurosci. 2005, 8, 79–84. [Google Scholar] [CrossRef]
- Manczak, M.; Anekonda, T.S.; Henson, E.; Park, B.S.; Quinn, J.; Reddy, P.H. Mitochondria are a direct site of A beta accumulation in Alzheimer’s disease neurons: Implications for free radical generation and oxidative damage in disease progression. Hum. Mol. Genet. 2006, 15, 1437–1449. [Google Scholar] [CrossRef] [PubMed]
- Dragicevic, N.; Mamcarz, M.; Zhu, Y.; Buzzeo, R.; Tan, J.; Arendash, G.W.; Bradshaw, P.C. Mitochondrial amyloid-beta levels are associated with the extent of mitochondrial dysfunction in different brain regions and the degree of cognitive impairment in Alzheimer’s transgenic mice. J. Alzheimers Dis. 2010, 20 (Suppl. S2), S535–S550. [Google Scholar] [CrossRef] [Green Version]
- Kruk, J.; Kubasik-Kladna, K.; Aboul-Enein, H.Y. The Role Oxidative Stress in the Pathogenesis of Eye Diseases: Current Status and a Dual Role of Physical Activity. Mini Rev. Med. Chem. 2015, 16, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Beatty, S.; Koh, H.; Phil, M.; Henson, D.; Boulton, M. The role of oxidative stress in the pathogenesis of age-related macular degeneration. Surv. Ophthalmol. 2000, 45, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Ho, M.-C.; Peng, Y.-J.; Chen, S.-J.; Chiou, S.-H. Senile cataracts and oxidative stress. J. Clin. Gerontol. Geriatr. 2010, 1, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zheng, Y.; Wang, C.; Liu, Y. Glutathione depletion induces ferroptosis, autophagy, and premature cell senescence in retinal pigment epithelial cells. Cell Death Dis. 2018, 9, 753. [Google Scholar] [CrossRef] [Green Version]
- Hanus, J.; Zhang, H.; Wang, Z.; Liu, Q.; Zhou, Q.; Wang, S. Induction of necrotic cell death by oxidative stress in retinal pigment epithelial cells. Cell Death Dis. 2013, 4, e965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu El-Asrar, A.M.; Alam, K.; Garcia-Ramirez, M.; Ahmad, A.; Siddiquei, M.M.; Mohammad, G.; Mousa, A.; De Hertogh, G.; Opdenakker, G.; Simó, R. Association of HMGB1 with oxidative stress markers and regulators in PDR. Mol. Vis. 2017, 23, 853–871. [Google Scholar]
- Marazita, M.C.; Dugour, A.; Marquioni-Ramella, M.D.; Figueroa, J.M.; Suburo, A.M. Oxidative stress-induced premature senescence dysregulates VEGF and CFH expression in retinal pigment epithelial cells: Implications for Age-related Macular Degeneration. Redox Biol. 2016, 7, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Monés, J.; Leiva, M.; Peña, T.; Martínez, G.; Biarnés, M.; Garcia, M.; Serrano, A.; Fernandez, E. A Swine Model of Selective Geographic Atrophy of Outer Retinal Layers Mimicking Atrophic AMD: A Phase I Escalating Dose of Subretinal Sodium Iodate. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3974–3983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obata, R.; Yanagi, Y.; Tamaki, Y.; Hozumi, K.; Mutoh, M.; Tanaka, Y. Retinal degeneration is delayed by tissue factor pathway inhibitor-2 in RCS rats and a sodium-iodate-induced model in rabbits. Eye 2005, 19, 464–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, H.; Krebs, M.P.; Kaushal, S.; Scott, E.W. Enhanced retinal pigment epithelium regeneration after injury in MRL/MpJ mice. Exp. Eye Res. 2011, 93, 862–872. [Google Scholar] [CrossRef] [Green Version]
- Stine, W.B.; Jungbauer, L.; Yu, C.; LaDu, M.J. Preparing synthetic Aβ in different aggregation states. Methods Mol. Biol. 2011, 670, 13–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahalingam, S.; Shankar, G.; Mooney, B.P.; Singh, K.; Santhoshkumar, P.; Sharma, K.K. Deletion of Specific Conserved Motifs from the N-Terminal Domain of αB-Crystallin Results in the Activation of Chaperone Functions. Int. J. Mol. Sci. 2022, 23, 1099. https://doi.org/10.3390/ijms23031099
Mahalingam S, Shankar G, Mooney BP, Singh K, Santhoshkumar P, Sharma KK. Deletion of Specific Conserved Motifs from the N-Terminal Domain of αB-Crystallin Results in the Activation of Chaperone Functions. International Journal of Molecular Sciences. 2022; 23(3):1099. https://doi.org/10.3390/ijms23031099
Chicago/Turabian StyleMahalingam, Sundararajan, Goutham Shankar, Brian P. Mooney, Kamal Singh, Puttur Santhoshkumar, and Krishna K. Sharma. 2022. "Deletion of Specific Conserved Motifs from the N-Terminal Domain of αB-Crystallin Results in the Activation of Chaperone Functions" International Journal of Molecular Sciences 23, no. 3: 1099. https://doi.org/10.3390/ijms23031099
APA StyleMahalingam, S., Shankar, G., Mooney, B. P., Singh, K., Santhoshkumar, P., & Sharma, K. K. (2022). Deletion of Specific Conserved Motifs from the N-Terminal Domain of αB-Crystallin Results in the Activation of Chaperone Functions. International Journal of Molecular Sciences, 23(3), 1099. https://doi.org/10.3390/ijms23031099