
Polycondensed Peptide Carriers Modified with Cyclic RGD Ligand for Targeted Suicide Gene Delivery to Uterine Fibroid Cells



Abstract
:1. Introduction
2. Results and Discussion
2.1. Carrier Design and Molecular Weight Characterization
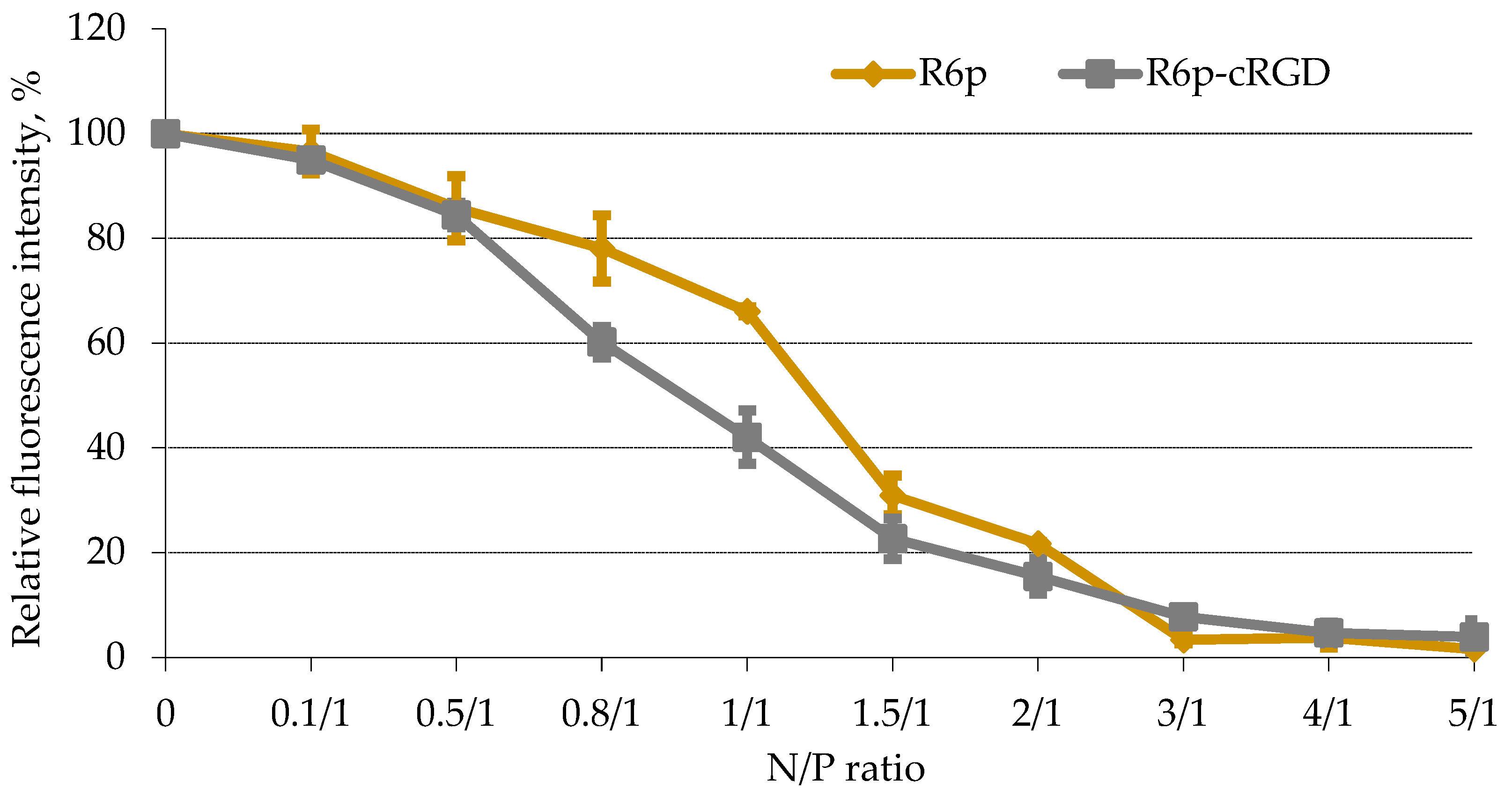
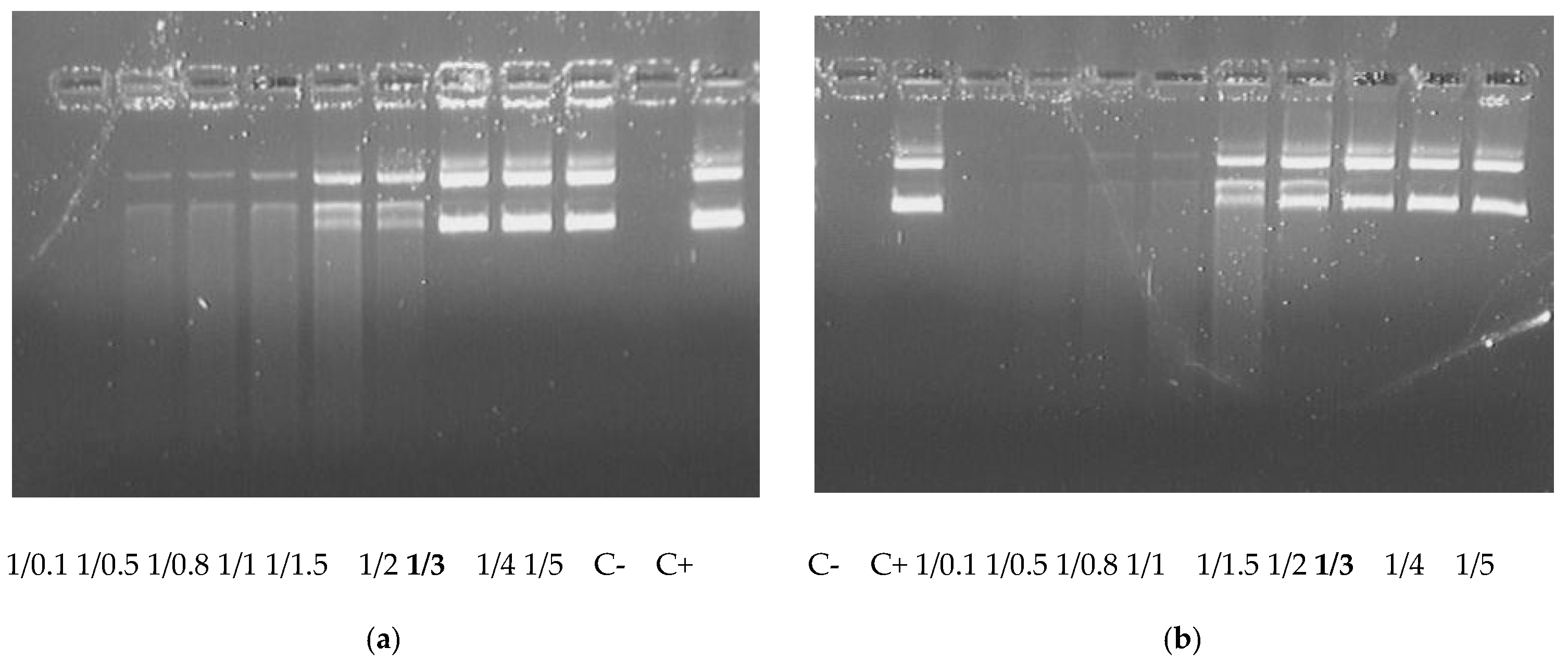
2.2. DNA-Binding and DNA Protection Properties of the Carrier
2.3. Size and ʐ-Potential of the Carrier/DNA Complexes
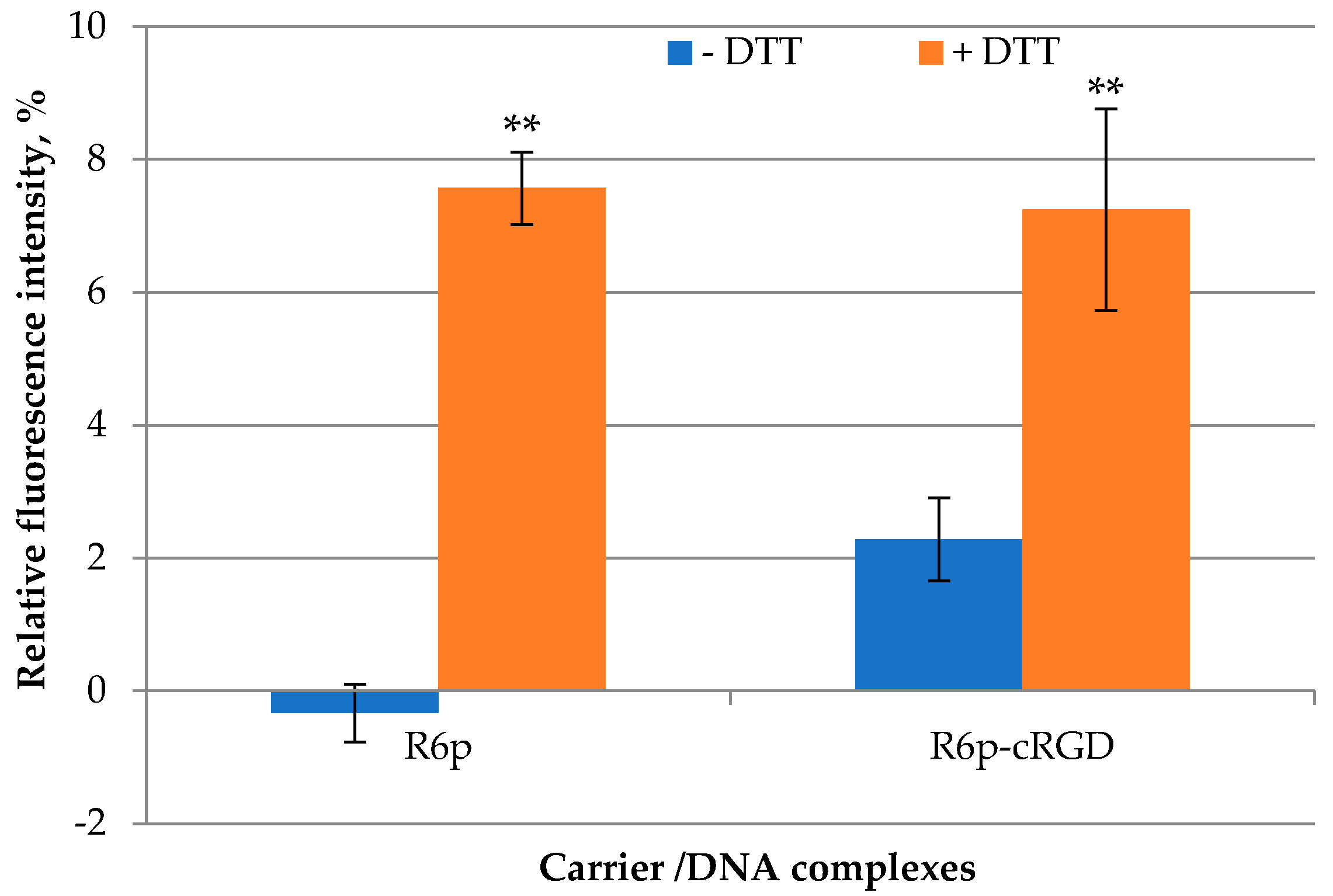
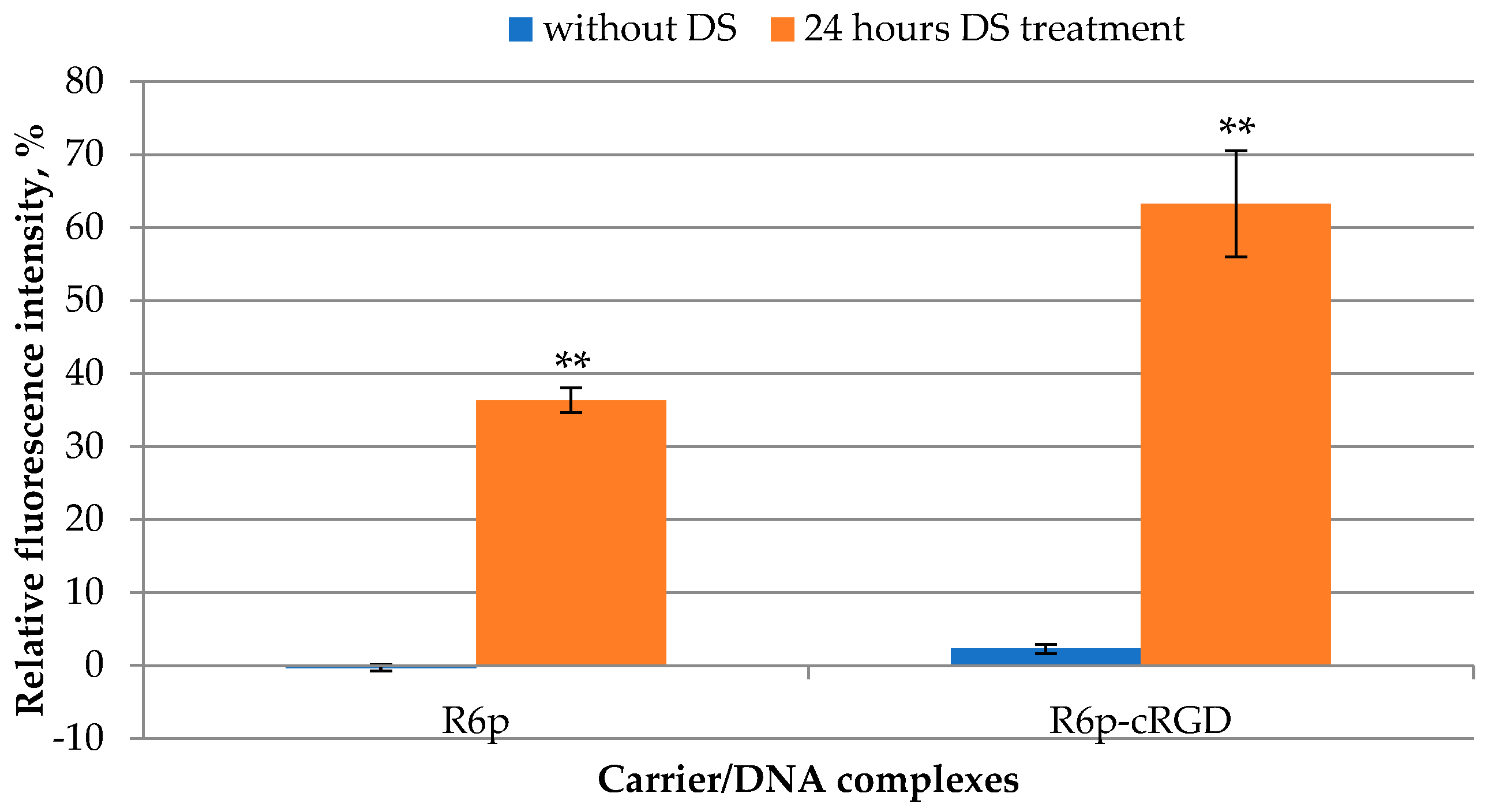
2.4. DTT and DS Treatment of the Complexes
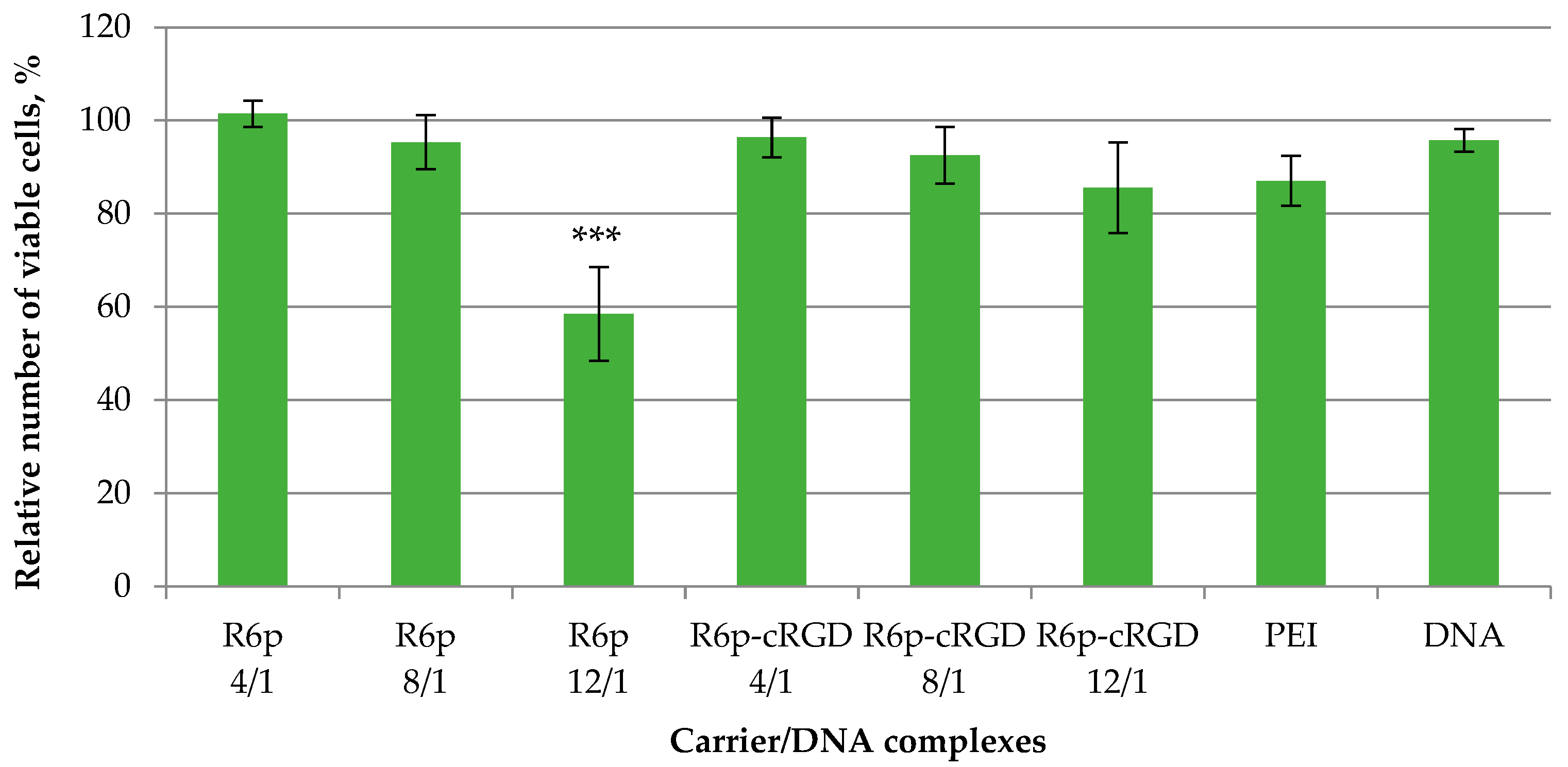
2.5. Cytotoxicity of Peptide/DNA Complexes
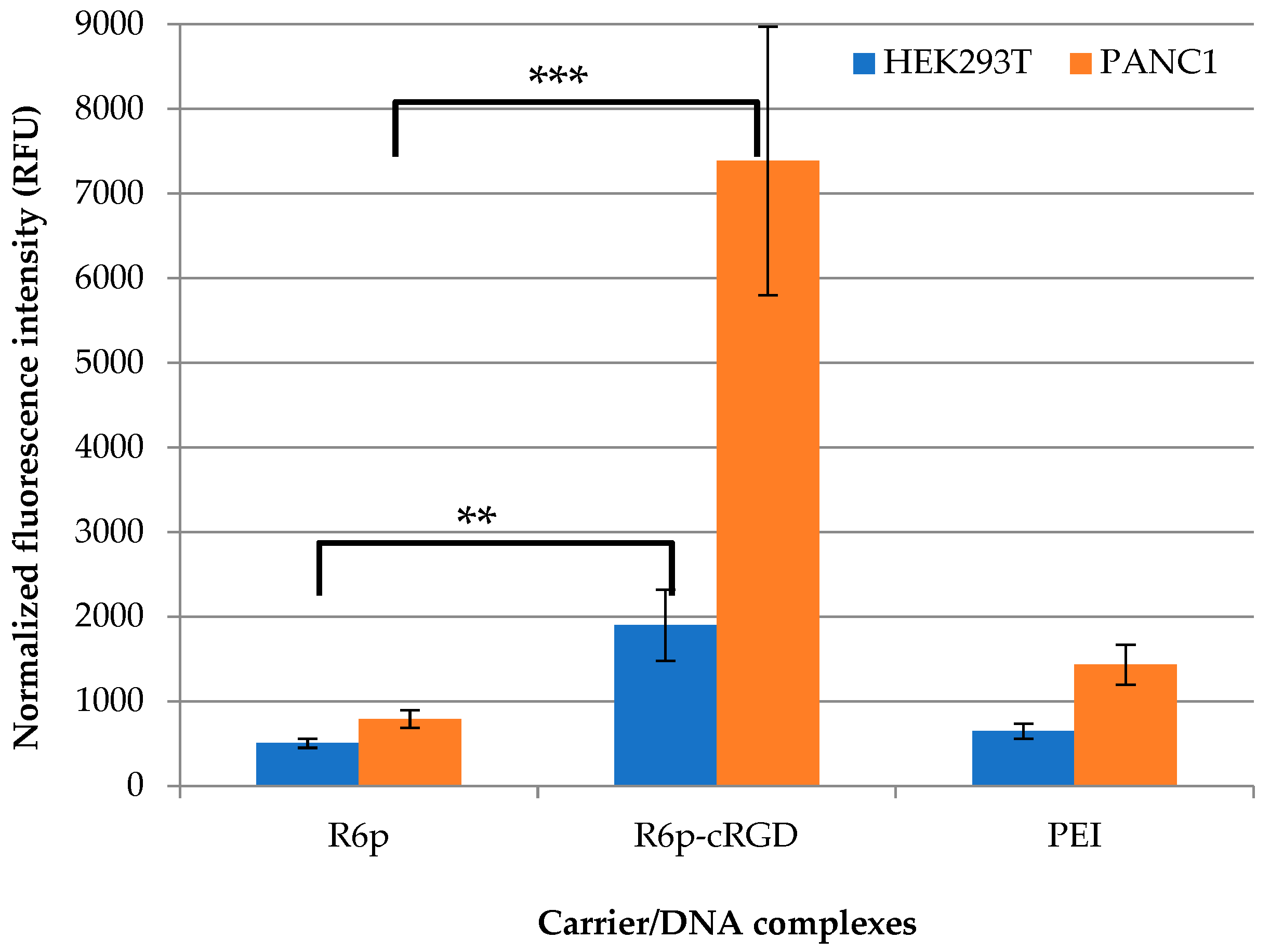
2.6. Cellular Uptake
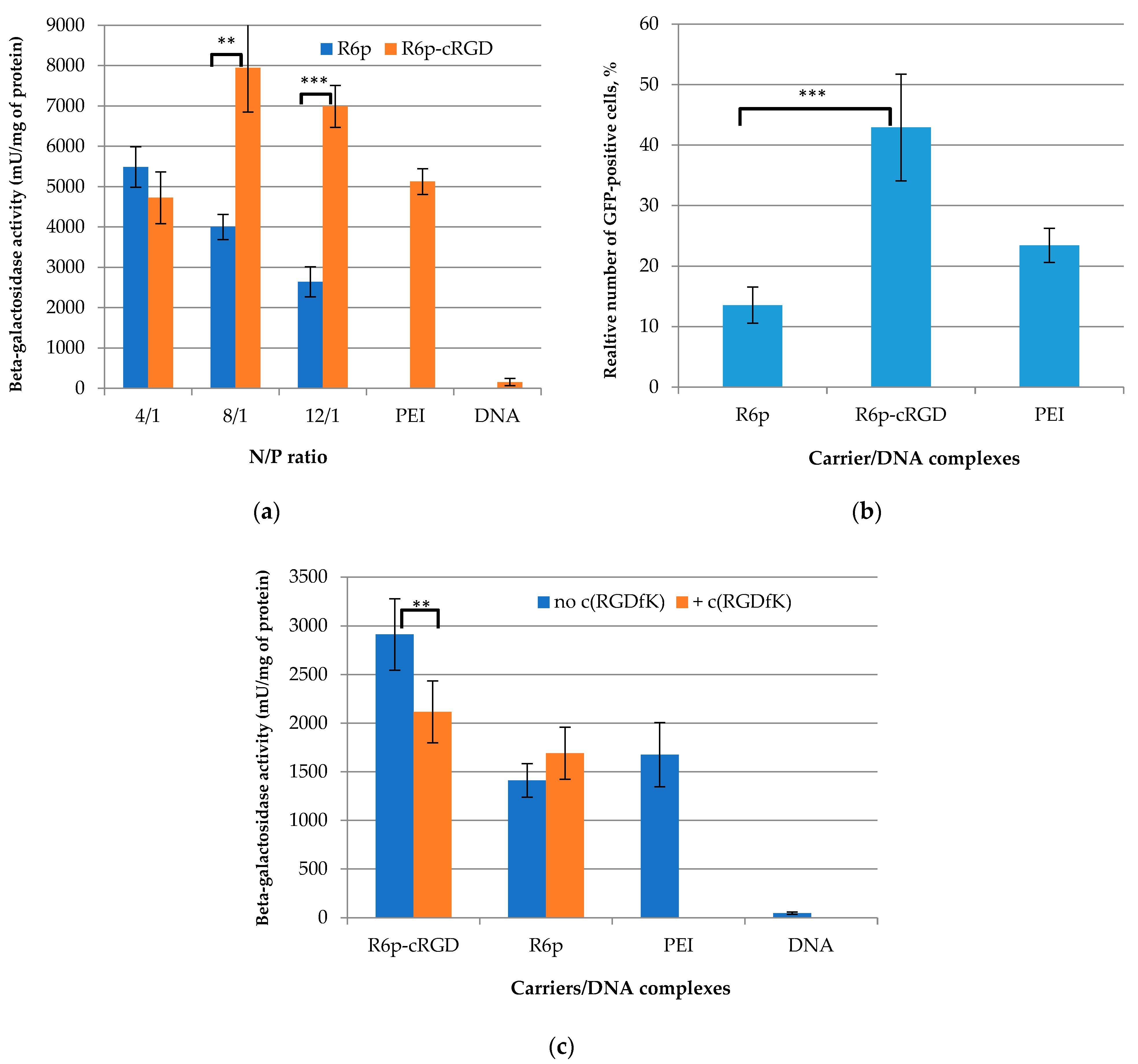
2.7. Transfection Studies
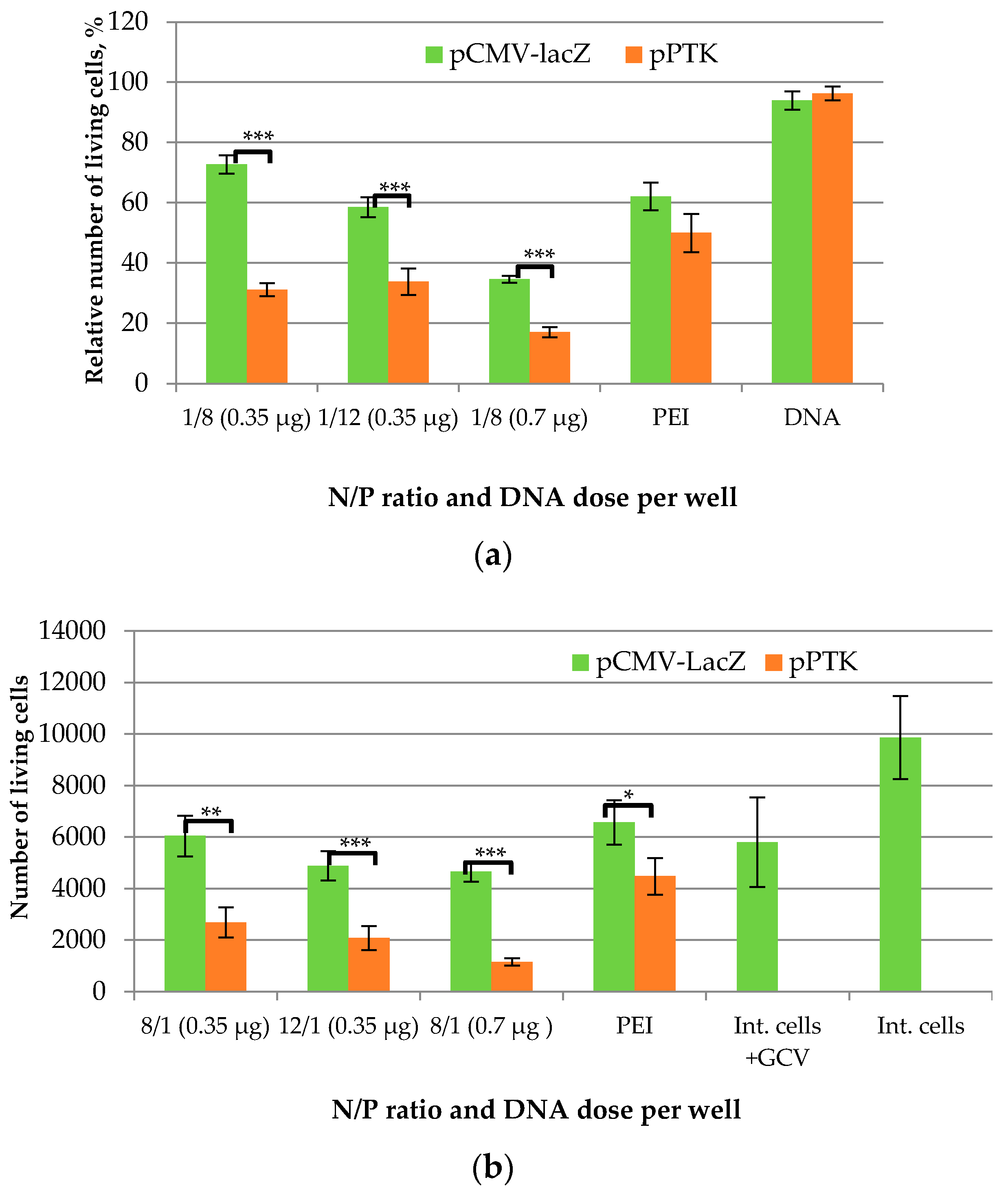
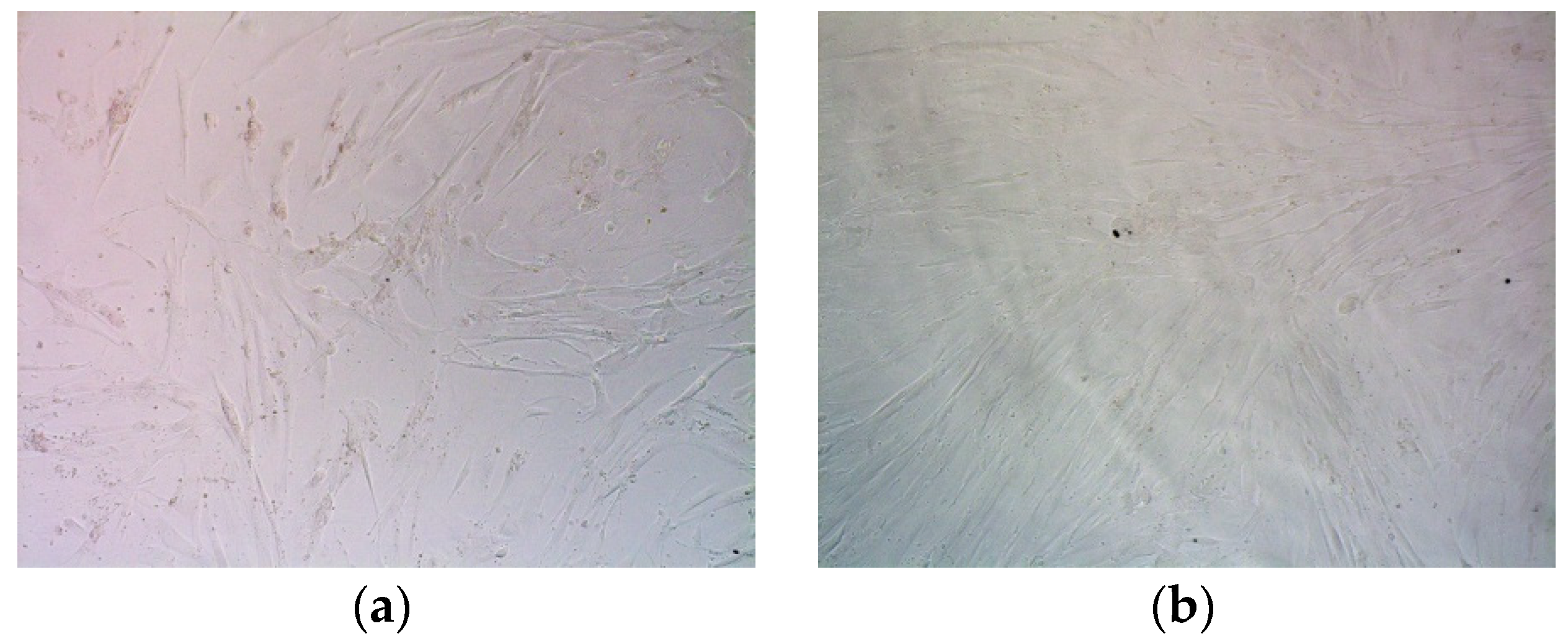
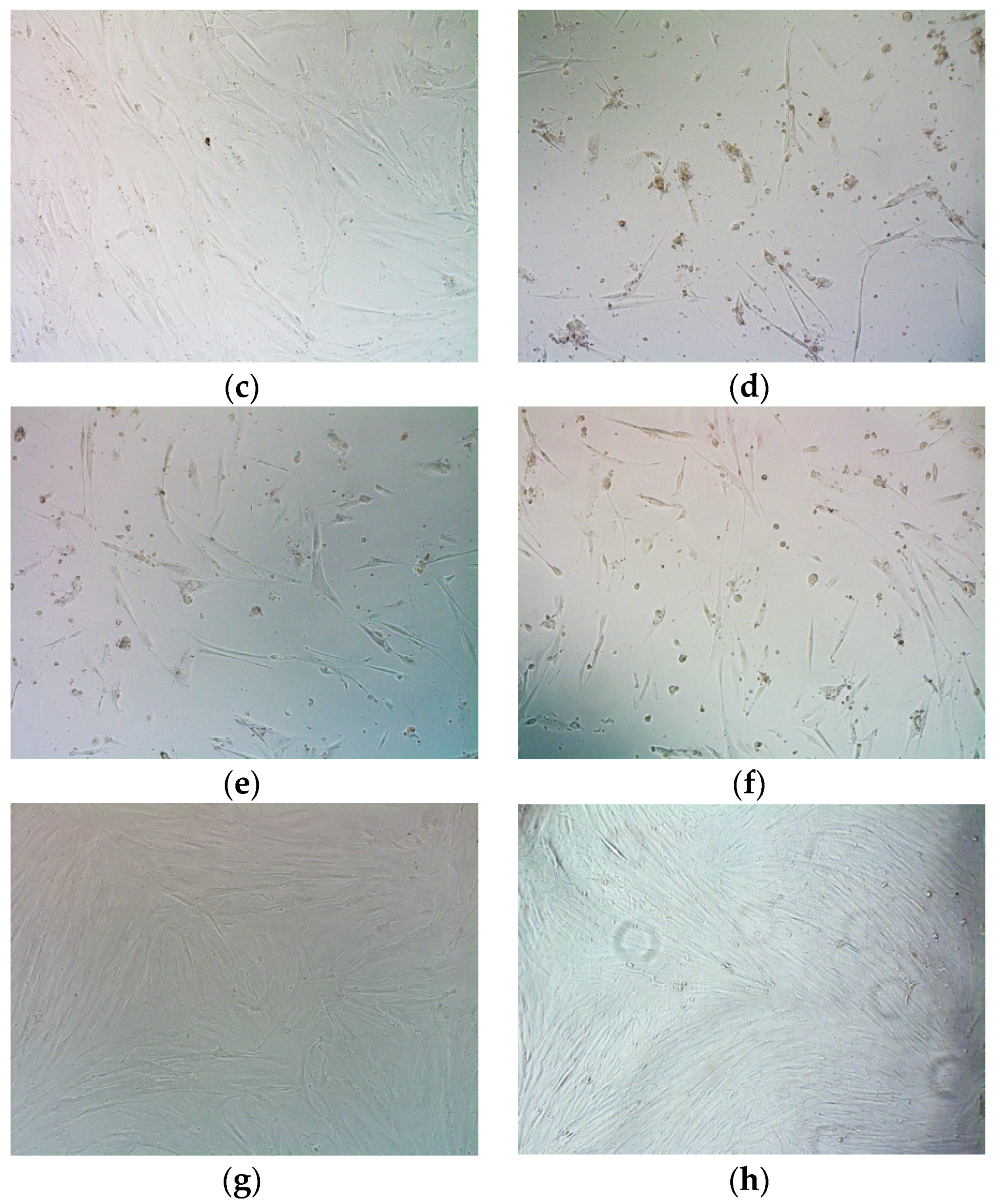
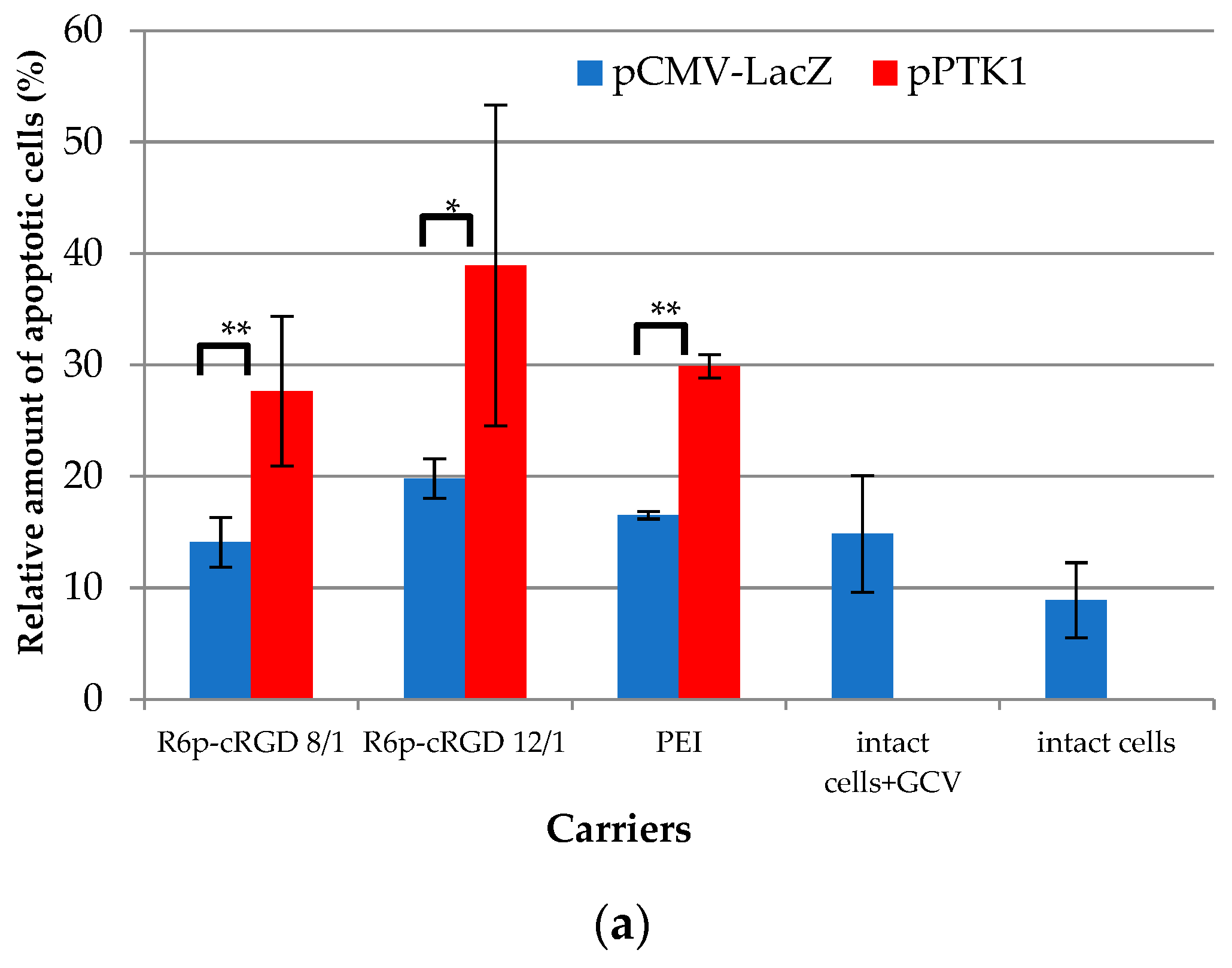
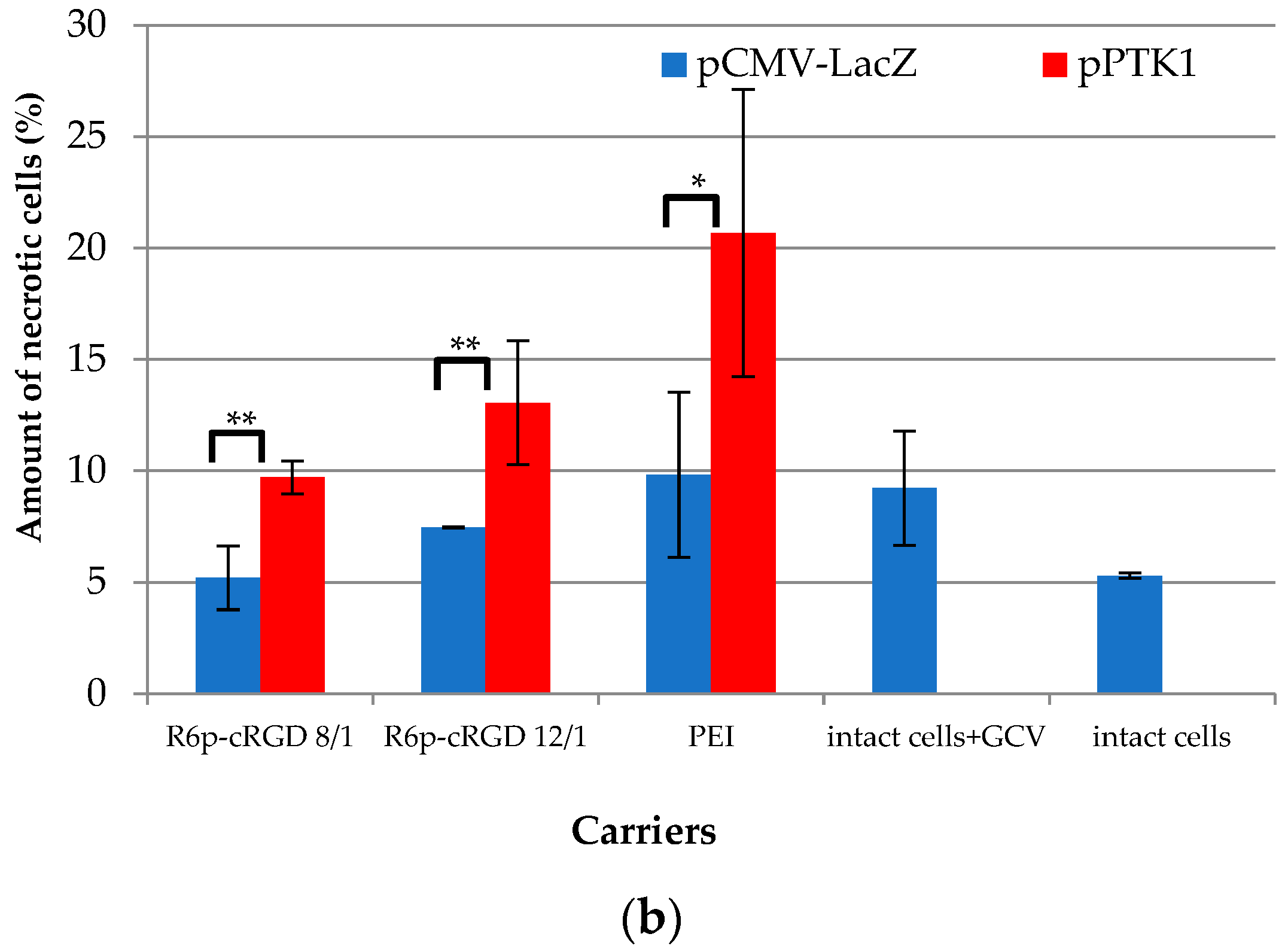
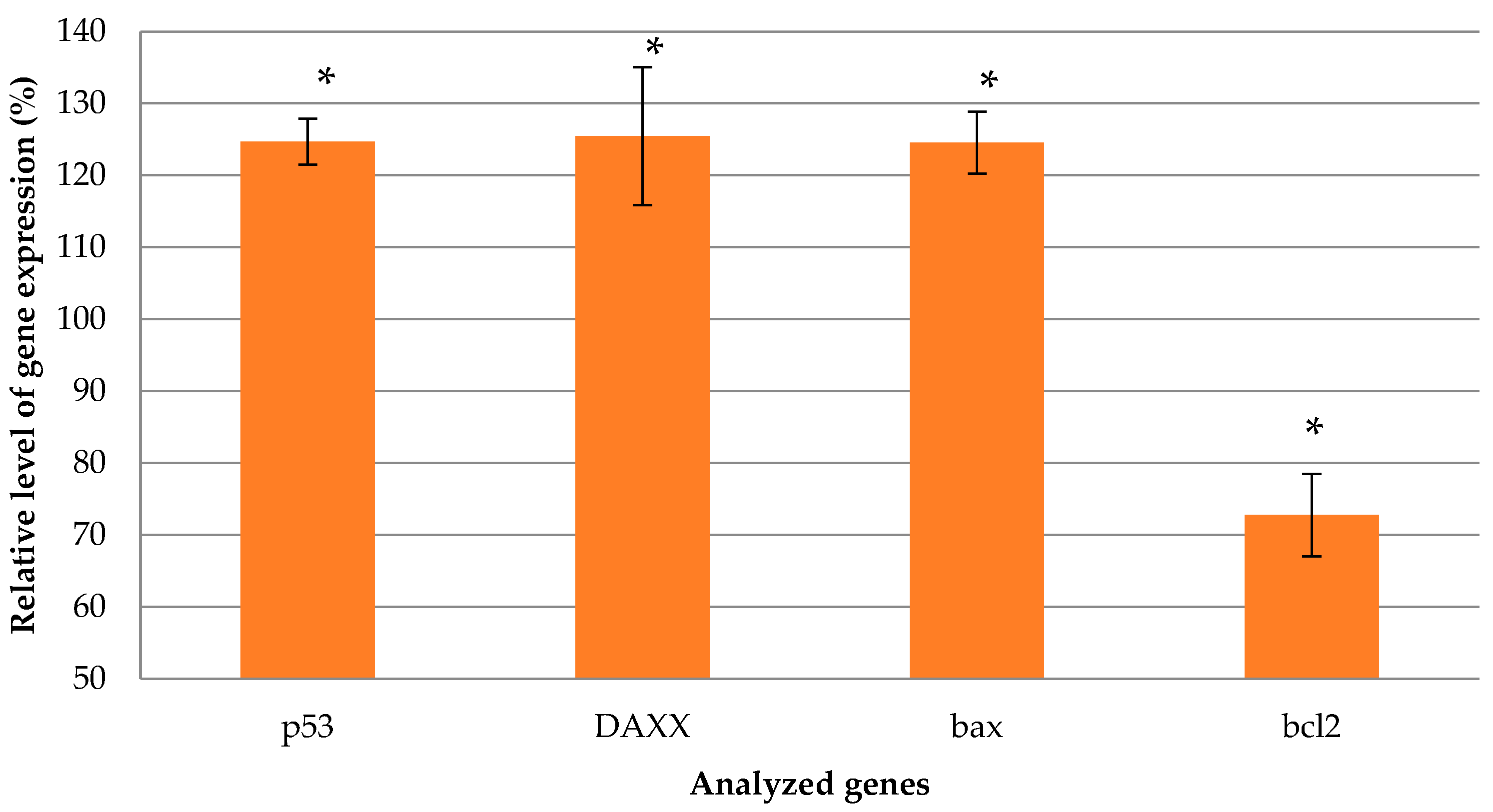
2.8. Assessment of Suicide Gene Therapy Effect in GSV-Treated Uterine Fibroid Cells after R6p-cRGD/pPTK1 Polyplex Transfection
3. Materials and Methods
3.1. Cell Lines
3.2. Peptide Synthesis and Characterization
3.3. Reporter Plasmids
3.4. Complex Preparation, DNA-Binding and Protection Assays, DS and DTT Treatment
3.5. Measurement of Size and ʐ-Potential of Peptide/DNA Complexes
3.6. Gene Transfer
3.7. Cellular Uptake of Peptide/DNA Complexes
3.8. Cytotoxicity Assay
3.9. Suicide Gene Therapy
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wirth, T.; Parker, N.; Ylä-Herttuala, S. History of gene therapy. Gene 2013, 525, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.J.; Barton-Smith, P. Long-term follow up of uterine artery embolisation-an effective alternative in the treatment of fibroids. BJOG Int. J. Obstet. Gynaecol. 2006, 113, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Stewart, E.A. Uterine fibroids. Lancet 2001, 357, 293–298. [Google Scholar] [CrossRef]
- Dubuisson, J. The current place of mini-invasive surgery in uterine leiomyoma management. J. Gynecol. Obstet. Hum. Reprod. 2019, 48, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Donnez, J.; Dolmans, M.-M. Hormone therapy for intramural myoma-related infertility from ulipristal acetate to GnRH antagonist: A review. Reprod. Biomed. Online 2020, 41, 431–442. [Google Scholar] [CrossRef]
- Cook, H.; Ezzati, M.; Segars, J.H.; McCarthy-Keith, D. The impact of uterine leiomyomas on reproductive outcomes. Minerva Ginecol. 2010, 62, 225–236. [Google Scholar]
- Shtykalova, S.V.; Egorova, A.A.; Maretina, M.A.; Freund, S.A.; Baranov, V.S.; Kiselev, A.V. Molecular Genetic Basis and Prospects of Gene Therapy of Uterine Leiomyoma. Russ. J. Genet. 2021, 57, 1002–1016. [Google Scholar] [CrossRef]
- Moolten, F.L.; Wells, J.M. Curability of Tumors Bearing Herpes Thymidine Kinase Genes Transfered by Retroviral Vectors. JNCI J. Natl. Cancer Inst. 1990, 82, 297–300. [Google Scholar] [CrossRef]
- Andersen, J.; Grine, E.; Eng, C.L.Y.; Zhao, K.; Barbieri, R.L.; Chumas, J.C.; Brink, P.R. Expression of connexin-43 in human myometrium and leiomyoma. Am. J. Obstet. Gynecol. 1993, 169, 1266–1276. [Google Scholar] [CrossRef]
- Salama, S.A.; Kamel, M.; Christman, G.; Wang, H.Q.; Fouad, H.M.; Al-Hendy, A. Gene therapy of uterine leiomyoma: Adenovirus-mediated herpes simplex virus thymidine kinase/ganciclovir treatment inhibits growth of human and rat leiomyoma cells in vitro and in a nude mouse model. Gynecol. Obstet. Investig. 2007, 63, 61–70. [Google Scholar] [CrossRef]
- Bakhtiar, A.; Chowdhury, E.H. PH-responsive strontium nanoparticles for targeted gene therapy against mammary carcinoma cells. Asian J. Pharm. Sci. 2021, 16, 236–252. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.H.; Khatoon, N.; Curiel, D.T.; Hamada, F.M.; Arafa, H.M.; Al-Hendy, A. Toward gene therapy of uterine fibroids: Targeting modified adenovirus to human leiomyoma cells. Hum. Reprod. 2008, 23, 514–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, S.; Curiel, D.T.; Rajaratnam, V.; Thota, C.; Al-Hendy, A. Targeting adenoviral vectors for enhanced gene therapy of uterine leiomyomas. Hum. Reprod. 2013, 28, 2398–2406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelaziz, M.; Sherif, L.; Elkhiary, M.; Nair, S.; Shalaby, S.; Mohamed, S.; Eziba, N.; El-Lakany, M.; Curiel, D.; Ismail, N.; et al. Targeted Adenoviral Vector Demonstrates Enhanced Efficacy for in Vivo Gene Therapy of Uterine Leiomyoma. Reprod. Sci. 2016, 23, 464–474. [Google Scholar] [CrossRef] [Green Version]
- Shalaby, S.M.; Khater, M.K.; Perucho, A.M.; Mohamed, S.A.; Helwa, I.; Laknaur, A.; Lebedyeva, I.; Liu, Y.; Diamond, M.P.; Al-Hendy, A.A. Magnetic nanoparticles as a new approach to improve the efficacy of gene therapy against differentiated human uterine fibroid cells and tumor-initiating stem cells. Fertil. Steril. 2016, 105, 1638–1648.e8. [Google Scholar] [CrossRef] [Green Version]
- Lukashev, A.N.; Zamyatnin, A.A. Viral vectors for gene therapy: Current state and clinical perspectives. Biochemistry 2016, 81, 700–708. [Google Scholar] [CrossRef]
- Niu, H. Nonviral Vector-Mediated Thymidine Kinase Gene Transfer and Ganciclovir Treatment in Leiomyoma Cells. Obstet. Gynecol. 1998, 91, 735–740. [Google Scholar] [CrossRef]
- Kato, T.; Yamashita, H.; Misawa, T.; Nishida, K.; Kurihara, M.; Tanaka, M.; Demizu, Y.; Oba, M. Plasmid DNA delivery by arginine-rich cell-penetrating peptides containing unnatural amino acids. Bioorg. Med. Chem. 2016, 24, 2681–2687. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, J.; Xu, D. Cell-penetrating peptides as noninvasive transmembrane vectors for the development of novel multifunctional drug-delivery systems. J. Control. Release 2016, 229, 130–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, E.; Dragojevic, S.; Price, A.; Raucher, D. Thermally Targeted p50 Peptide Inhibits Proliferation and Induces Apoptosis of Breast Cancer Cell Lines. Macromol. Biosci. 2020, 20, 2000170. [Google Scholar] [CrossRef]
- Zhou, S.; Watanabe, K.; Koide, S.; Kitamatsu, M.; Ohtsuki, T. Minimization of apoptosis-inducing CPP-Bim peptide. Bioorg. Med. Chem. Lett. 2021, 36, 127811. [Google Scholar] [CrossRef] [PubMed]
- Kosuge, M.; Takeuchi, T.; Nakase, I.; Jones, A.T.; Futaki, S. Cellular Internalization and Distribution of Arginine-Rich Peptides as a Function of Extracellular Peptide Concentration, Serum, and Plasma Membrane Associated Proteoglycans. Bioconjug. Chem. 2008, 19, 656–664. [Google Scholar] [CrossRef] [PubMed]
- van Asbeck, A.H.; Beyerle, A.; McNeill, H.; Bovee-Geurts, P.H.M.; Lindberg, S.; Verdurmen, W.P.R.; Hällbrink, M.; Langel, Ü.; Heidenreich, O.; Brock, R. Molecular Parameters of siRNA–Cell Penetrating Peptide Nanocomplexes for Efficient Cellular Delivery. ACS Nano 2013, 7, 3797–3807. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Guo, Z.; Du, Z.; Fang, R.; Wu, H.; Zeng, X.; Wang, C.; Feng, M.; Pan, S. Serum tolerance and endosomal escape capacity of histidine-modified pDNA-loaded complexes based on polyamidoamine dendrimer derivatives. Biomaterials 2012, 33, 8111–8121. [Google Scholar] [CrossRef]
- Chou, S.-T.; Hom, K.; Zhang, D.; Leng, Q.; Tricoli, L.J.; Hustedt, J.M.; Lee, A.; Shapiro, M.J.; Seog, J.; Kahn, J.D.; et al. Enhanced silencing and stabilization of siRNA polyplexes by histidine-mediated hydrogen bonds. Biomaterials 2014, 35, 846–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, Z.; Wang, X.; Tian, J.; Gao, Y.; Zhang, L.; Yao, C.; Wu, X.; Zhang, W.; Zhu, Q.; Gao, S. Biodegradable Stearylated Peptide with Internal Disulfide Bonds for Efficient Delivery of siRNA In Vitro and In Vivo. Biomacromolecules 2015, 16, 1119–1130. [Google Scholar] [CrossRef]
- McKenzie, D.L.; Kwok, K.Y.; Rice, K.G. A potent new class of reductively activated peptide gene delivery agents. J. Biol. Chem. 2000, 275, 9970–9977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oupický, D.; Parker, A.L.; Seymour, L.W. Laterally stabilized complexes of DNA with linear reducible polycations: Strategy for triggered intracellular activation of DNA delivery vectors. J. Am. Chem. Soc. 2002, 124, 8–9. [Google Scholar] [CrossRef]
- Soundara Manickam, D.; Bisht, H.S.; Wan, L.; Mao, G.; Oupicky, D. Influence of TAT-peptide polymerization on properties and transfection activity of TAT/DNA polyplexes. J. Control. Release 2005, 102, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, A.; Egorova, A.; Laukkanen, A.; Baranov, V.; Urtti, A. Characterization of reducible peptide oligomers as carriers for gene delivery. Int. J. Pharm. 2013, 441, 736–747. [Google Scholar] [CrossRef]
- Egorova, A.A.; Shtykalova, S.V.; Maretina, M.A.; Selyutin, A.V.; Shved, N.Y.; Krylova, N.V.; Ilina, A.V.; Pyankov, I.A.; Freund, S.A.; Selkov, S.A.; et al. Cys-Flanked Cationic Peptides For Cell Delivery of the Herpes Simplex Virus Thymidine Kinase Gene for Suicide Gene Therapy of Uterine Leiomyoma. Mol. Biol. 2020, 54, 436–448. [Google Scholar] [CrossRef]
- Liu, J.; Cheng, X.; Tian, X.; Guan, D.; Ao, J.; Wu, Z.; Huang, W.; Le, Z. Design and synthesis of novel dual-cyclic RGD peptides for αvβ3 integrin targeting. Bioorg. Med. Chem. Lett. 2019, 29, 896–900. [Google Scholar] [CrossRef]
- Malik, M.; Segars, J.; Catherino, W.H. Integrin β1 regulates leiomyoma cytoskeletal integrity and growth. Matrix Biol. 2012, 31, 389–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egorova, A.; Shtykalova, S.; Selutin, A.; Shved, N.; Maretina, M.; Selkov, S.; Baranov, V.; Kiselev, A. Development of irgd-modified peptide carriers for suicide gene therapy of uterine leiomyoma. Pharmaceutics 2021, 13, 202. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, T.; Endo, T.; Arima, N.; Ibaraki, H.; Takashima, Y.; Seta, Y. Systemic delivery of small interfering RNA targeting nuclear factor κB in mice with collagen-induced arthritis using arginine-histidine-cysteine based oligopeptide-modified polymer nanomicelles. Int. J. Pharm. 2016, 515, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Ibaraki, H.; Kanazawa, T.; Takashima, Y.; Okada, H.; Seta, Y. Transdermal anti-nuclear kappaB siRNA therapy for atopic dermatitis using a combination of two kinds of functional oligopeptide. Int. J. Pharm. 2018, 542, 213–220. [Google Scholar] [CrossRef]
- Egorova, A.; Selutin, A.; Maretina, M.; Selkov, S.; Baranov, V.; Kiselev, A. Characterization of iRGD-Ligand Modified Arginine-Histidine-Rich Peptides for Nucleic Acid Therapeutics Delivery to αvβ3 Integrin-Expressing Cancer Cells. Pharmaceuticals 2020, 13, 300. [Google Scholar] [CrossRef] [PubMed]
- Spears, R.J.; McMahon, C.; Chudasama, V. Cysteine protecting groups: Applications in peptide and protein science. Chem. Soc. Rev. 2021, 50, 11098–11155. [Google Scholar] [CrossRef]
- Lechardeur, D.; Sohn, K.-J.; Haardt, M.; Joshi, P.B.; Monck, M.; Graham, R.W.; Beatty, B.; Squire, J.; O’Brodovich, H.; Lukacs, G.L. Metabolic instability of plasmid DNA in the cytosol: A potential barrier to gene transfer. Gene Ther. 1999, 6, 482–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, S.; Tong, H.; Shi, Q.; Fernandes, J.C.; Jin, T.; Dai, K.; Zhang, X. Uptake mechanisms of non-viral gene delivery. J. Control. Release 2012, 158, 371–378. [Google Scholar] [CrossRef]
- Khalil, I.A.; Kogure, K.; Akita, H.; Harashima, H. Uptake pathways and subsequent intracellular trafficking in nonviral gene delivery. Pharmacol. Rev. 2006, 58, 32–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosse, S.; Aron, Y.; Thévenot, G.; François, D.; Monsigny, M.; Fajac, I. Potocytosis and cellular exit of complexes as cellular pathways for gene delivery by polycations. J. Gene Med. 2005, 7, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Litzinger, D.C.; Buiting, A.M.J.; van Rooijen, N.; Huang, L. Effect of liposome size on the circulation time and intraorgan distribution of amphipathic poly(ethylene glycol)-containing liposomes. Biochim. Biophys. Acta Biomembr. 1994, 1190, 99–107. [Google Scholar] [CrossRef]
- Ganbold, T.; Han, S.; Hasi, A.; Baigude, H. Receptor-mediated delivery of therapeutic RNA by peptide functionalized curdlan nanoparticles. Int. J. Biol. Macromol. 2019, 126, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Egorova, A.; Kiselev, A. Peptide modules for overcoming barriers of nucleic acids transport to cells. Curr. Top. Med. Chem. 2016, 16, 330–342. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.D.; Tandiono, F.; Wiradharma, N.; Khor, D.; Tan, C.G.; Khan, M.; Qian, Y.; Yang, Y.-Y. Cationic micelles self-assembled from cholesterol-conjugated oligopeptides as an efficient gene delivery vector. Biomaterials 2008, 29, 4838–4846. [Google Scholar] [CrossRef]
- Yue, Y.; Wu, C. Progress and perspectives in developing polymeric vectors for in vitro gene delivery. Biomater. Sci. 2013, 1, 152–170. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, J.-H.; Jeon, O.; Kwon, I.C.; Park, K. Engineered polymers for advanced drug delivery. Eur. J. Pharm. Biopharm. 2009, 71, 420–430. [Google Scholar] [CrossRef] [Green Version]
- Belting, M.; Petersson, P. Protective role for proteoglycans against cationic lipid cytotoxicity allowing optimal transfection efficiency in vitro. Biochem. J. 1999, 342, 281. [Google Scholar] [CrossRef]
- Ruponen, M.; Ylä-Herttuala, S.; Urtti, A. Interactions of polymeric and liposomal gene delivery systems with extracellular glycosaminoglycans: Physicochemical and transfection studies. Biochim. Biophys. Acta Biomembr. 1999, 1415, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Ruponen, M.; Honkakoski, P.; Tammi, M.; Urtti, A. Cell-surface glycosaminoglycans inhibit cation-mediated gene transfer. J. Gene Med. 2004, 6, 405–414. [Google Scholar] [CrossRef]
- Schaffer, D.V.; Fidelman, N.A.; Dan, N.; Lauffenburger, D.A. Vector unpacking as a potential barrier for receptor-mediated polyplex gene delivery. Biotechnol. Bioeng. 2000, 67, 598–606. [Google Scholar] [CrossRef]
- Saar, K.; Lindgren, M.; Hansen, M.; Eiríksdóttir, E.; Jiang, Y.; Rosenthal-Aizman, K.; Sassian, M.; Langel, Ü. Cell-penetrating peptides: A comparative membrane toxicity study. Anal. Biochem. 2005, 345, 55–65. [Google Scholar] [CrossRef] [PubMed]
- El-Andaloussi, S.; Järver, P.; Johansson, H.J.; Langel, Ü. Cargo-dependent cytotoxicity and delivery efficacy of cell-penetrating peptides: A comparative study. Biochem. J. 2007, 407, 285–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiradharma, N.; Khan, M.; Tong, Y.W.; Wang, S.; Yang, Y.-Y. Self-assembled Cationic Peptide Nanoparticles Capable of Inducing Efficient Gene Expression In Vitro. Adv. Funct. Mater. 2008, 18, 943–951. [Google Scholar] [CrossRef]
- Deng, K.; Tian, H.; Zhang, P.; Zhong, H.; Ren, X.; Wang, H. PH-temperature responsive poly(HPA-Co-AMHS) hydrogel as a potential drug-release carrier. J. Appl. Polym. Sci. 2009, 114, 176–184. [Google Scholar] [CrossRef]
- Kargaard, A.; Sluijter, J.P.G.; Klumperman, B. Polymeric siRNA gene delivery—Transfection efficiency versus cytotoxicity. J. Control. Release 2019, 316, 263–291. [Google Scholar] [CrossRef]
- Schiffelers, R.M. Cancer siRNA therapy by tumor selective delivery with ligand-targeted sterically stabilized nanoparticle. Nucleic Acids Res. 2004, 32, e149. [Google Scholar] [CrossRef]
- Al-Hendy, A.; Salama, S. Gene therapy and uterine leiomyoma: A review. Hum. Reprod. Update 2006, 12, 385–400. [Google Scholar] [CrossRef] [Green Version]
- Freeman, S.M.; Abboud, C.N.; Whartenby, K.A.; Packman, C.H.; Koeplin, D.S.; Moolten, F.L.; Abraham, G.N. The “Bystander Effect”: Tumor Regression When a Fraction of the Tumor Mass Is Genetically Modified. Cancer Res. 1993, 53, 5274–5283. [Google Scholar]
- Andrade-Rozental, A.F.; Rozental, R.; Hopperstad, M.G.; Wu, J.K.; Vrionis, F.D.; Spray, D.C. Gap junctions: The “kiss of death” and the “kiss of life”. Brain Res. Rev. 2000, 32, 308–315. [Google Scholar] [CrossRef]
- Krammer, P.H.; Kamiński, M.; Kießling, M.; Gülow, K. No Life Without Death. In Advances in Cancer Research; Academic Press Inc. Elsevier Science: San Diego, CA, USA, 2007; Volume 97, pp. 111–138. [Google Scholar]
- Cheng, K.; Kothapalli, S.-R.; Liu, H.; Koh, A.L.; Jokerst, J.V.; Jiang, H.; Yang, M.; Li, J.; Levi, J.; Wu, J.C.; et al. Construction and Validation of Nano Gold Tripods for Molecular Imaging of Living Subjects. J. Am. Chem. Soc. 2014, 136, 3560–3571. [Google Scholar] [CrossRef] [PubMed]
- Crowley, L.C.; Scott, A.P.; Marfell, B.J.; Boughaba, J.A.; Chojnowski, G.; Waterhouse, N.J. Measuring Cell Death by Propidium Iodide Uptake and Flow Cytometry. Cold Spring Harb. Protoc. 2016, 2016, pdb.prot087163. [Google Scholar] [CrossRef] [PubMed]
- Fischer, D. In Vivo Fate of Polymeric Gene Carriers; CRC Press: Boca Raton, FL, USA, 2004; ISBN 9780203492321. [Google Scholar]
- Haupt, S.; Berger, M.; Goldberg, Z.; Haupt, Y. Apoptosis—The p53 network. J. Cell Sci. 2003, 116, 4077–4085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jan, R.; Chaudhry, G.-E.-S. Understanding apoptosis and apoptotic pathways targeted cancer therapeutics. Adv. Pharm. Bull. 2019, 9, 205–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shved, N.; Egorova, A.; Osinovskaya, N.; Kiselev, A. Development of primary monolayer cell model m odel and organotypic model of uterine leiomyoma. Methods Protoc. 2021, 41, 1–12. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar] [CrossRef]
- Kiselev, A.V.; Il’ina, P.L.; Egorova, A.A.; Baranov, A.N.; Guryanov, I.A.; Bayanova, N.V.; Tarasenko, I.I.; Lesina, E.A.; Vlasov, G.P.; Baranov, V.S. Lysine dendrimers as vectors for delivery of genetic constructs to eukaryotic cells. Russ. J. Genet. 2007, 43, 593–600. [Google Scholar] [CrossRef]
- Egorova, A.; Bogacheva, M.; Shubina, A.; Baranov, V.; Kiselev, A. Development of a receptor-targeted gene delivery system using CXCR4 ligand-conjugated cross-linking peptides. J. Gene Med. 2014, 16, 336–351. [Google Scholar] [CrossRef]
- Egorova, A.; Petrosyan, M.; Maretina, M.; Balashova, N.; Polyanskih, L.; Baranov, V.; Kiselev, A. Anti-angiogenic treatment of endometriosis via anti-VEGFA siRNA delivery by means of peptide-based carrier in a rat subcutaneous model. Gene Ther. 2018, 25, 548–555. [Google Scholar] [CrossRef]
- Saed, G.M.; Jiang, Z.; Fletcher, N.M.; Diamond, M.P. Modulation of the BCL-2/BAX ratio by interferon-γ and hypoxia in human peritoneal and adhesion fibroblasts. Fertil. Steril. 2008, 90, 1925–1930. [Google Scholar] [CrossRef] [PubMed]
- Weglarz, L.; Molin, I.; Orchel, A.; Parfiniewicz, B.; Dzierzewicz, Z. Quantitative analysis of the level of p53 and p21(WAF1) mRNA in human colon cancer HT-29 cells treated with inositol hexaphosphate. Acta Biochim. Pol. 2006, 53, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Xu, G.; Zhang, J.; Tian, L.; Xue, J.; Chen, J.; Jia, W. Daxx interacts with HIV-1 integrase and inhibits lentiviral gene expression. Biochem. Biophys. Res. Commun. 2008, 373, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ma, Y.; Mu, L.; Chen, X.; Zheng, W. The expression of Bcl-2 in adenomyosis and its effect on proliferation, migration, and apoptosis of endometrial stromal cells. Pathol. Res. Pract. 2019, 215, 152477. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Composition | |
|---|---|---|
| R6 | CHRRRRRRHC | |
| Monomers | cRGD | C(Npys)RGDy |___________| |
| Carriers | R6p R6p-cRGD | (CHRRRRRRHC)n cRGD-(CHRRRRRRHC)n-cRGD |
| Carrier | N/P Ratio | Size (nm) ± SD | ʐ -Potential (mV) ± SD |
|---|---|---|---|
| R6p | 8/1 | 120.9 ± 0.17 | 35.6 ± 0.21 |
| R6p | 12/1 | 110.3 ± 9.12 | 35.5 ± 0.81 |
| R6p-cRGD | 8/1 | 104.5 ± 0.15 | 32.0 ± 1.17 |
| R6p-cRGD | 12/1 | 178.4 ± 24.32 | 31.5 ± 0.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Egorova, A.; Shtykalova, S.; Maretina, M.; Selutin, A.; Shved, N.; Deviatkin, D.; Selkov, S.; Baranov, V.; Kiselev, A. Polycondensed Peptide Carriers Modified with Cyclic RGD Ligand for Targeted Suicide Gene Delivery to Uterine Fibroid Cells. Int. J. Mol. Sci. 2022, 23, 1164. https://doi.org/10.3390/ijms23031164
Egorova A, Shtykalova S, Maretina M, Selutin A, Shved N, Deviatkin D, Selkov S, Baranov V, Kiselev A. Polycondensed Peptide Carriers Modified with Cyclic RGD Ligand for Targeted Suicide Gene Delivery to Uterine Fibroid Cells. International Journal of Molecular Sciences. 2022; 23(3):1164. https://doi.org/10.3390/ijms23031164
Chicago/Turabian StyleEgorova, Anna, Sofia Shtykalova, Marianna Maretina, Alexander Selutin, Natalia Shved, Dmitriy Deviatkin, Sergey Selkov, Vladislav Baranov, and Anton Kiselev. 2022. "Polycondensed Peptide Carriers Modified with Cyclic RGD Ligand for Targeted Suicide Gene Delivery to Uterine Fibroid Cells" International Journal of Molecular Sciences 23, no. 3: 1164. https://doi.org/10.3390/ijms23031164
APA StyleEgorova, A., Shtykalova, S., Maretina, M., Selutin, A., Shved, N., Deviatkin, D., Selkov, S., Baranov, V., & Kiselev, A. (2022). Polycondensed Peptide Carriers Modified with Cyclic RGD Ligand for Targeted Suicide Gene Delivery to Uterine Fibroid Cells. International Journal of Molecular Sciences, 23(3), 1164. https://doi.org/10.3390/ijms23031164