
Fine Regulation during Wound Healing by Mast Cells, a Physiological Role Not Yet Clarified


Abstract
:1. Introduction
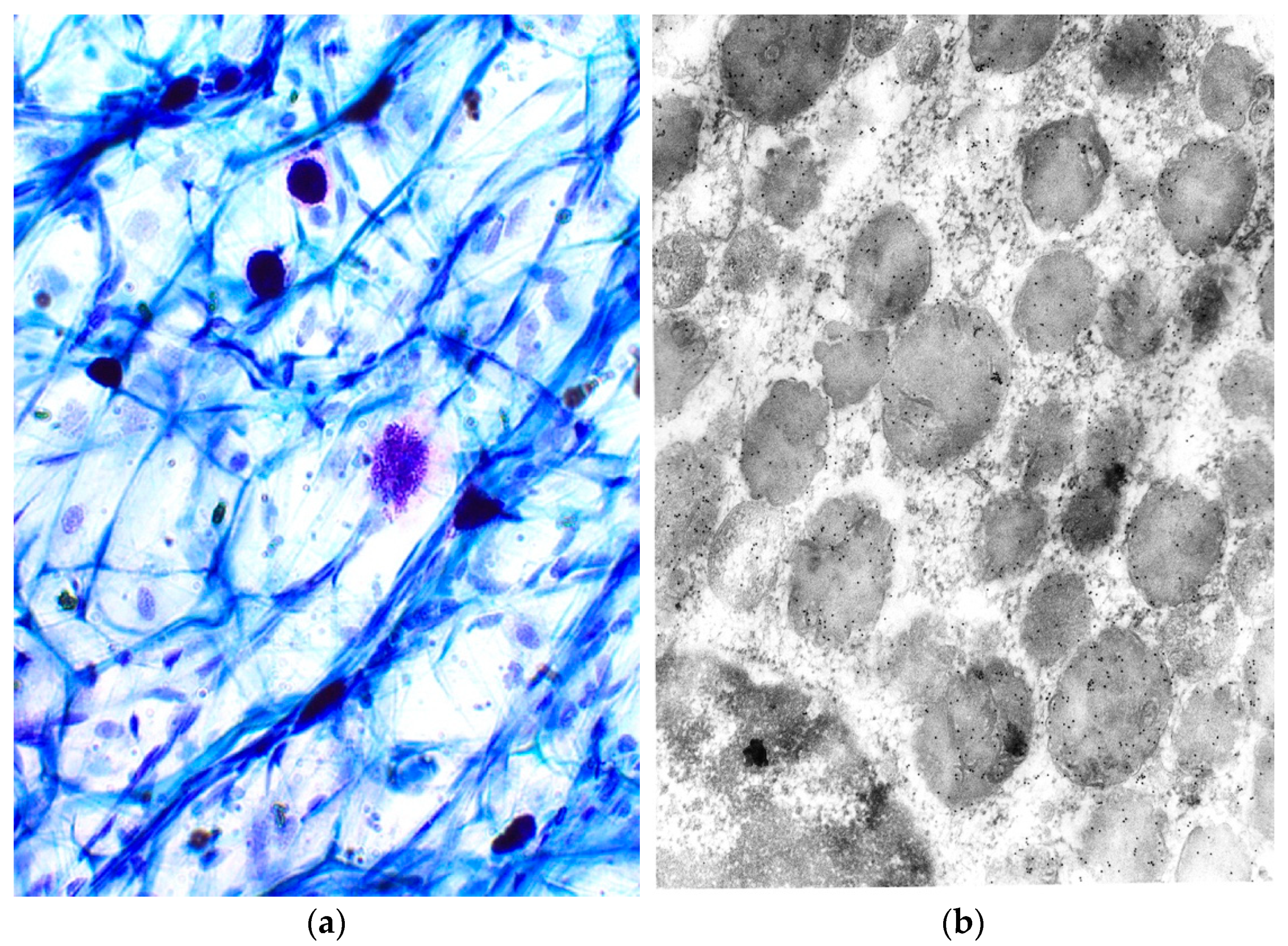
2. Structure of Mast Cells
3. Origin and Survival of Mast Cells
4. Mast Cell Heterogeneity
5. Mast Cell Relationships to Basophils
6. Mast Cell Activation
7. The Secretory Products of Mast Cells
8. Mast Cells as Effector Cells in the Immune System
9. The Wound Healing
10. Mast Cells and Wound Repair
11. Mast Cells and Scars
12. Mast Cells and Chronic Wounds
13. Neurogenic Stimuli and Mast Cells in Chronic Would Healing
14. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MCs | Mast cells |
| FcεRI | High-affinity IgE receptor |
| TNF | Tumor necrosis factor |
| VCAM | Vascular cell adhesion molecule |
| SCF | Stem cell factor |
| NGF | Nerve growth factor |
| TRAIL | TNF-related apoptosis inducing ligand |
| MCT | MCs containing only tryptase |
| MCTC | MCs containing both tryptase and chymase |
| MCC | MCs containing only chymase |
| TGF | Transforming growth factor |
| CD | Cluster of differentiation |
| IL | Interleukin |
| LYN | Lck/yes-related tyrosine kinase |
| SYK | Spleen associate tyrosine kinase |
| LAT | Linker for activation of T cells |
| NTAL | Non-T cell activation linker |
| VIP | Vaso active intestinal peptide |
| NO | Nitric oxide |
| FGF | Fibroblast growth factor |
| MCP-1 | Monocyte chemoattractant protein 1 |
| TGF | Transforming growth factor |
| VEGF | Vascular endothelial growth factor |
References
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D. The Staining of Mast Cells: A Historical Overview. Int. Arch. Allergy Immunol. 2018, 176, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Galli, J.S.; Gaudenzio, N.; Tsai, M. Mast cells in inflammation and disease: Recent progress and ongoing concerns. Annu. Rev. Immunol. 2020, 38, 49–77. [Google Scholar] [CrossRef] [PubMed]
- Saccheri, P.; Travan, L.; Ribatti, D.; Crivellato, E. Mast cells, an evolutionary approach. Ital. J. Anat. Embryol. 2020, 124, 271–287. [Google Scholar] [CrossRef]
- Fong, M.; Crane, J.S. Histology, Mast cells. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK499904 (accessed on 29 November 2021).
- Bacci, S.; Bonelli, A.; Romagnoli, P. Mast cells in injury response. In Cell Movement: New Research Trends; Abreu, T., Silva, G., Eds.; Nova Science Publishers: New York, NY, USA, 2009; pp. 81–121. [Google Scholar]
- Kirshenbaum, A.S.; Goff, J.P.; Semere, T.; Foster, B.; Scott, L.M.; Metcalfe, D.D. Demonstration that human mast cells arise from a progenitor cell population that is CD34(+), C-Kit(+), and expresses aminopeptidase N (CD13). Blood 1999, 94, 2333–2342. [Google Scholar] [CrossRef]
- Wright, H.V.; Bailey, D.; Kashyap, M.; Kepley, C.L.; Drutskaya, M.S.; Nedospasov, S.A.; Ryan, J.J. IL-3-mediated TNF production is necessary for mast cell development. J. Immunol. 2006, 176, 2114–2121. [Google Scholar] [CrossRef] [Green Version]
- Bacci, S.; Romagnoli, P. Drugs acting on mast cells function. A cell biological perspective. Inflamm. Allergy Drug Targets 2010, 9, 214–228. [Google Scholar] [CrossRef]
- Alcaide, P.; Jones, T.G.; Lord, G.M.; Glimcher, L.H.; Hallgren, J.; Arinobu, Y.; Akashi, K.; Paterson, A.M.; Gurish, M.A.; Luscinskas, F.W. Dendritic cell expression of the transcription factor T-beta regulates mast cell progenitor homing to mucosal tissue. J. Exp. Med. 2007, 204, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Gentek, R.; Ghigo, C.; Hoeffel, G.; Bulle, M.J.; Msallam, R.; Gautier, G.; Launay, P.; Chen, J.; Ginhoux, F.; Bajenoff, M. Hemogenic endothelial fate mapping reveals dual developmental origin of mast cells. Immunity 2018, 48, 1160–1171. [Google Scholar] [CrossRef] [Green Version]
- Ochi, H.; Hirani, W.M.; Yuan, Q.; Friend, D.S.; Austen, K.F.; Boyce, J.A. T helper cell type 2 cytokine-mediated comitogenic responses and CCR3 expression during differentiation of human mast cells in vitro. J. Exp. Med. 1999, 190, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Nakahata, T.; Toru, H. Cytokines regulate development of human mast cells from hematopoietic progenitors. Int. J. Hematol. 2002, 75, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Kanbe, N.; Kurosawa, M.; Miyachi, Y.; Kanbe, M.; Saitoh, H.; Matsuda, H. Nerve growth factor prevents apoptosis of cord blood-derived human cultured mast cells synergistically with stem cell factor. Clin. Exp. Allergy 2000, 30, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Berent-Maoz, B.; Piliponsky, A.M.; Daigle, I.; Simon, H.U.; Levi-Schaffer, F. Human mast cells undergo TRAIL-induced apoptosis. J. Immunol. 2006, 176, 2272–2278. [Google Scholar] [CrossRef] [PubMed]
- Okayama, Y.; Kawakami, T. Development, migration, and survival of mast cells. Immunol. Res. 2006, 34, 97–115. [Google Scholar] [CrossRef]
- da Silva, E.Z.; Jamur, M.C.; Oliver, C. Mast cell function: A new vision of an old cell. J. Histochem. Cytochem. 2014, 62, 698–738. [Google Scholar] [CrossRef] [PubMed]
- Crivellato, E.; Feltrami, C.A.; Mallardi, F.; Ribatti, D. The mast cell: An active participant or an innocent bystander? Histol. Histopathol. 2004, 19, 259–270. [Google Scholar] [CrossRef]
- Elieh Ali Komi, D.; Wöhrl, S.; Bielory, L. Mast Cell Biology at Molecular Level: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2020, 58, 342–365. [Google Scholar] [CrossRef]
- Varricchi, G.; Pecoraro, A.; Loffredo, S.; Poto, R.; Rivellese, F.; Genovese, A.; Marone, G.; Spadaro, G. Heterogeneity of human mast cells with respect to MRGPR2 receptor expression and function. Front. Cell. Neurosci. 2019, 13, 299. [Google Scholar] [CrossRef] [Green Version]
- Kocabas, C.N.; Yavuz, A.S.; Lipsky, P.E.; Metcalfe, D.D.; Akin, C. Analysis of the lineage relationship between mast cells and basophils using the C-Kit D816V mutation as a biologic signature. J. Allergy Clin. Immunol. 2005, 115, 1155–1161. [Google Scholar] [CrossRef]
- Crivellato, E.; Travan, L.; Ribatti, D. Mast cells and basophils: A potential link in promoting angiogenesis during allergic inflammation. Int. Arch. Allergy Immunol. 2010, 151, 89–97. [Google Scholar] [CrossRef]
- Voehringer, D. Protective and pathological roles of mast cells and basophils. Nat. Rev. Immunol. 2013, 13, 362–375. [Google Scholar] [CrossRef]
- Kubo, M. Mast cells and basophils in allergic inflammation. Curr. Opin. Immunol. 2018, 54, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Varricchi, G.; Raap, U.; Rivellese, F.; Marone, G.; Gibbs, B.F. Human mast cells and basophils-How are they similar how are they different? Immunol. Rev. 2018, 282, 8–34. [Google Scholar] [CrossRef] [PubMed]
- Moon, T.C.; Befus, A.D.; Kulka, M. Mast cell mediators: Their differential release and the secretory pathways involved. Front. Immunol. 2014, 5, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Blokhuis, B.R.; Garssen, J.; Redegeld, F.A. Non-IgE mediated mast cell activation. Eur. J. Pharmavol. 2016, 778, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Tsilioni, I.; Ren, H. Recent advances in our understanding of mast cell activation—Or should it be mast cell mediator disorders? Expert Rev. Clin. Immunol. 2019, 15, 639–656. [Google Scholar] [CrossRef]
- Castells, M.C. Mast cells mediators in allergic inflammation and mastocytosis. Immunol. Allergy Clin. North Am. 2006, 26, 465–485. [Google Scholar] [CrossRef]
- González-de-Olano, D.; Álvarez-Twose, I. Mast Cells as Key Players in Allergy and Inflammation. J. Investig. Allergol. Clin. Immunol. 2018, 28, 365–378. [Google Scholar] [CrossRef]
- Monument, M.J.; Hart, D.A.; Salo, P.T.; Befus, A.D.; Hildebrand, K.A. Neuroinflammatory Mechanisms of Connective Tissue Fibrosis: Targeting Neurogenic and Mast Cell Contributions. Adv. Wound Care 2015, 4, 137–151. [Google Scholar] [CrossRef] [Green Version]
- Wernersson, S.; Pejler, G. Mast cell secretory granules: Armed for battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef]
- Pastwińska, J.; Agier, J.; Dastych, J.; Brzezińska-Błaszczyk, E. Mast cells as the strength of the inflammatory process. Pol. J. Pathol. 2017, 68, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Bin, N.R.; Sugita, S. Diverse exocytic pathways for mast cell mediators. Biochem. Soc. Trans. 2018, 46, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Agier, J.; Pastwińska, J.; Brzezińska-Błaszczyk, E. An overview of mast cell pattern recognition receptors. Inflamm. Res. 2018, 67, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Castells, C.M.; Bankova, L. Mast Cells: Surface Receptors and Signal Transduction, 2021. Uptodate. Available online: https://www.uptodate.com/contents/mast-cells-surface-receptors-and-signal-transduction (accessed on 29 November 2021).
- Varricchi, G.; Rossi, F.W.; Galdiero, M.R.; Granata, F.; Criscuolo, G.; Spadaro, G.; de Paulis, A.; Marone, G. Physiological Roles of Mast Cells: Collegium Internationale Allergologicum Update 2019. Int. Arch. Allergy Immunol. 2019, 179, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. New Eng. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Darby, I.A.; Laverdet, B.; Bonté, F.; Desmoulière, A. Fibroblasts and myofibroblasts in wound healing. Clin. Cosmet. Investig. Dermatol. 2014, 7, 301–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, A.C.; Costa, T.F.; Andrade, Z.A.; Medrado, A.R. Wound healing—A literature review. An. Bras. De Dermatol. 2016, 91, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Cañedo-Dorantes, L.; Cañedo-Ayala, M. Skin Acute Wound Healing: A Comprehensive Review. Int. J. Inflam. 2019, 2019, 3706315. [Google Scholar] [CrossRef]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound Healing: A Cellular Perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef]
- Visha, M.G.; Karunagaran, M. A review on wound healing. Int. J. Clinicopathol. Correl. 2019, 3, 50–59. [Google Scholar] [CrossRef]
- Bacci, S. Cutaneous wound healing: Cellular mechanisms and therapies (an update). Med. Res. Arch. 2019, 7, 12. [Google Scholar]
- Tottoli, E.M.; Dorati, R.; Genta, I.; Chiesa, E.; Pisani, S.; Conti, B. Skin Wound Healing Process and New Emerging Technologies for Skin Wound Care and Regeneration. Pharmaceutics 2020, 12, 735. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, H.N.; Hardman, M.J. Wound healing: Cellular mechanisms and pathological outcomes. Open Biol. 2020, 10, 200223–200236. [Google Scholar] [CrossRef]
- Weller, K.; Foitzik, K.; Paus, R.; Syska, W.; Maurer, M. Mast cells are required for normal healing of skin wounds in mice. FASEB J. 2006, 20, 2366–2368. [Google Scholar] [CrossRef] [PubMed]
- Maurer, M.; Opitz, M.; Henz, B.M.; Paus, R. The mast cell products histamine and serotonin stimulate and TNF-alpha inhibits the proliferation of murine epidermal keratinocytes in situ. J. Dermatol. Sci. 1997, 16, 79–84. [Google Scholar] [CrossRef]
- Gailit, J.; Marchese, M.J.; Kew, R.R.; Gruber, B.L. The differentiation and function of myofibroblasts is regulated by mast cell mediators. J. Investig. Dermatol. 2001, 117, 1113–1119. [Google Scholar] [CrossRef] [Green Version]
- Levi-Schaffer, F.; Kupietzky, A. Mast cells enhance migration and proliferation of fibroblasts into an in vitro wound. Exp. Cell. Res. 1990, 188, 42–49. [Google Scholar] [CrossRef]
- Norrby, K.; Jakobsson, A.; Sörbo, J. Mast-cell-mediated angiogenesis: A novel experimental model using the rat mesentery. Virchows Arch. B 1986, 52, 195–206. [Google Scholar] [CrossRef]
- Egozi, E.I.; Ferreira, A.M.; Burns, A.L.; Gamelli, R.L.; Dipietro, L.A. Mast cells modulate the inflammatory but not the proliferative response in healing wounds. Wound Repair Regen. 2003, 11, 46–54. [Google Scholar] [CrossRef]
- Artuc, M.; Hermes, B.; Steckelings, U.M.; Grützkau, A.; Henz, B.M. Mast cells and their mediators in cutaneous wound healing--active participants or innocent bystanders? Exp. Dermatol. 1999, 8, 1–16. [Google Scholar] [CrossRef]
- Baum, C.L.; Arpey, C.J. Normal cutaneous wound healing: Clinical correlation with cellular and molecular events. Dermatol. Surg. 2005, 31, 674–686. [Google Scholar] [CrossRef] [PubMed]
- Bonelli, A.; Bacci, S.; Norelli, G.A. Affinity cytochemistry analysis of mast cells in skin lesions: A possible tool to assess the timing of lesions after death. Int. J. Leg. Med. 2003, 117, 331–334. [Google Scholar] [CrossRef] [PubMed]
- Trautmann, A.; Toksoy, A.; Engelhardt, E.; Bröcker, E.B.; Gillitzer, R. Mast cell involvement in normal human skin wound healing; expression of monocyte chemoattractant protein-1 is correlated with recruitment of mast cells which synthesize interleukin-4 in vivo. J. Pathol. 2000, 190, 100–106. [Google Scholar] [CrossRef]
- Oehmichen, M.; Gronki, T.; Meissner, C.; Anlauf, M.; Schwark, T. Mast cell reactivity at the margin of human skin wounds: An early cell marker of wound survival? Forensic Sci. Int. 2009, 191, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bacci, S.; Romagnoli, P.; Norelli, G.A.; Forestieri, A.L.; Bonelli, A. Early increase in TNF-alpha-containing mast cells in skin lesions. Int. J. Leg. Med. 2006, 120, 138–142. [Google Scholar] [CrossRef]
- Steinhoff, M.; Buddenkotte, J.; Shpacovitch, V.; Rattenholl, A.; Moormann, C.; Vergnolle, N.; Luger, T.A.; Hollenberg, M.D. Proteinase-activated receptors: Transducers of proteinase-mediated signaling in inflammation and immune response. Endocr. Rev. 2005, 26, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Terakawa, M.; Tomimori, Y.; Goto, M.; Fukuda, Y. Mast cell chymase induces expression of chemokines for neutrophils in eosinophilic EoL-1 cells and mouse peritonitis eosinophils. Eur. J. Pharmacol. 2006, 538, 175–181. [Google Scholar] [CrossRef]
- Abdel-Majid, R.M.; Marshall, J.S. Prostaglandin E2 induces degranulation-independent production of vascular endothelial growth factor by human mast cells. J. Immunol. 2004, 172, 1227–1236. [Google Scholar] [CrossRef] [Green Version]
- Muramatsu, M.; Katada, J.; Hattori, M.; Hayashi, I.; Majima, M. Chymase mediates mast cell-induced angiogenesis in hamster sponge granulomas. Eur. J. Pharmacol. 2000, 18, 181–191. [Google Scholar] [CrossRef]
- Doggrell, S.A.; Wanstall, J.C. Vascular chymase: Pathophysiological role and therapeutic potential of inhibition. Cardiovasc. Res. 2004, 61, 653–662. [Google Scholar] [CrossRef] [Green Version]
- Somasundaram, P.; Ren, G.; Nagar, H.; Kraemer, D.; Mendoza, L.; Michael, L.H.; Caughey, G.H.; Entman, M.L.; Frangogiannis, N.G. Mast cell tryptase may modulate endothelial cell phenotype in healing myocardial infarcts. J. Pathol. 2005, 205, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Presta, M.; Leali, D.; Stabile, H.; Ronca, R.; Camozza, M.; Coco, L.; Moroni, E.; Liekens, S.; Rusnati, M. Heparin derivatives as angiogenesis inhibitors. Curr. Pharm. Des. 2003, 9, 553–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, M.; Kurosawa, M.; Ishikawa, O.; Miyachi, Y.; Kido, H. Mast cell tryptase stimulates both human dermal fibroblast proliferation and type I collagen production. Clin. Exp. Allergy 1998, 28, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Komi, D.; Khomtchouk, K.; Santa Maria, P.L. A review of the contribution of mast cells in wound healing: Involved molecular and cellular mechanisms. Clin. Rev. Allergy Immunol. 2020, 58, 298–312. [Google Scholar] [CrossRef]
- van der Veer, W.M.; Bloemen, M.C.; Ulrich, M.M.; Molema, G.; van Zuijlen, P.P.; Middelkoop, E.; Niessen, F.B. Potential cellular and molecular causes of hypertrophic scar formation. Burns 2009, 35, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Wulff, B.C.; Parent, A.E.; Meleski, M.A.; DiPietro, L.A.; Schrementi, M.E.; Wilgus, T.A. Mast cells contribute to scar formation during fetal wound healing. J. Investig. Dermatol. 2012, 132, 458–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ud-Din, S.; Wilgus, T.A.; Bayat, A. Mast cells in skin scarring: A review of animal and human research. Front. Immunol. 2020, 11, 552205. [Google Scholar] [CrossRef]
- Wilgus, T.A.; Ud-Din, S.; Bayat, A. A Review of the Evidence for and against a role for mast cells in cutaneous scarring and fibrosis. Int. J. Molec. Sci. 2020, 21, 9673. [Google Scholar] [CrossRef]
- Chen, L.; Schrementi, M.E.; Ranzer, M.J.; Wilgus, T.A.; DiPietro, L.A. Blockade of mast cell activation reduces cutaneous scar formation. PLoS ONE 2014, 22, e85226. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.C.; Zhao, W.Y.; Cao, Y.; Liu, Y.Q.; Sun, Q.; Shi, P.; Cai, J.Q.; Shen, X.Z.; Tan, W.Q. The Roles of Inflammation in keloid and hypertrophic scars. Front. Immunol. 2020, 11, 603187. [Google Scholar] [CrossRef]
- Kyaw, B.M.; Järbrink, K.; Martinengo, L.; Car, J.; Harding, K.; Schmidtchen, A. Need for Improved Definition of “Chronic Wounds” in Clinical Studies. Acta Derm. Venereol. 2018, 98, 157–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottrup, F. A specialized wound-healing center concept: Importance of a multidisciplinary department structure and surgical treatment facilities in the treatment of chronic wounds. Am. J. Surg. 2004, 187, 8S–43S. [Google Scholar] [CrossRef]
- Huttunen, M.; Aalto, M.L.; Harvima, R.J.; Horsmanheimo, M.; Harvima, I.T. Alterations in mast cells showing tryptase and chymase activity in epithelializating and chronic wounds. Exp. Dermatol. 2000, 9, 258–265. [Google Scholar] [CrossRef]
- Huttunen, M.; Naukkarinen, A.; Horsmanheimo, M.; Harvima, I.T. Transient production of stem cell factor in dermal cells but increasing expression of Kit receptor in mast cells during normal wound healing. Arch. Dermatol. Res. 2002, 294, 324–330. [Google Scholar] [CrossRef]
- Corsi, A.; Lecci, P.P.; Bacci, S.; Cappugi, P.; Pimpinelli, N. Early activation of fibroblasts during PDT treatment in leg ulcers. G. Ital. Dermatol. Venereol. 2016, 151, 223–229. [Google Scholar]
- Grandi, V.; Bacci, S.; Corsi, A.; Sessa, M.; Puliti, E.; Murciano, N.; Scavone, F.; Cappugi, P.; Pimpinelli, N. ALA-PDT exerts beneficial effects on chronic venous ulcers by inducing changes in inflammatory microenvironment, especially through increased TGF-beta release: A pilot clinical and translational study. Photodiagnosis Photodyn. Ther. 2018, 21, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Grandi, V.; Paroli, G.; Puliti, E.; Bacci, S.; Pimpinelli, N. Single ALA-PDT irradiation induces increase in mast cells degranulation and neuropeptide acute response in chronic venous ulcers: A pilot study. Photodiagnosis Photodyn. Ther 2021, 34, 102222. [Google Scholar] [CrossRef]
- Bacci, S. Cellular Mechanisms and Therapies in Wound Healing: Looking toward the Future. Biomed. 2021, 9, 1611. [Google Scholar] [CrossRef]


{kind=link}
| MCT | MCTC | |
|---|---|---|
| Shape | Small, sparsely granulated | Large, densely granulated |
| Enzyme content of granules | Tryptase and others | Chymase, tryptase, and others |
| Response to compound 48/80 | Proliferative | Secretory |
| Sensitivity to disodium-cromoglycate | None | Yes |
| Life span | Short | Long |
| Preferential location in humans | Lung, bowel | Skin, bowel |
| Antigens | CD50 | CD32 |
| Cytokines | GM-CSF, TGF-β | IL3 |
| Complement receptor | None | CD88 (C5aR) |
| Basophils | MC | |
|---|---|---|
| Diameter | 8–9 μm | 13–14 μm |
| Granules | Few | Numerous |
| Compound exocytosis | Rare | Common |
| Tryptase in granules | No | Yes |
| Expression of C-Kit | No | Yes |
| Mediators | Functions |
|---|---|
| Histamine | Activation of a group of suppressor T cells |
| Prostaglandin D2 | Inhibition of the activity of helper T cells, stimulation of the differentiation and function of suppressor T cells, inhibition of IgE production |
| Leukotrienes | Similar function of prostaglandin D2 and inhibition of the differentiation of plasma cells. |
| VIP | Inhibition of the secretory and proliferative responses of at least some of the subgroups of T and B cells. |
| Heparin (low concentration) | Activation of macrophages to produce IL-1, which in turn affects both the macrophages themselves and the surrounding cells and the whole organism |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bacci, S. Fine Regulation during Wound Healing by Mast Cells, a Physiological Role Not Yet Clarified. Int. J. Mol. Sci. 2022, 23, 1820. https://doi.org/10.3390/ijms23031820
Bacci S. Fine Regulation during Wound Healing by Mast Cells, a Physiological Role Not Yet Clarified. International Journal of Molecular Sciences. 2022; 23(3):1820. https://doi.org/10.3390/ijms23031820
Chicago/Turabian StyleBacci, Stefano. 2022. "Fine Regulation during Wound Healing by Mast Cells, a Physiological Role Not Yet Clarified" International Journal of Molecular Sciences 23, no. 3: 1820. https://doi.org/10.3390/ijms23031820
APA StyleBacci, S. (2022). Fine Regulation during Wound Healing by Mast Cells, a Physiological Role Not Yet Clarified. International Journal of Molecular Sciences, 23(3), 1820. https://doi.org/10.3390/ijms23031820