
Interdisciplinary Study of the Effects of Dipeptidyl-Peptidase III Cancer Mutations on the KEAP1-NRF2 Signaling Pathway

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Peptidase Activity of DPP III Variants
2.2. MST Study of the Effects of DPP III Mutations on its Binding Affinity for the Kelch Domain
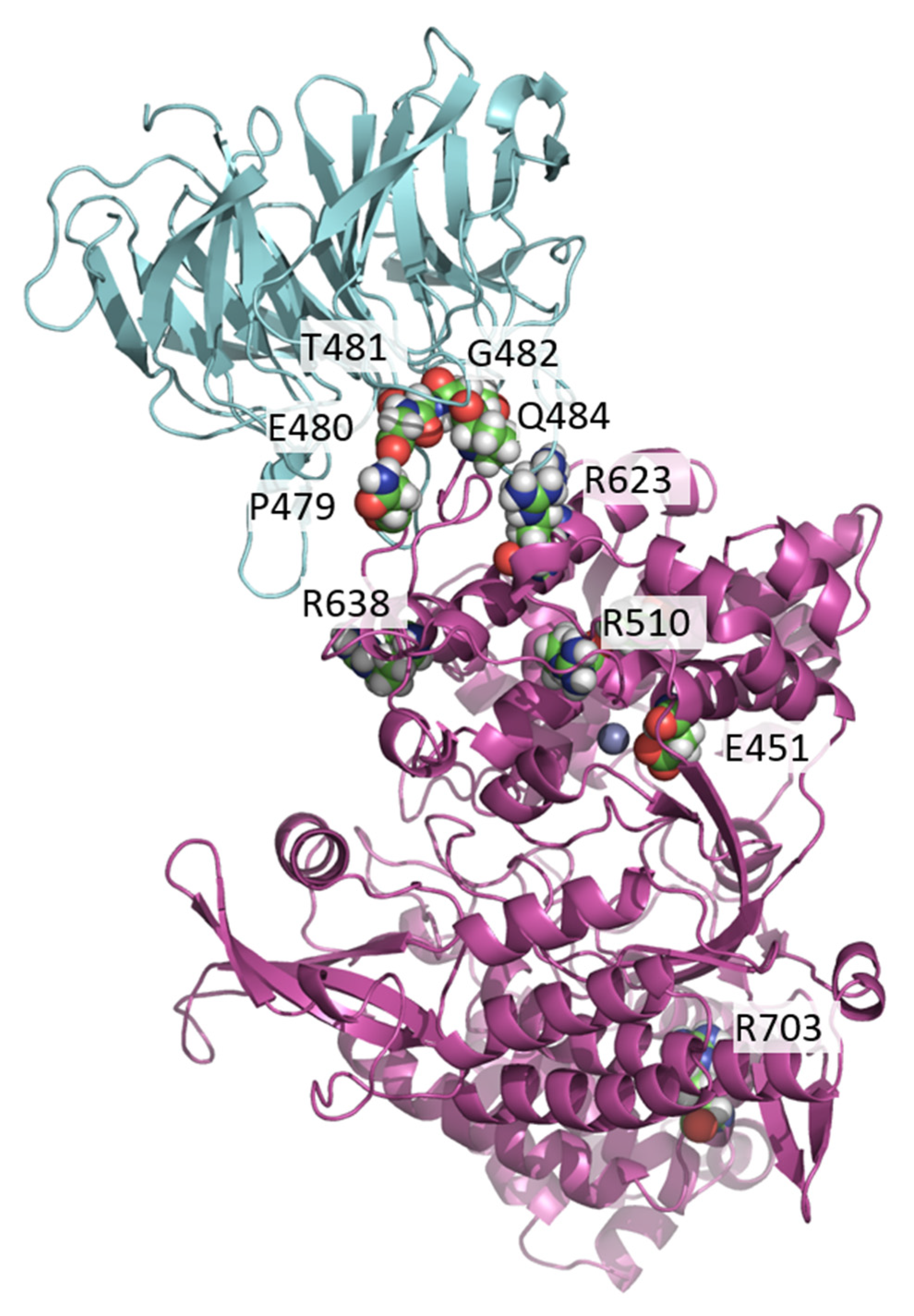
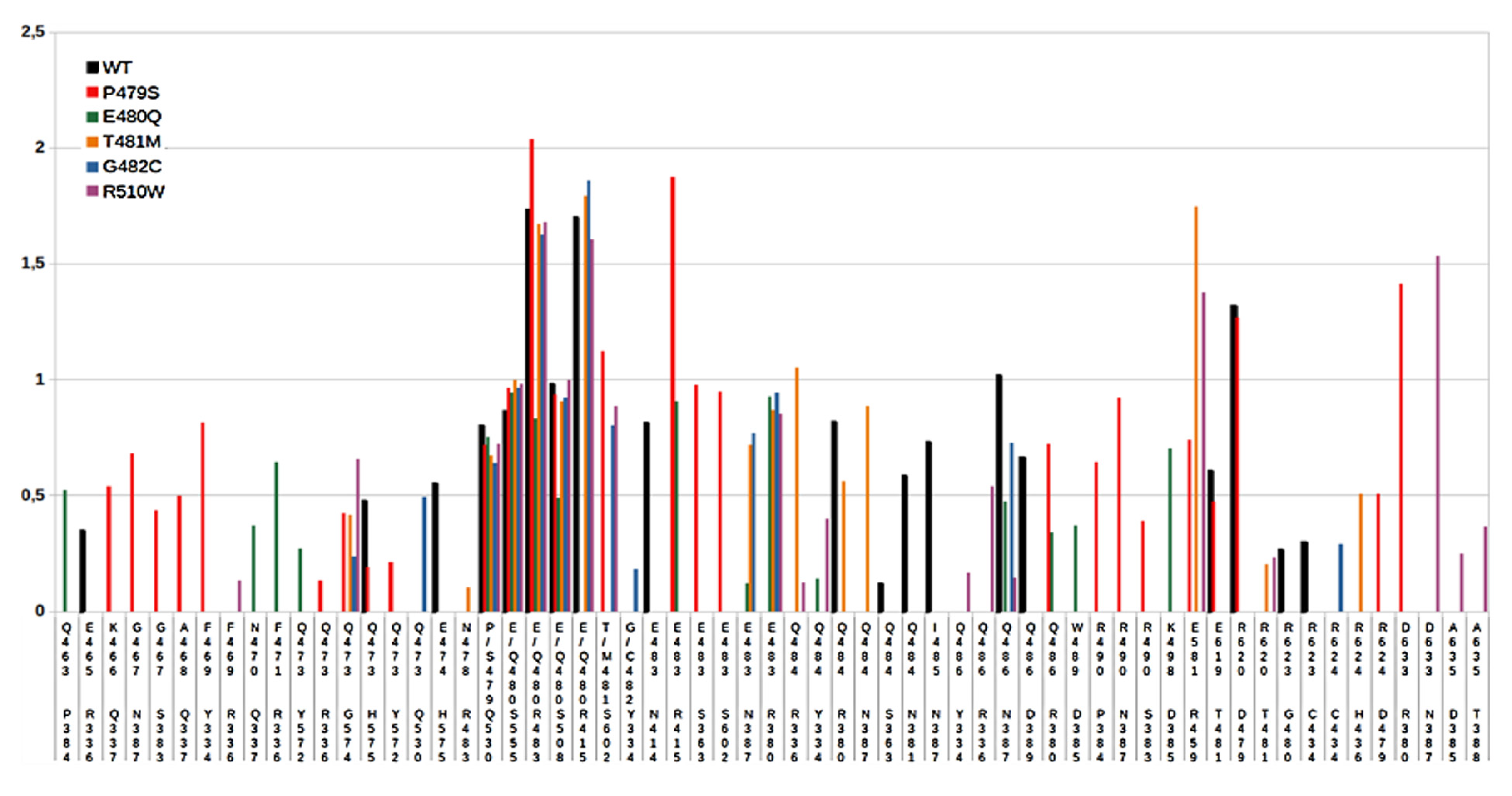
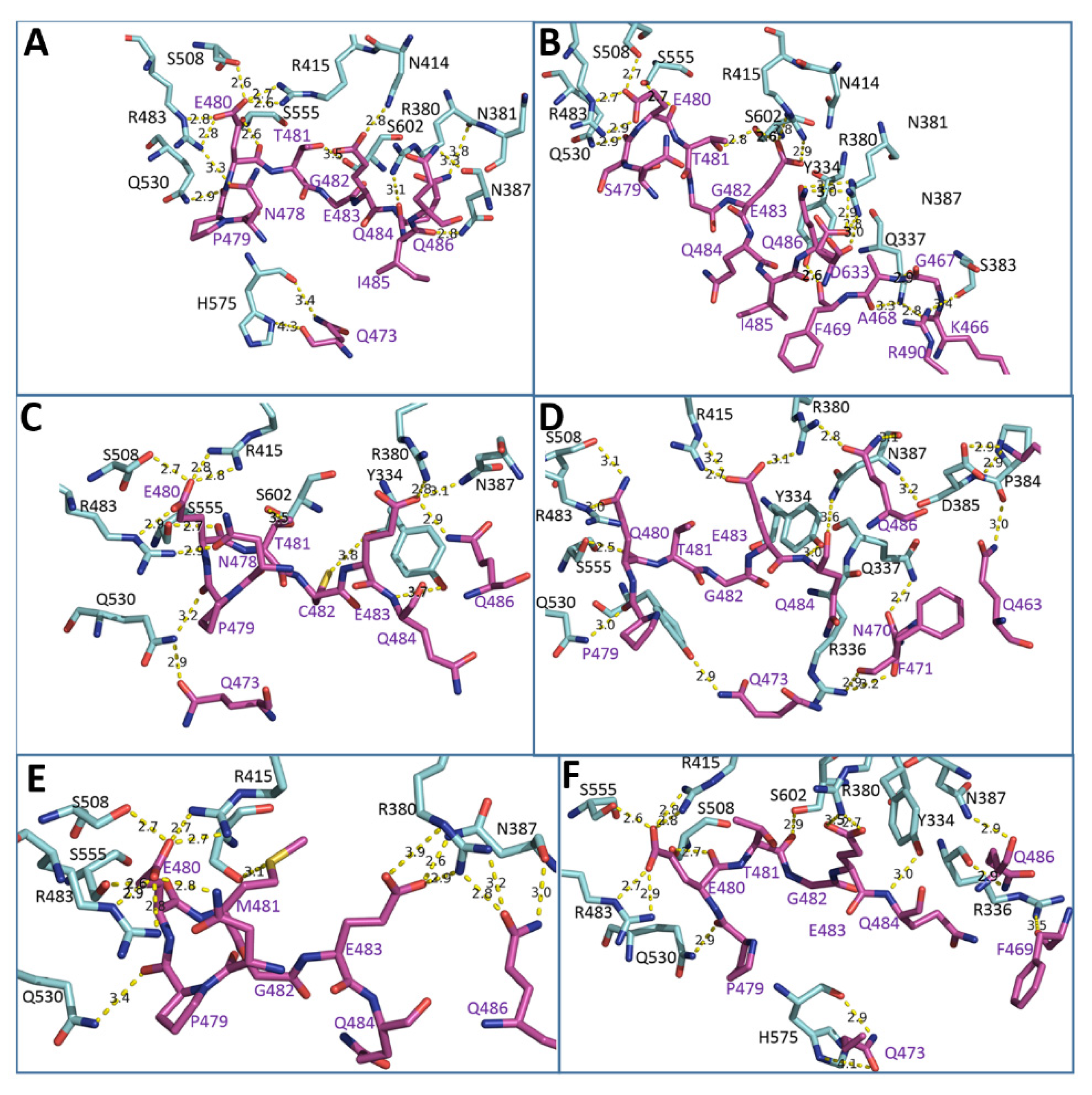
2.3. MD Simulations and Computational Analysis
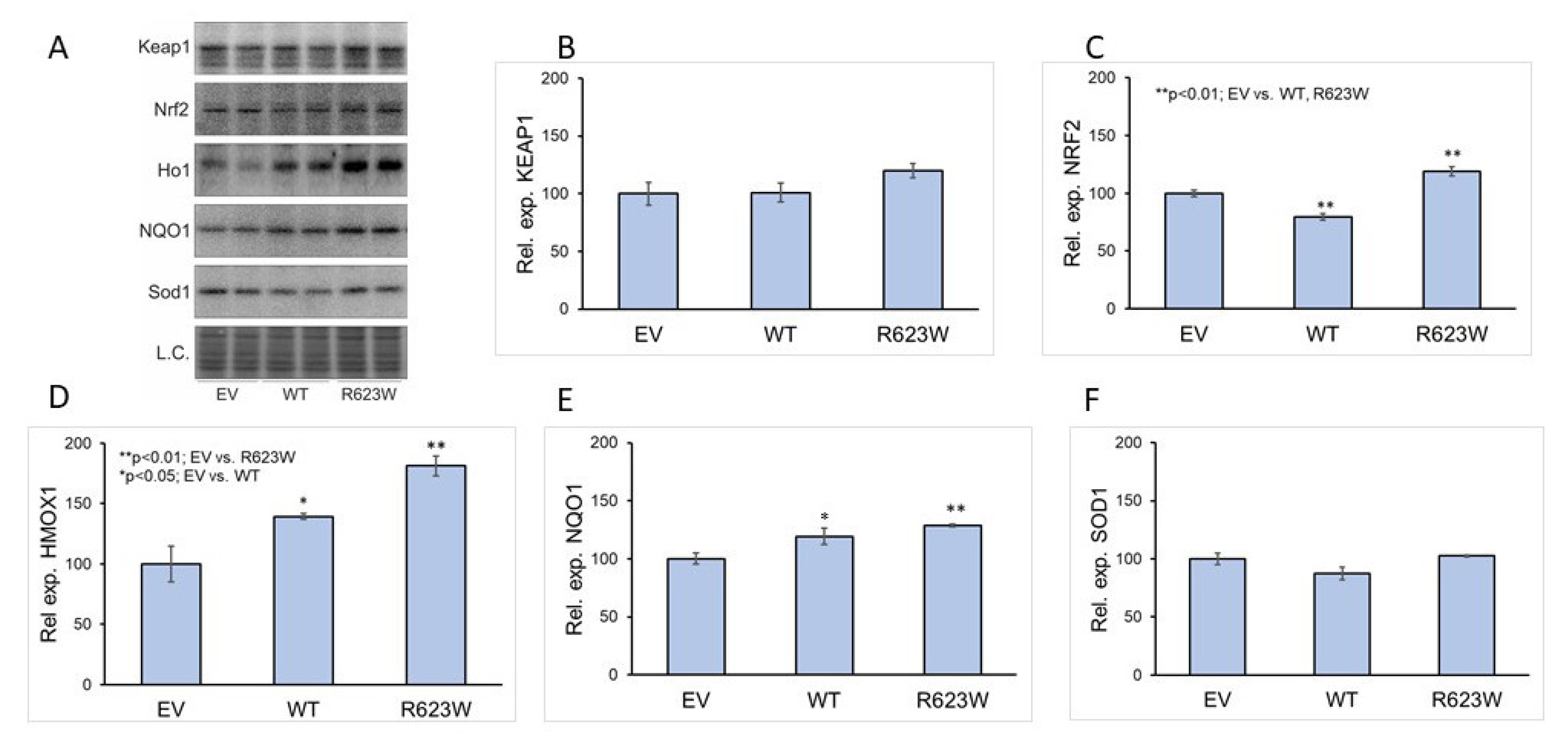
2.4. Overexpression of DPP III Induces the Expression of NQO1
3. Discussion
4. Materials and Methods
4.1. Protein Expression and Purification
4.2. Enzyme Kinetic
4.3. Binding Affinity
4.4. Computational Study
4.4.1. System Preparations, Parameterization and Simulations
4.4.2. Classical MD Simulations
4.4.3. Data Analysis
4.5. Cloning and Cell Culture
4.6. RNA Isolation and RT-qPCR
4.7. RNA Isolation Protein Isolation and Western Blot
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Abramić, M.; Zubanović, M.; Vitale, L. Dipeptidyl Peptidase III from Human Erythrocytes. Biol. Chem. Hoppe-Seyler. 1988, 369, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.J.; Chen, J.-M. Dipeptidyl-peptidase III. In Handbook of Proteolytic Enzymes; Rawlings, N.D., Salvesen, G., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2013; ISBN 9780123822192. [Google Scholar]
- Šimaga, Š.; Babić, D.; Osmak, M.; Ilić-Forko, J.; Vitale, L.; Miličić, D.; Abramić, M. Dipeptidyl peptidase III in malignant and non-malignant gynaecological tissue. Eur. J. Cancer 1998, 34, 399–405. [Google Scholar] [CrossRef]
- Šimaga, Š.; Babić, D.; Osmak, M.; Šprem, M.; Abramić, M. Tumor cytosol dipeptidyl peptidase III activity is increased with histological aggressiveness of ovarian primary carcinomas. Gynecol. Oncol. 2003, 91, 194–200. [Google Scholar] [CrossRef]
- Gamrekelashvili, J.; Greten, T.F.; Korangy, F. Immunogenicity of necrotic cell death. Cell. Mol. Life Sci. 2015, 72, 273–283. [Google Scholar] [CrossRef]
- Gamrekelashvili, J.; Kapanadze, T.; Han, M.; Wissing, J.; Ma, C.; Jaensch, L.; Manns, M.P.; Armstrong, T.; Jaffee, E.; White, A.O.; et al. Peptidases released by necrotic cells control CD8+ T cell cross-priming. J. Clin. Investig. 2013, 123, 4755–4768. [Google Scholar] [CrossRef] [Green Version]
- Miettinen, J.J.; Kumari, R.; Traustadottir, G.A.; Huppunen, M.-E.; Sergeev, P.; Majumder, M.M.; Schepsky, A.; Gudjonsson, T.; Lievonen, J.; Bazou, D.; et al. Aminopeptidase expression in multiple myeloma associates with disease progression and sensitivity to melflufen. Cancers 2021, 13, 1527. [Google Scholar] [CrossRef]
- Hast, B.E.; Goldfarb, D.; Mulvaney, K.M.; Hast, M.A.; Siesser, P.F.; Yan, F.; Hayes, D.N.; Major, M.B. Proteomic analysis of ubiquitin ligase KEAP1 reveals associated proteins that inhibit NRF2 ubiquitination. Cancer Res. 2013, 73, 2199–2210. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.; Alcivar, A.L.; Ma, J.; Foo, T.K.; Zywea, S.; Mahdi, A.; Huo, Y.; Kensler, T.W.; Gatza, M.L.; Xia, B. NRF2 induction supporting breast cancer cell survival is enabled by oxidative stress-induced DPP3-KEAP1 interaction. Cancer Res. 2017, 77, 2881–2892. [Google Scholar] [CrossRef] [Green Version]
- Tong, K.I.; Katoh, Y.; Kusunoki, H.; Itoh, K.; Tanaka, T.; Yamamoto, M. Keap1 Recruits Neh2 through Binding to ETGE and DLG Motifs: Characterization of the Two-Site Molecular Recognition Model. Mol. Cell. Biol. 2006, 26, 2887–2900. [Google Scholar] [CrossRef] [Green Version]
- Rojo de la Vega, M.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef]
- Jaramillo, M.C.; Zhang, D.D. The emerging role of the Nrf2—Keap1 signaling pathway in cancer. Genes Dev. 2013, 27, 2179–2191. [Google Scholar] [CrossRef] [Green Version]
- Taguchi, K.; Yamamoto, M. The KEAP1–NRF2 System in Cancer. Front. Oncol. 2017, 7, 85. [Google Scholar] [CrossRef]
- Menale, C.; Robinson, L.J.; Palagano, E.; Rigoni, R.; Erreni, M.; Almarza, A.J.; Strina, D.; Mantero, S.; Lizier, M.; Forlino, A.; et al. Absence of Dipeptidyl Peptidase 3 Increases Oxidative Stress and Causes Bone Loss. J. Bone Miner. Res. 2019, 34, 2133–2148. [Google Scholar] [CrossRef]
- Ren, X.; Yu, J.; Guo, L.; Ma, H. Dipeptidyl-peptidase 3 protects oxygen-glucose deprivation/reoxygenation-injured hippocampal neurons by suppressing apoptosis, oxidative stress and inflammation via modulation of Keap1/Nrf2 signaling. Int. Immunopharmacol. 2021, 96, 107595. [Google Scholar] [CrossRef]
- Gundić, M.; Tomić, A.; Wade, R.C.R.C.; Matovina, M.; Karačić, Z.; Kazazić, S.; Tomić, S. Human DPP III—Keap1 interactions: A combined experimental and computational study. Croat. Chem. Acta 2016, 89, 217–228. [Google Scholar] [CrossRef]
- Matić, S.; Kekez, I.; Tomin, M.; Bogár, F.; Šupljika, F.; Kazazić, S.; Hanić, M.; Jha, S.; Brkić, H.; Bourgeois, B.; et al. Binding of dipeptidyl peptidase III to the oxidative stress cell sensor Kelch-like ECH-associated protein 1 is a two-step process. J. Biomol. Struct. Dyn. 2021, 39, 6870–6881. [Google Scholar] [CrossRef]
- Matovina, M.; Agić, D.; Abramić, M.; Matić, S.; Karačić, Z.; Tomić, S. New findings about human dipeptidyl peptidase III based on mutations found in cancer. RSC Adv. 2017, 7, 36326–36334. [Google Scholar] [CrossRef] [Green Version]
- Zhong, M.; Lynch, A.; Muellers, S.N.; Jehle, S.; Luo, L.; Hall, D.R.; Iwase, R.; Carolan, J.P.; Egbert, M.; Wakefield, A.; et al. Interaction Energetics and Druggability of the Protein-Protein Interaction between Kelch-like ECH-Associated Protein 1 (KEAP1) and Nuclear Factor Erythroid 2 like 2 (Nrf2). Biochemistry 2020, 59, 563–581. [Google Scholar] [CrossRef]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonenn, A.L. The Keap1-Nrf2 pathway: Mechanisms of activation and dysregulation in cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Bailey, M.H.; Tokheim, C.; Porta-Pardo, E.; Sengupta, S.; Bertrand, D.; Weerasinghe, A.; Colaprico, A.; Wendl, M.C.; Kim, J.; Reardon, B.; et al. Comprehensive Characterization of Cancer Driver Genes and Mutations. Cell 2018, 173, 371–385. [Google Scholar] [CrossRef] [Green Version]
- Cino, E.A.; Killoran, R.C.; Karttunen, M.; Choy, W.Y. Binding of disordered proteins to a protein hub. Sci. Rep. 2013, 3, 2305. [Google Scholar] [CrossRef] [Green Version]
- Jha, S.; Taschler, U.; Domenig, O.; Poglitsch, M.; Bourgeois, B.; Pollheimer, M.; Pusch, L.M.; Malovan, G.; Frank, S.; Madl, T.; et al. Dipeptidyl peptidase 3 modulates the renin-Angiotensin system in mice. J. Biol. Chem. 2020, 295, 13711–13723. [Google Scholar] [CrossRef]
- Suzuki, T.; Muramatsu, A.; Saito, R.; Iso, T.; Shibata, T.; Kuwata, K.; Kawaguchi, S.; Iwawaki, T.; Adachi, S.; Suda, H.; et al. Molecular Mechanism of Cellular Oxidative Stress Sensing by Keap1. Cell Rep. 2019, 28, 746–758. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.C.; Lin, R.J.; Cheng, J.Y.; Wang, S.H.; Yu, J.C.; Wu, J.C.; Liang, Y.J.; Hsu, H.M.; Yu, J.; Yu, A.L. FAM129B, an antioxidative protein, reduces chemosensitivity by competing with Nrf2 for Keap1 binding. EBioMedicine 2019, 45, 25. [Google Scholar] [CrossRef] [Green Version]
- Špoljarić, J.; Tomić, A.; Vukelić, B.; Salopek-Sondi, B.; Agić, D.; Tomić, S.; Abramić, M. Human Dipeptidyl Peptidase III: The Role of Asn406 in Ligand Binding and Hydrolysis. Croat. Chem. Acta 2011, 84, 259–268. [Google Scholar] [CrossRef]
- Abramić, M.; Šimaga, Š.; Osmak, M.; Čičin-Šain, L.; Vukelić, B.; Vlahoviček, K.; Dolovčak, L. Highly reactive cysteine residues are part of the substrate binding site of mammalian dipeptidyl peptidases III. Int. J. Biochem. Cell Biol. 2004, 36, 434–446. [Google Scholar] [CrossRef]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [Green Version]
- Tomić, A.; Horvat, G.; Ramek, M.; Agić, D.; Brkić, H.; Tomić, S. New Zinc Ion Parameters Suitable for Classical MD Simulations of Zinc Metallopeptidases. J. Chem. Inf. Model. 2019, 59, 3437–3453. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Joung, I.S.; Cheatham, T.E. Determination of alkali and halide monovalent ion parameters for use in explicitly solvated biomolecular simulations. J. Phys. Chem. B 2008, 112, 9020–9041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Case, D.A.; Betz, R.; Botello-Smith, W.; Cerutti, D.S.; Cheatham, T.E.; Darden, T.A.; Duke, R.E.; TGiese, T.J.; Gohlke, H.; Goetz, A.W.; et al. AMBER 2016; University California: San Francisco, CA, USA, 2016. [Google Scholar]
- Miyamoto, S.; Kollman, P.A. Settle: An analytical version of the SHAKE and RATTLE algorithm for rigid water models. J. Comput. Chem. 1992, 13, 952–962. [Google Scholar] [CrossRef]
- Ryckaert, J.P.; Ciccotti, G.; Berendsen, H.J.C. Numerical integration of the cartesian equations of motion of a system with constraints: Molecular dynamics of n-alkanes. J. Comput. Phys. 1977, 23, 327–341. [Google Scholar] [CrossRef] [Green Version]
- Berendsen, H.J.C.; Postma, J.P.M.; van Gunsteren, W.F.; Dinola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef] [Green Version]
- Loncharich, R.J.; Brooks, B.R.; Pastor, R.W. Langevin dynamics of peptides: The frictional dependence of isomerization rates of N-acetylalanyl-N′-methylamide. Biopolymers 1992, 32, 523–535. [Google Scholar] [CrossRef]
- Roe, D.R.; Cheatham, T.E. PTRAJ and CPPTRAJ: Software for processing and analysis of molecular dynamics trajectory data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef]
- Miller, B.R.; McGee, T.D.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA.py: An efficient program for end-state free energy calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef]
- Tomić, A.; Kovačević, B.; Tomić, S. Concerted nitrogen inversion and hydrogen bonding to Glu451 are responsible for protein-controlled suppression of the reverse reaction in human DPP III. Phys. Chem. Chem. Phys. 2016, 18, 27245–27256. [Google Scholar] [CrossRef] [Green Version]
- Tomić, A.; Tomić, S. Hunting the human DPP III active conformation: Combined thermodynamic and QM/MM calculations. Dalt. Trans. 2014, 43, 15503–15514. [Google Scholar] [CrossRef]
- Sung, H.; Kang, S.K.; Bae, Y.J.; Hong, J.T.; Chung, Y.B.; Lee, C.K.; Song, S. PCR-based detection of Mycoplasma species. J. Microbiol. 2006, 44, 42–49. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT | P479S | E480Q | T481M | G482C | Q484H | R620C | R623W | R623L | R638L | R638W | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| kcat (s−1) | 70.2 ± 3.6 | 53.9 ± 1.5 | 52.3 ± 1.6 | 26.5 ± 0.9 | 62.5 ± 1.0 | 45.1 ± 1.8 | 63.0 ± 1.6 | 41.1 ± 0.6 | 55.7 ± 1.8 | 15.6 ± 0.4 | 8.6 ± 0.3 |
| Km (µM) | 12.7 ± 1.8 | 8.3 ± 0.8 | 10.4 ± 1.0 | 10.0 ± 1.1 | 8.8 ± 0.4 | 6.6 ± 1.0 | 11.2 ± 0.8 | 7.6 ± 0.4 | 9.8 ± 1. 0 | 3.5 ± 0.42 | 3.5 ± 0.5 |
| kcat/Km (s−1/µM) | 5.5 | 6.5 | 5.0 | 2.6 | 7.1 | 6.8 | 5.6 | 5.4 | 5.7 | 4.5 | 2.5 |
| DPP III | Kd(WT)/Kd(mut) a |
|---|---|
| WT | 1.0 |
| E451K | 2.1 |
| P479S | 18.4 |
| E480Q | 0.1 |
| T481M | 0.1 |
| G482C | 0.8 |
| Q484H | 2.1 |
| R510W | 0.3 |
| R623W | 160.0 b |
| R638L | 2.0 |
| R638W | 2.0 |
| R703C | 1.7 |
| DPP III-Variant | Simulations | Time/ns | MM-GBSA-Min (kcal/mol) |
|---|---|---|---|
| WT | 1 | 300 | −51 |
| 2 | 200 | −80 | |
| 3 | 200 | −73 | |
| total | 700 | −80 | |
| P479S | 1 | 300 | −93 |
| 2 | 200 | −116 | |
| 3 | 200 | −85 | |
| total | 700 | −116 | |
| E480Q | 1 | 300 | −42 |
| 2 | 200 | −36 | |
| 3 | 200 | −42 | |
| total | 700 | −42 | |
| T481M | 1 | 300 | −63 |
| 2 | 200 | −52 | |
| 3 | 200 | −63 | |
| total | 700 | −63 | |
| G482C | 1 | 300 | −55 |
| 2 | 200 | −56 | |
| 3 | 200 | −45 | |
| total | 700 | −56 | |
| R510W | 1 | 300 | −69 |
| 2 | 200 | −64 | |
| 3 | 200 | −49 | |
| total | 700 | −69 | |
| R623W | 1 | 300 | −51 |
| 2 | 200 | −50 | |
| 3 | 200 | −65 | |
| total | 700 | −65 |
| DPP III Variant | Total H-Bonds |
|---|---|
| WT | 14.7 |
| P479S | 20.6 |
| E480Q | 8.8 |
| T481M | 13.1 |
| G482C | 10.4 |
| R510W | 13.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matić, S.; Tomašić Paić, A.; Sobočanec, S.; Pinterić, M.; Pipalović, G.; Martinčić, M.; Matovina, M.; Tomić, S. Interdisciplinary Study of the Effects of Dipeptidyl-Peptidase III Cancer Mutations on the KEAP1-NRF2 Signaling Pathway. Int. J. Mol. Sci. 2022, 23, 1994. https://doi.org/10.3390/ijms23041994
Matić S, Tomašić Paić A, Sobočanec S, Pinterić M, Pipalović G, Martinčić M, Matovina M, Tomić S. Interdisciplinary Study of the Effects of Dipeptidyl-Peptidase III Cancer Mutations on the KEAP1-NRF2 Signaling Pathway. International Journal of Molecular Sciences. 2022; 23(4):1994. https://doi.org/10.3390/ijms23041994
Chicago/Turabian StyleMatić, Sara, Ana Tomašić Paić, Sandra Sobočanec, Marija Pinterić, Goran Pipalović, Monika Martinčić, Mihaela Matovina, and Sanja Tomić. 2022. "Interdisciplinary Study of the Effects of Dipeptidyl-Peptidase III Cancer Mutations on the KEAP1-NRF2 Signaling Pathway" International Journal of Molecular Sciences 23, no. 4: 1994. https://doi.org/10.3390/ijms23041994
APA StyleMatić, S., Tomašić Paić, A., Sobočanec, S., Pinterić, M., Pipalović, G., Martinčić, M., Matovina, M., & Tomić, S. (2022). Interdisciplinary Study of the Effects of Dipeptidyl-Peptidase III Cancer Mutations on the KEAP1-NRF2 Signaling Pathway. International Journal of Molecular Sciences, 23(4), 1994. https://doi.org/10.3390/ijms23041994