

Stressor-Induced Increases in Circulating, but Not Colonic, Cytokines Are Related to Anxiety-like Behavior and Hippocampal Inflammation in a Murine Colitis Model

Abstract
:1. Introduction
2. Results
2.1. Stressor Exposure Increases Anxiety-like Behavior in Mice Given DSS despite the Lack of Stressor Effects on the Colonic Disease-Activity Index
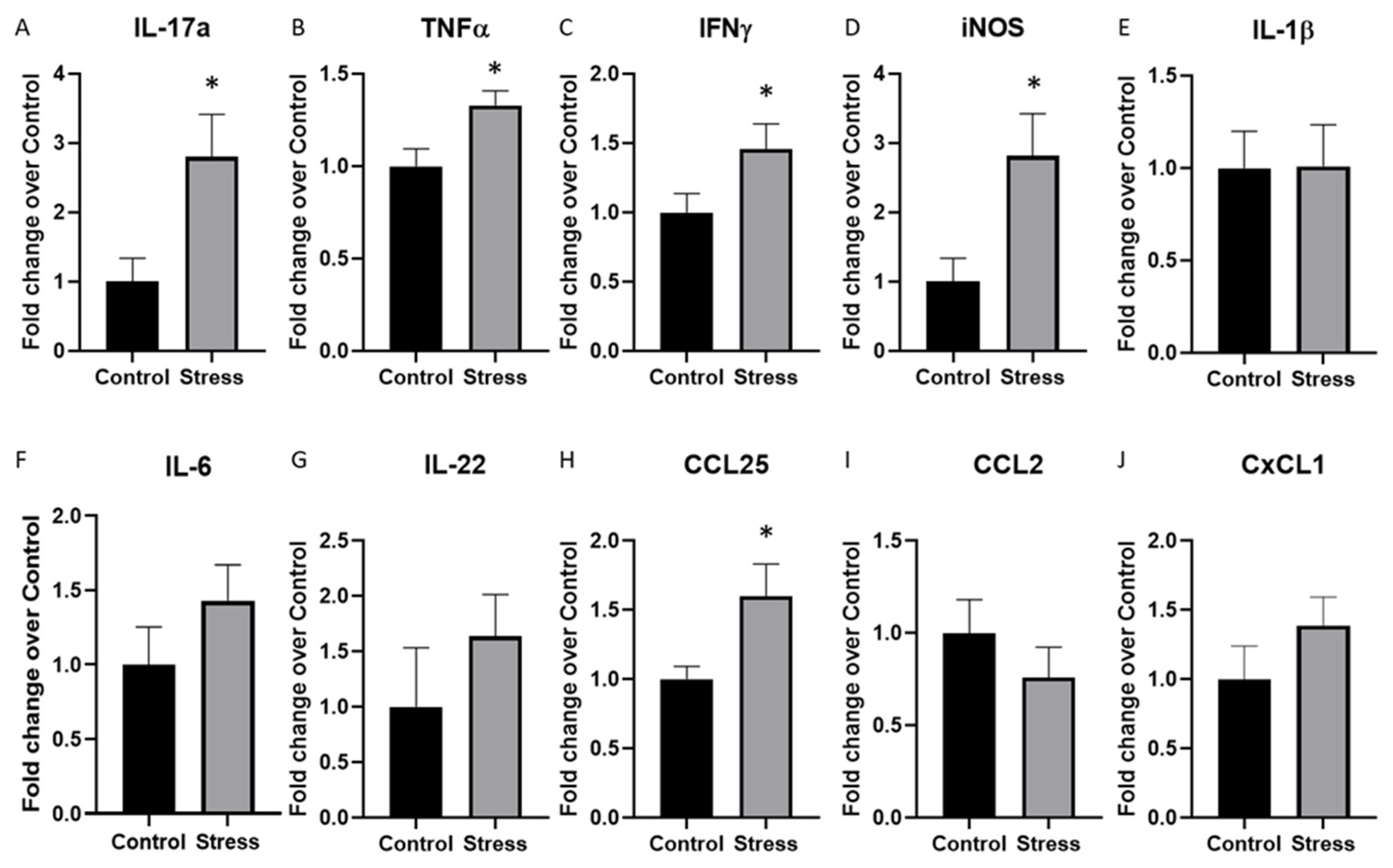
2.2. Cytokine Expression in the Hippocampus Was Significantly Altered by Stressor Exposure in Mice Given DSS
2.3. Serum Cytokine Levels Were Significantly Affected by Stressor Exposure in Mice Given DSS
2.4. Stressor-Exposed Mice Given DSS Have Significant Increases in Cytokines in the Mesenteric Lymph Nodes but Few Differences in the Colon
2.5. Neuroinflammation Is Highly Interrelated to Colonic Immune Activation
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Design
4.3. Behavioral Tests
4.3.1. Light/Dark Exploration
4.3.2. Open Field Exploration
4.4. Histopathology
4.5. Semiquantitative Real-Time PCR
4.6. Serum Measurement of Cytokines
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sartor, R.B. Mechanisms of disease: Pathogenesis of Crohn’s disease and ulcerative colitis. Nat. Clin. Pract. Gastroenterol. Hepatol. 2006, 3, 390–407. [Google Scholar] [CrossRef] [PubMed]
- Bamias, G.; Kaltsa, G.; Ladas, S.D. Cytokines in the pathogenesis of ulcerative colitis. Discov. Med. 2011, 11, 459–467. [Google Scholar] [PubMed]
- Olsen, T.; Rismo, R.; Cui, G.; Goll, R.; Christiansen, I.; Florholmen, J. TH1 and TH17 interactions in untreated inflamed mucosa of inflammatory bowel disease, and their potential to mediate the inflammation. Cytokine 2011, 56, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Shih, D.Q.; Targan, S.R. Immunopathogenesis of inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 390–400. [Google Scholar] [CrossRef]
- Fujino, S.; Andoh, A.; Bamba, S.; Ogawa, A.; Hata, K.; Araki, Y.; Bamba, T.; Fujiyama, Y. Increased expression of interleukin 17 in inflammatory bowel disease. Gut 2003, 52, 65–70. [Google Scholar] [CrossRef]
- Sartor, R.B. Genetics and environmental interactions shape the intestinal microbiome to promote inflammatory bowel disease versus mucosal homeostasis. Gastroenterology 2010, 139, 1816–1819. [Google Scholar] [CrossRef]
- Sartor, R.B. Key questions to guide a better understanding of host-commensal microbiota interactions in intestinal inflammation. Mucosal Immunol. 2011, 4, 127–132. [Google Scholar] [CrossRef]
- Bernstein, C.N.; Singh, S.; Graff, L.A.; Walker, J.R.; Miller, N.; Cheang, M. A prospective population-based study of triggers of symptomatic flares in IBD. Am. J. Gastroenterol. 2010, 105, 1994–2002. [Google Scholar] [CrossRef]
- Bitton, A.; Dobkin, P.L.; Edwardes, M.D.; Sewitch, M.J.; Meddings, J.B.; Rawal, S.; Cohen, A.; Vermeire, S.; Dufresne, L.; Franchimont, D.; et al. Predicting relapse in Crohn’s disease: A biopsychosocial model. Gut 2008, 57, 1386–1392. [Google Scholar] [CrossRef]
- Sexton, K.A.; Walker, J.R.; Graff, L.A.; Bernstein, M.T.; Beatie, B.; Miller, N.; Sargent, M.; Targownik, L.E. Evidence of Bidirectional Associations Between Perceived Stress and Symptom Activity: A Prospective Longitudinal Investigation in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2017, 23, 473–483. [Google Scholar] [CrossRef]
- Maltz, R.M.; Keirsey, J.; Kim, S.C.; Mackos, A.R.; Gharaibeh, R.Z.; Moore, C.C.; Xu, J.; Bakthavatchalu, V.; Somogyi, A.; Bailey, M.T. Prolonged restraint stressor exposure in outbred CD-1 mice impacts microbiota, colonic inflammation, and short chain fatty acids. PLoS ONE 2018, 13, e0196961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maltz, R.M.; Keirsey, J.; Kim, S.C.; Mackos, A.R.; Gharaibeh, R.Z.; Moore, C.C.; Xu, J.; Somogyi, A.; Bailey, M.T. Social Stress Affects Colonic Inflammation, the Gut Microbiome, and Short-chain Fatty Acid Levels and Receptors. J. Pediatric Gastroenterol. Nutr. 2019, 68, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Egger, B.; Bajaj-Elliott, M.; MacDonald, T.T.; Inglin, R.; Eysselein, V.E.; Buchler, M.W. Characterisation of acute murine dextran sodium sulphate colitis: Cytokine profile and dose dependency. Digestion 2000, 62, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Melgar, S.; Karlsson, A.; Michaelsson, E. Acute colitis induced by dextran sulfate sodium progresses to chronicity in C57BL/6 but not in BALB/c mice: Correlation between symptoms and inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G1328–G1338. [Google Scholar] [CrossRef]
- Reinoso Webb, C.; den Bakker, H.; Koboziev, I.; Jones-Hall, Y.; Rao Kottapalli, K.; Ostanin, D.; Furr, K.L.; Mu, Q.; Luo, X.M.; Grisham, M.B. Differential Susceptibility to T Cell-Induced Colitis in Mice: Role of the Intestinal Microbiota. Inflamm. Bowel Dis. 2018, 24, 361–379. [Google Scholar] [CrossRef]
- Rojas-Cartagena, C.; Flores, I.; Appleyard, C.B. Role of tumor necrosis factor receptors in an animal model of acute colitis. Cytokine 2005, 32, 85–93. [Google Scholar] [CrossRef]
- Reber, S.O.; Obermeier, F.; Straub, R.H.; Falk, W.; Neumann, I.D. Chronic intermittent psychosocial stress (social defeat/overcrowding) in mice increases the severity of an acute DSS-induced colitis and impairs regeneration. Endocrinology 2006, 147, 4968–4976. [Google Scholar] [CrossRef] [Green Version]
- Veenema, A.H.; Reber, S.O.; Selch, S.; Obermeier, F.; Neumann, I.D. Early life stress enhances the vulnerability to chronic psychosocial stress and experimental colitis in adult mice. Endocrinology 2008, 149, 2727–2736. [Google Scholar] [CrossRef] [Green Version]
- Ng, Q.X.; Soh, A.Y.S.; Loke, W.; Lim, D.Y.; Yeo, W.S. The role of inflammation in irritable bowel syndrome (IBS). J. Inflamm. Res. 2018, 11, 345–349. [Google Scholar] [CrossRef] [Green Version]
- Greenley, R.N.; Hommel, K.A.; Nebel, J.; Raboin, T.; Li, S.H.; Simpson, P.; Mackner, L. A meta-analytic review of the psychosocial adjustment of youth with inflammatory bowel disease. J. Pediatr. Psychol. 2010, 35, 857–869. [Google Scholar] [CrossRef]
- Mackner, L.M.; Greenley, R.N.; Szigethy, E.; Herzer, M.; Deer, K.; Hommel, K.A. Psychosocial issues in pediatric inflammatory bowel disease: Report of the North American Society for Pediatric Gastroenterology, Hepatology, and Nutrition. J. Pediatric Gastroenterol. Nutr. 2013, 56, 449–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Li, L.; Xie, R.; Wang, B.; Jiang, K.; Cao, H. Stress Triggers Flare of Inflammatory Bowel Disease in Children and Adults. Front. Pediatr. 2019, 7, 432. [Google Scholar] [CrossRef] [PubMed]
- Leff Gelman, P.; Mancilla-Herrera, I.; Flores-Ramos, M.; Saravia Takashima, M.F.; Cruz Coronel, F.M.; Cruz Fuentes, C.; Perez Molina, A.; Hernandez-Ruiz, J.; Silva-Aguilera, F.S.; Farfan-Labonne, B.; et al. The cytokine profile of women with severe anxiety and depression during pregnancy. BMC Psychiatry 2019, 19, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ho, R.C.; Mak, A. The role of interleukin (IL)-17 in anxiety and depression of patients with rheumatoid arthritis. Int. J. Rheum. Dis. 2012, 15, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Wohleb, E.S.; Hanke, M.L.; Corona, A.W.; Powell, N.D.; Stiner, L.M.; Bailey, M.T.; Nelson, R.J.; Godbout, J.P.; Sheridan, J.F. Beta-Adrenergic receptor antagonism prevents anxiety-like behavior and microglial reactivity induced by repeated social defeat. J. Neurosci. 2011, 31, 6277–6288. [Google Scholar] [CrossRef] [Green Version]
- Niraula, A.; Witcher, K.G.; Sheridan, J.F.; Godbout, J.P. Interleukin-6 Induced by Social Stress Promotes a Unique Transcriptional Signature in the Monocytes That Facilitate Anxiety. Biol. Psychiatry 2019, 85, 679–689. [Google Scholar] [CrossRef]
- Leppkes, M.; Becker, C.; Ivanov, I.I.; Hirth, S.; Wirtz, S.; Neufert, C.; Pouly, S.; Murphy, A.J.; Valenzuela, D.M.; Yancopoulos, G.D.; et al. RORgamma-expressing Th17 cells induce murine chronic intestinal inflammation via redundant effects of IL-17A and IL-17F. Gastroenterology 2009, 136, 257–267. [Google Scholar] [CrossRef]
- Zhang, Z.; Zheng, M.; Bindas, J.; Schwarzenberger, P.; Kolls, J.K. Critical role of IL-17 receptor signaling in acute TNBS-induced colitis. Inflamm. Bowel Dis. 2006, 12, 382–388. [Google Scholar] [CrossRef]
- Alves de Lima, K.; Rustenhoven, J.; Da Mesquita, S.; Wall, M.; Salvador, A.F.; Smirnov, I.; Martelossi Cebinelli, G.; Mamuladze, T.; Baker, W.; Papadopoulos, Z.; et al. Meningeal gammadelta T cells regulate anxiety-like behavior via IL-17a signaling in neurons. Nat. Immunol. 2020, 21, 1421–1429. [Google Scholar] [CrossRef]
- McHugh, S.B.; Deacon, R.M.; Rawlins, J.N.; Bannerman, D.M. Amygdala and ventral hippocampus contribute differentially to mechanisms of fear and anxiety. Behav. Neurosci. 2004, 118, 63–78. [Google Scholar] [CrossRef]
- Lu, Q.; Xiang, H.; Zhu, H.; Chen, Y.; Lu, X.; Huang, C. Intranasal lipopolysaccharide administration prevents chronic stress-induced depression- and anxiety-like behaviors in mice. Neuropharmacology 2021, 200, 108816. [Google Scholar] [CrossRef] [PubMed]
- Tarr, A.J.; Powell, N.D.; Reader, B.F.; Bhave, N.S.; Roloson, A.L.; Carson, W.E., 3rd; Sheridan, J.F. beta-Adrenergic receptor mediated increases in activation and function of natural killer cells following repeated social disruption. Brain Behav. Immun. 2012, 26, 1226–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuendorf, R.; Harding, A.; Stello, N.; Hanes, D.; Wahbeh, H. Depression and anxiety in patients with Inflammatory Bowel Disease: A systematic review. J. Psychosom. Res. 2016, 87, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Adam, R.; Colombel, J.F.; Bian, Z.X. A characterization of pro-inflammatory cytokines in dextran sulfate sodium-induced chronic relapsing colitis mice model. Int. Immunopharmacol. 2018, 60, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.I.; Ressler, K.J.; Binder, E.; Nemeroff, C.B. The neurobiology of anxiety disorders: Brain imaging, genetics, and psychoneuroendocrinology. Psychiatr. Clin. N. Am. 2009, 32, 549–575. [Google Scholar] [CrossRef] [Green Version]
- Kolbinger, F.; Huppertz, C.; Mir, A.; Padova, F.D. IL-17A and Multiple Sclerosis: Signaling Pathways, Producing Cells and Target Cells in the Central Nervous System. Curr. Drug Targets 2016, 17, 1882–1893. [Google Scholar] [CrossRef]
- Kostic, M.; Dzopalic, T.; Zivanovic, S.; Zivkovic, N.; Cvetanovic, A.; Stojanovic, I.; Vojinovic, S.; Marjanovic, G.; Savic, V.; Colic, M. IL-17 and glutamate excitotoxicity in the pathogenesis of multiple sclerosis. Scand. J. Immunol. 2014, 79, 181–186. [Google Scholar] [CrossRef]
- Kebir, H.; Kreymborg, K.; Ifergan, I.; Dodelet-Devillers, A.; Cayrol, R.; Bernard, M.; Giuliani, F.; Arbour, N.; Becher, B.; Prat, A. Human TH17 lymphocytes promote blood-brain barrier disruption and central nervous system inflammation. Nat. Med. 2007, 13, 1173–1175. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.M.; Jiang, G.X.; Li, Q.W.; Zhou, Z.M.; Cheng, Q. Increased serum levels of interleukin-18, -23 and -17 in Chinese patients with Alzheimer’s disease. Dement. Geriatr. Cogn. Disord. 2014, 38, 321–329. [Google Scholar] [CrossRef]
- Behairi, N.; Belkhelfa, M.; Mesbah-Amroun, H.; Rafa, H.; Belarbi, S.; Tazir, M.; Touil-Boukoffa, C. All-trans-retinoic acid modulates nitric oxide and interleukin-17A production by peripheral blood mononuclear cells from patients with Alzheimer’s disease. Neuroimmunomodulation 2015, 22, 385–393. [Google Scholar] [CrossRef]
- Liu, Z.; Huang, Y.; Cao, B.B.; Qiu, Y.H.; Peng, Y.P. Th17 Cells Induce Dopaminergic Neuronal Death via LFA-1/ICAM-1 Interaction in a Mouse Model of Parkinson’s Disease. Mol. Neurobiol. 2017, 54, 7762–7776. [Google Scholar] [CrossRef] [PubMed]
- Rocha, N.P.; Assis, F.; Scalzo, P.L.; Vieira, E.L.M.; Barbosa, I.G.; de Souza, M.S.; Christo, P.P.; Reis, H.J.; Teixeira, A.L. Reduced Activated T Lymphocytes (CD4+CD25+) and Plasma Levels of Cytokines in Parkinson’s Disease. Mol. Neurobiol. 2018, 55, 1488–1497. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Qiu, A.W.; Huang, Y.; Yang, Y.; Chen, J.N.; Gu, T.T.; Cao, B.B.; Qiu, Y.H.; Peng, Y.P. IL-17A exacerbates neuroinflammation and neurodegeneration by activating microglia in rodent models of Parkinson’s disease. Brain Behav. Immun. 2019, 81, 630–645. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Mao, X.; Zhou, T.; Cheng, X.; Lin, Y. IL-17A contributes to brain ischemia reperfusion injury through calpain-TRPC6 pathway in mice. Neuroscience 2014, 274, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Chou, Y.C.; Chen, H.C.; Lu, C.C.; Chang, D.M. Interleukin-6 and interleukin-17 are related to depression in patients with rheumatoid arthritis. Int. J. Rheum. Dis. 2019, 22, 980–985. [Google Scholar] [CrossRef]
- Kim, J.; Suh, Y.H.; Chang, K.A. Interleukin-17 induced by cumulative mild stress promoted depression-like behaviors in young adult mice. Mol. Brain 2021, 14, 11. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, N.; Mo, Z.M.; Lin, S.P.; Zhang, F. IL-17A Enhances Microglial Response to OGD by Regulating p53 and PI3K/Akt Pathways with Involvement of ROS/HMGB1. Front. Mol. Neurosci. 2017, 10, 271. [Google Scholar] [CrossRef] [Green Version]
- Wohleb, E.S. Neuron-Microglia Interactions in Mental Health Disorders: “For Better, and For Worse”. Front. Immunol. 2016, 7, 544. [Google Scholar] [CrossRef] [Green Version]
- Wohleb, E.S.; McKim, D.B.; Sheridan, J.F.; Godbout, J.P. Monocyte trafficking to the brain with stress and inflammation: A novel axis of immune-to-brain communication that influences mood and behavior. Front. Neurosci. 2014, 8, 447. [Google Scholar] [CrossRef]
- Wohleb, E.S.; McKim, D.B.; Shea, D.T.; Powell, N.D.; Tarr, A.J.; Sheridan, J.F.; Godbout, J.P. Re-establishment of anxiety in stress-sensitized mice is caused by monocyte trafficking from the spleen to the brain. Biol. Psychiatry 2014, 75, 970–981. [Google Scholar] [CrossRef] [Green Version]
- Cua, D.J.; Tato, C.M. Innate IL-17-producing cells: The sentinels of the immune system. Nat. Rev. Immunol. 2010, 10, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.T.; Dowd, S.E.; Parry, N.M.; Galley, J.D.; Schauer, D.B.; Lyte, M. Stressor exposure disrupts commensal microbial populations in the intestines and leads to increased colonization by Citrobacter rodentium. Infect. Immun. 2010, 78, 1509–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galley, J.D.; Mackos, A.R.; Varaljay, V.A.; Bailey, M.T. Stressor exposure has prolonged effects on colonic microbial community structure in Citrobacter rodentium-challenged mice. Sci. Rep. 2017, 7, 45012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.G.; Kamada, N.; Shaw, M.H.; Warner, N.; Chen, G.Y.; Franchi, L.; Nunez, G. The Nod2 sensor promotes intestinal pathogen eradication via the chemokine CCL2-dependent recruitment of inflammatory monocytes. Immunity 2011, 34, 769–780. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.S.; Yang, H.; Yang, J.Y.; Kim, Y.; Lee, S.H.; Kim, J.H.; Jang, Y.J.; Vallance, B.A.; Kweon, M.N. Interleukin-1 (IL-1) signaling in intestinal stromal cells controls KC/ CXCL1 secretion, which correlates with recruitment of IL-22- secreting neutrophils at early stages of Citrobacter rodentium infection. Infect. Immun. 2015, 83, 3257–3267. [Google Scholar] [CrossRef] [Green Version]
- Roberts, C.J.; Morgan, G.R.; Danford, N. Effect of hormones on the variation of radiosensitivity in females as measured by induction of chromosomal aberrations. Environ. Health Perspect. 1997, 105 (Suppl. S6), 1467–1471. [Google Scholar] [CrossRef]
- Moochhala, S.; Chhatwal, V.J.; Chan, S.T.; Ngoi, S.S.; Chia, Y.W.; Rauff, A. Nitric oxide synthase activity and expression in human colorectal cancer. Carcinogenesis 1996, 17, 1171–1174. [Google Scholar] [CrossRef] [Green Version]
- Beckman, J.S.; Beckman, T.W.; Chen, J.; Marshall, P.A.; Freeman, B.A. Apparent hydroxyl radical production by peroxynitrite: Implications for endothelial injury from nitric oxide and superoxide. Proc. Natl. Acad. Sci. USA 1990, 87, 1620–1624. [Google Scholar] [CrossRef] [Green Version]
- Marletta, M.A.; Yoon, P.S.; Iyengar, R.; Leaf, C.D.; Wishnok, J.S. Macrophage oxidation of L-arginine to nitrite and nitrate: Nitric oxide is an intermediate. Biochemistry 1988, 27, 8706–8711. [Google Scholar] [CrossRef]
- Witthoft, T.; Eckmann, L.; Kim, J.M.; Kagnoff, M.F. Enteroinvasive bacteria directly activate expression of iNOS and NO production in human colon epithelial cells. Am. J. Physiol. 1998, 275, G564–G571. [Google Scholar] [CrossRef]
- Kolios, G.; Rooney, N.; Murphy, C.T.; Robertson, D.A.; Westwick, J. Expression of inducible nitric oxide synthase activity in human colon epithelial cells: Modulation by T lymphocyte derived cytokines. Gut 1998, 43, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.C.; Vazquez-Torres, A.; Xu, Y. The transcriptional regulator SoxS is required for resistance of Salmonella typhimurium to paraquat but not for virulence in mice. Infect. Immun. 1997, 65, 5371–5375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, G.; Victor Fon, G.; Meixner, W.; Creekmore, A.; Zong, Y.; Dame, M.K.; Colacino, J.; Dedhia, P.H.; Hong, S.; Wiley, J.W. Chronic stress and intestinal barrier dysfunction: Glucocorticoid receptor and transcription repressor HES1 regulate tight junction protein Claudin-1 promoter. Sci. Rep. 2017, 7, 4502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, J.M.; Jaggers, R.M.; Solden, L.M.; Loman, B.R.; Davies, R.H.; Mackos, A.R.; Ladaika, C.A.; Berg, B.M.; Chichlowski, M.; Bailey, M.T. Dietary Oligosaccharides Attenuate Stress-Induced Disruptions in Immune Reactivity and Microbial B-Vitamin Metabolism. Front. Immunol. 2019, 10, 1774. [Google Scholar] [CrossRef] [Green Version]
- Bailey, M.T.; Dowd, S.E.; Galley, J.D.; Hufnagle, A.R.; Allen, R.G.; Lyte, M. Exposure to a social stressor alters the structure of the intestinal microbiota: Implications for stressor-induced immunomodulation. Brain Behav. Immun. 2011, 25, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Bailey, M.T.; Engler, H.; Sheridan, J.F. Stress induces the translocation of cutaneous and gastrointestinal microflora to secondary lymphoid organs of C57BL/6 mice. J. Neuroimmunol. 2006, 171, 29–37. [Google Scholar] [CrossRef]
- Sakuraba, A.; Sato, T.; Kamada, N.; Kitazume, M.; Sugita, A.; Hibi, T. Th1/Th17 immune response is induced by mesenteric lymph node dendritic cells in Crohn’s disease. Gastroenterology 2009, 137, 1736–1745. [Google Scholar] [CrossRef]
- Alshammari, M.A.; Khan, M.R.; Majid Mahmood, H.; Alshehri, A.O.; Alasmari, F.F.; Alqahtani, F.M.; Alasmari, A.F.; Alsharari, S.D.; Alhossan, A.; Ahmad, S.F.; et al. Systemic TNF-alpha blockade attenuates anxiety and depressive-like behaviors in db/db mice through downregulation of inflammatory signaling in peripheral immune cells. Saudi Pharm. J. 2020, 28, 621–629. [Google Scholar] [CrossRef]
- Bailey, M.T.; Kinsey, S.G.; Padgett, D.A.; Sheridan, J.F.; Leblebicioglu, B. Social stress enhances IL-1beta and TNF-alpha production by Porphyromonas gingivalis lipopolysaccharide-stimulated CD11b+ cells. Physiol. Behav. 2009, 98, 351–358. [Google Scholar] [CrossRef] [Green Version]
- Chandrashekara, S.; Jayashree, K.; Veeranna, H.B.; Vadiraj, H.S.; Ramesh, M.N.; Shobha, A.; Sarvanan, Y.; Vikram, Y.K. Effects of anxiety on TNF-alpha levels during psychological stress. J. Psychosom. Res. 2007, 63, 65–69. [Google Scholar] [CrossRef]
- Hou, R.; Garner, M.; Holmes, C.; Osmond, C.; Teeling, J.; Lau, L.; Baldwin, D.S. Peripheral inflammatory cytokines and immune balance in Generalised Anxiety Disorder: Case-controlled study. Brain Behav. Immun. 2017, 62, 212–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camara, R.J.; Ziegler, R.; Begre, S.; Schoepfer, A.M.; von Kanel, R.; Swiss Inflammatory Bowel Disease Cohort Study. The role of psychological stress in inflammatory bowel disease: Quality assessment of methods of 18 prospective studies and suggestions for future research. Digestion 2009, 80, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Mardini, H.E.; Kip, K.E.; Wilson, J.W. Crohn’s disease: A two-year prospective study of the association between psychological distress and disease activity. Dig. Dis. Sci. 2004, 49, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Traue, H.C.; Kosarz, P. Everyday stress and Crohn’s disease activity: A time series analysis of 20 single cases. Int. J. Behav. Med. 1999, 6, 101–119. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, H.; Neurath, M.F.; Atreya, R. Role of the IL23/IL17 Pathway in Crohn’s Disease. Front. Immunol. 2021, 12, 622934. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.T.; Kierstein, S.; Sharma, S.; Spaits, M.; Kinsey, S.G.; Tliba, O.; Amrani, Y.; Sheridan, J.F.; Panettieri, R.A.; Haczku, A. Social stress enhances allergen-induced airway inflammation in mice and inhibits corticosteroid responsiveness of cytokine production. J. Immunol. 2009, 182, 7888–7896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackos, A.R.; Galley, J.D.; Eubank, T.D.; Easterling, R.S.; Parry, N.M.; Fox, J.G.; Lyte, M.; Bailey, M.T. Social stress-enhanced severity of Citrobacter rodentium-induced colitis is CCL2-dependent and attenuated by probiotic Lactobacillus reuteri. Mucosal Immunol. 2015, 9, 515–526. [Google Scholar] [CrossRef] [Green Version]
- Rath, H.C.; Herfarth, H.H.; Ikeda, J.S.; Grenther, W.B.; Hamm, T.E., Jr.; Balish, E.; Taurog, J.D.; Hammer, R.E.; Wilson, K.H.; Sartor, R.B. Normal luminal bacteria, especially Bacteroides species, mediate chronic colitis, gastritis, and arthritis in HLA-B27/human beta2 microglobulin transgenic rats. J. Clin. Investig. 1996, 98, 945–953. [Google Scholar] [CrossRef] [Green Version]
- Oldak, B.; Cruz-Rivera, M.; Flisser, A.; Mendlovic, F. RNA Purity, Real-Time PCR Sensitivity, and Colon Segment Influence mRNA Relative Expression in Murine Dextran Sodium Sulfate Experimental Colitis. J. Biomol. Tech. 2018, 29, 61–70. [Google Scholar] [CrossRef]
- Head, C.C.; Farrow, M.J.; Sheridan, J.F.; Padgett, D.A. Androstenediol reduces the anti-inflammatory effects of restraint stress during wound healing. Brain Behav. Immun. 2006, 20, 590–596. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Stress | p Value | |
|---|---|---|---|
| IL-17a | 1.0 + 1.36 | 0.52 + 1.6 | 0.66 |
| TNFα | 1.0 ± 0.13 | 0.9 ± 0.21 | 0.51 |
| IL-22 | 1.0 ± 0.91 | 0.59 ± 0.47 | 0.52 |
| IL-6 | 1.0 ± 0.14 | 0.9 ± 0.36 | 0.67 |
| BDNF | 1.0 ± 0.16 | 1.46 ± 0.47 | 0.27 |
| CCL25 | 1.0 ± 0.59 | 0.88 ± 0.55 | 0.817 |
| CCL2 | 1.0 ± 0.77 | 1.49 ± 0.38 | 0.52 |
| CxCL1 | 1.0 ± 0.61 | 0.75 ± 0.66 | 0.65 |
| Control | Stress | p Value | |
|---|---|---|---|
| IL-17A | 0.86 ± 0.28 | 0.49 ± 0.17 | 0.29 |
| TNFα | 14.50 ± 5.76 | 10.35 ± 2.16 | 0.51 |
| CCL20 | 27.16 ± 8.24 | 32.71 ± 6.99 | 0.62 |
| IL-21 | 2.70 ± 1.09 | 1.27 ± 0.32 | 0.26 |
| IL-22 | 0.72 ± 0.33 (3) | 0.41 ± 0.44 (2) | 0.54 |
| IL-17E/IL-25 | 0.18 ± 0.05 (3) | 0.10 ± 0.03 (6) | 0.20 |
| IFNγ | 0.86 ± 0.41 (3) | 0.39 ± 0.13 (5) | 0.23 |
| GM-CSF | 0.94 ± 0.47 (3) | 0.06 ± 0.02 (4) | 0.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maltz, R.M.; Marte-Ortiz, P.; Rajasekera, T.A.; Loman, B.R.; Gur, T.L.; Bailey, M.T. Stressor-Induced Increases in Circulating, but Not Colonic, Cytokines Are Related to Anxiety-like Behavior and Hippocampal Inflammation in a Murine Colitis Model. Int. J. Mol. Sci. 2022, 23, 2000. https://doi.org/10.3390/ijms23042000
Maltz RM, Marte-Ortiz P, Rajasekera TA, Loman BR, Gur TL, Bailey MT. Stressor-Induced Increases in Circulating, but Not Colonic, Cytokines Are Related to Anxiety-like Behavior and Hippocampal Inflammation in a Murine Colitis Model. International Journal of Molecular Sciences. 2022; 23(4):2000. https://doi.org/10.3390/ijms23042000
Chicago/Turabian StyleMaltz, Ross M., Pedro Marte-Ortiz, Therese A. Rajasekera, Brett R. Loman, Tamar L. Gur, and Michael T. Bailey. 2022. "Stressor-Induced Increases in Circulating, but Not Colonic, Cytokines Are Related to Anxiety-like Behavior and Hippocampal Inflammation in a Murine Colitis Model" International Journal of Molecular Sciences 23, no. 4: 2000. https://doi.org/10.3390/ijms23042000
APA StyleMaltz, R. M., Marte-Ortiz, P., Rajasekera, T. A., Loman, B. R., Gur, T. L., & Bailey, M. T. (2022). Stressor-Induced Increases in Circulating, but Not Colonic, Cytokines Are Related to Anxiety-like Behavior and Hippocampal Inflammation in a Murine Colitis Model. International Journal of Molecular Sciences, 23(4), 2000. https://doi.org/10.3390/ijms23042000