Author Contributions
Conceptualization, C.D., G.S., F.M., A.U. and A.O.; Methodology, C.D., F.V., D.V.R. and F.M.; formal analysis, F.M., G.A.S. and F.V.; investigation, C.D., F.M., D.V.R., G.S., G.A.S., G.L.R., A.U. and A.O.; data curation, F.M. and C.D.; Writing—original draft preparation, F.M..; Writing—review and editing, C.D., G.S., A.U. and A.O.; supervision, C.D. and G.S. All authors have read and agreed to the published version of the manuscript.
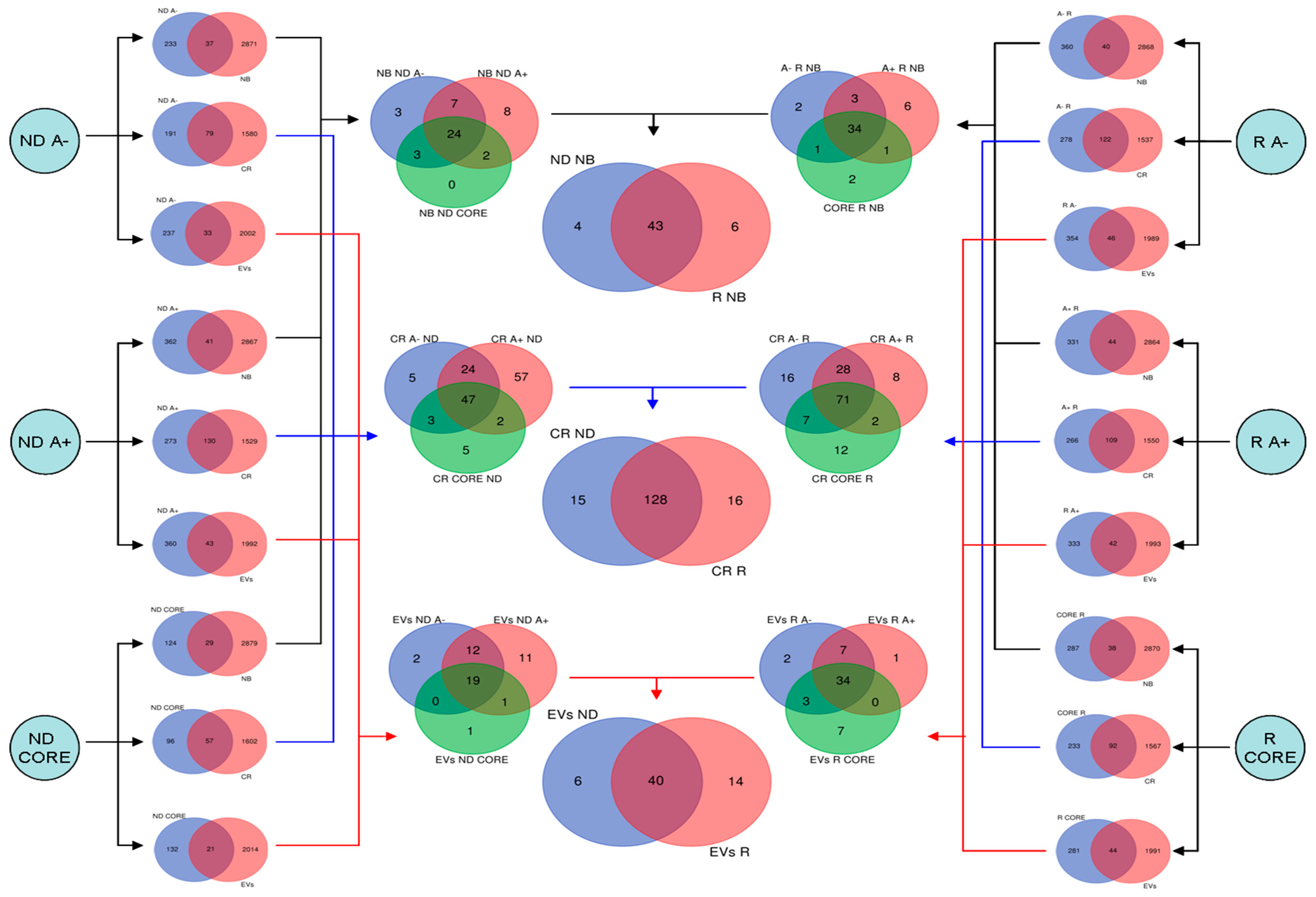
Figure 1.
Workflow applied for the computational study of GBM sub-proteomes based on different tumor zones, i.e., CORE, A+ and A−, and GBM diagnosis, i.e., ND- and R-GBMs.
Figure 1.
Workflow applied for the computational study of GBM sub-proteomes based on different tumor zones, i.e., CORE, A+ and A−, and GBM diagnosis, i.e., ND- and R-GBMs.
Figure 2.
(A) NB proteins found in ND-GBM pool, (B) NB proteins found in R-GBM pool and the relative zone of identifications.
Figure 2.
(A) NB proteins found in ND-GBM pool, (B) NB proteins found in R-GBM pool and the relative zone of identifications.
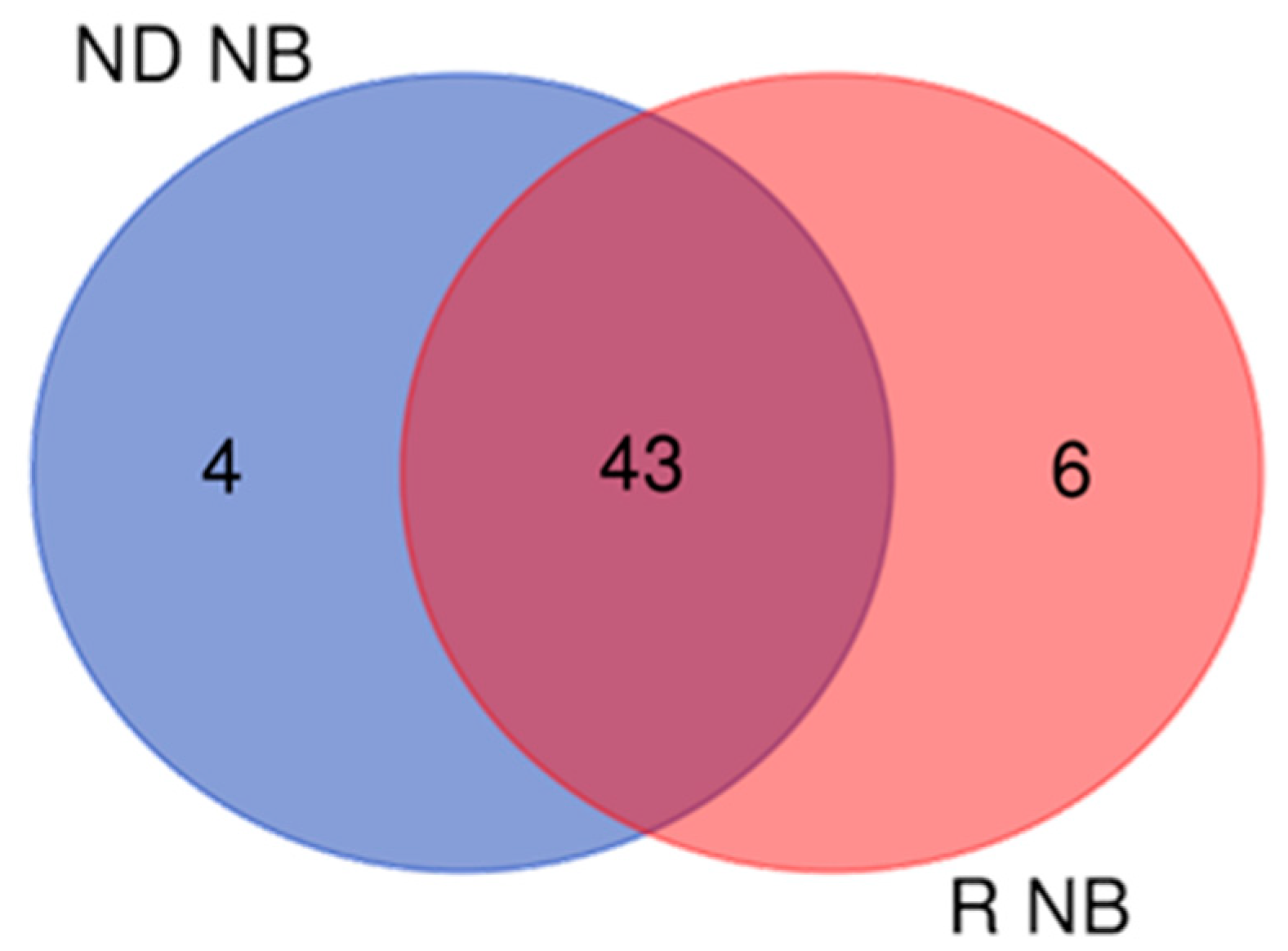
Figure 3.
Venn diagram between Newly Diagnosed (ND) and Recurrent (R) non-brain (NB) proteins: 43 are common to both GBM pools, 4 and 6 are exclusive of ND- and R-GBM, respectively.
Figure 3.
Venn diagram between Newly Diagnosed (ND) and Recurrent (R) non-brain (NB) proteins: 43 are common to both GBM pools, 4 and 6 are exclusive of ND- and R-GBM, respectively.
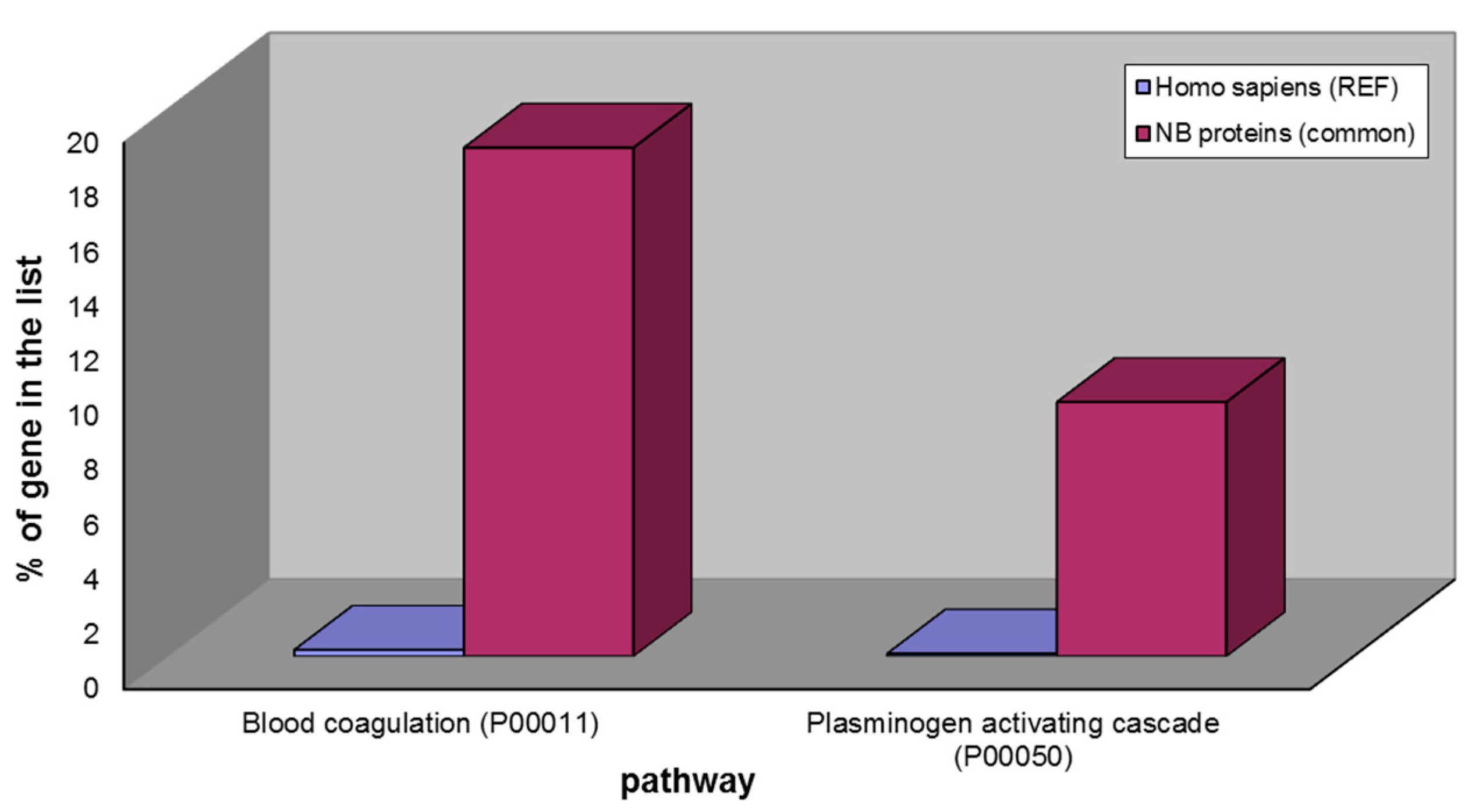
Figure 4.
Over-represented pathways of the 43 NB proteins common to ND- and R-GBM pools: blood coagulation and plasminogen activating cascade.
Figure 4.
Over-represented pathways of the 43 NB proteins common to ND- and R-GBM pools: blood coagulation and plasminogen activating cascade.
Figure 5.
Venn diagram of the 53 NB proteins of the samples and blood proteins of Human Protein Atlas.
Figure 5.
Venn diagram of the 53 NB proteins of the samples and blood proteins of Human Protein Atlas.
Figure 6.
Protein–protein functional interaction network (highest confidence) and clusters of the 43 NB proteins common to Newly Diagnosed and Recurrent GBM pools.
Figure 6.
Protein–protein functional interaction network (highest confidence) and clusters of the 43 NB proteins common to Newly Diagnosed and Recurrent GBM pools.
Figure 7.
Grouping analyses of the CR proteins found in ND-GBM (A) and R-GBM (B) pools and their zone of identification.
Figure 7.
Grouping analyses of the CR proteins found in ND-GBM (A) and R-GBM (B) pools and their zone of identification.
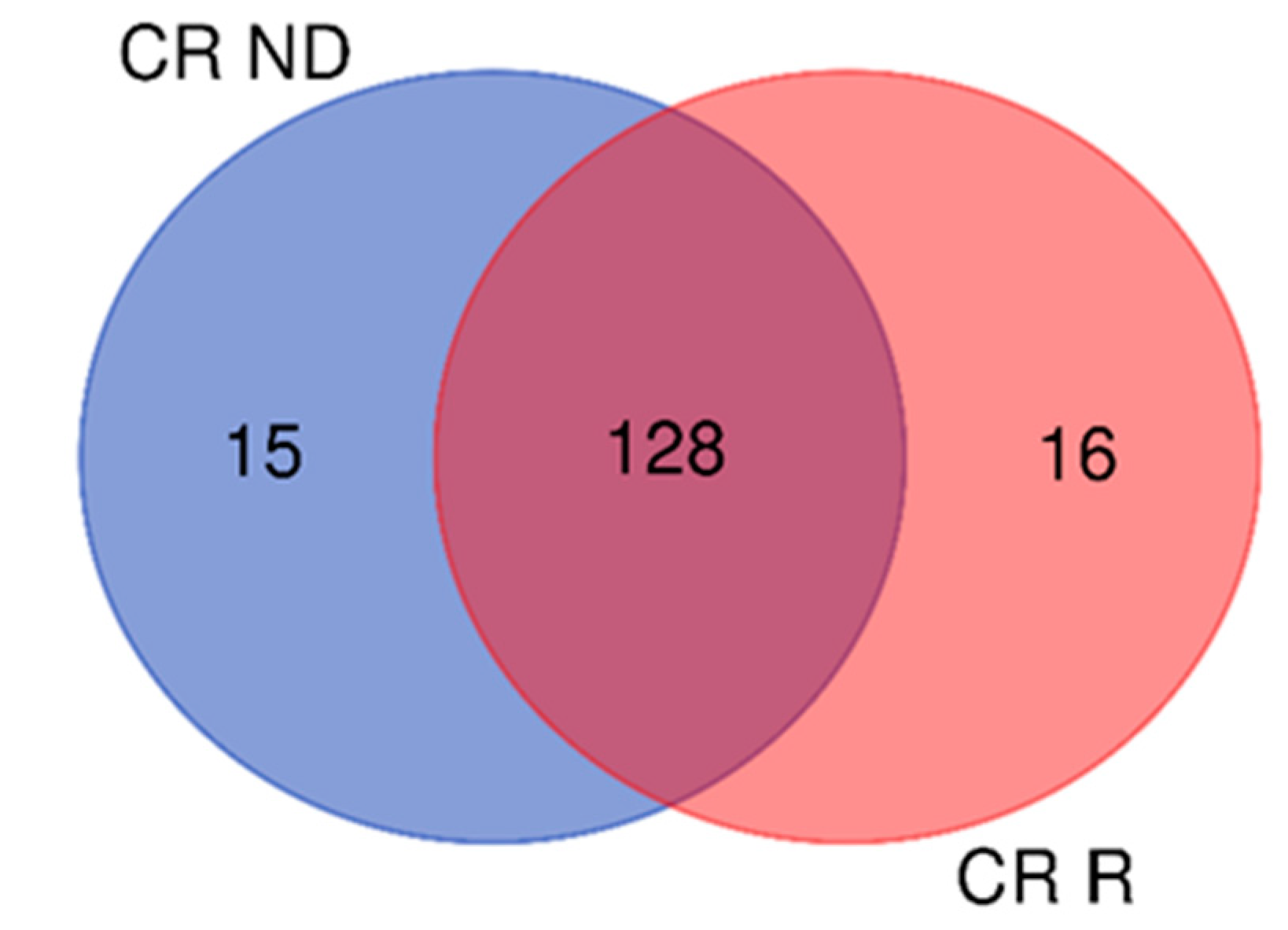
Figure 8.
Venn diagram of CR proteins of the samples: 128 elements were common to both GBM pools, while 15 and 16 were typical of ND- and R-GBMs, respectively.
Figure 8.
Venn diagram of CR proteins of the samples: 128 elements were common to both GBM pools, while 15 and 16 were typical of ND- and R-GBMs, respectively.
Figure 9.
Over-represented pathways of 128 CR proteins common to Newly Diagnosed and Recurrent GBMs. From the left: asparagine and aspartate biosynthesis, blood coagulation, de novo purine biosynthesis, fructose galactose metabolism, glycolysis, Parkinson disease, plasminogen activating cascade, pyruvate metabolism, TCA cycle.
Figure 9.
Over-represented pathways of 128 CR proteins common to Newly Diagnosed and Recurrent GBMs. From the left: asparagine and aspartate biosynthesis, blood coagulation, de novo purine biosynthesis, fructose galactose metabolism, glycolysis, Parkinson disease, plasminogen activating cascade, pyruvate metabolism, TCA cycle.
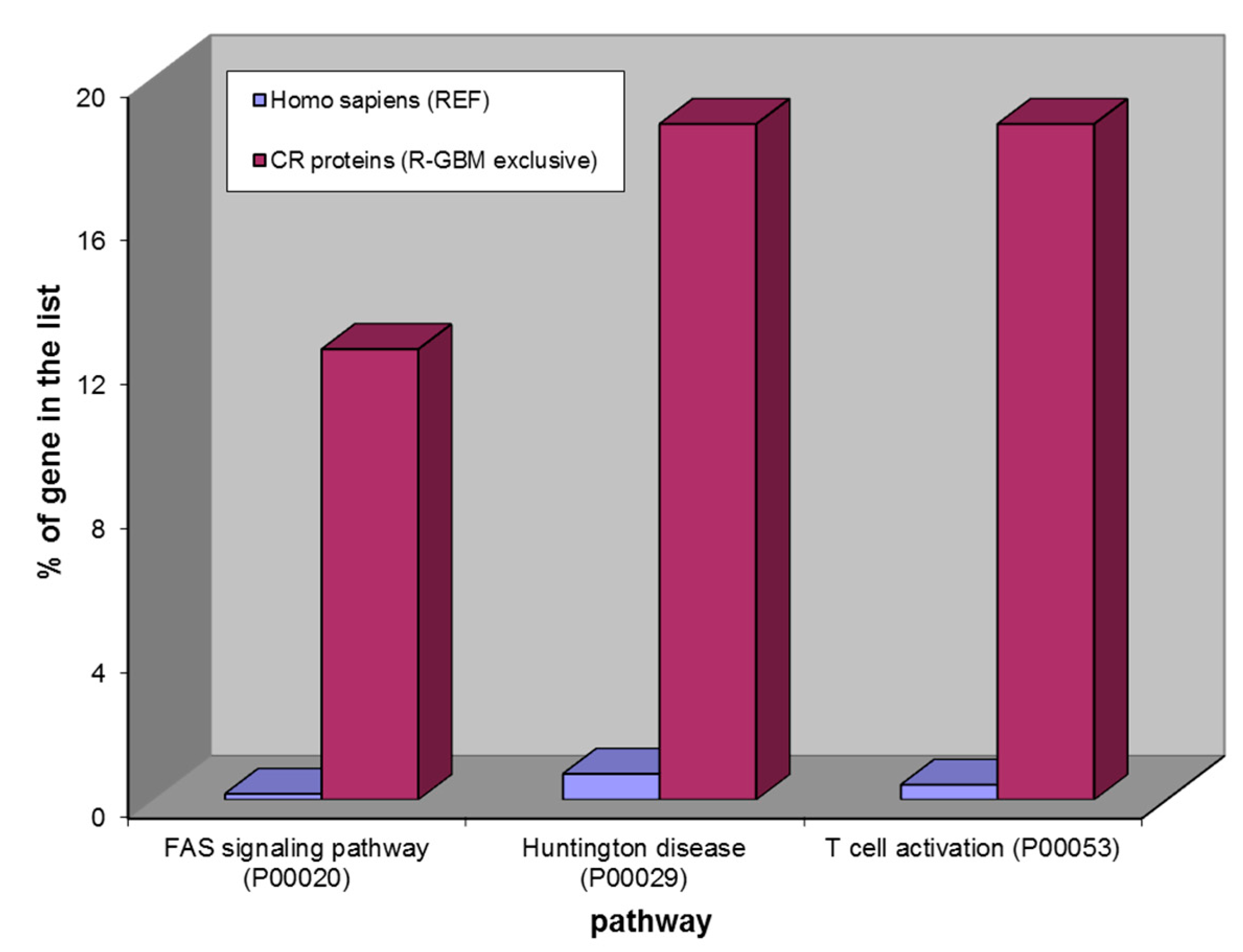
Figure 10.
Over-represented pathways of 16 CR proteins exclusively found in Recurrent GBM. From the left: FAS signaling pathway, Huntington disease and T cell activation.
Figure 10.
Over-represented pathways of 16 CR proteins exclusively found in Recurrent GBM. From the left: FAS signaling pathway, Huntington disease and T cell activation.
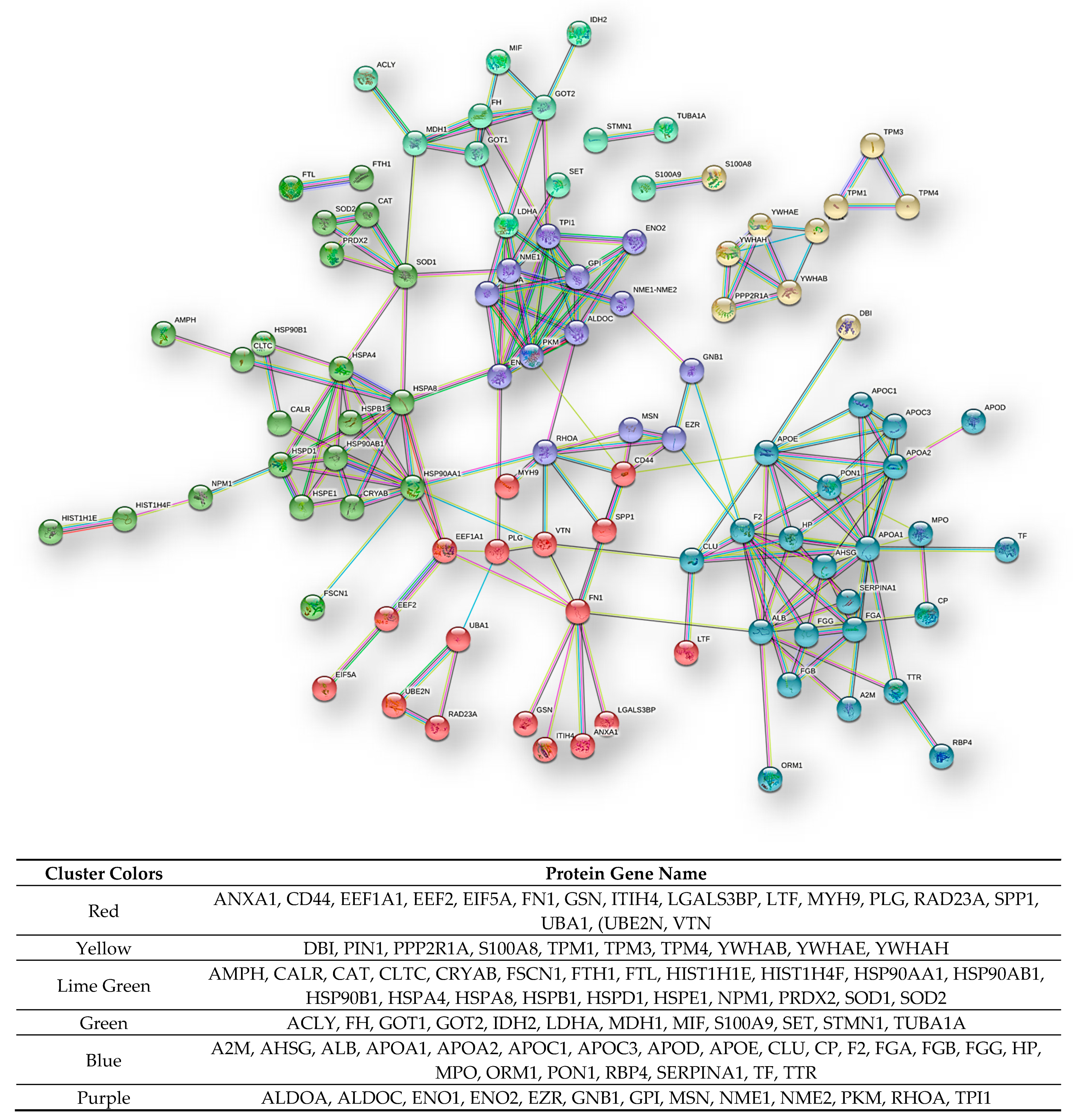
Figure 11.
STRING functional interaction network (highest confidence) and clusters of the 128 CR proteins common to both GBMs pools. At the bottom the list of the gene names of the proteins enclosed in the different clusters is reported.
Figure 11.
STRING functional interaction network (highest confidence) and clusters of the 128 CR proteins common to both GBMs pools. At the bottom the list of the gene names of the proteins enclosed in the different clusters is reported.
Figure 12.
Grouping analyses of the EVs proteins found in ND-GBM (A) and in R-GBM (B) pools and the relative zone of identification.
Figure 12.
Grouping analyses of the EVs proteins found in ND-GBM (A) and in R-GBM (B) pools and the relative zone of identification.
Figure 13.
Venn diagram of EVs proteins of the samples: 40 were present in both ND- and R-GBM pools; 6 only in ND-GBMs and 14 in R-GBMs.
Figure 13.
Venn diagram of EVs proteins of the samples: 40 were present in both ND- and R-GBM pools; 6 only in ND-GBMs and 14 in R-GBMs.
Figure 14.
(A) Results of the PANTHER pathway over-representation analysis of 40 EVs common proteins to ND- and R-GBM pools (glycolysis) and (B) of 14 EVs proteins exclusive to R-GBM pool (Huntington disease).
Figure 14.
(A) Results of the PANTHER pathway over-representation analysis of 40 EVs common proteins to ND- and R-GBM pools (glycolysis) and (B) of 14 EVs proteins exclusive to R-GBM pool (Huntington disease).
Figure 15.
STRING tool analysis (highest confidence) of protein–protein interaction and clusters of the 40 EVs proteins found in Newly Diagnosed and Recurrent GBMs.
Figure 15.
STRING tool analysis (highest confidence) of protein–protein interaction and clusters of the 40 EVs proteins found in Newly Diagnosed and Recurrent GBMs.
Figure 16.
Venn diagram of all proteins identified: 31 were only Extracellular Vesicles proteins, 28 only non-brain proteins and 123 only Cancer Related proteins; 6 were both EVs and NB proteins, 13 both CR and NB proteins, 17 both EVs and CR proteins. Finally, 6 were identified as EVs, NB and CR proteins.
Figure 16.
Venn diagram of all proteins identified: 31 were only Extracellular Vesicles proteins, 28 only non-brain proteins and 123 only Cancer Related proteins; 6 were both EVs and NB proteins, 13 both CR and NB proteins, 17 both EVs and CR proteins. Finally, 6 were identified as EVs, NB and CR proteins.
Figure 17.
STRING analyses of the six elements common to the three sub-proteomes, i.e., NB, CR and EVs proteins with (A) medium confidence and (B) high confidence.
Figure 17.
STRING analyses of the six elements common to the three sub-proteomes, i.e., NB, CR and EVs proteins with (A) medium confidence and (B) high confidence.
Figure 18.
Quantitative trend of the five proteins common to NB, CR and EVs sub-proteome identified in ND-GBM pool, with enlargement to appreciate the trends of VTN and ITIH4, and with p-value expression.
Figure 18.
Quantitative trend of the five proteins common to NB, CR and EVs sub-proteome identified in ND-GBM pool, with enlargement to appreciate the trends of VTN and ITIH4, and with p-value expression.
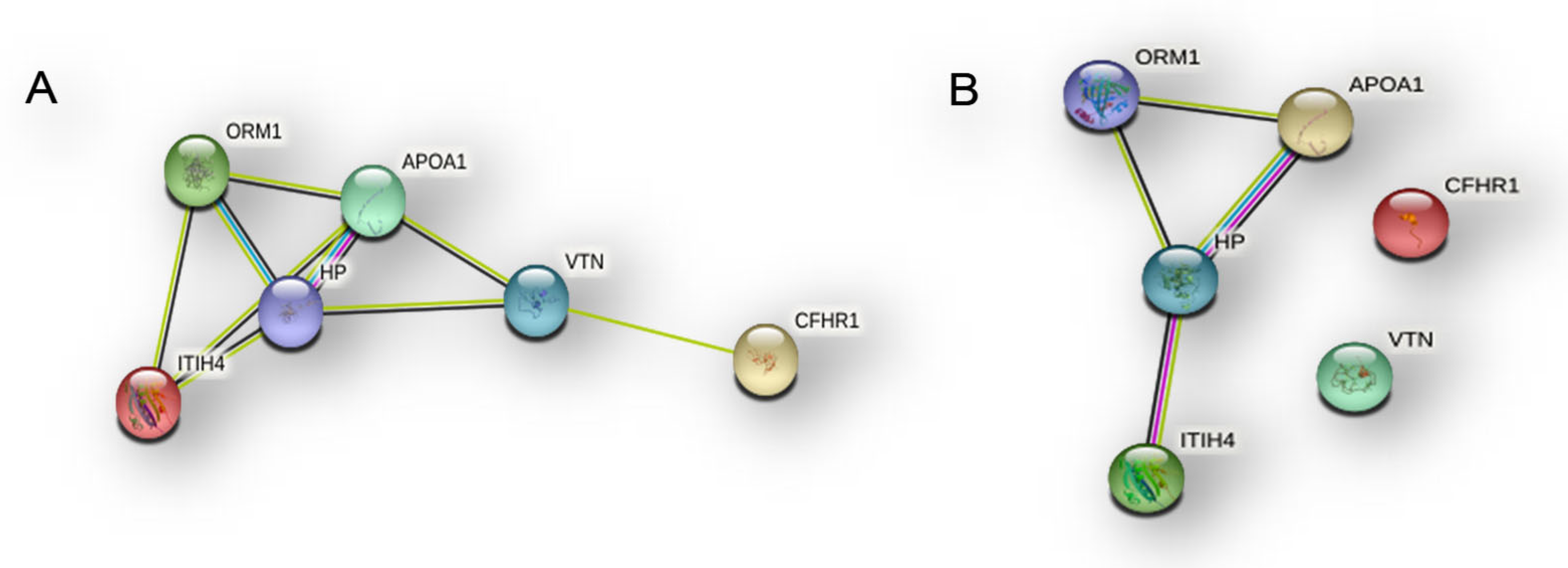
Figure 19.
Quantitative trend of the six proteins common to NB, CR and EVs sub-proteome identified in R-GBM pool, with enlargement to appreciate the trends of ITIH4, VTN and CFHR, and with p-value expression.
Figure 19.
Quantitative trend of the six proteins common to NB, CR and EVs sub-proteome identified in R-GBM pool, with enlargement to appreciate the trends of ITIH4, VTN and CFHR, and with p-value expression.
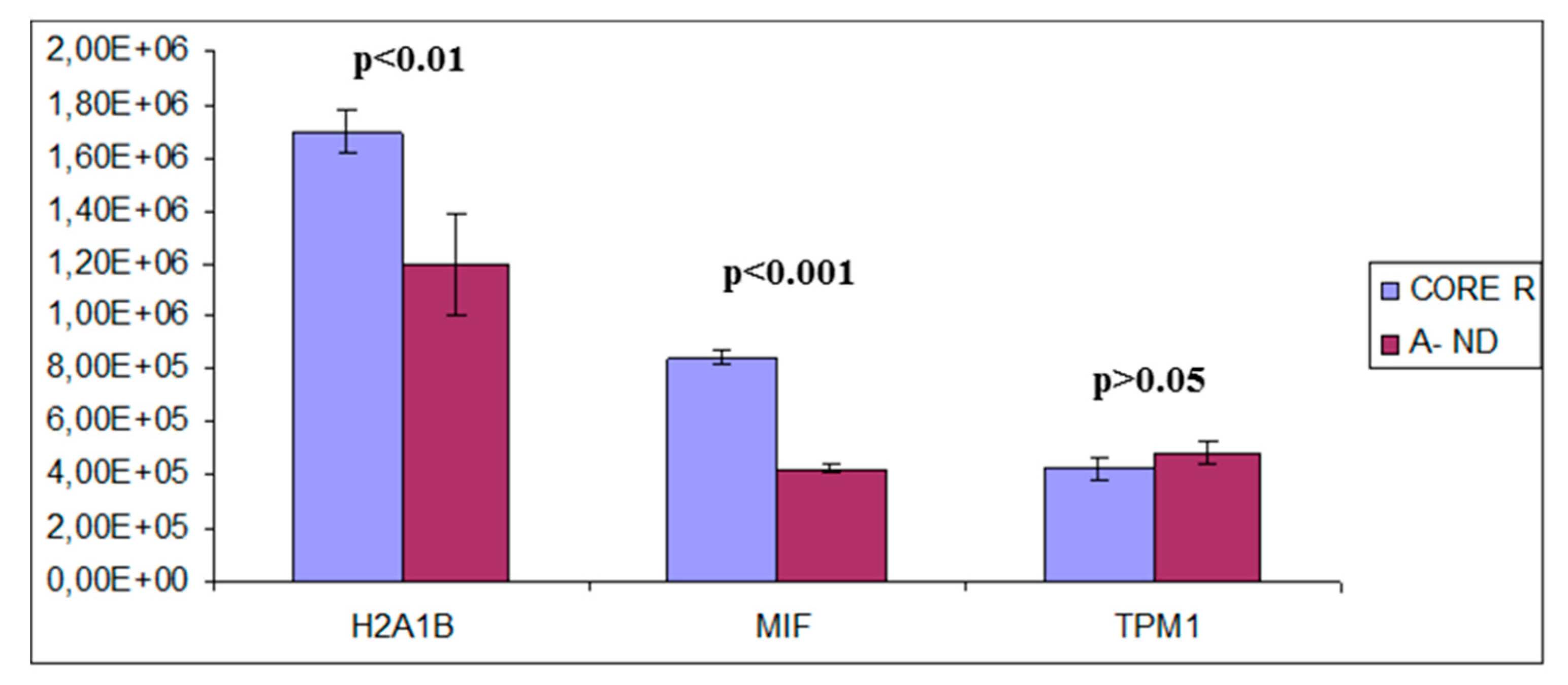
Figure 20.
Quantitative analysis of H2A1B, MIF and TPM1, proteins detected in A− ND-GBM and in CORE R-GBM. The statistical significant differences are additionally reported (p-value, p).
Figure 20.
Quantitative analysis of H2A1B, MIF and TPM1, proteins detected in A− ND-GBM and in CORE R-GBM. The statistical significant differences are additionally reported (p-value, p).
Table 1.
List of 4 NB proteins exclusively identified in ND-GBM pool.
Table 1.
List of 4 NB proteins exclusively identified in ND-GBM pool.
| Uniprot | Protein Name | ND Zones |
|---|
| Q96PD5 | N-acetylmuramoyl-L-alanine amidase | A+ |
| Q562R1 | Beta-actin-like protein 2 | A+ |
| P35527 | Keratin, type I cytoskeletal 9 | A+ |
| P08697 | Alpha-2-antiplasmin | A+ |
Table 2.
List of 6 NB proteins exclusively identified in R-GBM pool.
Table 2.
List of 6 NB proteins exclusively identified in R-GBM pool.
| Uniprot | Protein Name | R Zones |
|---|
| P35542 | Serum amyloid A-4 protein | A+ |
| Q06033 | Inter-alpha-trypsin inhibitor heavy chain H3 | A+ |
| P0DJI8 | Serum amyloid A-1 protein | A+ CORE |
| Q8WZ42 | Titin | A+ |
| Q03591 | Complement factor H-related protein 1 | A+ CORE A− |
| P02748 | Complement component C9 | A+ |
Table 3.
Description of the 5 NB non-blood proteins found in Newly Diagnosed and Recurrent GBMs pools.
Table 3.
Description of the 5 NB non-blood proteins found in Newly Diagnosed and Recurrent GBMs pools.
| Uniprot | Protein Name | ND Zones | R Zones |
|---|
| P23527 | Histone H2B type 1-O | CORE A− | CORE |
| Q96KK5 | Histone H2A type 1-H | CORE A+ A− | CORE A+ A− |
| P62805 | Histone H4 | A+ | A+ |
| P04908 | Histone H2A type 1-B/E | A− | CORE |
| P0DJI8 | Serum amyloid A-1 protein | | A+ CORE |
Table 4.
List of the 15 CR proteins exclusively identified in ND-GBM pool.
Table 4.
List of the 15 CR proteins exclusively identified in ND-GBM pool.
| Uniprot | Protein Name | ND Zones |
|---|
| P00533 | Epidermal growth factor receptor | A+ |
| Q15836 | Vesicle-associated membrane protein 3 | A+ |
| P20073 | Annexin A7 | A+ |
| Q15056 | Eukaryotic translation initiation factor 4H | A+ |
| Q15365 | Poly(rC)-binding protein 1 | A+ |
| P13987 | CD59 glycoprotein | CORE |
| P23297 | Protein S100-A1 | A+ |
| P07355 | Annexin A2 | A+ |
| P36222 | Chitinase-3-like protein 1 | A+ |
| P07951 | Tropomyosin beta chain | CORE |
| P26447 | Protein S100-A4 | A+ |
| P17931 | Galectin-3 | A+ |
| P27824 | Calnexin | A+ |
| P54727 | UV excision repair protein RAD23 homolog B | A+ |
| P13010 | X-ray repair cross-complementing protein 5 | A+ |
Table 5.
List of the 16 CR proteins only detected in R-GBM pool.
Table 5.
List of the 16 CR proteins only detected in R-GBM pool.
| Uniprot | Protein Name | R Zones |
|---|
| P31751 | RAC-beta serine/threonine-protein kinase | A− |
| Q00577 | Transcriptional activator protein Pur-alpha | A− |
| P02545 | Prelamin-A/C | CORE A− |
| P38159 | RNA-binding motif protein, X chromosome | CORE |
| P67809 | Nuclease-sensitive element-binding protein 1 | CORE |
| Q99961 | Endophilin-A2 | CORE |
| P19367 | Hexokinase-1 | A− |
| Q02952 | A-kinase anchor protein 12 | CORE A− |
| P54652 | Heat shock-related 70 kDa protein 2 | A+ A− |
| Q15843 | NEDD8 | A+ A− |
| Q03591 | Complement factor H-related protein 1 | CORE A+ A− |
| P63000 | Ras-related C3 botulinum toxin substrate 1 | CORE A− |
| P84103 | Serine/arginine-rich splicing factor 3 | CORE |
| P68402 | Platelet-activating factor acetylhydrolase IB subunit beta | CORE A+ A− |
| P63167 | Dynein light chain 1, cytoplasmic | A+ A− |
| P61769 | Beta-2-microglobulin | A− |
Table 6.
List of the 6 EVs proteins exclusive to Newly Diagnosed GBM pool.
Table 6.
List of the 6 EVs proteins exclusive to Newly Diagnosed GBM pool.
| Uniprot | Protein Name | ND Zones |
|---|
| P36222 | Chitinase-3-like protein 1 | A+ |
| Q9GZV4 | Eukaryotic translation initiation factor 5A-2 | A− |
| P27105 | Erythrocyte band 7 integral membrane protein | A+ A− |
| P13987 | CD59 glycoprotein | CORE |
| Q9H4G4 | Golgi-associated plant pathogenesis-related protein 1 | A+ |
| Q15181 | Inorganic pyrophosphatase | A+ |
Table 7.
List of 14 EVs proteins typical of Recurrent GBM pool.
Table 7.
List of 14 EVs proteins typical of Recurrent GBM pool.
| Uniprot | Protein Name | R Zones |
|---|
| P31751 | RAC-beta serine/threonine-protein kinase | A− |
| P67809 | Nuclease-sensitive element-binding protein 1 | CORE |
| P47756 | F-actin-capping protein subunit beta | A− CORE |
| Q6PUV4 | Complexin-2 | CORE |
| Q03591 | Complement factor H-related protein 1 | CORE A+ A− |
| Q9BY11 | Protein kinase C and casein kinase substrate in neurons protein 1 | CORE A+ A− |
| P25398 | 40S ribosomal protein S12 | CORE |
| P09497 | Clathrin light chain B | CORE |
| Q8N6N7 | Acyl-CoA-binding domain-containing protein 7 | CORE |
| P54652 | Heat shock-related 70 kDa protein 2 | A+ A− |
| P00739 | Haptoglobin-related protein | A+ |
| Q9UHG2 | ProSAAS | CORE |
| O14745 | Na(+)/H(+) exchange regulatory cofactor NHE-RF1 | A− CORE |
| Q9NRV9 | Heme-binding protein 1 | A− |
Table 8.
Cancer Related proteins exclusive to ND-GBM and their role in cancer disease.
Table 8.
Cancer Related proteins exclusive to ND-GBM and their role in cancer disease.
| Gene Name | ND Zones | Protein Function and References | GBM Identification |
|---|
| EGFR | A+ | Promote tumor survival, proliferation and invasion [17] | Yes |
| VAMP3 | A+ | Support EGFR in its functions [18] | Yes |
| ANXA7 | A+ | Tumor suppressor, downregulated in cancer [19] | Yes |
| IF4H | A+ | Associated to poor prognosis; promote the helicases activity [20] | Yes |
| PCBP1 | A+ | Inhibitor of tumor onset and metastasis; downregulated in cancers [21] | No |
| CD59 | CORE | Overexpressed; regulate the functions and the infiltration of immune cells in tumor [22] | No |
| S10A1 | A+ | Anti-apoptotic [23] | No |
| ANXA2 | A+ | Over-represented in higher grade GBM; stimulate angiogenesis, proliferation and invasiveness [24] | Yes |
| CH3L1 | A+ | Promoting of tumor growth, proliferation, invasion and metastasis; overexpressed in cancers [25] | Yes |
| TPM2 | CORE | Modulate invasion and migration [26] | No |
| S10A4 | A+ | Overexpressed in cancers; correlated with the occurrence of metastasis and associated to poor prognosis [27] | No |
| LEG3 | A+ | Induce endothelial cell differentiation [28] | Yes |
| CALX | A+ | Inhibits the infiltration and the functions of T cells [29] | No |
| RAD23B | A+ | Involved in genome nucleotide excision repair [30] | No |
| XRCC5 | A+ | Mutagenic factor [31] | No |
Table 9.
Cancer Related proteins exclusive to R-GBM and their role in cancer disease.
Table 9.
Cancer Related proteins exclusive to R-GBM and their role in cancer disease.
| Gene Name | R Zones | Protein Function and References | GBM Identification |
|---|
| AKT2 | A− | Involved in metabolism, cellular growth and survival, angiogenesis [31] | No |
| PURA | A− | Transcriptional activator protein overexpressed in GBM, where interact with tumor associated genes [32] | Yes |
| LMNA | CORE A− | Possible biomarkers of aggressiveness; associated to poor prognosis [33] | Yes |
| RBMX | CORE | All RNA binding proteins are overexpressed in GBM and associated with poor prognosis, but RBMX protein has never been detected [34] | No |
| YBOX1 | CORE | Like LMNA, it is a transcriptional activator protein overexpressed in GBM, where interact with tumor associated genes [32] | Yes |
| SH3G1 | CORE | Involved in oncogenesis; its expression is directly proportional to tumor progression [35] | Yes |
| HXK1 | A− | Increased glycolysis and oncogenesis; its isoform HXK2 has been found in GBM [36] | No |
| AKA12 | CORE A− | Downregulated in GBM; tumor suppressor [37] | Yes |
| HSP72 | A+ A− | Chaperones; inhibits proliferation of T cells [38] | Yes |
| NEDD8 | A+ A− | Inhibits the immune activity; it is a target for therapy under study [39] | Yes |
| CFHR1 | CORE A+ A− | Inhibitor of complement alternative pathway [40] | Yes |
| RAC1 | CORE A− | Promote radio resistance in GBM [41] | Yes |
| SRSF3 | CORE | Upregulated; associated to tumor progression and poor prognosis [42] | Yes |
| PA1B2 | CORE A+ A− | Involved in migration, invasion and metastasis [43] | No |
| DYL1 | A+ A− | Overexpressed; promote proliferation and invasion [44] | Yes |
| B2MG | A− | Correlated with malignancy and immune signatures [45] | Yes |
Table 10.
EVs proteins correlated with GBM and other brain tumors.
Table 10.
EVs proteins correlated with GBM and other brain tumors.
| Uniprot | Protein Name | ND Zones | R Zones |
|---|
| P04406 | Glyceraldehyde-3-phosphate dehydrogenase | CORE A+ A− | CORE A+ A− |
| P07602 | Prosaposin | A+ | CORE A+ A− |
| P36222 | Chitinase-3-like protein 1 | A+ | Not detected |
Table 11.
List of the six common proteins to all three sub-proteomes.
Table 11.
List of the six common proteins to all three sub-proteomes.
| Uniprot | Protein Name | ND Zones | R Zones |
|---|
| P04004 | Vitronectin | CORE A+ A− | CORE A+ A− |
| P02647 | Apolipoprotein A-I | CORE A+ A− | CORE A+ A− |
| P00738 | Haptoglobin | CORE A+ A− | CORE A+ A− |
| Q03591 | Complement factor H-related protein 1 | | CORE A+ A− |
| Q14624 | Inter-alpha-trypsin inhibitor heavy chain H4 | A+ A− | CORE A+ A− |
| P02763 | Alpha-1-acid glycoprotein 1 | CORE A+ A− | CORE A+ A− |
Table 12.
List of protein elements identified in the A− zone of ND-GBM pool and identified in R-GBM and their zone of identification in R-GBM.
Table 12.
List of protein elements identified in the A− zone of ND-GBM pool and identified in R-GBM and their zone of identification in R-GBM.
| Uniprot | Protein Name | R-GBM Zones | Sub-Proteome |
|---|
| P07451 | Carbonic anhydrase 3 | CORE A+ A− | NB |
| P04908 | Histone H2A type 1-B/E | CORE | NB |
| P13671 | Complement component C6 | A+ | NB/CR |
| P14174 | Macrophage migration inhibitory factor | CORE A+ | CR |
| P20810 | Calpastatin | A+ | CR |
| P09493 | Tropomyosin alpha-1 chain | CORE | CR |
| P05109 | Protein S100-A8 | A+ A− | CR |
| P07998 | Ribonuclease pancreatic | CORE A+ A− | EVs |


 ,
, 






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}