
The Unitary Micro-Immunotherapy Medicine Interferon-γ (4 CH) Displays Similar Immunostimulatory and Immunomodulatory Effects than Those of Biologically Active Human Interferon-γ on Various Cell Types




Abstract
:1. Introduction
2. Results
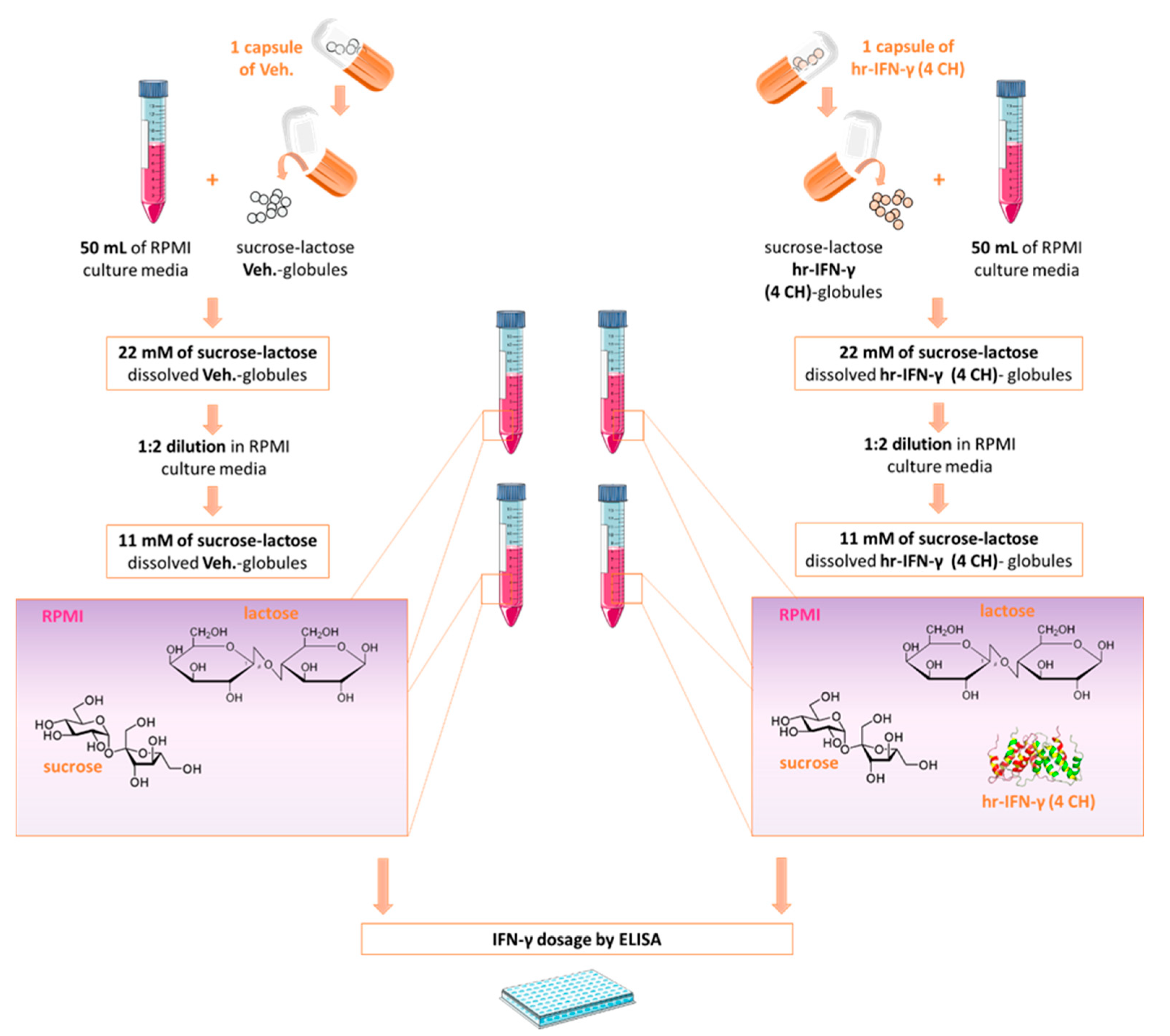
2.1. Few Picograms per mL of Human IFN-γ Are Contained in the Homeopathically Prepared IFN-γ (4 CH)-Impregnated Sucrose-Lactose Pillules
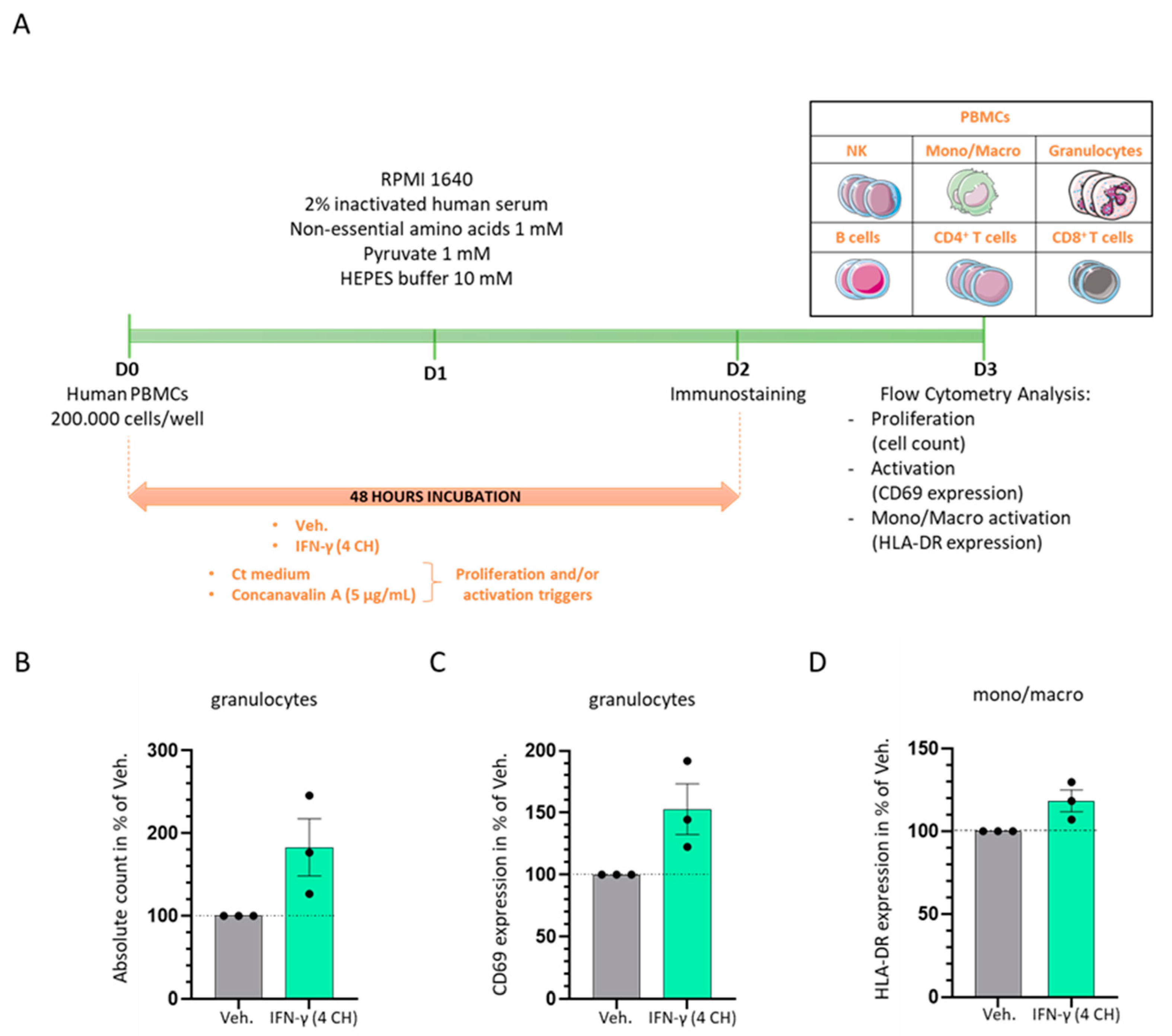
2.2. In Vitro, IFN-γ (4 CH) Increases the Proliferation and the Activation of Granulocytes and It Enhances the Expression of HLA-DR in Monocytes/Macrophages
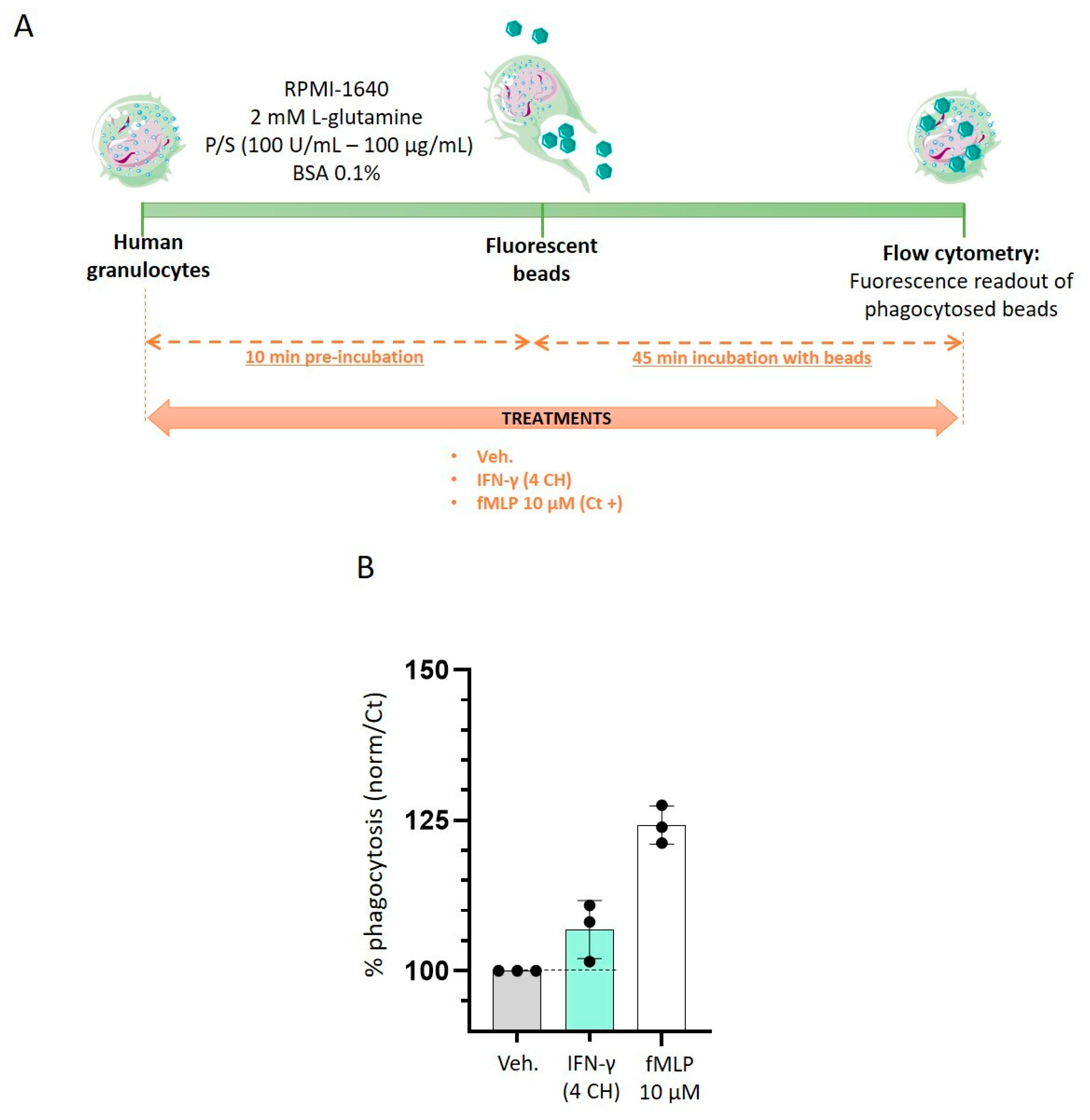
2.3. IFN-γ (4 CH) Stimulates Granulocytes Phagocytosis Capabilities In Vitro
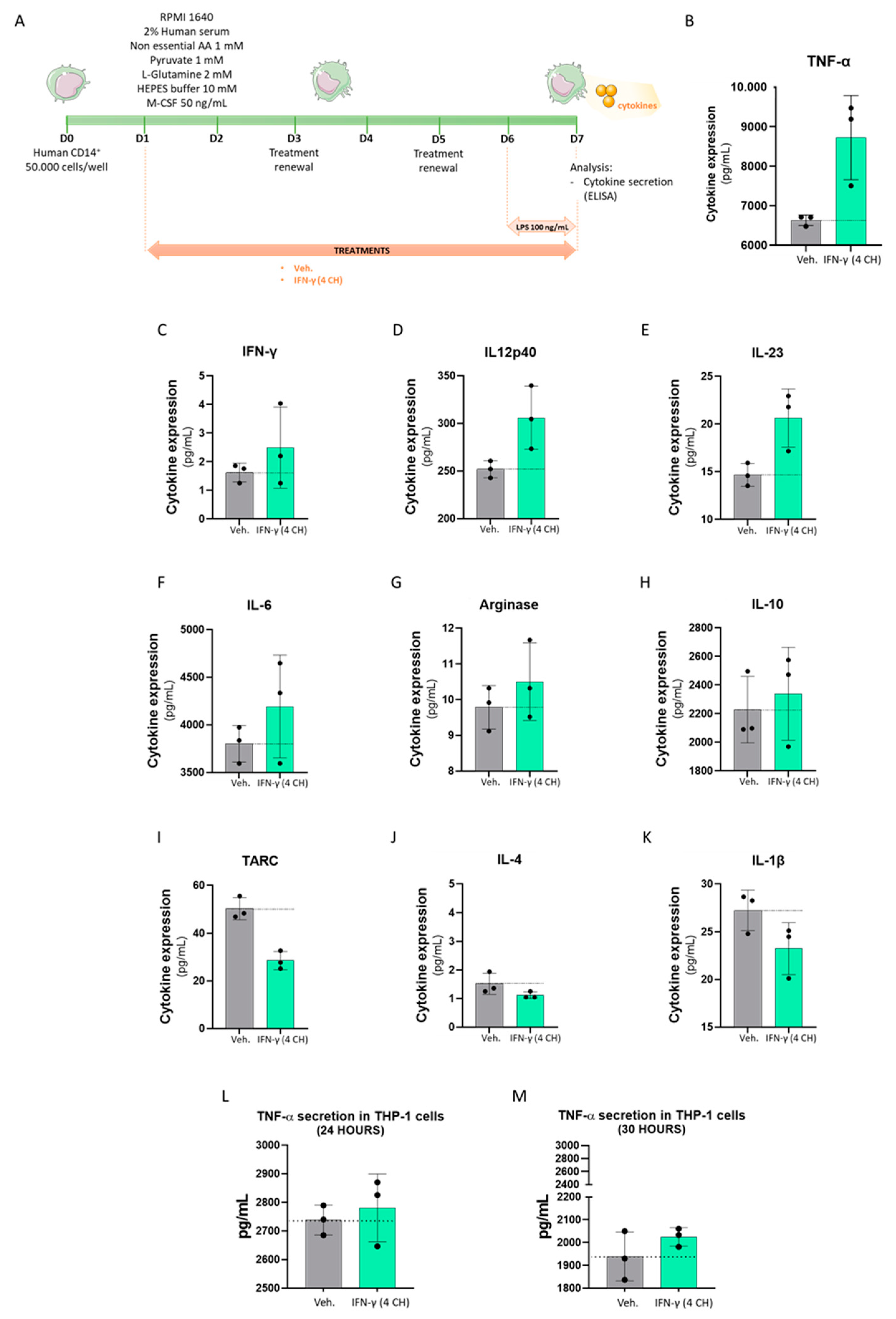
2.4. IFN-γ (4 CH) Increases the TNF-α Secretion in CD14+-Derived Macrophages and THP-1 Cells, and It Modulates the Cytokines’ Secretion Profile in CD14+-Derived Macrophages
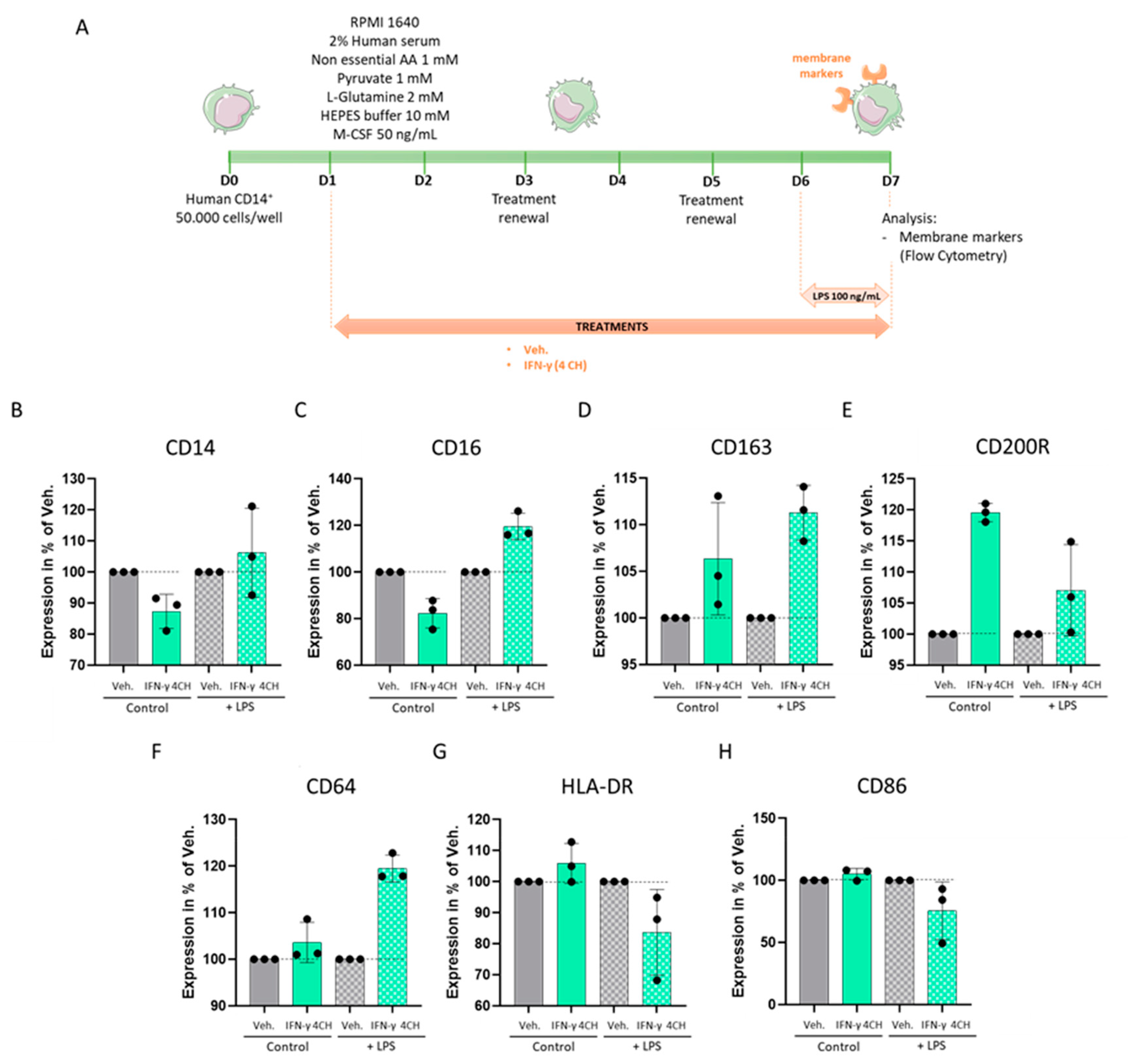
2.5. IFN-γ (4 CH) Modulates the Expression of Membrane Markers in an In Vitro Model of CD14+-Derived Macrophages
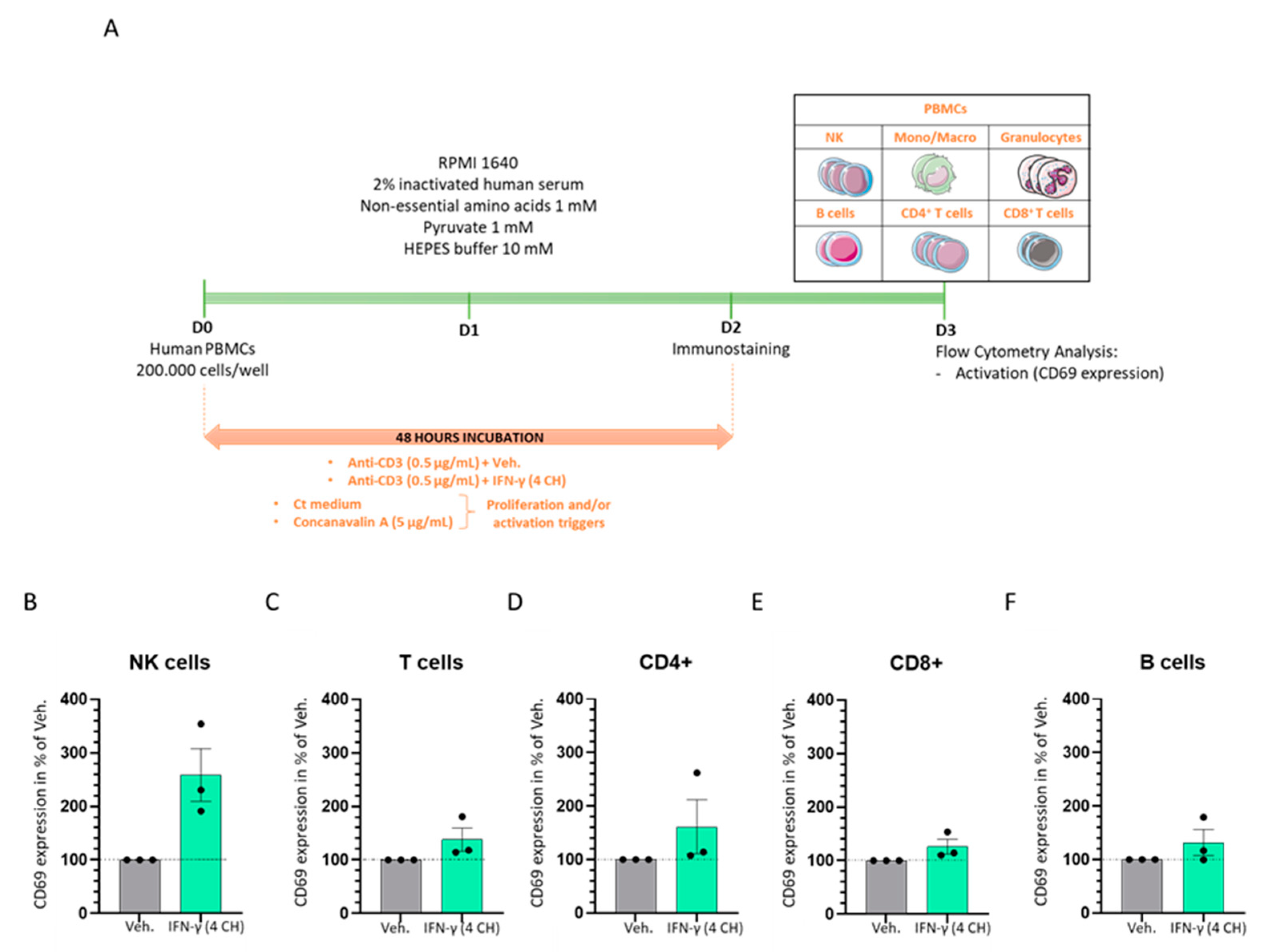
2.6. IFN-γ (4 CH) Enhances the Activation of CD3-Pre-Primed Immune Cells In Vitro
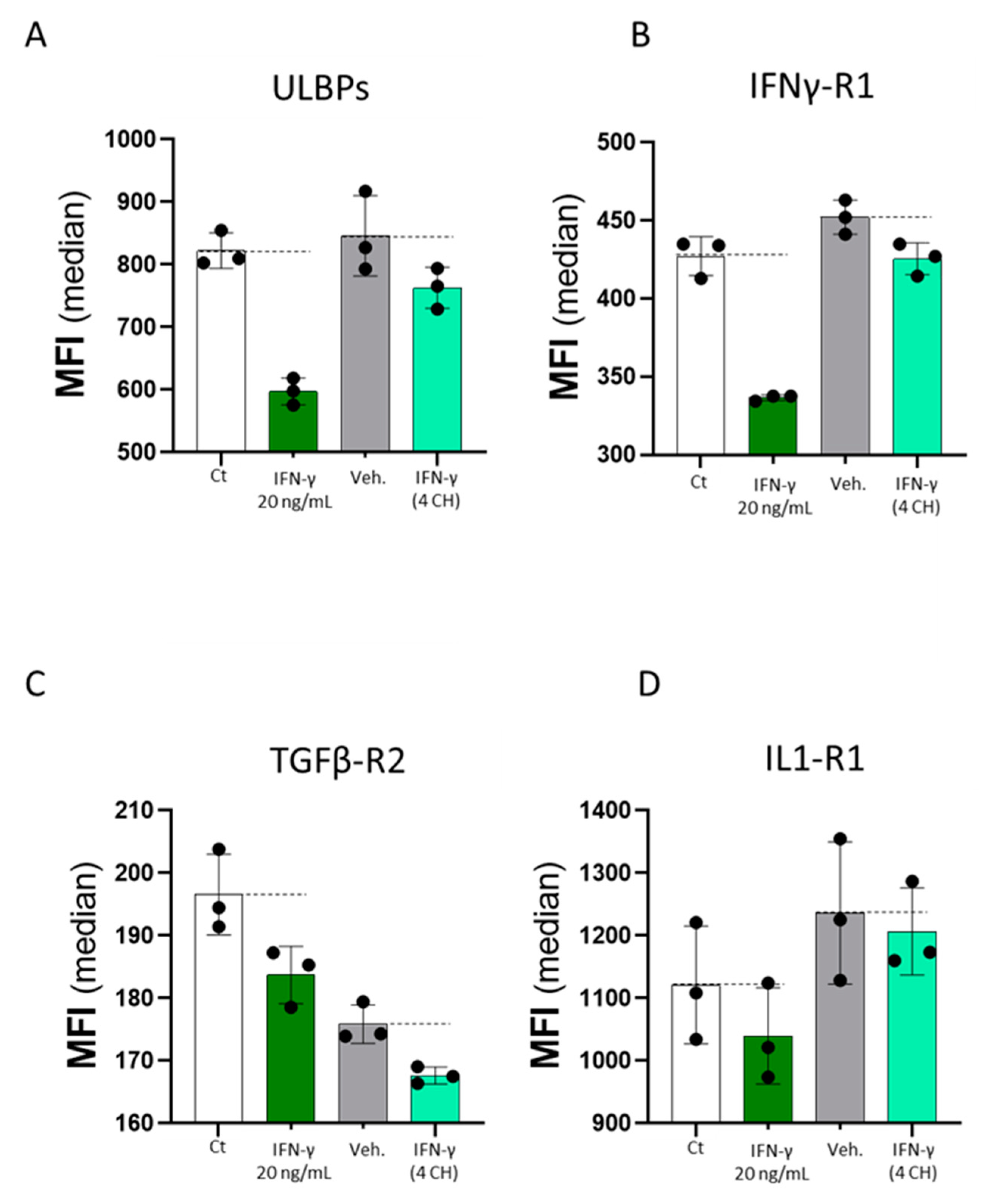
2.7. Commercially Available Human Recombinant IFN-γ and Homeopathically Prepared IFN-γ (4 CH) (Each Compared to Their Own Controls) Have Both DownRegulated the Expression of the Four Tested Endothelial Cell Surface Markers In Vitro
3. Discussion
4. Materials and Methods
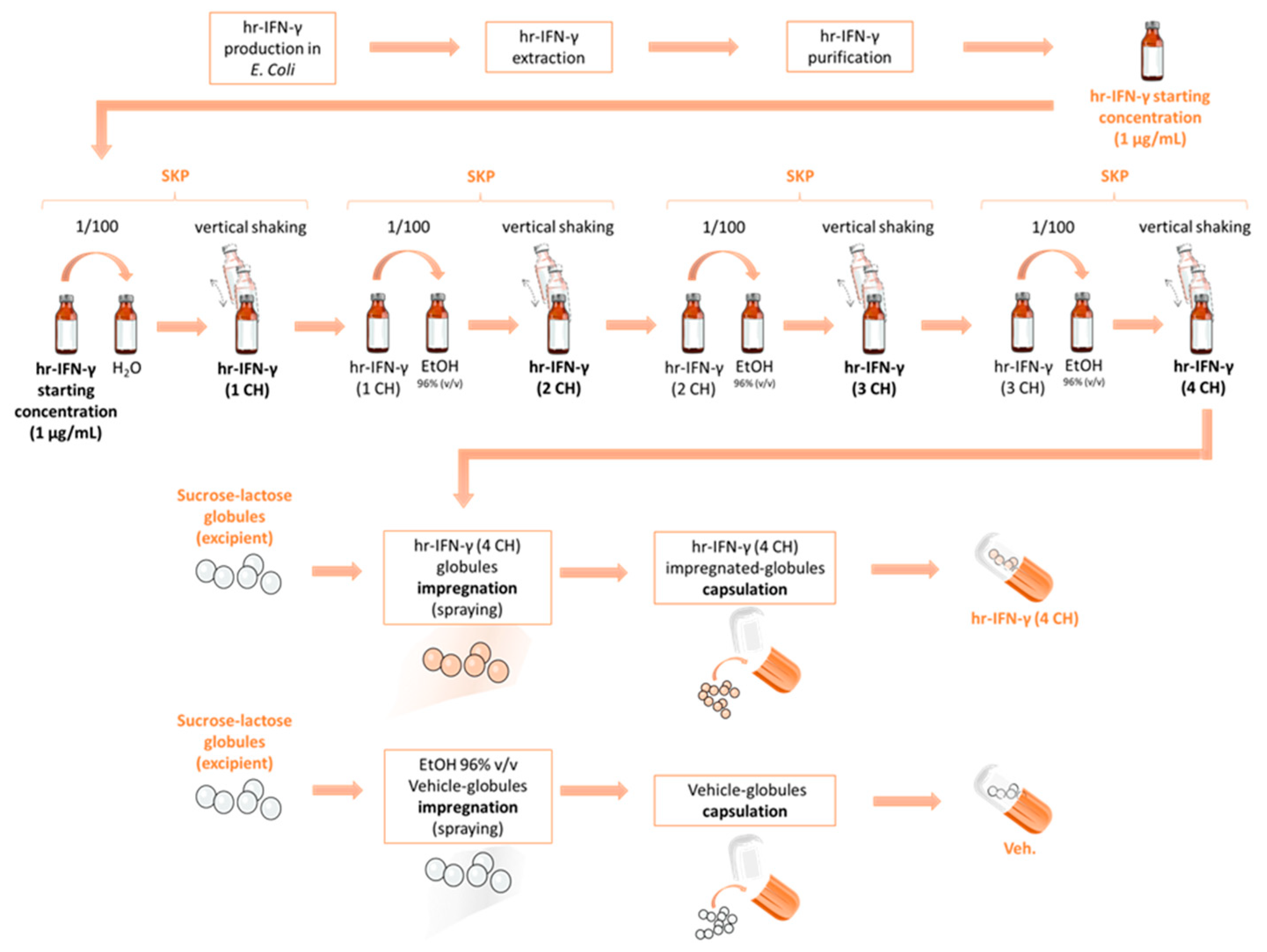
4.1. Tested Item and Experimental Control
4.2. Evaluation of IFN-γ Concentration in Basal Medium Containing the Tested Diluted Pillules
4.3. Evaluation of Cell Surface Activation Markers Expression and Proliferation of PBMCs Subpopulations by Flow Cytometry
4.4. Evaluation of Cell Surface Activation Markers Expression of PBMCs Subpopulations by Flow Cytometry, under CD3 Pre-Primed Condition
4.5. Phagocytosis
4.6. Macrophage Cell Surface Marker Expression and Cytokine Secretion Evaluation
4.7. Evaluation of Cytokine Secretion in THP-1 Cells
4.8. HUVECs Immunoprofiling by Flow Cytometry
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Boehm, U.; Klamp, T.; Groot, M.; Howard, J.C. Cellular responses to interferon-gamma. Annu. Rev. Immunol. 1997, 15, 749–795. [Google Scholar] [CrossRef] [PubMed]
- Keskinen, P.; Ronni, T.; Matikainen, S.; Lehtonen, A.; Julkunen, I. Regulation of HLA class I and II expression by interferons and influenza A virus in human peripheral blood mononuclear cells. Immunology 1997, 91, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, K.; Kagami, M.; Nakazawa, T.; Sugiyama, T.; Sueishi, M.; Ito, M.; Tomioka, H. Regulation of CD69 expression on eosinophil precursors by interferon-gamma. Int. Arch. Allergy Immunol. 2000, 122 (Suppl. 1), 28–32. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, S.F.; Ramsdell, F.; Alderson, M.R. The activation antigen CD69. Stem Cells Dayt Ohio 1994, 12, 456–465. [Google Scholar] [CrossRef]
- Schoenborn, J.R.; Wilson, C.B. Regulation of interferon-gamma during innate and adaptive immune responses. Adv. Immunol. 2007, 96, 41–101. [Google Scholar] [CrossRef] [PubMed]
- Pestka, S.; Kotenko, S.V.; Muthukumaran, G.; Izotova, L.S.; Cook, J.R.; Garotta, G. The interferon gamma (IFN-gamma) receptor: A paradigm for the multichain cytokine receptor. Cytokine Growth Factor Rev. 1997, 8, 189–206. [Google Scholar] [CrossRef]
- Schindler, C.; Plumlee, C. Inteferons pen the JAK–STAT pathway. Semin. Cell Dev. Biol. 2008, 19, 311–318. [Google Scholar] [CrossRef] [Green Version]
- Dawson, R.; Condos, R.; Tse, D.; Huie, M.L.; Ress, S.; Tseng, C.-H.; Brauns, C.; Weiden, M.; Hoshino, Y.; Bateman, E.; et al. Immunomodulation with recombinant interferon-gamma1b in pulmonary tuberculosis. PLoS ONE 2009, 4, e6984. [Google Scholar] [CrossRef] [Green Version]
- van Laarhoven, A.; Kurver, L.; Overheul, G.J.; Kooistra, E.J.; Abdo, W.F.; van Crevel, R.; Duivenvoorden, R.; Kox, M.; Oever, J.T.; Schouten, J.; et al. Interferon gamma immunotherapy in five critically ill COVID-19 patients with impaired cellular immunity: A case series. Medicine 2021, 2, 1163–1170.e2. [Google Scholar] [CrossRef]
- Miller, C.H.; Maher, S.; Young, H.A. Clinical Use of Interferon-γ. Ann. N Y Acad. Sci. 2009, 1182, 69–79. [Google Scholar] [CrossRef]
- Khorana, A.A.; Rosenblatt, J.D.; Sahasrabudhe, D.M.; Evans, T.; Ladrigan, M.; Marquis, D.; Rosell, K.; Whiteside, T.; Phillippe, S.; Acres, B.; et al. A phase I trial of immunotherapy with intratumoral adenovirus-interferon-gamma (TG1041) in patients with malignant melanoma. Cancer Gene Ther. 2003, 10, 251–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dummer, R.; Hassel, J.C.; Fellenberg, F.; Eichmüller, S.; Maier, T.; Slos, P.; Acres, B.; Bleuzen, P.; Bataille, V.; Squiban, P.; et al. Adenovirus-mediated intralesional interferon-gamma gene transfer induces tumor regressions in cutaneous lymphomas. Blood 2004, 104, 1631–1638. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Makino, S.; Araki, Y.; Imamura, T.; Seita, M. Recombinant interferon beta and gamma in the treatment of adult T-cell leukemia. Cancer 1987, 59, 1059–1062. [Google Scholar] [CrossRef]
- Suárez-Méndez, R.; García-García, I.; Fernández-Olivera, N.; Valdés-Quintana, M.; Milanés-Virelles, M.T.; Carbonell, D.; Machado-Molina, D.; Valenzuela-Silva, C.M.; López-Saura, P.A. Adjuvant interferon gamma in patients with drug—Resistant pulmonary tuberculosis: A pilot study. BMC Infect. Dis. 2004, 4, 44. [Google Scholar] [CrossRef] [Green Version]
- Lau, J.Y.N.; Lai, C.L.; Wu, P.C.; Chung, H.T.; Lok, A.S.F.; Lin, H.J. A randomised controlled trial of recombinant interferon-gamma in chinese patients with chronic hepatitis B virus infection. J. Med. Virol. 1991, 34, 184–187. [Google Scholar] [CrossRef]
- Soza, A.; Heller, T.; Ghany, M.; Lutchman, G.; Liang, T.J.; Germain, J.; Hsu, H.H.; Park, Y.; Hoofnagle, J.H. Pilot study of interferon gamma for chronic hepatitis C. J. Hepatol. 2005, 43, 67–71. [Google Scholar] [CrossRef]
- Hansen, M.B. Interleukin-6 signaling requires only few IL-6 molecules: Relation to physiological concentrations of extracellular IL-6. Immun. Inflamm. Dis. 2020, 8, 170–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabrese, E.J.; Giordano, J. Ultra low doses and biological amplification: Approaching Avogadro’s number. Pharmacol. Res. 2021, 170, 105738. [Google Scholar] [CrossRef] [PubMed]
- Manchanda, R.K.; Gupta, M.; Gupta, A.; van Haselen, R. The Clinical and Biological Effects of Homeopathically Prepared Signaling Molecules: A Scoping Review. Homeopath. J. Fac. Homeopath. 2021. [Google Scholar] [CrossRef] [PubMed]
- Jacques, C.; Floris, I.; Lejeune, B. Ultra-Low Dose Cytokines in Rheumatoid Arthritis, Three Birds with One Stone as the Rationale of the 2LARTH® Micro-Immunotherapy Treatment. Int. J. Mol. Sci. 2021, 22, 6717. [Google Scholar] [CrossRef] [PubMed]
- Floris, I.; Rose, T.; Rojas, J.A.C.; Appel, K.; Roesch, C.; Lejeune, B. Pro-Inflammatory Cytokines at Ultra-Low Dose Exert Anti-Inflammatory Effect In Vitro: A Possible Mode of Action Involving Sub-Micron Particles? Dose-Response 2020, 18, 1559325820961723. [Google Scholar] [CrossRef] [PubMed]
- Floris, I.; García-González, V.; Palomares, B.; Appel, K.; Lejeune, B. The Micro-Immunotherapy Medicine 2LARTH® Reduces Inflammation and Symptoms of Rheumatoid Arthritis In Vivo. Int. J. Rheumatol. 2020, 2020, 1594573. [Google Scholar] [CrossRef] [Green Version]
- Jacques, C.; Chatelais, M.; Fekir, K.; Fauconnier, L.; Mellier, M.; Togbe, D.; Floris, I. The Micro-Immunotherapy Medicine 2LEID Exhibits an Immunostimulant Effect by Boosting Both Innate and Adaptive Immune Responses. Int. J. Mol. Sci. 2022, 23, 110. [Google Scholar] [CrossRef] [PubMed]
- Dorward, D.A.; Lucas, C.; Chapman, G.B.; Haslett, C.; Dhaliwal, K.; Rossi, A.G. The Role of Formylated Peptides and Formyl Peptide Receptor 1 in Governing Neutrophil Function during Acute Inflammation. Am. J. Pathol. 2015, 185, 1172–1184. [Google Scholar] [CrossRef] [Green Version]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.; Kim, J.-H.; Jang, H.-J.; Park, J.-W.; Lee, J.-W.; Kwon, O.-K.; Kim, J.-H.; Ahn, K.-S. Anti-Inflammatory Effects of the Fraction from the Leaves of Pyrus pyrifolia on LPS-Stimulated THP-1 Cells. Evid.-Based Complement. Altern. Med. 2021, 2021, 4946241. [Google Scholar] [CrossRef] [PubMed]
- Sharif, O.; Bolshakov, V.N.; Raines, S.; Newham, P.; Perkins, N.D. Transcriptional profiling of the LPS induced NF-kappaB response in macrophages. BMC Immunol. 2007, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decker, M.-L.; Gotta, V.; Wellmann, S.; Ritz, N. Cytokine profiling in healthy children shows association of age with cytokine concentrations. Sci. Rep. 2017, 7, 17842. [Google Scholar] [CrossRef] [Green Version]
- Ellis, T.N.; Beaman, B.L. Interferon-γ activation of polymorphonuclear neutrophil function. Immunology 2004, 112, 2–12. [Google Scholar] [CrossRef]
- Ivashkiv, L.B. IFNγ: Signalling, epigenetics and roles in immunity, metabolism, disease and cancer immunotherapy. Nat. Rev. Immunol. 2018, 18, 545–558. [Google Scholar] [CrossRef]
- Atzeni, F.; Schena, M.; Ongari, A.M.; Carrabba, M.; Bonara, P.; Minonzio, F.; Capsoni, F. Induction of CD69 activation molecule on human neutrophils by GM-CSF, IFN-gamma, and IFN-alpha. Cell. Immunol. 2002, 220, 20–29. [Google Scholar] [CrossRef]
- Hu, X.; Ivashkiv, L.B. Cross-regulation of signaling pathways by interferon-gamma: Implications for immune responses and autoimmune diseases. Immunity. Immunology 2009, 31, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Sol, V.V.-D.; Punzón, C.; Fresno, M. IFN-γ-Induced TNF-α Expression Is Regulated by Interferon Regulatory Factors 1 and 8 in Mouse Macrophages. J. Immunol. 2008, 181, 4461–4470. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Boyanapalli, R.; McPhillips, K.A.; Frasch, S.C.; Janssen, W.J.; Dinauer, M.C.; Riches, D.W.H.; Henson, P.M.; Byrne, A.; Bratton, D.L. Impaired Phagocytosis of Apoptotic Cells by Macrophages in Chronic Granulomatous Disease Is Reversed by IFN-γ in a Nitric Oxide-Dependent Manner. J. Immunol. 2010, 185, 4030–4041. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, R.W.; Rodriguez, P.C.; Toque, H.A.; Narayanan, S.P. Arginase: A Multifaceted Enzyme Important in Health and Disease. Physiol. Rev. 2018, 98, 641–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ming, X.; Rajapakse, A.G.; Yepuri, G.; Xiong, Y.; Carvas, J.M.; Ruffieux, J.; Scerri, I.; Wu, Z.; Popp, K.; Li, J.; et al. Arginase II Promotes Macrophage Inflammatory Responses Through Mitochondrial Reactive Oxygen Species, Contributing to Insulin Resistance and Atherogenesis. J. Am. Hearth Assoc. 2012, 1, e000992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langrish, C.L.; McKenzie, B.S.; Wilson, N.J.; Malefyt, R.D.W.; Kastelein, R.A.; Cua, D.J. IL-12 and IL-23: Master regulators of innate and adaptive immunity. Immunol. Rev. 2004, 202, 96–105. [Google Scholar] [CrossRef]
- Ma, X.; Chow, J.M.; Gri, G.; Carra, G.; Gerosa, F.; Wolf, S.F.; Dzialo, R.; Trinchieri, G. The interleukin 12 p40 gene promoter is primed by interferon gamma in monocytic cells. J. Exp. Med. 1996, 183, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Chujor, C.S.N.; Klein, L.; Lam, C. Selective inhibition of interleukin-1 beta gene expression in activated RAW 264.7 macrophages by interferon-gamma. Eur. J. Immunol. 1996, 26, 1253–1259. [Google Scholar] [CrossRef]
- Mishra, B.B.; Rathinam, V.A.K.; Martens, G.W.; Martinot, A.J.; Kornfeld, H.; A Fitzgerald, K.; Sassetti, C.M. Nitric oxide controls tuberculosis immunopathology by inhibiting NLRP3 inflammasome–dependent IL-1β processing. Nat. Immunol. 2013, 14, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Imai, T.; Nagira, M.; Takagi, S.; Kakizaki, M.; Nishimura, M.; Wang, J.; Gray, P.W.; Matsushima, K.; Yoshie, O. Selective recruitment of CCR4-bearing Th2 cells toward antigen-presenting cells by the CC chemokines thymus and activation-regulated chemokine and macrophage-derived chemokine. Int. Immunol. 1999, 11, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Liddiard, K.; Welch, J.S.; Lozach, J.; Heinz, S.; Glass, C.K.; Greaves, D.R. Interleukin-4 induction of the CC chemokine TARC (CCL17) in murine macrophages is mediated by multiple STAT6 sites in the TARC gene promoter. BMC Mol. Biol. 2006, 7, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, J.B.; Nichols, F.C.; Peluso, J.F. The effects of interferon-gamma and bacterial lipopolysaccharide on CD14 expression in human monocytes. J. Interf. Res. 1992, 12, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Zanoni, I.; Granucci, F. Role of CD14 in host protection against infections and in metabolism regulation. Front. Cell. Infect. Microbiol. 2013, 3, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanoni, I.; Ostuni, R.; Marek, L.R.; Barresi, S.; Barbalat, R.; Barton, G.; Granucci, F.; Kagan, J.C. CD14 controls the LPS-induced endocytosis of Toll-like receptor 4. Cell 2011, 147, 868–880. [Google Scholar] [CrossRef] [Green Version]
- Quero, L.; Hanser, E.; Manigold, T.; Tiaden, A.N.; Kyburz, D. TLR2 stimulation impairs anti-inflammatory activity of M2-like macrophages, generating a chimeric M1/M2 phenotype. Arthritis Res. Ther. 2017, 19, 245. [Google Scholar] [CrossRef] [Green Version]
- Crisler, W.J.; Lenz, L.L. Crosstalk between type I and II interferons in regulation of myeloid cell responses during bacterial infection. Curr. Opin. Immunol. 2018, 54, 35–41. [Google Scholar] [CrossRef]
- Yeap, W.H.; Wong, K.L.; Shimasaki, N.; Teo, E.C.Y.; Quek, J.K.S.; Yong, H.X.; Diong, C.P.; Bertoletti, A.; Linn, Y.C.; Wong, S.C. CD16 is indispensable for antibody-dependent cellular cytotoxicity by human monocytes. Sci. Rep. 2016, 6, 34310. [Google Scholar] [CrossRef]
- Hartnell, A.; Kay, A.B.; Wardlaw, A.J. IFN-gamma induces expression of Fc gamma RIII (CD16) on human eosinophils. J. Immunol. Baltim. Md. 1950 1992, 148, 1471–1478. [Google Scholar]
- Waller, K.; James, C.; de Jong, A.; Blackmore, L.; Ma, Y.; Stagg, A.; Kelsell, D.; O’Dwyer, M.; Hutchins, R.; Alazawi, W. ADAM17-Mediated Reduction in CD14++CD16+ Monocytes ex vivo and Reduction in Intermediate Monocytes With Immune Paresis in Acute Pancreatitis and Acute Alcoholic Hepatitis. Front. Immunol. 2019, 10, 1902. [Google Scholar] [CrossRef] [Green Version]
- Barros, M.H.M.; Hauck, F.; Dreyer, J.H.; Kempkes, B.; Niedobitek, G. Macrophage polarisation: An immunohistochemical approach for identifying M1 and M2 macrophages. PLoS ONE 2013, 8, e80908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristiansen, M.; Graversen, J.H.; Jacobsen, C.; Sonne, O.; Hoffman, H.-J.; Law, S.K.A.; Moestrup, S.K. Identification of the haemoglobin scavenger receptor. Nature 2001, 409, 198–201. [Google Scholar] [CrossRef] [PubMed]
- Snelgrove, R.J.; Goulding, J.; Didierlaurent, A.M.; Lyonga, D.; Vekaria, S.; Edwards, L.; Gwyer, E.; Sedgwick, J.D.; Barclay, A.N.; Hussell, T. A critical function for CD200 in lung immune homeostasis and the severity of influenza infection. Nat. Immunol. 2008, 9, 1074–1083. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Hao, B.; Yang, X.F.; Chen, W.Q. Decreased CD200R expression on monocyte-derived macrophages correlates with Th17/Treg imbalance and disease activity in rheumatoid arthritis patients. Inflamm Res. 2014, 63, 441–450. [Google Scholar] [CrossRef]
- Kotwica-Mojzych, K.; Jodłowska-Jędrych, B.; Mojzych, M. CD200:CD200R Interactions and Their Importance in Immunoregulation. Int. J. Mol. Sci. 2021, 22, 1602. [Google Scholar] [CrossRef]
- Chen, Z.; Marsden, P.A.; Gorczynski, R.M. Role of a distal enhancer in the transcriptional responsiveness of the human CD200 gene to interferon-gamma and tumor necrosis factor-alpha. Mol. Immunol. 2009, 46, 1951–1963. [Google Scholar] [CrossRef]
- Koning, N.; Van Eijk, M.; Pouwels, W.; Brouwer, M.S.; Voehringer, D.; Huitinga, I.; Hoek, R.M.; Raes, G.; Hamann, J. Expression of the inhibitory CD200 receptor is associated with alternative macrophage activation. J. Innate Immun. 2010, 2, 195–200. [Google Scholar] [CrossRef] [Green Version]
- van der Poel, C.E.; Spaapen, R.M.; van de Winkel, J.G.J.; Leusen, J.H.W. Functional characteristics of the high affinity IgG receptor, FcγRI. J. Immunol. Baltim. Md. 2011, 186, 2699–2704. [Google Scholar] [CrossRef] [Green Version]
- Akinrinmade, O.A.; Chetty, S.; Daramola, A.K.; Islam, M.-U.; Thepen, T.; Barth, S. CD64: An Attractive Immunotherapeutic Target for M1-type Macrophage Mediated Chronic Inflammatory Diseases. Biomedicines 2017, 5, 56. [Google Scholar] [CrossRef] [Green Version]
- Antoniades, C.G.; Berry, P.A.; Wendon, J.A.; Vergani, D. The importance of immune dysfunction in determining outcome in acute liver failure. J. Hepatol. 2008, 49, 845–861. [Google Scholar] [CrossRef] [Green Version]
- Bolandi, N.; Derakhshani, A.; Hemmat, N.; Baghbanzadeh, A.; Asadzadeh, Z.; Nour, M.A.; Brunetti, O.; Bernardini, R.; Silvestris, N.; Baradaran, B. The Positive and Negative Immunoregulatory Role of B7 Family: Promising Novel Targets in Gastric Cancer Treatment. Int. J. Mol. Sci. 2021, 22, 10719. [Google Scholar] [CrossRef] [PubMed]
- Delneste, Y.; Charbonnier, P.; Herbault, N.; Magistrelli, G.; Caron, G.; Bonnefoy, J.-Y.; Jeannin, P. Interferon-gamma switches monocyte differentiation from dendritic cells to macrophages. Blood 2003, 101, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Wolk, K.; Döcke, W.-D.; von Baehr, V.; Volk, H.-D.; Sabat, R. Impaired antigen presentation by human monocytes during endotoxin tolerance. Blood 2000, 96, 218–223. [Google Scholar] [CrossRef] [PubMed]
- O’Carroll, C.; Fagan, A.; Shanahan, F.; Carmody, R.J. Identification of a unique hybrid macrophage-polarization state following recovery from lipopolysaccharide tolerance. J. Immunol. Baltim. Md. 1950 2014, 192, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Fontecha, A.; Thomsen, L.L.; Brett, S.; Gerard, C.; Lipp, M.; Lanzavecchia, A.; Sallusto, F. Induced recruitment of NK cells to lymph nodes provides IFN-gamma for T(H)1 priming. Nat. Immunol. 2004, 5, 1260–1265. [Google Scholar] [CrossRef] [PubMed]
- Mai, J.; Virtue, A.; Shen, J.; Wang, H.; Yang, X.-F. An evolving new paradigm: Endothelial cells—Conditional innate immune cells. J. Hematol. Oncol. J. Hematol. Oncol. 2013, 6, 61. [Google Scholar] [CrossRef] [Green Version]
- van Hinsbergh, V.W.M. Endothelium—Role in regulation of coagulation and inflammation. Semin. Immunopathol. 2012, 34, 93–106. [Google Scholar] [CrossRef] [Green Version]
- Ferrà-Cañellas, M.D.M.; Munar-Bestard, M.; Garcia Sureda, L.; Lejeune, B.; Ramis, J.M.; Monjo, M. BMP4 micro—#immunotherapy increases collagen deposition and reduces PGE2 release in human gingival fibroblasts and increases tissue viability of engineered 3D gingiva under inflammatory conditions. J. Periodontol. 2021, 92, 1448–1459. [Google Scholar] [CrossRef]
- Floris, I.; Appel, K.; Rose, T.; Lejeune, B. 2LARTH®, a micro-immunotherapy medicine, exerts anti-inflammatory effects in vitro and reduces TNF-α and IL-1β secretion. J. Inflamm. Res. 2018, 11, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Lilli, N.L.; Révy, D.; Robelet, S.; Lejeune, B. Effect of the micro-immunotherapy medicine 2LPARK® on rat primary dopaminergic neurons after 6-OHDA injury: Oxidative stress and survival evaluation in an in vitro model of Parkinson’s disease. Degener. Neurol. Neuromuscul. Dis. 2019, 9, 79–88. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IFN-γ (4 CH) Sucrose-Lactose Pillules Concentrations | ||
|---|---|---|
| Parameters (pg/mL) | 11 mM | 22 mM |
| Mean | 1.63 | 1.91 |
| SD | 0.43 | 0.69 |
| Median | 1.6 | 1.72 |
| Max | 2.12 | 2.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacques, C.; Chatelais, M.; Fekir, K.; Brulefert, A.; Floris, I. The Unitary Micro-Immunotherapy Medicine Interferon-γ (4 CH) Displays Similar Immunostimulatory and Immunomodulatory Effects than Those of Biologically Active Human Interferon-γ on Various Cell Types. Int. J. Mol. Sci. 2022, 23, 2314. https://doi.org/10.3390/ijms23042314
Jacques C, Chatelais M, Fekir K, Brulefert A, Floris I. The Unitary Micro-Immunotherapy Medicine Interferon-γ (4 CH) Displays Similar Immunostimulatory and Immunomodulatory Effects than Those of Biologically Active Human Interferon-γ on Various Cell Types. International Journal of Molecular Sciences. 2022; 23(4):2314. https://doi.org/10.3390/ijms23042314
Chicago/Turabian StyleJacques, Camille, Mathias Chatelais, Karim Fekir, Adrien Brulefert, and Ilaria Floris. 2022. "The Unitary Micro-Immunotherapy Medicine Interferon-γ (4 CH) Displays Similar Immunostimulatory and Immunomodulatory Effects than Those of Biologically Active Human Interferon-γ on Various Cell Types" International Journal of Molecular Sciences 23, no. 4: 2314. https://doi.org/10.3390/ijms23042314
APA StyleJacques, C., Chatelais, M., Fekir, K., Brulefert, A., & Floris, I. (2022). The Unitary Micro-Immunotherapy Medicine Interferon-γ (4 CH) Displays Similar Immunostimulatory and Immunomodulatory Effects than Those of Biologically Active Human Interferon-γ on Various Cell Types. International Journal of Molecular Sciences, 23(4), 2314. https://doi.org/10.3390/ijms23042314